
Dietary Habit Is Associated with Depression and Intelligence: An Observational and Genome-Wide Environmental Interaction Analysis in the UK Biobank Cohort

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Definition of Depression and Intelligence in the UK Biobank Samples
2.2. Genotyping, Imputation and Quality Control in the UK Biobank
2.3. GWAS Summary Data of Dietary Habits
2.4. Dietary Habit Related PRS Calculation and Association Analysis
2.5. Genome-Wide Environmental Interaction (GWEI) Study
3. Results
3.1. Associations of Dietary Habits with Depression and Fluid Intelligence
3.2. Interaction Analysis of Dietary Habits with Depression and Fluid Intelligence
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Dietary Habits | OR | p |
|---|---|---|
| Cereal type: cornflakes/frosties vs. any other | 0.96 | 5.66 × 10−12 |
| Among current drinkers, drinks usually with meals: yes vs. no | 0.97 | 4.24 × 10−7 |
| Coffee type: decaffeinated vs. any other | 0.98 | 1.78 × 10−6 |
| Bread type: white vs. any other | 0.98 | 1.82 × 10−6 |
| Never eat wheat vs. no wheat restrictions | 0.98 | 7.40 × 10−6 |
| Overall cheese intake | 0.98 | 1.70 × 10−5 |
| Frequency of adding salt to food | 1.02 | 1.79 × 10−4 |
| Among current drinkers. Drinks usually with meals: yes vs. no | 1.02 | 3.45 × 10−4 |
| Among current drinkers, drinks usually with meals: yes, it varies, no | 0.98 | 3.57 × 10−4 |
| Cups of tea per day | 1.02 | 5.07 × 10−4 |
| Champagne/white wine glasses per month | 0.98 | 6.56 × 10−4 |
| Total drinks of alcohol per month | 0.98 | 6.86 × 10−4 |
| Bowls of cereal per week | 0.98 | 8.66 × 10−4 |
| Bread type: white vs. wholemeal/wholegrain + brown | 1.02 | 1.42 × 10−3 |
| Tablespoons of vegetables per day | 0.99 | 3.94 × 10−3 |
| Bread type: wholemeal/wholegrain vs. white + brown | 0.99 | 4.88 × 10−3 |
| Slices of bread per week | 0.99 | 6.76 × 10−3 |
| Among current drinkers, drinks usually with meals: yes + it varies vs. no | 0.99 | 7.41 × 10−3 |
| Overall oily fish intake | 0.99 | 8.04 × 10−3 |
| Red wine glasses per month | 0.99 | 8.75 × 10−3 |
| Milk type: soy milk vs. never | 0.99 | 8.76 × 10−3 |
| Never eat sugar vs. no sugar restrictions | 1.01 | 1.09 × 10−2 |
| Never eat wheat vs. no eggs, dairy, wheat, or sugar restrictions | 0.99 | 1.14 × 10−2 |
| Overall non-oily fish intake | 0.99 | 1.17 × 10−2 |
| Milk type: skimmed vs. never | 0.99 | 1.66 × 10−2 |
| Cereal type: muesli vs. any other | 1.01 | 2.83 × 10−2 |
| Tablespoons of cooked vegetables per day | 1.01 | 2.87 × 10−2 |
| Overall alcohol intake | 0.99 | 3.60 × 10−2 |
| Alcohol drinker status: current + former vs. never | 0.99 | 4.15 × 10−2 |
| Slices of bread per week | 0.99 | 4.21 × 10−2 |
| Overall poultry intake | 0.99 | 4.41 × 10−2 |
| Cereal type: muesli vs. any other | 0.99 | 4.87 × 10−2 |
| Dietary Habits | Beta | p |
|---|---|---|
| Bread type: white vs. any other | 0.03 | 1.30 × 10−32 |
| Bread type: whole grain/whole meal vs. white bread | 0.02 | 3.13 × 10−23 |
| Overall cheese intake | 0.02 | 1.20 × 10−22 |
| Red wine glasses per month | 0.02 | 3.35 × 10−19 |
| Among current drinkers. drinks usually with meals: yes vs. no | −0.02 | 1.21 × 10−13 |
| Cereal type: cornflakes/frosties vs. any other | 0.01 | 8.36 × 10−10 |
| Overall alcohol intake | 0.01 | 8.31 × 10−8 |
| Champagne/white wine glasses per month | 0.01 | 1.07 × 10−6 |
| Temperature of hot drinks | −0.01 | 1.18 × 10−6 |
| Among current drinkers, drinks usually with meals: yes vs. no | 0.01 | 2.85 × 10−6 |
| Never eat sugar vs. no sugar restrictions | 0.01 | 6.91 × 10−6 |
| Bread type: wholemeal/wholegrain vs. any other | 0.01 | 9.75 × 10−6 |
| Overall non-oily fish intake | 0.01 | 1.24 × 10−5 |
| Bread type: wholemeal/wholegrain vs. white + brown | 0.01 | 1.51 × 10−5 |
| Spread type: flora + benecol vs. never | 0.01 | 4.50 × 10−5 |
| Overall oily fish intake | −0.01 | 7.96 × 10−5 |
| Spread type: tub margarine vs. never | −0.01 | 1.96 × 10−4 |
| Never eat wheat vs. no wheat restrictions | 0.01 | 3.43 × 10−4 |
| Spread type: butter and butter-like spreads vs. oil-based spreads | 0.01 | 3.99 × 10−4 |
| Never eat sugar vs. no sugar restrictions | −0.01 | 5.99 × 10−4 |
| Spread type: butter and margarine spreads vs. oil-based spreads | −0.01 | 7.58 × 10−4 |
| Bowls of cereal per week | 0.01 | 8.10 × 10−4 |
| Cups of tea per day | −0.01 | 1.43 × 10−3 |
| Tablespoons of vegetables per day | 0.01 | 3.54 × 10−3 |
| Cereal type: comflakes/frosties vs. any other | 0.01 | 5.97 ×10−3 |
| Spread type: low fat spread vs. never | −0.01 | 5.97 × 10−3 |
| Beer/cider glasses per month | −0.01 | 7.68 × 10−3 |
| Cereal type: biscuit cereal vs. any other | 0.01 | 8.49 × 10−3 |
| Coffee type: decaffeinated vs. any other | 0.01 | 8.77 × 10−3 |
| Never eat wheat vs. no eggs, dairy, wheat, or sugar restrictions | 0.01 | 9.77 × 10−3 |
| Among current drinkers, drinks usually with meals: yes, it varies, no | 0.01 | 1.09 × 10−2 |
| Bread type: white vs. any other | −0.01 | 1.16 × 10−2 |
| Pieces of dried fruit per day | 0.01 | 1.49 × 10−2 |
| Milk type: skimmed vs. never | 0.01 | 1.53 × 10−2 |
| Overall non-oily fish intake | −0.01 | 1.63 × 10−2 |
| Slices of bread per week | 0.01 | 1.98 × 10−2 |
| Alcohol drinker status: current + former vs. never | 0.01 | 2.09 × 10−2 |
| Overall beef intake | −0.01 | 2.33 × 10−2 |
| Cups of tea per day | −0.01 | 2.33 × 10−2 |
| Total drinks of alcohol per month | 0.01 | 2.92 × 10−2 |
| Overall lamb/mutton intake | 0.01 | 3.07 × 10−2 |
References
- Malhi, G.S.; Mann, J.J. Depression. Lancet 2018, 392, 2299–2312. [Google Scholar] [CrossRef]
- Kessler, R.C.; Bromet, E.J. The epidemiology of depression across cultures. Annu. Rev. Public Health 2013, 34, 119–138. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, E.; Shaywitz, B.A.; Holahan, J.M.; Marchione, K.; Shaywitz, S.E. Uncoupling of reading and iq over time: Empirical evidence for a definition of dyslexia. Psychol. Sci. 2010, 21, 93–101. [Google Scholar] [CrossRef]
- Benson, N.F.; Kranzler, J.H.; Floyd, R.G. Examining the integrity of measurement of cognitive abilities in the prediction of achievement: Comparisons and contrasts across variables from higher-order and bifactor models. J. Sch. Psychol. 2016, 58, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Jaycox, L.H.; Stein, B.D.; Paddock, S.; Miles, J.N.; Chandra, A.; Meredith, L.S.; Tanielian, T.; Hickey, S.; Burnam, M.A. Impact of teen depression on academic, social, and physical functioning. Pediatrics 2009, 124, e596–e605. [Google Scholar] [CrossRef] [Green Version]
- Rice, F.; Lifford, K.J.; Thomas, H.V.; Thapar, A. Mental health and functional outcomes of maternal and adolescent reports of adolescent depressive symptoms. J. Am. Acad. Child Adolesc. Psychiatry 2007, 46, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Barbe, R.P.; Bridge, J.; Birmaher, B.; Kolko, D.; Brent, D.A. Suicidality and its relationship to treatment outcome in depressed adolescents. Suicide Life-Threat. Behav. 2004, 34, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Haier, R.J.; Jung, R.E.; Yeo, R.A.; Head, K.; Alkire, M.T. Structural brain variation and general intelligence. NeuroImage 2004, 23, 425–433. [Google Scholar] [CrossRef]
- Neubauer, A.C.; Grabner, R.H.; Fink, A.; Neuper, C. Intelligence and neural efficiency: Further evidence of the influence of task content and sex on the brain-iq relationship. Brain Res. Cogn. Brain Res. 2005, 25, 217–225. [Google Scholar] [CrossRef]
- Wray, N.R.; Ripke, S.; Mattheisen, M.; Trzaskowski, M.; Byrne, E.M.; Abdellaoui, A.; Adams, M.J.; Agerbo, E.; Air, T.M.; Andlauer, T.M.F.; et al. Genome-wide association analyses identify 44 risk variants and refine the genetic architecture of major depression. Nat. Genet. 2018, 50, 668–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aichele, S.; Rabbitt, P.; Ghisletta, P. Illness and intelligence are comparatively strong predictors of individual differences in depressive symptoms following middle age. Aging Ment. Health 2019, 23, 122–131. [Google Scholar] [CrossRef]
- Aichele, S.; Ghisletta, P.; Corley, J.; Pattie, A.; Taylor, A.M.; Starr, J.M.; Deary, I.J. Fluid intelligence predicts change in depressive symptoms in later life: The lothian birth cohort 1936. Psychol. Sci. 2018, 29, 1984–1995. [Google Scholar] [CrossRef] [Green Version]
- Dobson, K.G.; Schmidt, L.A.; Saigal, S.; Boyle, M.H.; Van Lieshout, R.J. Childhood cognition and lifetime risk of major depressive disorder in extremely low birth weight and normal birth weight adults. J. Dev. Orig. Health Dis. 2016, 7, 574–580. [Google Scholar] [CrossRef]
- Masana, M.F.; Tyrovolas, S.; Kolia, N.; Chrysohoou, C.; Skoumas, J.; Haro, J.M.; Tousoulis, D.; Papageorgiou, C.; Pitsavos, C.; Panagiotakos, D.B. Dietary patterns and their association with anxiety symptoms among older adults: The attica study. Nutrients 2019, 11, 1250. [Google Scholar] [CrossRef] [Green Version]
- Leventakou, V.; Roumeliotaki, T.; Sarri, K.; Koutra, K.; Kampouri, M.; Kyriklaki, A.; Vassilaki, M.; Kogevinas, M.; Chatzi, L. Dietary patterns in early childhood and child cognitive and psychomotor development: The rhea mother-child cohort study in crete. Br. J. Nutr. 2016, 115, 1431–1437. [Google Scholar] [CrossRef] [Green Version]
- Velten, J.; Lavallee, K.L.; Scholten, S.; Meyer, A.H.; Zhang, X.C.; Schneider, S.; Margraf, J. Lifestyle choices and mental health: A representative population survey. BMC Psychol. 2014, 2, 58. [Google Scholar] [CrossRef] [PubMed]
- Merino, J.; Dashti, H.S. Genome-wide meta-analysis of macronutrient intake of 91,114 european ancestry participants from the cohorts for heart and aging research in genomic epidemiology consortium. Mol. Psychiatry 2019, 24, 1920–1932. [Google Scholar] [CrossRef] [PubMed]
- Bulik-Sullivan, B.; Finucane, H.K.; Anttila, V.; Gusev, A.; Day, F.R. An atlas of genetic correlations across human diseases and traits. Nat. Genet. 2015, 47, 1236–1241. [Google Scholar] [CrossRef] [Green Version]
- Purcell, S.M.; Wray, N.R.; Stone, J.L.; Visscher, P.M.; O’Donovan, M.C.; Sullivan, P.F.; Sklar, P. Common polygenic variation contributes to risk of schizophrenia and bipolar disorder. Nature 2009, 460, 748–752. [Google Scholar] [PubMed]
- Dudbridge, F. Polygenic epidemiology. Genet. Epidemiol. 2016, 40, 268–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, M.J.; Martin, J.; Lu, Y.; Brikell, I.; Lundström, S.; Larsson, H.; Lichtenstein, P. Association of genetic risk factors for psychiatric disorders and traits of these disorders in a swedish population twin sample. JAMA Psychiatry 2019, 76, 280–289. [Google Scholar] [CrossRef]
- Hunter, D.J. Gene-environment interactions in human diseases. Nat. Rev. Genet. 2005, 6, 287–298. [Google Scholar] [CrossRef]
- Zwicker, A.; Denovan-Wright, E.M.; Uher, R. Gene-environment interplay in the etiology of psychosis. Psychol. Med. 2018, 48, 1925–1936. [Google Scholar] [CrossRef] [PubMed]
- Nugent, N.R.; Tyrka, A.R.; Carpenter, L.L.; Price, L.H. Gene-environment interactions: Early life stress and risk for depressive and anxiety disorders. Psychopharmacology 2011, 214, 175–196. [Google Scholar] [CrossRef] [Green Version]
- Normann, C.; Buttenschøn, H.N. Gene-environment interactions between hpa-axis genes and stressful life events in depression: A systematic review. Acta Neuropsychiatr. 2019, 31, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Bycroft, C.; Freeman, C.; Petkova, D.; Band, G.; Elliott, L.T.; Sharp, K.; Motyer, A.; Vukcevic, D.; Delaneau, O.; O’Connell, J.; et al. The uk biobank resource with deep phenotyping and genomic data. Nature 2018, 562, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, R.C.; Andrews, G.; Mroczek, D.; Ustun, B.; Wittchen, H.U. The world health organization composite international diagnostic interview short-form (CIDI-SF). Int. J. Methods Psychiatr. Res. 1998, 7, 171–185. [Google Scholar] [CrossRef]
- Kroenke, K.; Spitzer, R.L.; Williams, J.B.W.; Löwe, B. The patient health questionnaire somatic, anxiety, and depressive symptom scales: A systematic review. Gen. Hosp. Psychiatry 2010, 32, 345–359. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.A.S.; Cullen, B.; Adams, M.; Brailean, A.; Breen, G.; Coleman, J.R.I.; Dregan, A.; Gaspar, H.A.; Hübel, C.; Lee, W.; et al. Indicators of mental disorders in uk biobank—A comparison of approaches. Int. J. Methods Psychiatr. Res. 2019, 28, e1796. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.B.; Florez, J.C.; Hirschhorn, J.N. Comprehensive genomic analysis of dietary habits in uk biobank identifies hundreds of genetic associations. Nat. Commun. 2020, 11, 1467. [Google Scholar] [CrossRef] [Green Version]
- Loh, P.R.; Tucker, G.; Bulik-Sullivan, B.K.; Vilhjálmsson, B.J.; Finucane, H.K.; Salem, R.M.; Chasman, D.I.; Ridker, P.M.; Neale, B.M.; Berger, B.; et al. Efficient bayesian mixed-model analysis increases association power in large cohorts. Nat. Genet. 2015, 47, 284–290. [Google Scholar] [CrossRef]
- Loh, P.R.; Kichaev, G.; Gazal, S. Mixed-model association for biobank-scale datasets. Nat. Genet. 2018, 50, 906–908. [Google Scholar] [CrossRef]
- Abraham, G.; Qiu, Y.; Inouye, M. Flashpca2: Principal component analysis of biobank-scale genotype datasets. Bioinformatics 2017, 33, 2776–2778. [Google Scholar] [CrossRef]
- Benner, C.; Havulinna, A.S.; Järvelin, M.R.; Salomaa, V.; Ripatti, S.; Pirinen, M. Prospects of fine-mapping trait-associated genomic regions by using summary statistics from genome-wide association studies. Am. J. Hum. Genet. 2017, 101, 539–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. Plink: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruim, R.J.; Welch, R.P.; Sanna, S.; Teslovich, T.M.; Chines, P.S.; Gliedt, T.P.; Boehnke, M.; Abecasis, G.R.; Willer, C.J. Locuszoom: Regional visualization of genome-wide association scan results. Bioinformatics 2010, 26, 2336–2337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasterling, J.J.; Duke, L.M.; Brailey, K.; Constans, J.I.; Allain, A.N., Jr.; Sutker, P.B. Attention, learning, and memory performances and intellectual resources in vietnam veterans: Ptsd and no disorder comparisons. Neuropsychology 2002, 16, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Yehuda, R.; Keefe, R.S.; Harvey, P.D.; Levengood, R.A.; Gerber, D.K.; Geni, J.; Siever, L.J. Learning and memory in combat veterans with posttraumatic stress disorder. Am. J. Psychiatry 1995, 152, 137–139. [Google Scholar]
- Keyes, K.M.; Platt, J.; Kaufman, A.S.; McLaughlin, K.A. Association of fluid intelligence and psychiatric disorders in a population-representative sample of us adolescents. JAMA Psychiatry 2017, 74, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Connor, J.P.; Haber, P.S.; Hall, W.D. Alcohol use disorders. Lancet 2016, 387, 988–998. [Google Scholar] [CrossRef] [Green Version]
- Awaworyi Churchill, S.; Farrell, L. Alcohol and depression: Evidence from the 2014 health survey for england. Drug Alcohol Depend. 2017, 180, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Batty, G.D.; Deary, I.J.; Schoon, I.; Emslie, C.; Hunt, K.; Gale, C.R. Childhood mental ability and adult alcohol intake and alcohol problems: The 1970 british cohort study. Am. J. Public Health 2008, 98, 2237–2243. [Google Scholar] [CrossRef]
- Batty, G.D.; Deary, I.J.; Macintyre, S. Childhood iq and life course socioeconomic position in relation to alcohol induced hangovers in adulthood: The aberdeen children of the 1950s study. J. Epidemiol. Community Health 2006, 60, 872–874. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, L.H.; Sørensen, T.I.; Grønbaek, M. Intelligence in relation to later beverage preference and alcohol intake. Addiction 2005, 100, 1445–1452. [Google Scholar] [CrossRef]
- Gangwisch, J.E.; Hale, L.; Garcia, L.; Malaspina, D.; Opler, M.G.; Payne, M.E.; Rossom, R.C.; Lane, D. High glycemic index diet as a risk factor for depression: Analyses from the women’s health initiative. Am. J. Clin. Nutr. 2015, 102, 454–463. [Google Scholar] [CrossRef] [Green Version]
- Knüppel, A.; Shipley, M.J.; Llewellyn, C.H.; Brunner, E.J. Sugar intake from sweet food and beverages, common mental disorder and depression: Prospective findings from the whitehall ii study. Sci. Rep. 2017, 7, 6287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quirk, S.E.; Williams, L.J.; O’Neil, A.; Pasco, J.A.; Jacka, F.N.; Housden, S.; Berk, M.; Brennan, S.L. The association between diet quality, dietary patterns and depression in adults: A systematic review. BMC Psychiatry 2013, 13, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.; Cheng, L.; Jiang, W. Sugar-sweetened beverages consumption and the risk of depression: A meta-analysis of observational studies. J. Affect. Disord. 2019, 245, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Westover, A.N.; Marangell, L.B. A cross-national relationship between sugar consumption and major depression? Depress. Anxiety 2002, 16, 118–120. [Google Scholar] [CrossRef]
- Li, R.; Diao, H.; Zhao, F.; Xiao, S.; El Zowalaty, A.E.; Dudley, E.A.; Mattson, M.P.; Ye, X. Olfactomedin 1 deficiency leads to defective olfaction and impaired female fertility. Endocrinology 2015, 156, 3344–3357. [Google Scholar] [CrossRef] [Green Version]
- Nakaya, N.; Sultana, A.; Lee, H.S.; Tomarev, S.I. Olfactomedin 1 interacts with the nogo a receptor complex to regulate axon growth. J. Biol. Chem. 2012, 287, 37171–37184. [Google Scholar] [CrossRef] [Green Version]
- Recabarren-Leiva, D.; Alarcón, M. New insights into the gene expression associated to amyotrophic lateral sclerosis. Life Sci. 2018, 193, 110–123. [Google Scholar] [CrossRef]
- Yu, Y.; Shintani, T.; Takeuchi, Y.; Shirasawa, T.; Noda, M. Protein tyrosine phosphatase receptor type j (ptprj) regulates retinal axonal projections by inhibiting eph and abl kinases in mice. J. Neurosci. 2018, 38, 8345–8363. [Google Scholar] [CrossRef] [PubMed]
- Wigner, P.; Synowiec, E.; Jóźwiak, P.; Czarny, P.; Bijak, M.; Białek, K.; Szemraj, J.; Gruca, P.; Papp, M.; Śliwiński, T. The effect of chronic mild stress and venlafaxine on the expression and methylation levels of genes involved in the tryptophan catabolites pathway in the blood and brain structures of rats. J. Mol. Neurosci. 2020, 70, 1425–1436. [Google Scholar] [CrossRef]
- Nehlig, A. Is caffeine a cognitive enhancer? J. Alzheimer’s Dis. 2010, 20 (Suppl. S1), S85–S94. [Google Scholar] [CrossRef] [Green Version]
- Cattell, R.B. The effects of alcohol and caffeine on intelligent and associative performance. Br. J. Med. Psychol. 1930, 10, 20–33. [Google Scholar] [CrossRef]
- Corley, J.; Jia, X.; Kyle, J.A.; Gow, A.J.; Brett, C.E.; Starr, J.M.; McNeill, G.; Deary, I.J. Caffeine consumption and cognitive function at age 70: The lothian birth cohort 1936 study. Psychosom. Med. 2010, 72, 206–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, K.; Allen, D.; Lader, M. The influences of age and caffeine on psychomotor and cognitive function. Psychopharmacology 1999, 145, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Irwin, C.; Khalesi, S.; Desbrow, B.; McCartney, D. Effects of acute caffeine consumption following sleep loss on cognitive, physical, occupational and driving performance: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2020, 108, 877–888. [Google Scholar] [CrossRef]
- Fagerberg, L.; Hallström, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol. Cell. Proteom. 2014, 13, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; You, X.; Li, S.; Long, Q.; Zhu, Y.; Teng, Z.; Zeng, Y. Peripheral blood leukocyte rna-seq identifies a set of genes related to abnormal psychomotor behavior characteristics in patients with schizophrenia. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2020, 26, e922426-1. [Google Scholar] [CrossRef] [PubMed]
- Kibbe, W.A.; Arze, C.; Felix, V.; Mitraka, E.; Bolton, E.; Fu, G.; Mungall, C.J.; Binder, J.X.; Malone, J.; Vasant, D.; et al. Disease ontology 2015 update: An expanded and updated database of human diseases for linking biomedical knowledge through disease data. Nucleic Acids Res. 2015, 43, D1071–D1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simard, S.; Coppola, G.; Rudyk, C.A.; Hayley, S.; McQuaid, R.J.; Salmaso, N. Profiling changes in cortical astroglial cells following chronic stress. Neuropsychopharmacology 2018, 43, 1961–1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catanzaro, I.; Naselli, F.; Saverini, M.; Giacalone, A.; Montalto, G.; Caradonna, F. Cytochrome p450 2e1 variable number tandem repeat polymorphisms and health risks: A genotype-phenotype study in cancers associated with drinking and/or smoking. Mol. Med. Rep. 2012, 6, 416–420. [Google Scholar] [CrossRef] [Green Version]
- Dannenberg, L.O.; Chen, H.J.; Tian, H.; Edenberg, H.J. Differential regulation of the alcohol dehydrogenase 1b (adh1b) and adh1c genes by DNA methylation and histone deacetylation. Alcohol. Clin. Exp. Res. 2006, 30, 928–937. [Google Scholar] [CrossRef]
- Edenberg, H.J. The genetics of alcohol metabolism: Role of alcohol dehydrogenase and aldehyde dehydrogenase variants. Alcohol Res. Health J. Natl. Inst. Alcohol Abus. Alcohol. 2007, 30, 5–13. [Google Scholar]
- Witkin, S.S.; Linhares, I.; Giraldo, P.; Jeremias, J.; Ledger, W.J. Individual immunity and susceptibility to female genital tract infection. Am. J. Obstet. Gynecol. 2000, 183, 252–256. [Google Scholar] [CrossRef]
- Naselli, F.; Catanzaro, I.; Bellavia, D.; Perez, A.; Sposito, L.; Caradonna, F. Role and importance of polymorphisms with respect to DNA methylation for the expression of cyp2e1 enzyme. Gene 2014, 536, 29–39. [Google Scholar] [CrossRef]
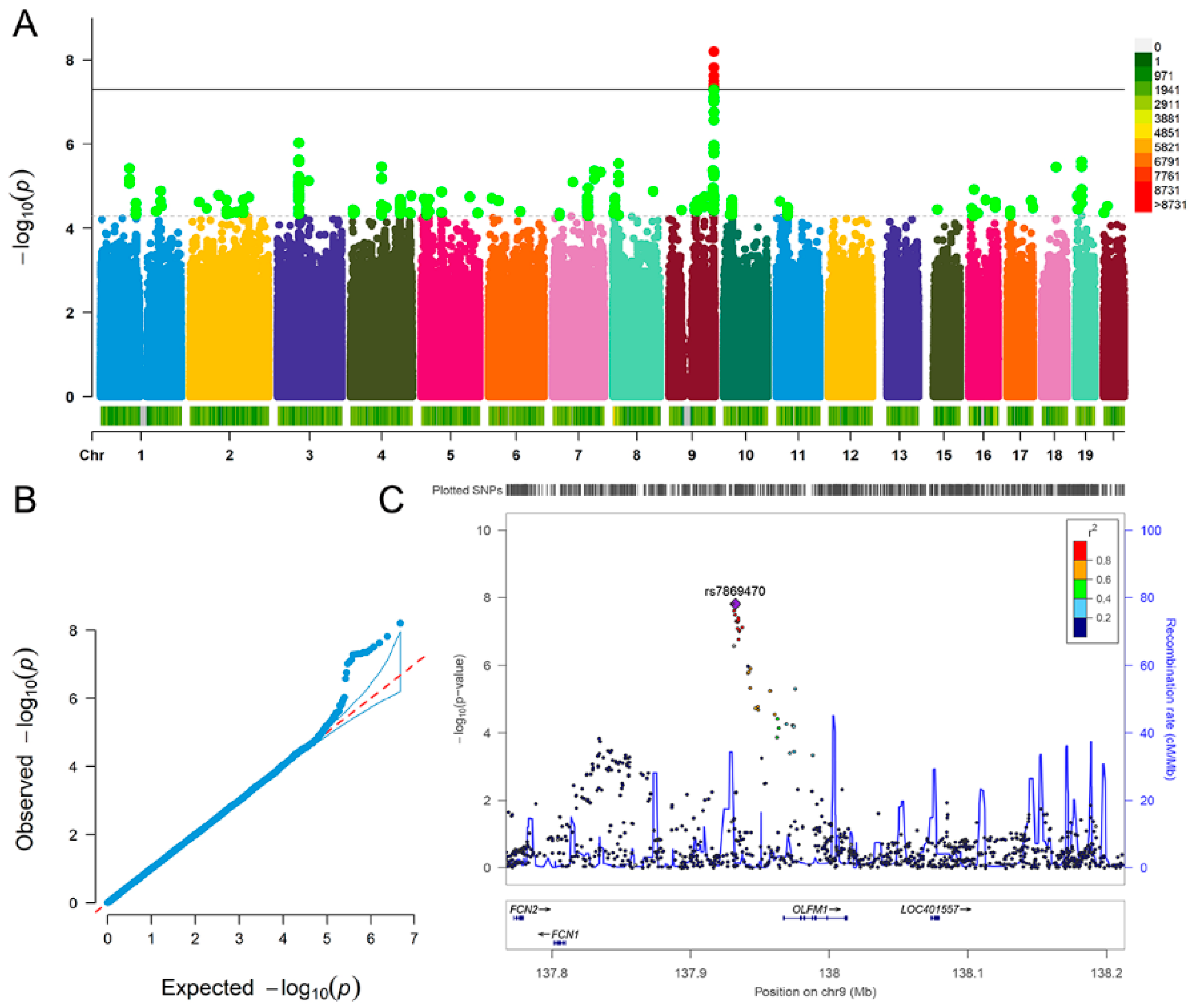
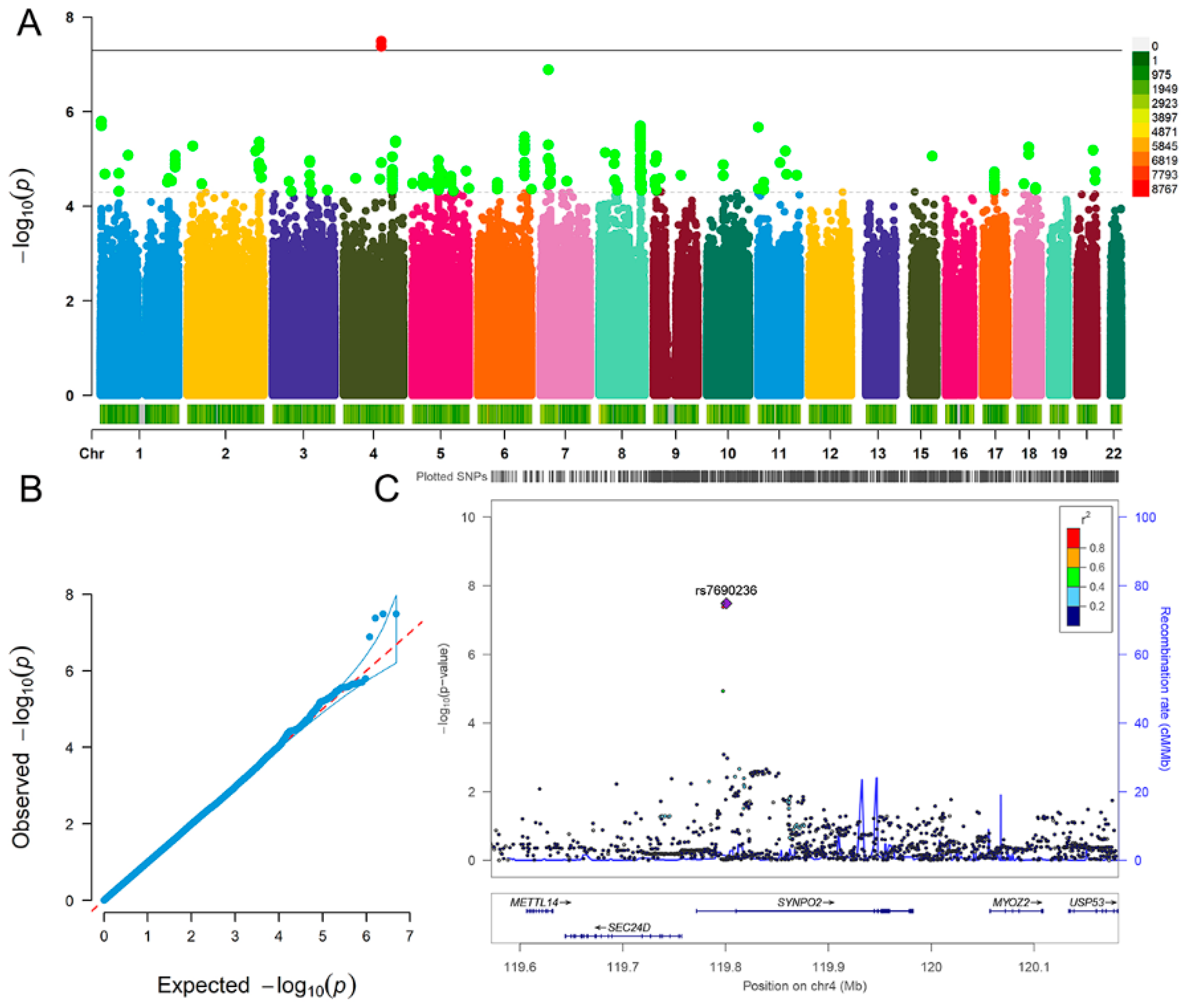
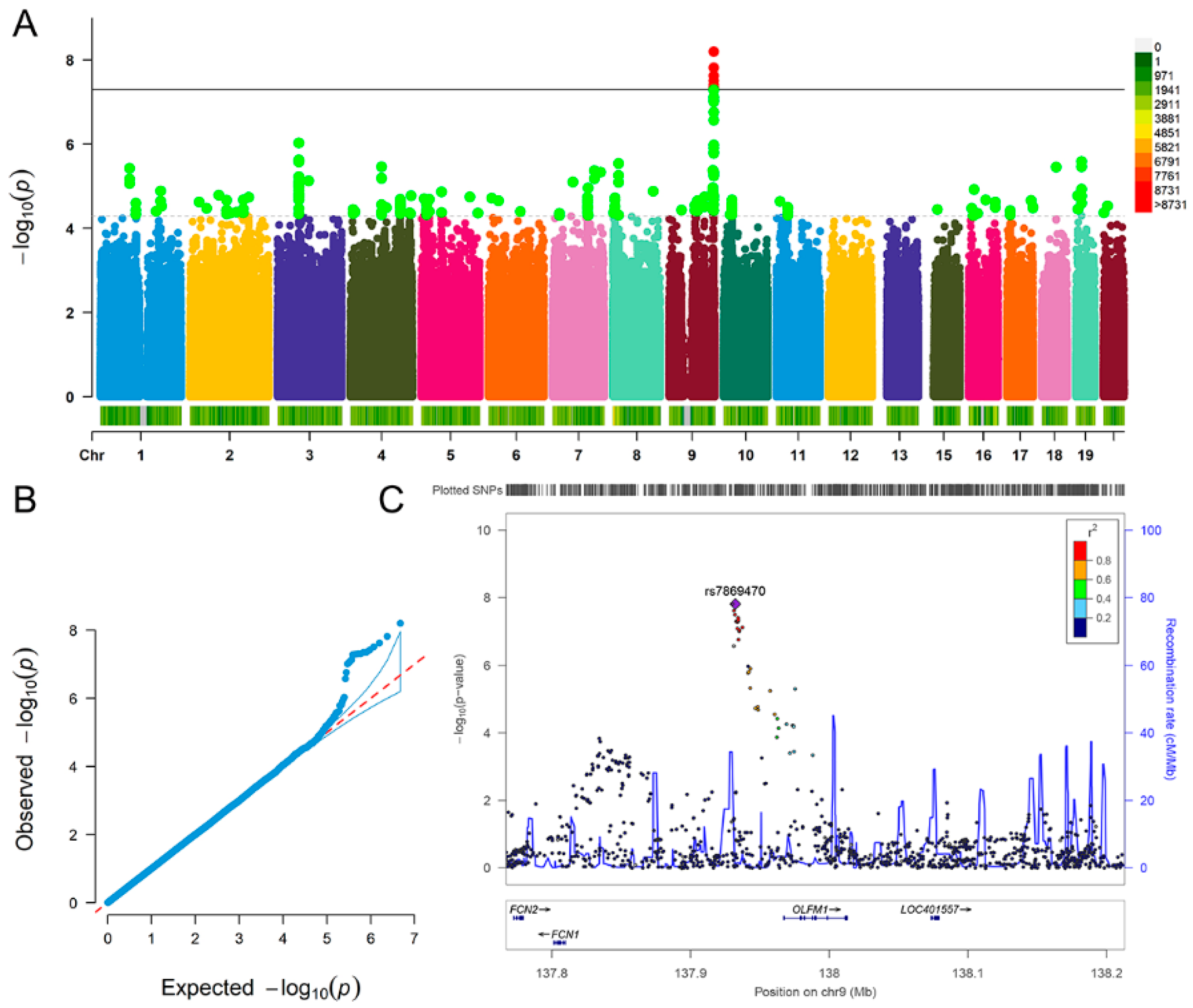
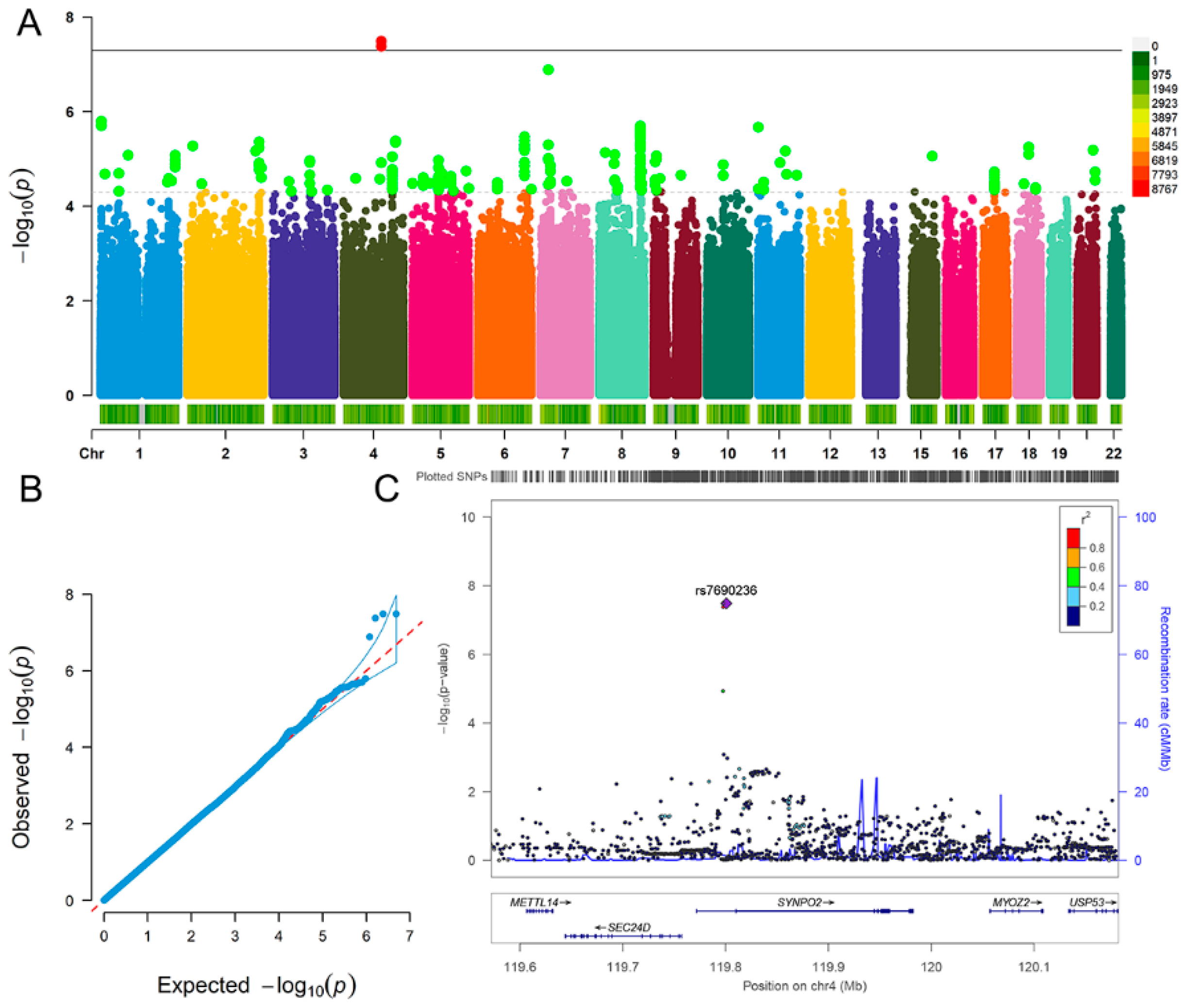
| Fluid Intelligence | Depression | |
|---|---|---|
| Participants | 160,121 | 153,549 (case = 74,579) |
| Sex (female) | 86,818 (54.22%) | 87,265 (56.83%) |
| Age (years) | 56.70 ± 8.15 | 56.12 ± 7.78 |
| SNP | ALT | A1 | Beta | SE | p |
|---|---|---|---|---|---|
| rs7869470 | A | G | 0.062 | 0.011 | 1.54 × 10−8 |
| rs34379422 | C | T | 0.061 | 0.011 | 2.39 × 10−8 |
| rs796938996 | G | GCG | 0.067 | 0.011 | 6.33 × 10−9 |
| rs17493408 | A | G | 0.061 | 0.011 | 3.13 × 10−8 |
| rs11103643 | T | C | 0.060 | 0.011 | 4.94 × 10−8 |
| rs113597793 | C | T | 0.060 | 0.011 | 4.92 × 10−8 |
| rs7036368 | A | C | 0.061 | 0.011 | 3.87 × 10−8 |
| rs7049100 | G | A | 0.060 | 0.011 | 4.58 × 10−8 |
| rs7040385 | T | A | 0.060 | 0.011 | 4.43 × 10−8 |
| SNP | ALT | A1 | Beta | SE | p |
|---|---|---|---|---|---|
| rs6846781 | T | T | 0.052 | 0.009 | 4.22 × 10−8 |
| rs7690236 | T | T | 0.052 | 0.009 | 3.27 × 10−8 |
| rs28378450 | A | A | 0.052 | 0.009 | 3.29 × 10−8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, B.; Chu, X.; Yang, X.; Wen, Y.; Jia, Y.; Liang, C.; Yao, Y.; Ye, J.; Cheng, S.; Liu, L.; et al. Dietary Habit Is Associated with Depression and Intelligence: An Observational and Genome-Wide Environmental Interaction Analysis in the UK Biobank Cohort. Nutrients 2021, 13, 1150. https://doi.org/10.3390/nu13041150
Cheng B, Chu X, Yang X, Wen Y, Jia Y, Liang C, Yao Y, Ye J, Cheng S, Liu L, et al. Dietary Habit Is Associated with Depression and Intelligence: An Observational and Genome-Wide Environmental Interaction Analysis in the UK Biobank Cohort. Nutrients. 2021; 13(4):1150. https://doi.org/10.3390/nu13041150
Chicago/Turabian StyleCheng, Bolun, Xiaomeng Chu, Xuena Yang, Yan Wen, Yumeng Jia, Chujun Liang, Yao Yao, Jing Ye, Shiqiang Cheng, Li Liu, and et al. 2021. "Dietary Habit Is Associated with Depression and Intelligence: An Observational and Genome-Wide Environmental Interaction Analysis in the UK Biobank Cohort" Nutrients 13, no. 4: 1150. https://doi.org/10.3390/nu13041150