
Herbal Remedies as Potential in Cartilage Tissue Engineering: An Overview of New Therapeutic Approaches and Strategies

 ,
, 

Abstract
1. Introduction
Data Acquisition
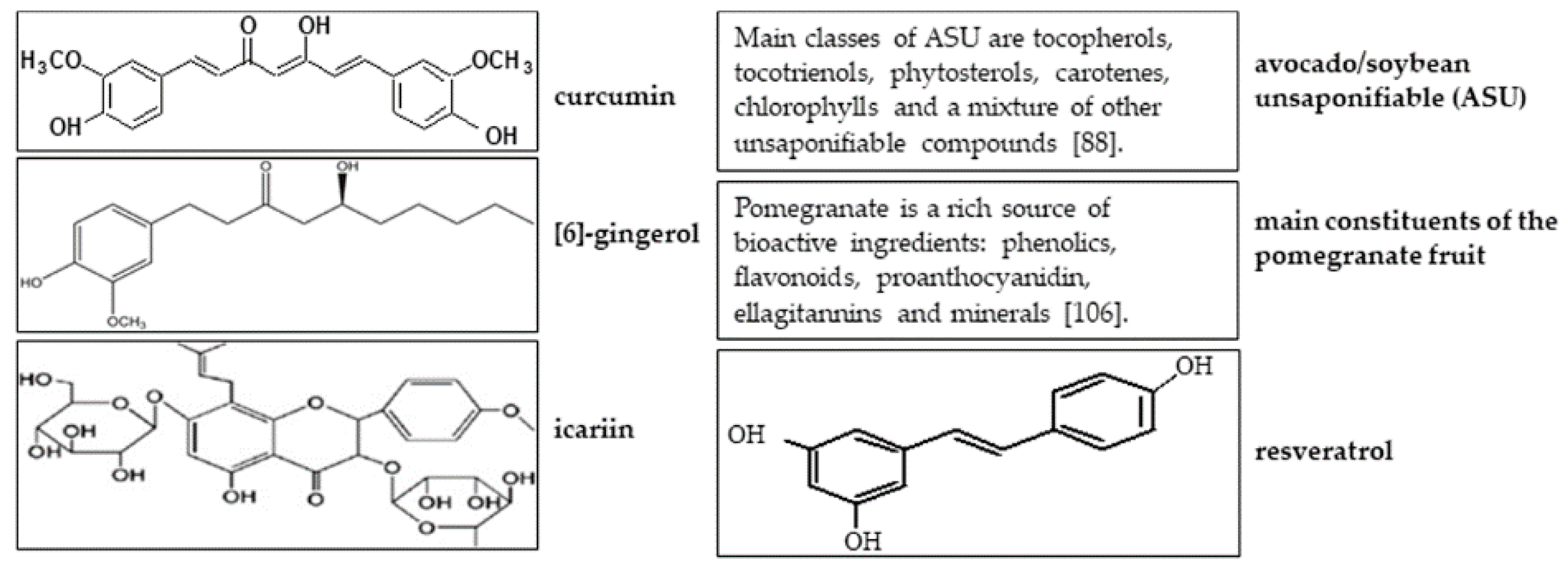
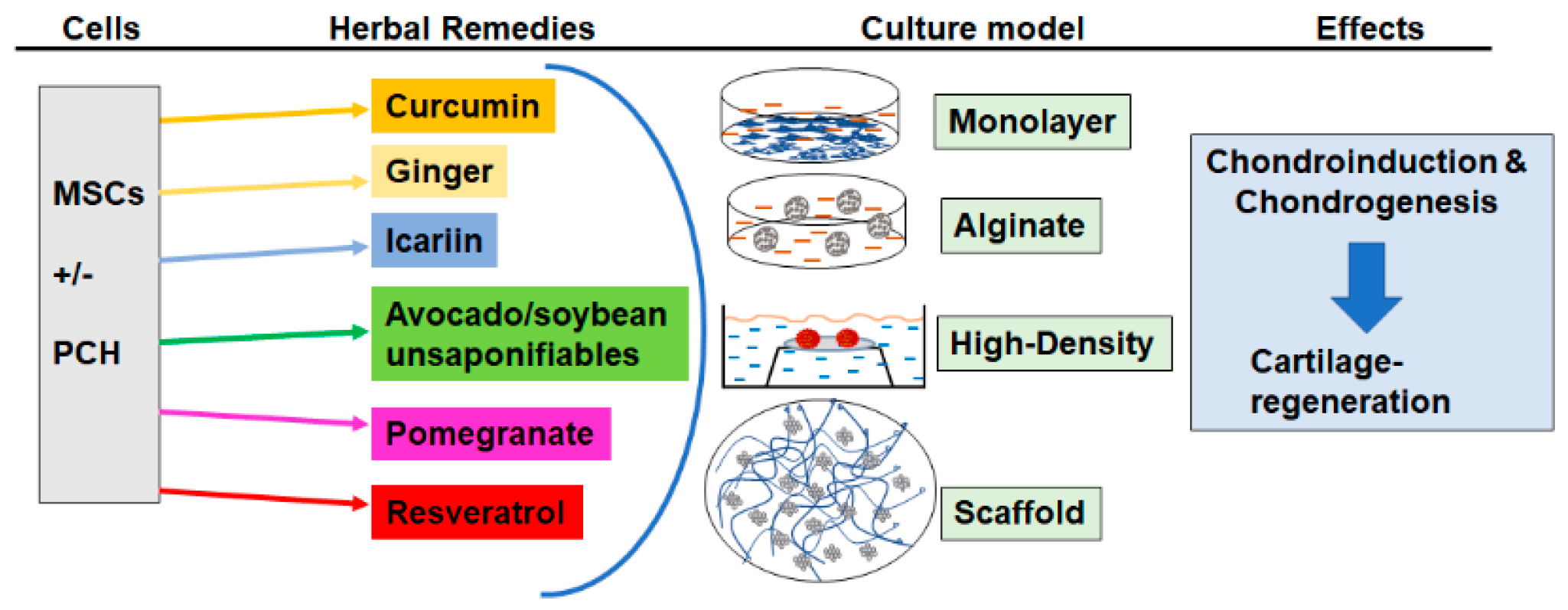
2. Curcumin
3. Ginger
4. Icariin
5. Avocado/Soybean Unsaponifiables
6. Pomegranate
7. Resveratrol
8. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3D | Three-Dimensional |
| ACI | Autologous Chondrocyte Implantation |
| ACLT | Anterior Cruciate Ligament Transection |
| AGC | Aggrecan |
| AGEs | Advanced Glycation End products |
| AMPK/mTOR | Adenosine Monophosphate-activated Protein Kinase/rapamycin |
| AP-1 | Activator Protein-1 |
| ASU | Avocado/Soybean Unsaponifiable (ASU) |
| BALB/c | Bagg Albino Laboratory-Bred strain c of the house mouse |
| BFP-MSCs | Buccal Fat Pad-Derived Mesenchymal Stem Cells |
| BMP-6 | Bone Morphogenic Protein-6 |
| CIA | Collagen-Induced Arthritis |
| COL2A1 | Collagen type II |
| COX-2 | Cyclooxygenase-2 |
| CS | Chitosan |
| CSG | Chitosan-Gelatin Scaffolds |
| CSPGs | Cartilage-Specific Proteoglycans |
| DJD | Degenerative Joint Disease |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| DMM | Destabilization of the Medial Meniscus |
| DMSO | Dimethyl Sulphoxide |
| ECM | Extracellular Matrix |
| EFSA | European Food Safety Authority |
| EM | Electron Microscopy |
| EMSA | Electrophoretic Mobility Shift Assay |
| FDA | US Food and Drug Administration |
| GA | Glutaraldehyde |
| GAGs | Glycosaminoglycans |
| GC | Gas Chromatography |
| Gel/HA | Gelatin/Hyaluronic Acid |
| GlcN | Glucosamine |
| GRE | Ginger Root Extract |
| H2O2 | Hydrogen peroxide |
| hADSCs | Human Adipose Tissue-Derived Mesenchymal Stem Cells |
| HIF | Hypoxia-Inducible Factor |
| HPG | Hyperbranched Polyglycerol |
| HPLS | High Pressure Liquid Chromatography |
| Ica | Icariin |
| IGF-1 | Insulin-Like Growth Factor-1 |
| IHC | Immunohistochemistry |
| Ihh | Indian Hedgehog |
| IkBα | Inhibitor of Kappa B Alpha |
| IKKβ | inhibitor of Nuclear Factor Kappa-B Kinase Subunit Beta (IKKβ), |
| IL-1β | Interleukin-1β |
| IGFBP-3 | Insulin-like-growth-factor-binding-protein-3 |
| iNOS | Inducible Nitric Oxide Synthase |
| JNK/ERK/AP-1 | c-Jun N-terminal kinase/Extracellular signal-Regulated Kinase/Activator Protein -1 pathways |
| LA | α-Lipoic Acid |
| LPS | Lipopolysaccharides |
| LTB | Leukotriene B |
| MAPK | Mitogen-Activated Protein Kinase |
| MKK3 | Mitogen-activated protein kinase kinase 3 |
| MMP | Matrix Metalloproteinase |
| MSCs | Mesenchymal Stem Cells |
| MTT | ((3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyl-Tetrazolium-Bromid)) |
| NF-κB | Nuclear Factor-Kappa B |
| NO | Nitric Oxide |
| NSAIDs | Non-Steroidal Anti-Inflammatory Drugs |
| OA | Osteoarthritis |
| Oxi-HA/Res | Oxidized Hyaluronic Acid/Resveratrol |
| PAI | Plasminogen Activator Inhibitor |
| PARP | Poly(ADP-ribose)-Polymerasen |
| PCH | Primary Human Chondrocytes |
| PCR | Polymerase Chain Reaction. |
| PFE | Pomegranate Fruit Extract |
| PG | Proteoglycan |
| PGE2 | Prostaglandin E2 |
| PHrP | Parathyroid Hormone-Related Proteins |
| PLA | Poly(Lactic Acid) |
| PLLA | Poly(L-lactic acid) |
| PVA | Poly(Vinyl Alcohol) |
| RA | Rheumatoid Arthritis |
| ROS | Reactive Oxygen Species |
| RUNX-2 | Runt-related transcription factor 2 |
| S-35 | 35-sulfate into proteoglycans |
| SIRT1 | Histone Deacetylase Sirtuin-1 |
| Sirtuin-1 | Silent Mating-Type Information Regulation-1 |
| SNP | sodium nitroprusside |
| SOX9 | (Sex-Determining Region Y)-Box |
| TE | Tissue Engineering |
| TGF-β | Transforming Growth Factor-β |
| TGF-βRI | Transforming growth factor beta receptor I |
| TNF-α | Tumor-Necrosis Factor |
| VEGF | Vascular Endothelial Growth Factor |
| WB | Western Immunoblotting |
References
- Aigner, T.; Rose, J.; Martin, J.; Buckwalter, J. Aging theories of primary osteoarthritis: From epidemiology to molecular biology. Rejuvenation Res. 2004, 7, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, R.C.; Felson, D.T.; Helmick, C.G.; Arnold, L.M.; Choi, H.; Deyo, R.A.; Gabriel, S.; Hirsch, R.; Hochberg, M.C.; Hunder, G.G.; et al. Estimates of the prevalence of arthritis and other rheumatic conditions in the United States. Part II. Arthritis Rheum. 2008, 58, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Cushnaghan, J.; Dieppe, P. Study of 500 patients with limb joint osteoarthritis. I. Analysis by age, sex, and distribution of symptomatic joint sites. Ann. Rheum. Dis. 1991, 50, 8–13. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, T.W.; McCabe, P.S.; McBeth, J. Update on the epidemiology, risk factors and disease outcomes of osteoarthritis. Best Pr. Res. Clin. Haematol. 2018, 32, 312–326. [Google Scholar] [CrossRef]
- Mobasheri, A. Intersection of inflammation and herbal medicine in the treatment of osteoarthritis. Curr. Rheumatol. Rep. 2012, 14, 604–616. [Google Scholar] [CrossRef]
- Roach, H.I.; Aigner, T.; Soder, S.; Haag, J.; Welkerling, H. Pathobiology of osteoarthritis: Pathomechanisms and potential therapeutic targets. Curr. Drug Targets 2007, 8, 271–282. [Google Scholar] [CrossRef]
- Goldring, M.B.; Goldring, S.R. Osteoarthritis. J. Cell. Physiol. 2007, 213, 626–634. [Google Scholar] [CrossRef]
- Sutton, S.; Clutterbuck, A.; Harris, P.; Gent, T.; Freeman, S.; Foster, N.; Barrett-Jolley, R.; Mobasheri, A. The contribution of the synovium, synovial derived inflammatory cytokines and neuropeptides to the pathogenesis of osteoarthritis. Vet. J. (Lond. Engl. 1997) 2009, 179, 10–24. [Google Scholar] [CrossRef]
- Ilyin, S.E.; Belkowski, S.M.; Plata-Salamán, C.R. Biomarker discovery and validation: Technologies and integrative approaches. Trends Biotechnol. 2004, 22, 411–416. [Google Scholar] [CrossRef]
- Buckwalter, J.A.; Mankin, H.J. Articular cartilage: Degeneration and osteoarthritis, repair, regeneration, and transplantation. Instr. Course Lect. 1998, 47, 487–504. [Google Scholar]
- Mobasheri, A.; Airley, R.; Foster, C.S.; Schulze-Tanzil, G.; Shakibaei, M. Post-genomic applications of tissue microarrays: Basic research, prognostic oncology, clinical genomics and drug discovery. Histol. Histopathol. 2004, 19, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Dieppe, P.A.; Lohmander, L.S. Pathogenesis and management of pain in osteoarthritis. Lancet (Lond. Engl.) 2005, 365, 965–973. [Google Scholar] [CrossRef]
- Messina, O.D.; Vidal Wilman, M.; Vidal Neira, L.F. Nutrition, osteoarthritis and cartilage metabolism. Aging Clin. Exp. Res. 2019, 31, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; Allaway, D.; Nebrich, S.; Mobasheri, A. Botanical extracts from rosehip (Rosa canina), willow bark (Salix alba), and nettle leaf (Urtica dioica) suppress IL-1β-Induced NF-κB activation in canine articular chondrocytes. Evid. -Based Complementary Altern. Med. Ecam 2012, 2012, 509383. [Google Scholar] [CrossRef] [PubMed]
- Cancedda, R.; Dozin, B.; Giannoni, P.; Quarto, R. Tissue engineering and cell therapy of cartilage and bone. Matrix Biol. J. Int. Soc. Matrix Biol. 2003, 22, 81–91. [Google Scholar] [CrossRef]
- Vachon, A.; Bramlage, L.R.; Gabel, A.A.; Weisbrode, S. Evaluation of the repair process of cartilage defects of the equine third carpal bone with and without subchondral bone perforation. Am. J. Vet. Res. 1986, 47, 2637–2645. [Google Scholar]
- Kuettner, K.E. Biochemistry of articular cartilage in health and disease. Clin. Biochem. 1992, 25, 155–163. [Google Scholar] [CrossRef]
- Qi, W.N.; Scully, S.P. Type II collagen modulates the composition of extracellular matrix synthesized by articular chondrocytes. J. Orthop. Res.: Off. Publ. Orthop. Res. Soc. 2003, 21, 282–289. [Google Scholar] [CrossRef]
- Mobasheri, A.; Csaki, C.; Clutterbuck, A.L.; Rahmanzadeh, M.; Shakibaei, M. Mesenchymal stem cells in connective tissue engineering and regenerative medicine: Applications in cartilage repair and osteoarthritis therapy. Histol. Histopathol. 2009, 24, 347–366. [Google Scholar] [CrossRef]
- Csaki, C.; Schneider, P.R.; Shakibaei, M. Mesenchymal stem cells as a potential pool for cartilage tissue engineering. Ann. Anat. Anat. Anz. Off. Organ Anat. Ges. 2008, 190, 395–412. [Google Scholar] [CrossRef]
- Huselstein, C.; Li, Y.; He, X. Mesenchymal stem cells for cartilage engineering. Bio-Med. Mater. Eng. 2012, 22, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Zhao, M.; Mundy, G.R. Bone morphogenetic proteins. Growth Factors (ChurSwitz.) 2004, 22, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Kabiri, A.; Esfandiari, E.; Hashemibeni, B.; Kazemi, M.; Mardani, M.; Esmaeili, A. Effects of FGF-2 on human adipose tissue derived adult stem cells morphology and chondrogenesis enhancement in Transwell culture. Biochem. Biophys. Res. Commun. 2012, 424, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.A.; Scherb, M.B.; Lembke, L.A.; Buckwalter, J.A. Damage control mechanisms in articular cartilage: The role of the insulin-like growth factor I axis. Iowa Orthop. J. 2000, 20, 1–10. [Google Scholar]
- Hutmacher, D.W. Scaffolds in tissue engineering bone and cartilage. Biomaterials 2000, 21, 2529–2543. [Google Scholar] [CrossRef]
- Loeser, R.F. Growth factor regulation of chondrocyte integrins. Differential effects of insulin-like growth factor 1 and transforming growth factor beta on alpha 1 beta 1 integrin expression and chondrocyte adhesion to type VI collagen. Arthritis Rheum. 1997, 40, 270–276. [Google Scholar] [CrossRef]
- Hollander, A.P.; Dickinson, S.C.; Sims, T.J.; Brun, P.; Cortivo, R.; Kon, E.; Marcacci, M.; Zanasi, S.; Borrione, A.; De Luca, C.; et al. Maturation of tissue engineered cartilage implanted in injured and osteoarthritic human knees. Tissue Eng. 2006, 12, 1787–1798. [Google Scholar] [CrossRef]
- Caplan, A.I.; Goldberg, V.M. Principles of tissue engineered regeneration of skeletal tissues. Clin. Orthop. Relat. Res. 1999, 367, S12–S16. [Google Scholar] [CrossRef]
- Solchaga, L.A.; Goldberg, V.M.; Caplan, A.I. Cartilage regeneration using principles of tissue engineering. Clin. Orthop. Relat. Res. 2001, 391, S161–S170. [Google Scholar] [CrossRef]
- Armiento, A.R.; Stoddart, M.J.; Alini, M.; Eglin, D. Biomaterials for articular cartilage tissue engineering: Learning from biology. Acta Biomater. 2018, 65, 1–20. [Google Scholar] [CrossRef]
- Caplan, A.I. Adult mesenchymal stem cells for tissue engineering versus regenerative medicine. J. Cell. Physiol. 2007, 213, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Grevenstein, D.; Mamilos, A.; Schmitt, V.H.; Niedermair, T.; Wagner, W.; Kirkpatrick, C.J.; Brochhausen, C. Excellent histological results in terms of articular cartilage regeneration after spheroid-based autologous chondrocyte implantation (ACI). Knee Surg. Sports Traumatol. Arthrosc. Off. J. Esska 2020. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Lu, Z.; Cai, P.; Zheng, L.; Zhao, J. Salidroside enhances proliferation and maintains phenotype of articular chondrocytes for autologous chondrocyte implantation (ACI) via TGF-beta/Smad3 Signal. Biomed. Pharm. Biomed. Pharmacother. 2020, 122, 109388. [Google Scholar] [CrossRef] [PubMed]
- Csaki, C.; Mobasheri, A.; Shakibaei, M. Synergistic chondroprotective effects of curcumin and resveratrol in human articular chondrocytes: Inhibition of IL-1beta-induced NF-kappaB-mediated inflammation and apoptosis. Arthritis Res. Ther. 2009, 11, R165. [Google Scholar] [CrossRef]
- Mobasheri, A.; Henrotin, Y.; Biesalski, H.K.; Shakibaei, M. Scientific evidence and rationale for the development of curcumin and resveratrol as nutraceutricals for joint health. Int. J. Mol. Sci. 2012, 13, 4202–4232. [Google Scholar] [CrossRef]
- Schulze-Tanzil, G.; Mobasheri, A.; Sendzik, J.; John, T.; Shakibaei, M. Effects of curcumin (diferuloylmethane) on nuclear factor kappaB signaling in interleukin-1beta-stimulated chondrocytes. Ann. N.Y. Acad. Sci. 2004, 1030, 578–586. [Google Scholar] [CrossRef]
- Shakibaei, M.; John, T.; Schulze-Tanzil, G.; Lehmann, I.; Mobasheri, A. Suppression of NF-kappaB activation by curcumin leads to inhibition of expression of cyclo-oxygenase-2 and matrix metalloproteinase-9 in human articular chondrocytes: Implications for the treatment of osteoarthritis. Biochem. Pharm. 2007, 73, 1434–1445. [Google Scholar] [CrossRef]
- Ameye, L.G.; Chee, W.S. Osteoarthritis and nutrition. From nutraceuticals to functional foods: A systematic review of the scientific evidence. Arthritis Res. Ther. 2006, 8, R127. [Google Scholar] [CrossRef]
- Henrotin, Y.; Lambert, C.; Couchourel, D.; Ripoll, C.; Chiotelli, E. Nutraceuticals: Do they represent a new era in the management of osteoarthritis? - a narrative review from the lessons taken with five products. Osteoarthr. Cartil. 2011, 19, 1–21. [Google Scholar] [CrossRef]
- Kalra, E.K. Nutraceutical-definition and introduction. Aaps Pharmsci 2003, 5, E25. [Google Scholar] [CrossRef]
- Hak, A.E.; Choi, H.K. Lifestyle and gout. Curr. Opin. Rheumatol. 2008, 20, 179–186. [Google Scholar] [CrossRef]
- Sale, J.E.; Gignac, M.; Hawker, G. The relationship between disease symptoms, life events, coping and treatment, and depression among older adults with osteoarthritis. J. Rheumatol. 2008, 35, 335–342. [Google Scholar]
- Henrotin, Y.; Clutterbuck, A.L.; Allaway, D.; Lodwig, E.M.; Harris, P.; Mathy-Hartert, M.; Shakibaei, M.; Mobasheri, A. Biological actions of curcumin on articular chondrocytes. Osteoarthr. Cartil. 2010, 18, 141–149. [Google Scholar] [CrossRef]
- Schulze-Tanzil, G.; de Souza, P.H.; Behnke, B.; Klingelhoefer, S.; Scheid, A.; Shakibaei, M. Effects of the antirheumatic remedy hox alpha-a new stinging nettle leaf extract-on matrix metalloproteinases in human chondrocytes in vitro. Histol. Histopathol. 2002, 17, 477–485. [Google Scholar] [CrossRef]
- Shen, C.L.; Smith, B.J.; Lo, D.F.; Chyu, M.C.; Dunn, D.M.; Chen, C.H.; Kwun, I.S. Dietary polyphenols and mechanisms of osteoarthritis. Nutr. Biochem. 2012, 23, 1367–1377. [Google Scholar] [CrossRef]
- Long, L.; Soeken, K.; Ernst, E. Herbal medicines for the treatment of osteoarthritis: A systematic review. Rheumatology (Oxf. Engl.) 2001, 40, 779–793. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Bordoloi, D.; Padmavathi, G.; Monisha, J.; Roy, N.K.; Prasad, S.; Aggarwal, B.B. Curcumin, the golden nutraceutical: Multitargeting for multiple chronic diseases. Br. J. Pharm.. 2017, 174, 1325–1348. [Google Scholar] [CrossRef]
- Buhrmann, C.; Shayan, P.; Aggarwal, B.B.; Shakibaei, M. Evidence that TNF-beta (lymphotoxin alpha) can activate the inflammatory environment in human chondrocytes. Arthritis Res. Ther. 2013, 15, R202. [Google Scholar] [CrossRef]
- Shakibaei, M.; Schulze-Tanzil, G.; John, T.; Mobasheri, A. Curcumin protects human chondrocytes from IL-l1beta-induced inhibition of collagen type II and beta1-integrin expression and activation of caspase-3: An immunomorphological study. Ann. Anat. Anat. Anz. Off. Organ Anat. Ges. 2005, 187, 487–497. [Google Scholar] [CrossRef]
- Wang, J.; Ma, J.; Gu, J.H.; Wang, F.Y.; Shang, X.S.; Tao, H.R.; Wang, X. Regulation of type II collagen, matrix metalloproteinase-13 and cell proliferation by interleukin-1beta is mediated by curcumin via inhibition of NF-kappaB signaling in rat chondrocytes. Mol. Med. Rep. 2017, 16, 1837–1845. [Google Scholar] [CrossRef]
- Clutterbuck, A.L.; Mobasheri, A.; Shakibaei, M.; Allaway, D.; Harris, P. Interleukin-1β–Induced extracellular matrix degradation and glycosaminoglycan release is inhibited by curcumin in an explant model of cartilage inflammation. Ann. N.Y. Acad. Sci. 2009, 1171, 428. [Google Scholar] [CrossRef] [PubMed]
- Toegel, S.; Wu, S.Q.; Piana, C.; Unger, F.M.; Wirth, M.; Goldring, M.B.; Gabor, F.; Viernstein, H. Comparison between chondroprotective effects of glucosamine, curcumin, and diacerein in IL-1beta-stimulated C-28/I2 chondrocytes. Osteoarthr. Cartil. 2008, 16, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Mobasheri, A.; Matis, U.; Shakibaei, M. Curcumin mediated suppression of nuclear factor-kappaB promotes chondrogenic differentiation of mesenchymal stem cells in a high-density co-culture microenvironment. Arthritis Res. Ther. 2010, 12, R127. [Google Scholar] [CrossRef]
- Chen, Y.; Lin, J.; Fei, Y.; Wang, H.; Gao, W. Preparation and characterization of electrospinning PLA/curcumin composite membranes. Fibers Polym. 2010, 11, 1128–1131. [Google Scholar] [CrossRef]
- Perumal, G.; Pappuru, S.; Chakraborty, D.; Maya Nandkumar, A.; Chand, D.K.; Doble, M. Synthesis and characterization of curcumin loaded PLA-Hyperbranched polyglycerol electrospun blend for wound dressing applications. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 76, 1196–1204. [Google Scholar] [CrossRef]
- Golchin, A.; Hosseinzadeh, S.; Staji, M.; Soleimani, M.; Ardeshirylajimi, A.; Khojasteh, A. Biological behavior of the curcumin incorporated chitosan/poly(vinyl alcohol) nanofibers for biomedical applications. J. Cell. Biochem. 2019, 120, 15410–15421. [Google Scholar] [CrossRef]
- Kim, D.K.; In Kim, J.; Sim, B.R.; Khang, G. Bioengineered porous composite curcumin/silk scaffolds for cartilage regeneration. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 78, 571–578. [Google Scholar] [CrossRef]
- Buhrmann, C.; Popper, B.; Aggarwal, B.B.; Shakibaei, M. Resveratrol downregulates inflammatory pathway activated by lymphotoxin alpha (TNF-beta) in articular chondrocytes: Comparison with TNF-alpha. PLoS ONE 2017, 12, e0186993. [Google Scholar] [CrossRef]
- Ali, B.H.; Blunden, G.; Tanira, M.O.; Nemmar, A. Some phytochemical, pharmacological and toxicological properties of ginger (Zingiber officinale Roscoe): A review of recent research. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2008, 46, 409–420. [Google Scholar] [CrossRef]
- Tjendraputra, E.; Tran, V.H.; Liu-Brennan, D.; Roufogalis, B.D.; Duke, C.C. Effect of ginger constituents and synthetic analogues on cyclooxygenase-2 enzyme in intact cells. Bioorg. Chem. 2001, 29, 156–163. [Google Scholar] [CrossRef]
- Bartels, E.M.; Folmer, V.N.; Bliddal, H.; Altman, R.D.; Juhl, C.; Tarp, S.; Zhang, W.; Christensen, R. Efficacy and safety of ginger in osteoarthritis patients: A meta-analysis of randomized placebo-controlled trials. Osteoarthr. Cartil. 2015, 23, 13–21. [Google Scholar] [CrossRef]
- Grzanna, R.; Lindmark, L.; Frondoza, C.G. Ginger-an herbal medicinal product with broad anti-inflammatory actions. J. Med. Food 2005, 8, 125–132. [Google Scholar] [CrossRef]
- van Breemen, R.B.; Tao, Y.; Li, W. Cyclooxygenase-2 inhibitors in ginger (Zingiber officinale). Fitoterapia 2011, 82, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Kiuchi, F.; Iwakami, S.; Shibuya, M.; Hanaoka, F.; Sankawa, U. Inhibition of prostaglandin and leukotriene biosynthesis by gingerols and diarylheptanoids. Chem. Pharm. Bull. 1992, 40, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Phan, P.V.; Sohrabi, A.; Polotsky, A.; Hungerford, D.S.; Lindmark, L.; Frondoza, C.G. Ginger extract components suppress induction of chemokine expression in human synoviocytes. J. Altern. Complementary Med. (N.Y.) 2005, 11, 149–154. [Google Scholar] [CrossRef]
- Shen, C.L.; Hong, K.J.; Kim, S.W. Effects of ginger (Zingiber officinale Rosc.) on decreasing the production of inflammatory mediators in sow osteoarthrotic cartilage explants. J. Med. Food 2003, 6, 323–328. [Google Scholar] [CrossRef]
- Shen, C.L.; Hong, K.J.; Kim, S.W. Comparative effects of ginger root (Zingiber officinale Rosc.) on the production of inflammatory mediators in normal and osteoarthrotic sow chondrocytes. J. Med. Food 2005, 8, 149–153. [Google Scholar] [CrossRef]
- Ruangsuriya, J.; Budprom, P.; Viriyakhasem, N.; Kongdang, P.; Chokchaitaweesuk, C.; Sirikaew, N.; Chomdej, S.; Nganvongpanit, K.; Ongchai, S. Suppression of cartilage degradation by zingerone involving the p38 and JNK MAPK signaling pathway. Planta Med. 2017, 83, 268–276. [Google Scholar] [CrossRef]
- Hosseinzadeh, A.; Bahrampour Juybari, K.; Fatemi, M.J.; Kamarul, T.; Bagheri, A.; Tekiyehmaroof, N.; Sharifi, A.M. Protective effect of ginger (Zingiber officinale Roscoe) extract against oxidative stress and mitochondrial apoptosis induced by interleukin-1β in cultured chondrocytes. Cells Tissues Organs 2017, 204, 241–250. [Google Scholar] [CrossRef]
- Ming, L.G.; Chen, K.M.; Xian, C.J. Functions and action mechanisms of flavonoids genistein and icariin in regulating bone remodeling. J. Cell. Physiol. 2013, 228, 513–521. [Google Scholar] [CrossRef]
- Mi, B.; Wang, J.; Liu, Y.; Liu, J.; Hu, L.; Panayi, A.C.; Liu, G.; Zhou, W. Icariin activates autophagy via Down-Regulation of the NF-κB Signaling-Mediated apoptosis in chondrocytes. Front. Pharm. 2018, 9, 605. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, T.; Huang, Y.; Wismeijer, D.; Liu, Y. Icariin: Does it have an osteoinductive potential for bone tissue engineering? Phytother. Res. Ptr 2014, 28, 498–509. [Google Scholar] [CrossRef]
- Wang, Z.C.; Sun, H.J.; Li, K.H.; Fu, C.; Liu, M.Z. Icariin promotes directed chondrogenic differentiation of bone marrow mesenchymal stem cells but not hypertrophy in vitro. Exp. Med. 2014, 8, 1528–1534. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, X.; Li, K.F.; Li, D.X.; Xiao, Y.M.; Fan, Y.J.; Zhang, X.D. Icariin promotes extracellular matrix synthesis and gene expression of chondrocytes in vitro. Phytother. Res. Ptr 2012, 26, 1385–1392. [Google Scholar] [CrossRef]
- Liu, M.H.; Sun, J.S.; Tsai, S.W.; Sheu, S.Y.; Chen, M.H. Icariin protects murine chondrocytes from lipopolysaccharide-induced inflammatory responses and extracellular matrix degradation. Nutr. Res. (N.Y.) 2010, 30, 57–65. [Google Scholar] [CrossRef]
- Xu, C.Q.; Liu, B.J.; Wu, J.F.; Xu, Y.C.; Duan, X.H.; Cao, Y.X.; Dong, J.C. Icariin attenuates LPS-induced acute inflammatory responses: Involvement of PI3K/Akt and NF-kappaB signaling pathway. Eur. J. Pharm. 2010, 642, 146–153. [Google Scholar] [CrossRef]
- Liu, N.; Zhang, T.; Cao, B.R.; Luan, F.Y.; Liu, R.X.; Yin, H.R.; Wang, W.B. Icariin possesses chondroprotective efficacy in a rat model of dexamethasone-induced cartilage injury through the activation of miR-206 targeting of cathepsin K. Int. J. Med. Mol. Adv. Sci. 2018, 41, 1039–1047. [Google Scholar] [CrossRef]
- Wang, W.; Sun, L.; Zhang, P.; Song, J.; Liu, W. An anti-inflammatory cell-free collagen/resveratrol scaffold for repairing osteochondral defects in rabbits. Acta Biomater. 2014, 10, 4983–4995. [Google Scholar] [CrossRef]
- Kankala, R.K.; Lu, F.J.; Liu, C.G.; Zhang, S.S.; Chen, A.Z.; Wang, S.B. Effect of Icariin on Engineered 3D-Printed Porous Scaffolds for Cartilage Repair. Materials. 2018, 11, 1390. [Google Scholar] [CrossRef]
- Yang, J.; Liu, Y.; He, L.; Wang, Q.; Wang, L.; Yuan, T.; Xiao, Y.; Fan, Y.; Zhang, X. Icariin conjugated hyaluronic acid/collagen hydrogel for osteochondral interface restoration. Acta Biomater. 2018, 74, 156–167. [Google Scholar] [CrossRef]
- Li, D.; Yuan, T.; Zhang, X.; Xiao, Y.; Wang, R.; Fan, Y.; Zhang, X. Icariin: A potential promoting compound for cartilage tissue engineering. Osteoarthr. Cartil. 2012, 20, 1647–1656. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, F.; He, Q.; Wang, J.; Shiu, H.T.; Shu, Y.; Tsang, W.P.; Liang, S.; Zhao, K.; Wan, C. Flavonoid compound icariin activates Hypoxia Inducible Factor-1α in chondrocytes and promotes articular cartilage repair. PLoS ONE 2016, 11, e0148372. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhang, Y.; Huang, Y. Icariin reduces cartilage degeneration in a mouse model of osteoarthritis and is associated with the changes in expression of Indian Hedgehog and Parathyroid Hormone-Related protein. Med Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 6695–6706. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, J.; Luo, Z.; Li, D.; Lu, J.; Wang, Q.; Xiao, Y.; Zhang, X. Development of an injectable thiolated icariin functionalized collagen/hyaluronic hydrogel to promote cartilage formation in vitro and in vivo. J. Mater. Chem. B 2019, 7, 2845–2854. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, D.; Wu, C.; Liu, A.; Zhang, C.; Jiao, J.; Shang, M. Icariin-conditioned serum engineered with hyaluronic acid promote repair of articular cartilage defects in rabbit knees. BMC Complementary Altern. Med. 2019, 19, 155. [Google Scholar] [CrossRef]
- Cameron, M.; Chrubasik, S. Oral herbal therapies for treating osteoarthritis. Cochrane Database Syst. Rev. 2014, Cd002947. [Google Scholar] [CrossRef]
- Salehi, B.; Rescigno, A.; Dettori, T.; Calina, D.; Docea, A.O.; Singh, L.; Cebeci, F.; Ozcelik, B.; Bhia, M.; Dowlati Beirami, A.; et al. Avocado-Soybean unsaponifiables: A panoply of potentialities to be exploited. Biomolecules 2020, 10, 130. [Google Scholar] [CrossRef]
- Henrotin, Y.E.; Labasse, A.H.; Jaspar, J.M.; De Groote, D.D.; Zheng, S.X.; Guillou, G.B.; Reginster, J.Y. Effects of three avocado/soybean unsaponifiable mixtures on metalloproteinases, cytokines and prostaglandin E2 production by human articular chondrocytes. Clin. Rheumatol. 1998, 17, 31–39. [Google Scholar] [CrossRef]
- Henrotin, Y.E.; Sanchez, C.; Deberg, M.A.; Piccardi, N.; Guillou, G.B.; Msika, P.; Reginster, J.Y. Avocado/soybean unsaponifiables increase aggrecan synthesis and reduce catabolic and proinflammatory mediator production by human osteoarthritic chondrocytes. J. Rheumatol. 2003, 30, 1825–1834. [Google Scholar]
- Khanna, D.; Sethi, G.; Ahn, K.S.; Pandey, M.K.; Kunnumakkara, A.B.; Sung, B.; Aggarwal, A.; Aggarwal, B.B. Natural products as a gold mine for arthritis treatment. Curr. Opin. Pharm. 2007, 7, 344–351. [Google Scholar] [CrossRef]
- Hashemibeni, B.; Valiani, A.; Esmaeli, M.; Kazemi, M.; Aliakbari, M.; Iranpour, F.G. Comparison of the efficacy of piascledine and transforming growth factor beta1 on chondrogenic differentiation of human adipose-derived stem cells in fibrin and fibrin-alginate scaffolds. Iran. J. Basic Med. Sci. 2018, 21, 212–218. [Google Scholar] [CrossRef]
- Altinel, L.; Saritas, Z.K.; Kose, K.C.; Pamuk, K.; Aksoy, Y.; Serteser, M. Treatment with unsaponifiable extracts of avocado and soybean increases TGF-beta1 and TGF-beta2 levels in canine joint fluid. Tohoku J. Exp. Med. 2007, 211, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Boumediene, K.; Felisaz, N.; Bogdanowicz, P.; Galera, P.; Guillou, G.B.; Pujol, J.P. Avocado/soya unsaponifiables enhance the expression of transforming growth factor beta1 and beta2 in cultured articular chondrocytes. Arthritis Rheum. 1999, 42, 148–156. [Google Scholar] [CrossRef]
- Maheu, E.; Mazières, B.; Valat, J.P.; Loyau, G.; Le Loët, X.; Bourgeois, P.; Grouin, J.M.; Rozenberg, S. Symptomatic efficacy of avocado/soybean unsaponifiables in the treatment of osteoarthritis of the knee and hip: A prospective, randomized, double-blind, placebo-controlled, multicenter clinical trial with a six-month treatment period and a two-month followup demonstrating a persistent effect. Arthritis Rheum. 1998, 41, 81–91. [Google Scholar] [CrossRef]
- Frondoza, C.G.; Fortuno, L.V.; Grzanna, M.W.; Ownby, S.L.; Au, A.Y.; Rashmir-Raven, A.M. α-Lipoic acid potentiates the anti-inflammatory activity of avocado/soybean unsaponifiables in chondrocyte cultures. Cartilage 2018, 9, 304–312. [Google Scholar] [CrossRef]
- Au, R.Y.; Al-Talib, T.K.; Au, A.Y.; Phan, P.V.; Frondoza, C.G. Avocado soybean unsaponifiables (ASU) suppress TNF-alpha, IL-1beta, COX-2, iNOS gene expression, and prostaglandin E2 and nitric oxide production in articular chondrocytes and monocyte/macrophages. Osteoarthr. Cartil. 2007, 15, 1249–1255. [Google Scholar] [CrossRef] [PubMed]
- Lippiello, L.; Nardo, J.V.; Harlan, R.; Chiou, T. Metabolic effects of avocado/soy unsaponifiables on articular chondrocytes. Evid.-Based Complementary Altern. Med.: Ecam 2008, 5, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Cake, M.A.; Read, R.A.; Guillou, B.; Ghosh, P. Modification of articular cartilage and subchondral bone pathology in an ovine meniscectomy model of osteoarthritis by avocado and soya unsaponifiables (ASU). Osteoarthr. Cartil. 2000, 8, 404–411. [Google Scholar] [CrossRef]
- Boileau, C.; Martel-Pelletier, J.; Caron, J.; Msika, P.; Guillou, G.B.; Baudouin, C.; Pelletier, J.P. Protective effects of total fraction of avocado/soybean unsaponifiables on the structural changes in experimental dog osteoarthritis: Inhibition of nitric oxide synthase and matrix metalloproteinase-13. Arthritis Res. Ther. 2009, 11, R41. [Google Scholar] [CrossRef]
- Fazelipour, S.; Jahromy, M.H.; Tootian, Z.; Kiaei, S.B.; Sheibani, M.T.; Talaee, N. The effect of chronic administration of methylphenidate on morphometric parameters of testes and fertility in male mice. J. Reprod. Infertil. 2012, 13, 232–236. [Google Scholar]
- Afaq, F.; Malik, A.; Syed, D.; Maes, D.; Matsui, M.S.; Mukhtar, H. Pomegranate fruit extract modulates UV-B-mediated phosphorylation of mitogen-activated protein kinases and activation of nuclear factor kappa B in normal human epidermal keratinocytes paragraph sign. Photochem. Photobiol. 2005, 81, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Gil, M.I.; Tomás-Barberán, F.A.; Hess-Pierce, B.; Holcroft, D.M.; Kader, A.A. Antioxidant activity of pomegranate juice and its relationship with phenolic composition and processing. J. Agric. Food. Chem. 2000, 48, 4581–4589. [Google Scholar] [CrossRef]
- Seeram, N.P.; Nair, M.G. Inhibition of lipid peroxidation and structure-activity-related studies of the dietary constituents anthocyanins, anthocyanidins, and catechins. J. Agric. Food. Chem. 2002, 50, 5308–5312. [Google Scholar] [CrossRef] [PubMed]
- Sreekumar, S.; Sithul, H.; Muraleedharan, P.; Azeez, J.M.; Sreeharshan, S. Pomegranate fruit as a rich source of biologically active compounds. Biomed. Res. Int. 2014, 2014, 686921. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; McClees, S.F.; Afaq, F. Pomegranate for prevention and treatment of cancer: An update. Molecules 2017, 22, 177. [Google Scholar] [CrossRef] [PubMed]
- Ajaikumar, K.B.; Asheef, M.; Babu, B.H.; Padikkala, J. The inhibition of gastric mucosal injury by Punicagranatum, L. (pomegranate) methanolic extract. J. Ethnopharmacol. 2005, 96, 171–176. [Google Scholar] [CrossRef]
- Akhtar, N.; Haqqi, T.M. Current nutraceuticals in the management of osteoarthritis: A review. Ther. Adv. Musculoskelet. Dis. 2012, 4, 181–207. [Google Scholar] [CrossRef]
- Seeram, N.P.; Adams, L.S.; Henning, S.M.; Niu, Y.; Zhang, Y.; Nair, M.G.; Heber, D. In vitro antiproliferative, apoptotic and antioxidant activities of punicalagin, ellagic acid and a total pomegranate tannin extract are enhanced in combination with other polyphenols as found in pomegranate juice. J. Nutr. Biochem. 2005, 16, 360–367. [Google Scholar] [CrossRef]
- Ahmed, S.; Wang, N.; Hafeez, B.B.; Cheruvu, V.K.; Haqqi, T.M. Punica granatum L. extract inhibits IL-1beta-induced expression of matrix metalloproteinases by inhibiting the activation of MAP kinases and NF-kappaB in human chondrocytes in vitro. J. Nutr. 2005, 135, 2096–2102. [Google Scholar] [CrossRef]
- Shukla, M.; Gupta, K.; Rasheed, Z.; Khan, K.A.; Haqqi, T.M. Bioavailable constituents/metabolites of pomegranate (Punica granatum L) preferentially inhibit COX2 activity ex vivo and IL-1beta-induced PGE2 production in human chondrocytes in vitro. J. Inflamm. (Lond. Engl.) 2008, 5, 9. [Google Scholar] [CrossRef]
- Haseeb, A.; Khan, N.M.; Ashruf, O.S.; Haqqi, T.M. A polyphenol-rich pomegranate fruit extract euppresses NF-κB and IL-6 expression by blocking the activation of IKKβ and NIK in primary human chondrocytes. Phytother. Res. Ptr 2017, 31, 778–782. [Google Scholar] [CrossRef]
- Rasheed, Z.; Akhtar, N.; Haqqi, T.M. Pomegranate extract inhibits the interleukin-1beta-induced activation of MKK-3, p38alpha-MAPK and transcription factor RUNX-2 in human osteoarthritis chondrocytes. Arthritis Res. Ther. 2010, 12, R195. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Khan, N.M.; Ashruf, O.S.; Haqqi, T.M. Inhibition of cartilage degradation and suppression of PGE (2) and MMPs expression by pomegranate fruit extract in a model of posttraumatic osteoarthritis. Nutrition (Burbank Los Angeles Cty. Calif.) 2017, 33, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hadipour-Jahromy, M.; Mozaffari-Kermani, R. Chondroprotective effects of pomegranate juice on monoiodoacetate-induced osteoarthritis of the knee joint of mice. Phytother. Res. Ptr 2010, 24, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Monsefi, M.; Parvin, F.; Talaei-Khozani, T. Effects of pomegranate extracts on cartilage, bone and mesenchymal cells of mouse fetuses. Br. J. Nutr. 2012, 107, 683–690. [Google Scholar] [CrossRef]
- Ghoochani, N.; Karandish, M.; Mowla, K.; Haghighizadeh, M.H.; Jalali, M.T. The effect of pomegranate juice on clinical signs, matrix metalloproteinases and antioxidant status in patients with knee osteoarthritis. J. Sci. Food Agric. 2016, 96, 4377–4381. [Google Scholar] [CrossRef]
- Shakibaei, M.; Harikumar, K.B.; Aggarwal, B.B. Resveratrol addiction: To die or not to die. Mol. Nutr. Food Res. 2009, 53, 115–128. [Google Scholar] [CrossRef]
- Takaoka, M. Resveratrol, a new phenolic compound, from Veratrum grandiflorum. Nippon Kagaku Kaishi 1939, 60, 1090–1100. [Google Scholar] [CrossRef]
- Borriello, A.; Bencivenga, D.; Caldarelli, I.; Tramontano, A.; Borgia, A.; Zappia, V.; Della Ragione, F. Resveratrol: From basic studies to bedside. Cancer Treat. Res. 2014, 159, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Csiszar, A. Anti-inflammatory effects of resveratrol: Possible role in prevention of age-related cardiovascular disease. Ann. N. Y. Acad. Sci. 2011, 1215, 117–122. [Google Scholar] [CrossRef]
- Csaki, C.; Keshishzadeh, N.; Fischer, K.; Shakibaei, M. Regulation of inflammation signalling by resveratrol in human chondrocytes in vitro. Biochem. Pharmacol. 2008, 75, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; Csaki, C.; Nebrich, S.; Mobasheri, A. Resveratrol suppresses interleukin-1beta-induced inflammatory signaling and apoptosis in human articular chondrocytes: Potential for use as a novel nutraceutical for the treatment of osteoarthritis. Biochem. Pharmacol. 2008, 76, 1426–1439. [Google Scholar] [CrossRef] [PubMed]
- Dave, M.; Attur, M.; Palmer, G.; Al-Mussawir, H.E.; Kennish, L.; Patel, J.; Abramson, S.B. The antioxidant resveratrol protects against chondrocyte apoptosis via effects on mitochondrial polarization and ATP production. Arthritis Rheum. 2008, 58, 2786–2797. [Google Scholar] [CrossRef]
- Kim, H.J.; Braun, H.J.; Dragoo, J.L. The effect of resveratrol on normal and osteoarthritic chondrocyte metabolism. Bone Jt. Res. 2014, 3, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.C.; Hung, L.F.; Wu, W.L.; Chang, D.M.; Huang, C.Y.; Lai, J.H.; Ho, L.J. Chondroprotective effects and mechanisms of resveratrol in advanced glycation end products-stimulated chondrocytes. Arthritis Res. Ther. 2010, 12, R167. [Google Scholar] [CrossRef]
- Meng, X.; Zhou, J.; Zhao, C.N.; Gan, R.Y.; Li, H.B. Health benefits and molecular mechanisms of resveratrol: A narrative review. Foods (Basel Switz.) 2020, 9, 340. [Google Scholar] [CrossRef]
- Frémont, L. Biological effects of resveratrol. Life Sci. 2000, 66, 663–673. [Google Scholar] [CrossRef]
- Pezzuto, J.M. Resveratrol: Twenty years of growth, development and controversy. Biomol. Ther. 2019, 27, 1–14. [Google Scholar] [CrossRef]
- Mobasheri, A.; Shakibaei, M. Osteogenic effects of resveratrol in vitro: Potential for the prevention and treatment of osteoporosis. Ann. N. Y. Acad. Sci. 2013, 1290, 59–66. [Google Scholar] [CrossRef]
- Xie, W.L.; Chipman, J.G.; Robertson, D.L.; Erikson, R.L.; Simmons, D.L. Expression of a mitogen-responsive gene encoding prostaglandin synthase is regulated by mRNA splicing. Proc. Natl. Acad. Sci. USA 1991, 88, 2692–2696. [Google Scholar] [CrossRef]
- Tseng, C.C.; Chen, Y.J.; Chang, W.A.; Tsai, W.C.; Ou, T.T.; Wu, C.C.; Sung, W.Y.; Yen, J.H.; Kuo, P.L. Dual Role of chondrocytes in rheumatoid arthritis: The chicken and the egg. Int. J. Mol. Sci. 2020, 21, 71. [Google Scholar] [CrossRef]
- Varela-Eirin, M.; Loureiro, J.; Fonseca, E.; Corrochano, S.; Caeiro, J.R.; Collado, M.; Mayan, M.D. Cartilage regeneration and ageing: Targeting cellular plasticity in osteoarthritis. Ageing Res. Rev. 2018, 42, 56–71. [Google Scholar] [CrossRef]
- Subbaramaiah, K.; Chung, W.J.; Michaluart, P.; Telang, N.; Tanabe, T.; Inoue, H.; Jang, M.; Pezzuto, J.M.; Dannenberg, A.J. Resveratrol inhibits cyclooxygenase-2 transcription and activity in phorbol ester-treated human mammary epithelial cells. J. Biol. Chem. 1998, 273, 21875–21882. [Google Scholar] [CrossRef] [PubMed]
- Im, H.J.; Li, X.; Chen, D.; Yan, D.; Kim, J.; Ellman, M.B.; Stein, G.S.; Cole, B.; Kc, R.; Cs-Szabo, G.; et al. Biological effects of the plant-derived polyphenol resveratrol in human articular cartilage and chondrosarcoma cells. J. Cell. Physiol. 2012, 227, 3488–3497. [Google Scholar] [CrossRef] [PubMed]
- Eo, S.H.; Cho, H.; Kim, S.J. Resveratrol inhibits nitric oxide-induced apoptosis via the NF-Kappa B pathway in rabbit articular chondrocytes. Biomol. Ther. 2013, 21, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Maepa, M.; Razwinani, M.; Motaung, S. Effects of resveratrol on collagen type II protein in the superficial and middle zone chondrocytes of porcine articular cartilage. J. Ethnopharmacol. 2016, 178, 25–33. [Google Scholar] [CrossRef]
- Elmali, N.; Esenkaya, I.; Harma, A.; Ertem, K.; Turkoz, Y.; Mizrak, B. Effect of resveratrol in experimental osteoarthritis in rabbits. Inflamm. Res. Off. J. Eur. Histamine Res. Soc. 2005, 54, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Cai, L.; Zhang, Y.; Cui, L.; Shen, G. Intra-articular resveratrol injection prevents osteoarthritis progression in a mouse model by activating SIRT1 and thereby silencing HIF-2α. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2015, 33, 1061–1070. [Google Scholar] [CrossRef]
- Buhrmann, C.; Busch, F.; Shayan, P.; Shakibaei, M. Sirtuin-1 (SIRT1) is required for promoting chondrogenic differentiation of mesenchymal stem cells. J. Biol. Chem. 2014, 289, 22048–22062. [Google Scholar] [CrossRef]
- Qin, N.; Wei, L.; Li, W.; Yang, W.; Cai, L.; Qian, Z.; Wu, S. Local intra-articular injection of resveratrol delays cartilage degeneration in C57BL/6 mice by inducing autophagy via AMPK/mTOR pathway. J. Pharmacol. Sci. 2017, 134, 166–174. [Google Scholar] [CrossRef]
- Jin, H.; Zhang, H.; Ma, T.; Lan, H.; Feng, S.; Zhu, H.; Ji, Y. Resveratrol protects murine chondrogenic ATDC5 cells ggainst LPS-Induced inflammatory injury through up-regulating MiR-146b. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 47, 972–980. [Google Scholar] [CrossRef]
- Wang, J.; Gao, J.S.; Chen, J.W.; Li, F.; Tian, J. Effect of resveratrol on cartilage protection and apoptosis inhibition in experimental osteoarthritis of rabbit. Rheumatol. Int. 2012, 32, 1541–1548. [Google Scholar] [CrossRef]
- Lei, M.; Liu, S.Q.; Liu, Y.L. Resveratrol protects bone marrow mesenchymal stem cell derived chondrocytes cultured on chitosan-gelatin scaffolds from the inhibitory effect of interleukin-1beta. Acta Pharmacol. Sin. 2008, 29, 1350–1356. [Google Scholar] [CrossRef] [PubMed]
- Sheu, S.Y.; Chen, W.S.; Sun, J.S.; Lin, F.H.; Wu, T. Biological characterization of oxidized hyaluronic acid/resveratrol hydrogel for cartilage tissue engineering. J. Biomed. Mater. Res. Part. A 2013, 101, 3457–3466. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Wang, L.; Li, H.; Ke, Y.; Yao, Y. Function of sustained released resveratrol on IL-1β-induced hBMSC MMP13 secretion inhibition and chondrogenic differentiation promotion. J. Biomater. Appl. 2016, 30, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; Mobasheri, A.; Buhrmann, C. Curcumin synergizes with resveratrol to stimulate the MAPK signaling pathway in human articular chondrocytes in vitro. Genes Nutr. 2011, 6, 171–179. [Google Scholar] [CrossRef] [PubMed]
- EFSA, European Food Safety Authority. Available online: www.efsa.europa.eu (accessed on 27 May 2020).
- FDA, US Food and Drug Administration. Available online: www.fda.gov (accessed on 27 May 2020).
- AHP, American Herbal Pharmacopoeia Monographs. Available online: https://herbal-ahp.org (accessed on 29 May 2020).
- ESCOP, European Scientific Cooperative on Phytotherapy Monographs. Available online: https://escop.com (accessed on 29 May 2020).



{kind=link}
{kind=link}
| First Author | References | Experimental Model | Study Type | Findings |
|---|---|---|---|---|
| Shakibaei | [37] | The effects of curcumin were investigated in primary human chondrocytes (PCH) treated with interleukin-1 beta (IL-1β), tumor necrosis factor alpha (TNF-α). 90% collagen type II (COL2A1), integrin β1, cyclooxygenase-2 (COX-2), matrix metalloproteinase 9 (MMP-9), and the phosphorylation and nuclear translocation of NF-kB were analyzed. | in vitro/human chondrocytes | Curcumin as a naturally occurring anti-inflammatory agent for the treatment of osteoarthritis (OA) by suppressing the nuclear factor kappa B (NF-kB)-mediated IL-1β/TNF-α catabolic pathways in PCH. |
| Buhrmann | [48] | PCH from human samples treated with diverse cytokines, such as TNF-β, TNF-α, and/or curcumin in monolayer cultures. | in vitro/human chondrocytes | TNF-β is involved in the inflammatory microenvironment in PCH, similar to TNF-α, which leads to an upregulation of NF-κB signaling. The curcumin-inhibited action of cytokines can be used to maintain the chondrogenic potential of chondrocytes. |
| Shakibaei | [49] | PCH were pretreated with IL-1β, in monolayer and 3D-cultures with curcumin. The ultrastructure of PCH was investigated by TEM. The production of COL2A1, the β1 integrin, caspase–3 was analyzed by immunohistochemistry (IHC), Western immunoblotting (WB). | in vitro/human chondrocytes | Curcumin exerted anti-apoptotic, anti-catabolic effects on IL-1β-stimulated PCH. Curcumin has potential as an adjunct nutraceutical chondroprotective agent for treating OA and related osteoarticular disorders. |
| Wang | [50] | Chondrocytes were treated with IL-1β and co-treated with various concentrations of curcumin. | in vitro/rat chondrocytes | Curcumin inhibited the IL-1β-induced activation of NF-κB by suppressing pIκBα and p65/RelA nuclear translocation, suppressed MMP-13, and upregulated COL2A1 expression. Curcumin as anti-inflammatory agent for the treatment of OA by inhibition of NF-κB signaling. |
| Buhrmann | [53] | Mesenchymal stem cells (MSCs) were cultured in a ratio of 1:1 with primary chondrocytes in 3D-high-density culture with/without curcumin and/or IL-1β. | in vitro/chondrocytes, MSCs | Curcumin established a microenvironment, in which the effects of cytokines were antagonized, thus facilitating chondrogenesis of MSCs. |
| Perumal | [55] | Curcumin incorporated electrospun nanofibers of a blend of PLA and HPG for wound healing applications. Both the polymers are fabricated by electrospinning technique. | in vitro/curcumin-loaded biomaterials | (Poly(lactic acid)) (PLA)/hyperbranched polyglycerol (HPG)/curcumin nanofibers can be a potential wound patch dressing for acute and chronic wound applications. |
| Golchin | [56] | Nanofibrous scaffolds fabricated by chitosan (CS), PVA, carbopol, and polycaprolactone using a dual electrospinning technique while curcumin incorporated inside of the CS/PVA fibers. | in vitro/buccal fat pad-derived mesenchymal stem cells (BFP-MSCs) | This nanofibrous scaffold has great potential, as simultaneous administration of curcumin and BFP-MSCs holds the promising potential for use in various regenerative medicine applications. |
| Kim | [57] | Scaffold composed of curcumin and silk fibroin as an appropriate clinical replacement for defected cartilage. The scaffolds were designed to have adequate pore size and mechanical strength for cartilage repair. | in vitro/in vivo rat chondrocytes | The curcumin/silk scaffold showed its biocompatibility, a favorable environment for post-transplant cartilage repair in vivo. A functional composite scaffold of curcumin/silk can be used in cartilage-tissue engineering (TE) and as a promising substrate for cartilage repair. |
| First Author | References | Experimental Model | Study Type | Findings |
|---|---|---|---|---|
| Bartels | [61] | Meta-analysis evaluating the clinical efficacy and safety of oral ginger consumption for the symptomatic treatment of OA. | in vivo/OA patients | Ginger lead to a statistically significant reduction in pain and OA symptoms compared to placebo control group and was well tolerated by the majority of OA patients. |
| Shen | [66,67] | Comparative effects of ginger on the synthesis of inflammatory mediators in normal and OA chondrocytes and cartilage explants | in vitro/chondrocytes, cartilage explants | Production of the pro-inflammatory mediators nitric oxide (NO) and prostaglandin E2 (PGE2) were significantly reduced with ginger extract in chondrocytes and cartilage explants. |
| Ruangsuriya | [68] | Zingerone was prepared in dimethyl sulphoxide (DMSO) and diluted to final concentrations in the culture media. | in vitro/chondrocytes | Zingerone suppressed the expression of TNF-α, IL-6, and IL-8 mRNA levels and reduced p38-mitogen-activated protein kinase (MAPK) and c-Jun n-terminal kinase phosphorylation, reducing cartilage inflammations and degradation. |
| Hosseinzadeh | [69] | Chondrocytes were pretreated with ginger extract and co-treated with IL-1β. | in vitro/chondrocytes | Ginger extract reduced IL-1β-induced oxidative stress, mitochondrial changes and apoptosis in chondrocytes. |
| First Author | References | Experimental Model | Study Type | Findings |
|---|---|---|---|---|
| Zhang | [72] | Natural extracellular matrix (ECM)/PLLA scaffolds loading Ica (icariin), Ica–2-hydroxypropyl-β-cyclodextrin were prepared via phase separation, solvent replacement and freeze drying. | in vitro/chondrocytes | Ica–2-hydroxypropyl-β-cyclodextrin inclusion complex-loaded PLLA scaffolds are suitable for cartilage TE. |
| Wang | [73] | Ica was added to the chondrogenic medium in bone marrow MSCs cultures. | in vitro/chondrocytes, MSCs | Ica is an accelerant of growth factors for cartilage TE by promoting chondrogenesis of bone marrow MSCs but not hypertrophy. |
| Zhang | [74] | Rabbit chondrocytes were isolated and cultured with different concentrations of Ica. | in vitro/rabbit chondrocytes | Ica is an effective accelerant for chondrogenesis by up-regulation of the expression of aggrecan (AGC), COL2A1, and SOX9 genes. Ica-loaded biomaterials have the potential for cartilage TE. |
| Liu | [75] | Murine chondrocytes were treated with lipopolysaccharides (LPS) and co-cultured with various concentrations of Ica. | in vitro/mouse chondrocytes | Ica is a safe anabolic agent of chondrocytes, inhibits NO and MMP synthesis and may exert its protective effects by inhibition of NO and MMP synthesis, reduces the ECM destruction. |
| Kankala | [79] | Fabricated porous sodium alginate and gelatin 3D scaffold by 3D printing method. Cells were incubated with Ica. | in vitro/chondrocytes | Ica significantly enhanced the proliferation of chondrocytes, suggesting application for cartilage TE. |
| Yang, Li | [80,81] | Ica conjugated hyaluronic acid/collagen hydrogel were used for osteochondral defect repair. | in vitro/chondrocytes | Ica–hyaluronic acid (HA)/Col constructs facilitated reconstruction of osteochondral interface in rabbit subchondral defects. Ica–HA/Col hydrogel is a promising scaffold for restoring of osteochondral defect, suggesting application for cartilage TE. |
| Wang | [82] | Cells were seeded in plates, maintained in normal medium with Ica. | in vitro/in vivo chondrocytes | Ica treatment upregulated mRNA levels of SOX9, COL2A1, and AGC in the 3D cultures. Ica significantly enhanced cartilage repair. |
| Luo | [83] | Ica was administered into the chondrogenic medium for micromass culture of mouse chondrocytes. | in vitro/in vivo mouse chondrocytes | In mouse model of OA and chondrocytes in micromass cultures, Ica treatment reduced destruction of cartilage, promoted chondrocyte differentiation, upregulated expression of parathyroid hormone related proteins (PHrP), down-regulated the expression of Indian hedgehog (Ihh). |
| Liu, Zhang | [84,85] | Injectable thiolated Ica functionalized Col/HA hydrogel to promote cartilage formation. Ica-conditioned serum engineered with hyaluronic acid, articular cartilage defects in rabbit knees. | in vitro/in vivo cartilage | Ica–Col/HA hydrogel had great potential for clinical application in articular cartilage repair. Ica-conditioned serum combined with HA promotes reparative response in cartilage defects, the possible application in bioactive material-based cartilage regeneration therapies. |
| First Author | References | Experimental Model | Study Type | Findings |
|---|---|---|---|---|
| Henrotin | [88] | Chondrocytes cultured for 72 h with/without IL-1β and co-treated with avocado/soybean unsaponifiables (ASUs) were analyzed by HPLC and mass spectrometry. Anti-inflammatory and anabolic activity was investigated. | in vitro/human chondrocytes | ASU increased the expression of COL2A1 and AGC genes, cell proliferation. ASU partially reversed IL-1β effects on chondrocytes. Reduction of IL-1β effects were consistent with chondroprotective activity. |
| Boumediene | [93] | Articular chondrocytes were treated with various concentrations of ASU, the expression of transforming growth factor-β (TGF-β1),2, and their receptors (TGF-βRI and RII) was determined by Northern blot and PCR. | in vitro/bovine chondrocytes | ASU induced stimulation of ECM synthesis by the ability to enhance TGF-β expression. ASU increased the production of plasminogen activator inhibitor (PAI-1), lead to inhibition of MMP, and induced matrix repair mechanisms in chondrocytes. |
| Frondoza | [95] | Evaluation of the effects of ASU/α-lipoic acid (LA) on production of PGE2 in equine chondrocyte stimulated with LPS, IL-1β, or H2O2 for 24 h, and supernatants were immunoassayed for PGE2. | in vitro/equine chondrocytes | Chondrocyte PGE2 production was inhibited by ASU/LA more effectively than either alone, which was associated with the suppression of NF-κB translocation. ASU/LA on PGE2 production has a potential for anti-inflammatory/antioxidant approach in OA. |
| Au | [96] | Evaluation of ASU on pro-inflammatory mediators in chondrocytes and monocyte/ macrophage-like cells. Cells were stimulated for 1 h with LPS, and analyzed for TNF-α, IL-1β, COX-2, inducible nitric oxide synthase (iNOS), and PGE2 expression. | in vitro/chondrocytes | ASU reduced TNF-α, IL-1β, COX-2, PGE2, and iNOS expression in LPS-stimulated chondrocytes, and anti-inflammatory effects of ASU were observed. ASU also reduced TNF-α and IL-1β expression in LPS-stimulated monocyte/ macrophage-like cells. |
| Lippiello | [97] | ASU samples were analyzed by HPLC, gas chromatography (GC) mass spectrometry, to clarify, if the sterol of ASU were the primary contributors to biological activity in chondrocytes. The sterol content was normalized between diverse samples prior to testing on chondrocytes. Anti-inflammatory activity was assayed by measuring of IL-1β-induced synthesis of PGE2, MMPs, release of S-35. | in vitro/chondrocytes | ASU samples exerted a time-dependent upregulation of the 35-sulphate uptake in chondrocytes. ASU were effective in the dose-dependent inhibition of IL-1β-induced MMP-3 activity, PGE2 synthesis. The upregulation of glycosaminoglycans (GAGs), collagen synthesis, and the reduction of IL-1β effects in cartilage were compatible with chondroprotective activity. |
| Hashemibeni | [91] | Isolated human adipose tissue-derived mesenchymal stem cells (hADSCs) were seeded in fibrin or fibrin-alginate scaffolds in chondrogenic medium containing Piascledin, TGF-β1, or both. | in vitro/hADSCs/chondrocytes | Piascledin was able to enhance the proliferation, survival of hADSCs in scaffolds. However, the expression of COL2A1 was higher in the TGF-β1 groups, and the expression of AGC was higher in TGF-β1 alone or with Piascledin in scaffolds. |
| First Author | References | Experimental Model | Study Type | Findings |
|---|---|---|---|---|
| Ahmed | [109] | OA chondrocytes or cartilage explants were pre-treated with pomegranate fruit extract (PFE), co-treated with IL-1β. The amounts of PG were measured with a colorimetric assay. The expression of MMPs, pIkBα, and MAPKs was determined by WB and NF-kB by electrophoretic mobility shift assay (EMSA). | in vitro/cartilage explants, chondrocytes | PFE inhibited the IL-1β-induced PG breakdown, MMPs expression on protein and mRNA level, p38-MAPK, phosphorylation of inhibitor of kappa B alpha (IkBα), and NF-kB binding to DNA in OA cartilage explants. |
| Haseeb | [111] | The potential of PFE to suppress IL-1β-stimulated expression of IL-6, reactive oxygen species (ROS), and NF-κB by analyzing the activation of the kinases upstream of IκBα in PCH by WB. | in vitro/human chondrocytes | PFE inhibited the mRNA and protein expression of IL-6, ROS, and IL-1β-mediated phosphorylation IKKβ, degradation of IκBα, and activation and nuclear translocation of NF-κB/p65 in human chondrocytes. PFE exerted chondroprotective effects by suppressing the NF-kB pathway. |
| Akhtar | [113] | OA was surgically induced in the tibiofemoral joints of rabbits. In one group, animals were fed PFE in water for 8 wks postsurgery. In the second group, animals were fed PFE for 2 wks before surgery and for 8 wks postsurgery. | in vitro/in vivo rabbit chondrocytes | PFE-fed rabbits had lower levels of IL-6, MMP-13, and PGE2 in synovial fluid/plasma and showed higher expression of AGC and COL2A1 mRNA. PFE treatment significantly reduced IL-1β-induced MAPK and NF-κB inhibitors, and PGE2 production, which highlighted the chondroprotective effect of PFE in the treatment of OA. |
| Monsefi | [115] | Pregnant BALB/c mice were given PFE to investigate the effect on chondrogenesis. Their embryos were stained with alizarin red S and alcian blue. Bone Ca content in pregnant mice was also measured. | in vitro/in vivo mouse chondrocytes, MSCs | PFE was able to enhance bone/ cartilage formation. MSCs from fetal limb buds were cultured, exposed to PFE, the number of viable cells was greater than in control cultures. The number of cartilage nodules and their diameters were greater in PFE-treated cultures. |
| Ghoochani | [116] | Patients with knee OA and control drank 200 mL PFE/daily for 6 weeks, and the effect of this intervention on clinical signs was evaluated. | in vivo/OA patients | Significant increases in physical function of decrease in breakdown of cartilage enzymes and increase of anti-oxidant status in patients with knee OA were observed in PFE group. |
| First Author | References | Experimental Model | Study Type | Findings |
|---|---|---|---|---|
| Csaki, Shakibaei | [34,146] | Investigation of the potential synergistic effects of resveratrol or/and curcumin on IL-1β-stimulated human PCH using WB and electron microscopy (EM). | in vitro/human chondrocytes | Both compounds targeted the NF-κB and MAPK pathways. Resveratrol inhibits the proteasome, while curcumin modulates inhibiting upstream kinases and MAPK. |
| Buhrmann | [58] | PCH were cultured in 3D-alginate cultures, resveratrol was prepared in ethanol, diluted in medium. Alginate cultures of PCH were treated with TNF-β, TNF-α, or T-lymphocytes and co-treated with resveratrol. | in vitro/human chondrocytes | Suppression of TNF-β-, similar to TNF-α- or T-lymphocytes-induced inflammatory microenvironment in PCH by resveratrol/histone deacetylase sirtuin-1 (SIRT1), might be a novel therapeutic approach for targeting inflammation during OA/RA. |
| Wang | [78] | Resveratrol grafted to polyacrylic acid to fabricate collagen/resveratrol scaffolds and chondrocytes seeded on the scaffolds. | in vitro/in vivo chondrocytes | Osteochondral defects were completely repaired by the collagen/resveratrol scaffold, and the neo-cartilage integrated well with its surrounding tissue. |
| Csaki, Shakibaei | [121,122] | Human PCH in monolayer cultures treated with IL-1β and co-treated with various concentrations of resveratrol and evaluated with ((3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium-bromid)) (MTT) assay, WB, and EM. | in vitro/human chondrocytes | Resveratrol inhibited the expression of VEGF, MMP-3, MMP-9, and COX-2 in PCH stimulated with IL-1β. Resveratrol exerted a chondroprotective capacity by suppression of IL-1β, ROS, p53-production. and apoptosis by down-regulation of NF-kB. |
| Dave | [123] | Human chondrocytes and cartilage explants were isolated from OA patients, treated with IL-1β and/or resveratrol. Evaluation of PGE2, leukotriene B (LTB4), COX, and MMP expression, as well as PG production. | in vitro/in vivo chondrocytes | Resveratrol inhibited chondrocyte apoptosis via inhibition of COX-2-derived PGE2 synthesis by suppression of mitochondrial membrane depolarization, depletion. Resveratrol protected against oxidant injury and apoptosis, which are main features of progressive OA. |
| Kim | [124] | Healthy and OA chondrocytes were incubated with various concentrations of resveratrol. Cell proliferation and gene expressions were evaluated. | in vitro/chondrocytes | Resveratrol treatment significantly upregulated SIRT1 gene expression in normal and OA chondrocytes. |
| Liu | [125] | Chondrocytes were isolated from pig joints; Resveratrol was prepared as a stock solution with ethanol. Activation of the IκBα, NF-kB, and MAPK, activator protein-1 (AP-1) pathways was assessed by EMSA, WB, and transfection assay. | in vitro/in vivo porcine chondrocytes | iNOS, COX-2, PGE2 were suppressed by resveratrol, mediated by inhibiting IKK-IκBα-NF-κB and JNK/ERK/AP-1pathways induced by advanced glycation end products (AGEs). Resveratrol could prevent AGEs-induced degradation of PG and AGC in cartilage explants. |
| Im | [134] | Isolated human PCH, cultured in monolayer or in 3D-alginate cultures and treated with resveratrol. | in vitro/human chondrocytes | Resveratrol improved the viability of PCH, antagonized protease production, and promoted PG synthesis as evaluated by (35) S-sulfate incorporation. Modulation of the downstream regulators of MAPK and NF-κB. Resveratrol exerted its chondroprotective actions partly by inhibition p53-induced apoptosis but not in chondrosarcoma. |
| Eo | [135] | The effect of resveratrol on NO-induced apoptosis in rabbit articular chondrocytes was investigated. | in vitro/chondrocytes | Resveratrol inhibited NO-induced apoptosis through the NF-κB signaling pathway in articular chondrocytes. |
| Maepa | [136] | Porcine articular chondrocytes were isolated cultured as monolayers, treated with resveratrol. | in vitro/porcine chondrocytes | Resveratrol stimulated the expression of collagen II, regulated collagen II protein in different zones of articular cartilage. |
| Elmali | [137] | OA model, a unilateral anterior cruciate ligament transection (ACLT) was performed in rabbits. Five weeks after the test group had been injected daily with 10 μM/kg resveratrol in DMSO into the knees for two weeks, the control group was injected DMSO into the knees. | in vivo/rabbit chondrocytes | Intraarticular injections of resveratrol starting at the onset of OA disease may protect cartilage against the development of experimentally induced OA. |
| Li | [138] | Intra-articular injection of resveratrol into a mouse model of OA. OA was induced in the mouse knee using the destabilization of the medial meniscus (DMM). Resveratrol was injected weekly into the operated knee starting 4 weeks after surgery. The OA phenotype was investigated by histological and IHC analyses. | in vitro/in vivo human chondrocytes | Injection of resveratrol significantly prevented the destruction of OA cartilage by stimulating SIRT1, thereby suppressing the expression of HIF-2α and catabolic factors. Activation of SIRT1, the inhibition of HIF-2α in cartilage tissue and in vitro in IL-1β-treated chondrocytes. |
| Jin | [141] | Mouse chondrogenic cells were treated with 30 μM resveratrol for 24 h and 10 μg/mL LPS for 12 h. Cell viability, apoptosis and the release of pro-inflammatory cytokines was assessed. | in vitro/mouse chondrocytes | Resveratrol supported chondrogenic cell line of LPS-induced inflammatory apoptotic effects by upregulation of miR-146b, and deactivation of NF-κB, p38-MAPK signaling. |
| Lei | [143] | MSC-derived chondrocytes cultured on CGS, treated with IL-1β and co-treated with resveratrol and evaluated ECM, MMPs, and NF-kB. | in vitro/MSCs chondrocytes | Resveratrol acted as a NF-kB inhibitor to protect MSC-derived chondrocytes on the CGS from IL-1β catabolism and these effects were mediated by β1-integrin. |
| Sheu | [144] | Fabricated and characterized an Oxi-HA/resveratrol hydrogel for future applications in cartilage TE. | in vitro/chondrocytes | Oxi-HA/resveratrol hydrogel up-regulated COL2A1, AGC, and SOX-9 genes, down-regulated IL-1β and MMPs genes. Oxi-HA/resveratrol hydrogel is biocompatible with chondrocytes, allows ECM synthesis, a potentially suitable cell carrier for chondrocyte in the treatment of cartilage defects |
| Wu | [145] | Resveratrol-loaded microspheres were fabricated using oil-in-water emulsion and solution-evaporation methods. Human bone marrow MSCs were treated with IL-1β and co-treated with resveratrol. | in vitro/hMSCs | Resveratrol inhibited the activity of IL-1β, thereby downregulating MMP-13 mRNA expression. Up-regulation of COL2A1, AGC, and Sox9 mRNA expression. Resveratrol maintained chondrogenic gene expression of cells when exposed to the inflammatory agents. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buhrmann, C.; Honarvar, A.; Setayeshmehr, M.; Karbasi, S.; Shakibaei, M.; Valiani, A. Herbal Remedies as Potential in Cartilage Tissue Engineering: An Overview of New Therapeutic Approaches and Strategies. Molecules 2020, 25, 3075. https://doi.org/10.3390/molecules25133075
Buhrmann C, Honarvar A, Setayeshmehr M, Karbasi S, Shakibaei M, Valiani A. Herbal Remedies as Potential in Cartilage Tissue Engineering: An Overview of New Therapeutic Approaches and Strategies. Molecules. 2020; 25(13):3075. https://doi.org/10.3390/molecules25133075
Chicago/Turabian StyleBuhrmann, Constanze, Ali Honarvar, Mohsen Setayeshmehr, Saeed Karbasi, Mehdi Shakibaei, and Ali Valiani. 2020. "Herbal Remedies as Potential in Cartilage Tissue Engineering: An Overview of New Therapeutic Approaches and Strategies" Molecules 25, no. 13: 3075. https://doi.org/10.3390/molecules25133075
APA StyleBuhrmann, C., Honarvar, A., Setayeshmehr, M., Karbasi, S., Shakibaei, M., & Valiani, A. (2020). Herbal Remedies as Potential in Cartilage Tissue Engineering: An Overview of New Therapeutic Approaches and Strategies. Molecules, 25(13), 3075. https://doi.org/10.3390/molecules25133075