
High Diversity and Low Coinfections of Pathogens in Ticks from Ruminants in Pakistan

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
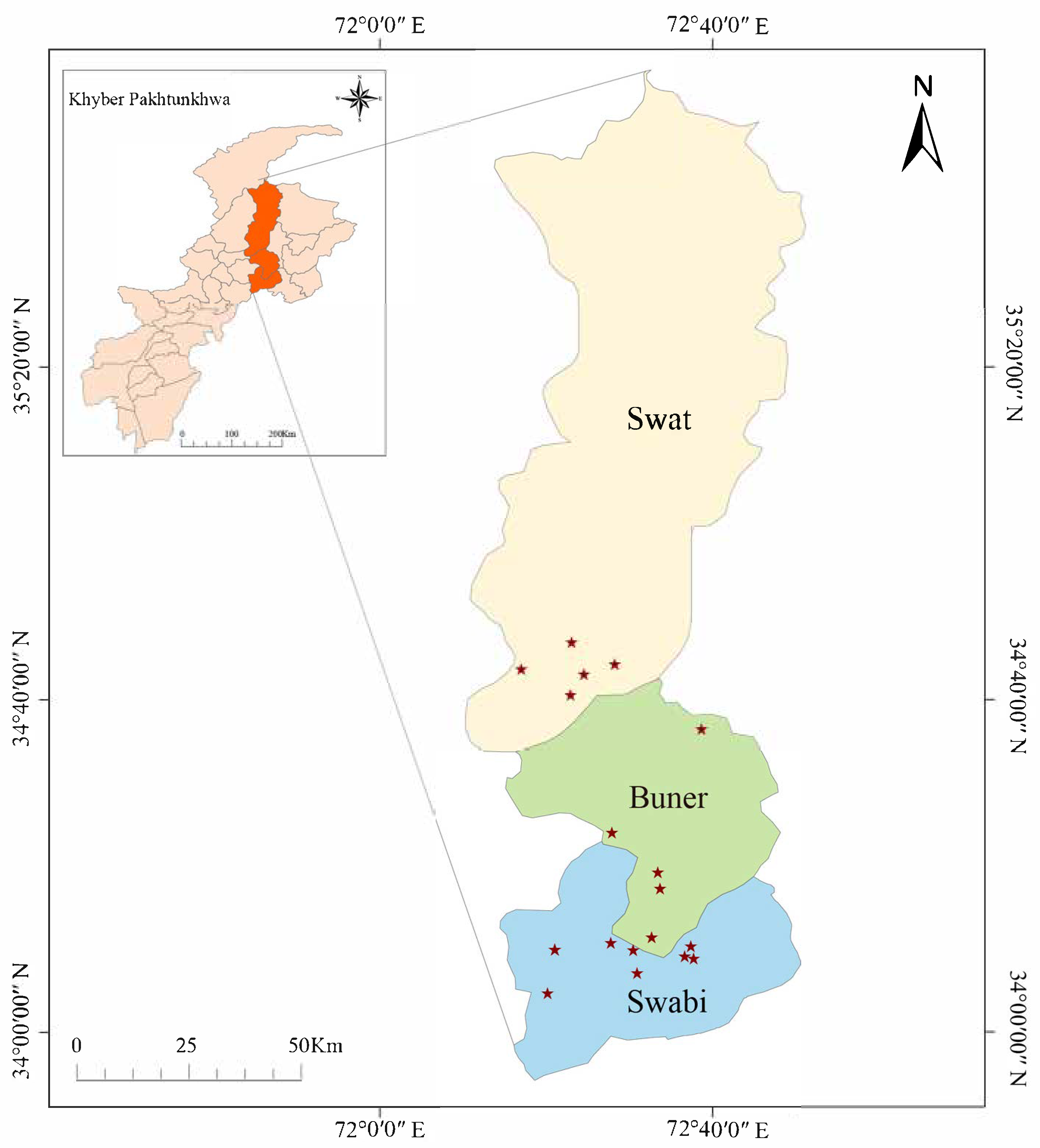
2.1. Study Areas and Sample Collection
2.2. DNA Extraction and PCR Assays
2.3. Phylogenetic Analyses
2.4. Statistical Analyses
2.5. Ethical Approval
3. Results
3.1. Collected Tick Samples
3.2. Detection and Phylogenetic Analysis of Anaplasma
3.3. Detection and Phylogenetic Analysis of Ehrlichia
3.4. Detection and Phylogenetic Analysis of Rickettsia
3.5. Detection and Phylogenetic Analysis of Babesia and Theilria
3.6. Prevalence and Co-Infections of Tick-Borne Agents
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nasirian, H. Hard tick species parasitism levels in domestic ruminants with their distribution and role as vectors: A detailed global meta-analysis and systematic review. Acta Parasitol. 2023, 69, 1–105. [Google Scholar] [CrossRef] [PubMed]
- Paules, C.I.; Marston, H.D.; Bloom, M.E.; Fauci, A.S. Tickborne diseases—Confronting a growing threat. N. Engl. J. Med. 2018, 379, 701–703. [Google Scholar] [PubMed]
- Rochlin, I.; Toledo, A. Emerging tick-borne pathogens of public health importance: A mini-review. J. Med. Microbiol. 2020, 69, 781–791. [Google Scholar] [CrossRef]
- Rahman, A.; Kashif, M.; Nasir, A.; Idress, A.; Jamil, M.; Qadir, Z.A.; Qasim, M.; Khan, I.; Aziz, H.; Qazi, I.; et al. A review of tick and tick control strategies in Pakistan. Pak. J. Med. Health Sci. 2022, 16, 652–655. [Google Scholar] [CrossRef]
- Mushtaq, A.; Shoukat, T.; Mumtaz, T.; Qasim, M.; Ajmal, K.; Fatima, N.; Khan, A.; Kouser, M.; Hussain, N.; Khan, S.S.; et al. Tick-borne diseases in sheep and goats in Pakistan: A systematic review and meta-analysis. Acta Parasitol. 2021, 66, 1316–1325. [Google Scholar] [CrossRef]
- Ghafar, A.; Abbas, T.; Rehman, A.; Sandhu, Z.-U.-D.; Cabezas-Cruz, A.; Jabbar, A. Systematic review of ticks and tick-borne pathogens of small ruminants in Pakistan. Pathogens 2020, 9, 937. [Google Scholar] [CrossRef]
- Ashraf, Q.U.; Khan, A.U.; Khattak, R.M.; Ali, M.; Shaikh, R.S.; Ali, M.; Iqbal, F. A report on the high prevalence of Anaplasma sp. in buffaloes from two provinces in Pakistan. Ticks Tick. Borne Dis. 2013, 4, 395–398. [Google Scholar] [CrossRef]
- Karim, S.; Budachetri, K.; Mukherjee, N.; Williams, J.; Kausar, A.; Hassan, M.J.; Adamson, S.; Dowd, S.E.; Apanskevich, D.; Arijo, A.; et al. A study of ticks and tick-borne livestock pathogens in Pakistan. PLoS Negl. Trop. Dis. 2017, 11, e0005681. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.; Roohi, N.; Rashid, M.I.; Ali, S.; Ul-Rehman, Z. Occurrence of ticks and tick-borne mixed parasitic microbiota in cross-bred cattle in District Lahore, Pakistan. Braz. J. Biol. 2022, 82, e266721. [Google Scholar]
- Zulfiqar, S.; Shahnawaz, S.; Ali, M. Detection of Babesia bovis in blood samples and its effect on the hematological and serum biochemical profile in large ruminants from Southern Punjab. Asian Pac. J. Trop. Biomed. 2012, 2, 104–108. [Google Scholar] [CrossRef]
- Ammar, M.; Moaaz, M.; Yue, C.; Fang, Y.; Zhang, Y.; Shen, S.; Deng, F. Emerging arboviral diseases in Pakistan: Epidemiology and public health implications. Viruses 2025, 17, 232. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Abbas, R.Z.; Alsayeqh, A. Distribution pattern of Crimean-Congo Hemorrhagic Fever in Asia and the Middle East. Front. Public. Health 2023, 11, 1093817. [Google Scholar] [CrossRef]
- Yasmeen, N.; Jabbar, A.; Shah, T. One Health Paradigm to Confront Zoonotic Health Threats: A Pakistan Prospective. Front. Microbiol. 2022, 12, 719334. [Google Scholar] [CrossRef]
- Ullah, S.; Alouffi, A.; Almutairi, M.M. First Report of Rickettsia conorii in Hyalomma kumari Ticks. Animal 2023, 13, 1488. [Google Scholar] [CrossRef]
- Seidi, S.; Omidi, A.H.; Esmaeili, S. Distribution of different Rickettsia species in countries of the WHO Eastern Mediterranean (WHO-EMRO) region: An overview. Travel. Med. Infect. Dis. 2024, 58, 102695. [Google Scholar]
- Gebrekidan, H.; Abbas, T.; Wajid, M.; Ali, A.; Gasser, R.B.; Jabbar, A. Molecular characterisation of Theileria orientalis in imported and native bovines from Pakistan. Infect. Genet. Evol. 2017, 47, 19–25. [Google Scholar] [CrossRef]
- Nasreen; Khan, A.; Niaz, S. Molecular detection of small ruminant piroplasmosis and first report of Theileria luwenshuni (Apicomplexa: Theileridae) in small ruminants of Pakistan. Exp. Parasitol. 2020, 212, 107872. [Google Scholar] [CrossRef] [PubMed]
- El Imam, A.H.; Hassan, S.M.; Gameel, A.A.; El Hussein, A.M.; Taha, K.M.; Oosthuizen, M.C. Molecular identification of different Theileria and Babesia species infecting sheep in Sudan. Ann. Parasitol. 2016, 62, 47–54. [Google Scholar] [PubMed]
- Zeb, J.; Shams, S.; Din, I.U. Molecular epidemiology and associated risk factors of Anaplasma marginale and Theileria annulata in cattle from North-western Pakistan. Vet. Parasitol. 2020, 279, 109044. [Google Scholar] [CrossRef]
- Khan, M.; Islam, N.; Khan, A. New records of Amblyomma gervaisi from Pakistan, with detection of a reptile-associated Borrelia sp. Ticks Tick. Borne Dis. 2022, 13, 102047. [Google Scholar] [CrossRef]
- Zeb, J.; Song, B.; Khan, M.A.; Senbill, H.; Aziz, M.U.; Hussain, S.; Sánchez, A.A.D.; Cabezas-Cruz, A.; Alzahrani, A.; Alshehri, M.; et al. Genetic diversity of tick-borne zoonotic pathogens in ixodid ticks collected from small ruminants in Northern Pakistan. Infect. Genet. Evol. 2024, 124, 105663. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, A.; Darzi, M.M.; Wani, B.M.; Shah, S.A.; Shabir, M.; Shafi, M. Climate change and infectious diseases of animals: A review. J. Entomol. Zool. Stud. 2017, 5, 1470–1477. [Google Scholar]
- Government of Pakistan (GOP). Pakistan Economic Survey 2019–2020. Ministry of Finance. Available online: https://www.finance.gov.pk/updates.html (accessed on 11 June 2020).
- Apanaskevich, D.A.; Horak, I.G. The genus Hyalomma Koch, 1844: V. re-evaluation of the taxonomic rank of taxa comprising the H. (Euhyalomma) marginatum koch complex of species (Acari: Ixodidae) with redescription of all parasitic stages and notes on biology. Int. J. Acarol. 2008, 34, 13–42. [Google Scholar] [CrossRef]
- Brahma, R.K.; Dixit, V.; Sangwan, A.K.; Doley, R. Identification and characterization of Rhipicephalus (Boophilus) microplus and Haemaphysalis bispinosa ticks (Acari: Ixodidae) of northeast India by ITS2 and 16S rDNA sequences and morphological analysis. Exp. Appl. Acarol. 2014, 62, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Nava, S.; Beati, L.; Venzal, J.M.; Labruna, M.B.; Szabó, M.P.J.; Petney, T.; Saracho-Bottero, M.N.; Tarragona, E.L.; Dantas-Torres, F.; Silva, M.M.S.; et al. Rhipicephalus sanguineus (Latreille, 1806): Neotype designation, morphological re-description of all parasitic stages and molecular characterization. Ticks Tick. Borne Dis. 2018, 9, 1573–1585. [Google Scholar] [CrossRef]
- Intirach, J.; Lv, X.; Han, Q.; Lv, Z.Y.; Chen, T. Morphological and molecular identification of hard Ticks in Hainan Island, China. Genes 2023, 14, 1592. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J.; Kjer, K.; Page, R.; Sullivan, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v6: Recent updates to the phylogenetic tree display and annotation tool. Nucl. Acids Res. 2024, 52, W78–W82. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S.; Battistuzzi, F.U. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Khan, M.A.; Zahid, H.; Yaseen, P.M.; Khan, M.Q.; Nawab, J.; Rehman, Z.U.; Ateeq, M.; Khan, S.; Ibrahim, M.; et al. Seasonal dynamics, record of ticks infesting humans, wild and domestic animals and molecular phylogeny of Rhipicephalus microplus in Khyber Pakhtunkhwa Pakistan. Front. Physiol. 2019, 10, 793. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Mulenga, A.; Vaz, I.S. Editorial: Tick and tick-borne pathogens: Molecular and immune targets for control strategies. Front. Physiol. 2020, 11, 744. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Shehla, S.; Alouffi, A.; Obaid, M.K.; Khan, A.Z.; Almutairi, M.M.; Numan, M.; Aiman, O.; Alam, S.; Ullah, S.; et al. Molecular survey and genetic characterization of Anaplasma marginale in ticks collected from livestock hosts in Pakistan. Animals 2022, 12, 1708. [Google Scholar] [CrossRef]
- Moura de Aguiar, D.; Pessoa Araújo Junior, J.; Nakazato, L.; Bard, E.; Aguilar-Bultet, L.; Vorimore, F.; Leonidovich Popov, V.; Moleta Colodel, E.; Cabezas-Cruz, A. Isolation and characterization of a novel pathogenic strain of Ehrlichia minasensis. Microorganisms 2019, 7, 528. [Google Scholar] [CrossRef]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.-E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [PubMed]
- Vitale, G.; Mansuelo, S.; Rolain, J.M.; Raoult, D. Rickettsia massiliae human isolation. Emerg. Infect. Dis. 2006, 12, 174–175. [Google Scholar] [CrossRef]
- Ali, A.; Ullah, S.; Numan, M.; Almutairi, M.M.; Alouffi, A.; Tanaka, T. First report on tick-borne pathogens detected in ticks infesting stray dogs near butcher shops. Front. Vet. Sci. 2023, 10, 1246871. [Google Scholar] [CrossRef]
- Khan, M.; Khan, M.; Khan, M.; Batool, S.; Tanaka, T.; Aloufi, A.; Almutairi, M.M.; Ali, A. Babesia bigemina and Theileria annulata infections in cattle: Molecular detection, phylogenetic analysis, and assessment of risk factors. Trop. Anim. Health Prod. 2024, 56, 282. [Google Scholar] [CrossRef]
- Gharbi, M.; Sassi, L.; Dorchies, P.; Darghouth, M.A. Infection of calves with Theileria annulata in Tunisia: Economic analysis and evaluation of the potential benefit of vaccination. Vet. Parasitol. 2006, 137, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Asif, M.; Said, M.B.; Parveen, A.; Ejaz, A.; Ikram, M.; Awais, M.M.; Ozubek, S.; Aktas, M.; Baber, M.; Iqbal, F. Seasonal survey, risk factor’s analysis and genotyping of Theileria annulata infecting cattle in Punjab province, Pakistan. Acta Trop. 2022, 234, 106587. [Google Scholar] [CrossRef]
- Roy, B.C.; Krücken, J.; Ahmed, J.S.; Majumder, S.; Baumann, M.P.; Clausen, P.H.; Nijhof, A.M. Molecular identification of tick-borne pathogens infecting cattle in Mymensingh district of Bangladesh reveals emerging species of Anaplasma and Babesia. Transbound. Emerg. Dis. 2018, 65, e231–e242. [Google Scholar] [CrossRef] [PubMed]
- Al-Hamidhi, S.; HTageldin, M.; Weir, W.; Al-Fahdi, A.; Johnson, E.H.; Bobade, P.; Alqamashoui, B.; Beja-Pereira, A.; Thompson, J.; Kinnaird, J.; et al. Genetic diversity and population structure of Theileria annulata in Oman. PLoS ONE 2015, 10, e0139581. [Google Scholar] [CrossRef]
- Zhu, W.-J.; Ye, R.-Z.; Tian, D.; Wang, N.; Gao, W.-Y.; Wang, B.-H.; Lin, Z.-T.; Liu, Y.-T.; Wang, Y.-F.; Zhu, D.-Y.; et al. The first direct detection of spotted fever group Rickettsia spp. diversity in ticks from Ningxia, northwestern China. PLoS Negl. Trop. Dis. 2025, 19, e0012729. [Google Scholar] [CrossRef] [PubMed]
- Nooroong, P.; Trinachartvanit, W.; Baimai, V.; Ahantarig, A. Phylogenetic studies of bacteria (Rickettsia, Coxiella, and Anaplasma) in Amblyomma and Dermacentor ticks in Thailand and their co-infection. Ticks Tick. Borne Dis. 2018, 9, 963–971. [Google Scholar] [CrossRef]
- Hoffmann, A.; Müller, T.; Fingerle, V.; Silaghi, C.; Noll, M. Co-infection of potential tick-borne pathogens of the order Rickettsiales and Borrelia burgdorferi s. l. and their link to season and area in Germany. Microorganisms 2023, 11, 157. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| District | Host | Tick Species | Total | ||||
|---|---|---|---|---|---|---|---|
| Rhipicephalus | Haemaphysalis | Hyalomma | |||||
| Rhipicephalus microplus | Rhipicephalus sanguineus | Rhipicephalus haemaphysaloides | Haemaphysalis bispinosa | Hyalomma marginatum | |||
| Swabi | Cattle | 156 (F122, M34) * | - | - | 3 (M3) | 155 (F105, M50) | 314 |
| Goat/Sheep | - | 17 (F9, M8) | 9 (F9) | 45 (F40, M5) | - | 71 | |
| Buner | Cattle | 114 (F114) | - | - | - | - | 114 |
| Sheep | 101 (F95, M6) | - | 4 (M4) | 1 (M1) | 2 (F2) | 108 | |
| Swat | Cattle | 382 (F371, M11) | - | - | - | - | 382 |
| Total | Sheep/Goat/Cattle | 753 | 17 | 13 | 49 | 157 | 989 |
| Tick Species | No. of Tested Ticks | Positive Rate % (95% CI) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Anaplasma | Ehrlichia | Rickettsia | |||||||
| A. marginale | A. ovis | E. minasensis | C. E. hyalommae | C. E. rhipicephalis | R. felis | R. massiliae | C. R. pakistanensis | ||
| Hyalomma marginatum | 157 | 0 | 26.1 (19.2–32.9) | 0.64 (0.02–3.5) | 1.92 (0.4–5.5) | 0 | 0 | 0 | 4.5 (1.2–7.7) |
| Rhipicephalus microplus | 753 | 6.8 (5.0–8.6) | 0 | 0 | 0 | 0.93 (0.2–1.6) | 0.1 (0.0–0.7) | 0.1 (0.0–0.7) | 0 |
| Rhipicephalus sanguineus | 17 | 0 | 17.6 (4.1–41.9) | 0 | 0 | 0 | 0 | 29.4 (10.3–55.8) | 0 |
| Rhipicephalus haemaphysaloides | 13 | 0 | 7.7 (0.2–35.8) | 0 | 0 | 0 | 0 | 0 | 0 |
| Haemaphysalis bispinosa | 49 | 0 | 10.2 (3.4–22.2) | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 989 | 5.2 (3.7–6.5) | 5.1 (3.7–6.4) | 0.1 (0.0–0.6) | 0.3 (0.1–0.8) | 0.7 (0.2–1.2) | 0.1 (0.0–0.6) | 0.6 (0.1–1.1) | 0.7 (0.2–1.2) |
| Tick Species | No. of Tested Ticks | Positive Rate % (95% CI) | |||||
|---|---|---|---|---|---|---|---|
| Babesia | Theileria | ||||||
| B. bigemina | T. annulata | T. buffeli | T. luwenshuni | T. orientalis | T. sinensis | ||
| Hyalomma marginatum | 157 | 0 | 0 | 0 | 4.5 (1.2–7.7) | 0.64 (0.02–3.5) | 1.92 (0.4–5.5) |
| Rhipicephalus microplus | 753 | 0.3 (0.0–0.9) | 5.7 (4.0–7.4) | 0.1 (0.0–0.7) | 0.3 (0.0–0.9) | 3.0 (1.7–4.1) | 0 |
| Rhipicephalus sanguineus | 17 | 0 | 0 | 0 | 0 | 0 | 0 |
| Rhipicephalus haemaphysaloides | 13 | 0 | 0 | 0 | 0 | 0 | 0 |
| Haemaphysalis bispinosa | 49 | 0 | 0 | 0 | 10.2 (3.4–22.2) | 2.0 (0.0–11.0) | 18.8 (7.6–29.2) |
| Total | 989 | 0.2 (0.0–0.5) | 4.3 (3.0–5.6) | 0.1 (0.0–0.6) | 1.4 (0.7–2.2) | 2.4 (1.5–3.4) | 1.2 (0.5–1.9) |
| Co-Infection Pathogens | Tick Species | Host | District | Number | Positive Rate % (95% CI) |
|---|---|---|---|---|---|
| A. marginale + B. bigemina | R. microplus | Cattle | Swat | 2 | 0.2 (0.0–0.5) |
| A. marginale + T. annulata | R. microplus | Cattle | Swat | 11 | 1.1 (0.5–1.7) |
| A. ovis + C. R. pakistanensis | Hy. marginatum | Goat/Sheep | Swabi | 1 | 0.1 (0.0–0.6) |
| A. ovis + T. luwenshuni | Hy. marginatum | Goat/Sheep | Swabi | 1 | 0.1 (0.0–0.6) |
| A. ovis + T. sinensis | Hae. bispinosa | Goat/Sheep | Swabi | 3 | 0.3 (0.1–0.8) |
| Hy. marginatum | Goat/Sheep | Swabi | 3 | 0.3 (0.1–0.8) | |
| C. R. pakistanensis + T. luwenshuni | Hy. marginatum | Goat/Sheep | Swabi | 1 | 0.1 (0.0–0.6) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jamil, L.; Li, C.; Wang, Y.; Jamil, J.; Tian, W.; Zhao, D.; Shen, S.; Sun, Y.; Zhao, L.; Cao, W. High Diversity and Low Coinfections of Pathogens in Ticks from Ruminants in Pakistan. Microorganisms 2025, 13, 1276. https://doi.org/10.3390/microorganisms13061276
Jamil L, Li C, Wang Y, Jamil J, Tian W, Zhao D, Shen S, Sun Y, Zhao L, Cao W. High Diversity and Low Coinfections of Pathogens in Ticks from Ruminants in Pakistan. Microorganisms. 2025; 13(6):1276. https://doi.org/10.3390/microorganisms13061276
Chicago/Turabian StyleJamil, Laila, Cheng Li, Yifei Wang, Jabran Jamil, Wenya Tian, Di Zhao, Shijing Shen, Yi Sun, Lin Zhao, and Wuchun Cao. 2025. "High Diversity and Low Coinfections of Pathogens in Ticks from Ruminants in Pakistan" Microorganisms 13, no. 6: 1276. https://doi.org/10.3390/microorganisms13061276
APA StyleJamil, L., Li, C., Wang, Y., Jamil, J., Tian, W., Zhao, D., Shen, S., Sun, Y., Zhao, L., & Cao, W. (2025). High Diversity and Low Coinfections of Pathogens in Ticks from Ruminants in Pakistan. Microorganisms, 13(6), 1276. https://doi.org/10.3390/microorganisms13061276