
Research Progresses on the Function and Detection Methods of Insect Gut Microbes

Tangshan Key Laboratory of Agricultural Pathogenic Fungi and Toxins, Department of Life Science, Tangshan Normal University, Tangshan 063000, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Microorganisms 2023, 11(5), 1208; https://doi.org/10.3390/microorganisms11051208
Submission received: 3 March 2023
/
Revised: 20 March 2023
/
Accepted: 27 April 2023
/
Published: 5 May 2023
(This article belongs to the Section Veterinary Microbiology)
Abstract
:The insect gut is home to an extensive array of microbes that play a crucial role in the digestion and absorption of nutrients, as well as in the protection against pathogenic microorganisms. The variety of these gut microbes is impacted by factors such as age, diet, pesticides, antibiotics, sex, and caste. Increasing evidence indicates that disturbances in the gut microbiota can lead to compromised insect health, and that its diversity has a far-reaching impact on the host’s health. In recent years, the use of molecular biology techniques to conduct rapid, qualitative, and quantitative research on the host intestinal microbial diversity has become a major focus, thanks to the advancement of metagenomics and bioinformatics technologies. This paper reviews the main functions, influencing factors, and detection methods of insect gut microbes, in order to provide a reference and theoretical basis for better research utilization of gut microbes and management of harmful insects.
1. Introduction
The insect microbial community plays an important role in nutrient absorption, development of immunity, and resistance to foreign pathogenic organisms [1,2,3]. Gut microbes and hosts are interdependent and mutually restrictive and play an important role in host health [4]. Alterations in the intestinal community may influence insecticide resistance, physiological functions, and the eating habits of the host. Insect intestinal microorganisms have shown great application potential in the fields of pest prevention and control, utilization of insect resources, and industrial production (such as degrading cellulose and providing trace elements) [5,6,7]. Currently, functional studies of insect intestinal microorganisms remain challenging, in part because of the complexity of their composition, which may vary widely among individuals [8,9]. Therefore, understanding the function and composition of insect intestinal microorganisms is very important for regulating the rise and fall of the population, studying the relevant evolutionary mechanisms, and excavating strains with special functions [7].
Historically, traditional culture-based methods have been used to identify insect gut microbial diversity with many drawbacks (such as the fact that many intestinal bacteria cannot be obtained through culture and cannot systematically reveal the characteristics of insect intestinal flora), hindering the in-depth study of gut microbiota. In recent years, the development and application of new technologies such as metagenomics and bioinformatics have considerably enhanced our understanding of the composition and variety of the insect gut microbiota. This review summarizes insect gut function and the factors that influence insect gut microbes. In addition, we compared past and present methods for detecting insect intestinal microorganisms.
2. Functional Roles of the Insect Gut Microbiota
There are complicated microbial communities in the gut of insects, including bacteria, archaea, fungi, and viruses, which play an important role in the host’s nutrient metabolism, immune defense, and drug resistance enhancement. Insect gut microbiota supply host nutrition and maintain gut microbial homeostasis by generating antimicrobial compounds (bacteriocins and lipopeptides), metabolites such as short-chain fatty acids and vitamins, and by degrading plastics or inducing immune responses [3,10,11,12]. Xiao et al. found that Drosophila gut microbial homeostasis is synergistically regulated by Duox-ROS, AMPs, and C-type lectins [12]. In addition, the intestinal flora of insects can also recycle urea and uric acid, help insects defend against the infection of foreign pathogenic microorganisms, maintain body homeostasis, and facilitate the regular physiological activities of the host and the progress of various life activities [13].
2.1. Nutrient Metabolism
The intestinal microbiota of insects can secrete a variety of digestive enzymes to participate in the digestion of food and supply nutrients for insects. Common insects such as Bombyx mori, Locusts, Aedes albopictus, Termite, etc., have intestinal microorganisms that can secrete a variety of digestive enzymes. Pseudomonas, Klebsiella pneumoniae, and Clostridium flexneri in the intestinal microbiota of B. mori can secrete cellulase and have the ability to degrade carbohydrates [14]. Cellulase secreted by Klebsiella pneumonia in the intestinal tract of Locusta migratoria manilensis can degrade grass to generate carbohydrates, amino acids, and sugars for the host to utilize [15]. After Aedes albopictus ingests plant nectar, part of it is directly digested and absorbed, and the other part is transferred to the gut, where it is transformed into a common energy source for the host and gut microbes under the action of gut bacteria or fungi [16]. Cellobiohydrolase, which is secreted by microorganisms in the gut of Termites, can digest cellulose to provide energy for the host [17]. In addition, Klebsiella, Proteus vulgaris, Erwinia sp., and Serratia liquefaciens were isolated from the gut of Diatraea saccharalis larvae by in vitro culture, which can utilize starch, xylanolytic, pectinolytic, and polysaccharide, respectively [18]. Zheng et al. reveal that honeybee intestinal bacteria can degrade plant polymers from pollen and that the resulting metabolites provide nutrients for the host [19]. In addition, some intestinal symbionts can provide the host with amino acids and vitamins [20]. Hu et al. reported that the gut bacteria of ants utilize reclaimed recycled N to recycle urea or uric acid to synthesize amino acids required by the host in large quantities [13]. Duplais et al. reported that intestinal microorganisms in Herbivorous turtle ants Cephalotes can recycle metabolic waste rich in nitrogen to enrich nutritional components and supply amino acids for the host [21]. Blow et al. confirmed that Acyrthosiphon pisum can obtain vitamins B2 and B5 through intestinal symbiotic bacteria, Buchnera aphidicola, and supplement micronutrients [6]. These results indicate that gut microbes are a promising source of various digestive enzymes and trace elements.
In addition, the intestinal flora participates in the nutrient metabolism of insects and can also influence the feeding behavior, growth, and development of the host. Because of its simple intestinal microorganism species and easy feeding, Drosophila is often used as a model to study the function of symbiotic bacteria [22]. Wong et al. inoculated Drosophila eggs on a feed comprising Acetobacter pomorum and Lactobacillus plantarum (bacteria isolated from the gut or body of Drosophila) to investigate the behavioral responses of Drosophila to microorganisms. The research found that Drosophila vaccinated with A. pomorum had a diminished tendency to choose a high-protein diet, while flies vaccinated with L. plantarum were more likely to eat foods with higher carbohydrates [23]. In addition, both adults and larvae of D. melanogaster were attracted to volatile compounds from Saccharomyces cerevisiae and L. plantarum but repelled by Acetobacter malorum in behavioral assays [24]. Another study found that the germ-free Rhynchophorus ferrugineus Olivier larval development and weight had considerably diminished. In addition, after the introduction of intestinal microbiota, the content of hemolymph protein, glucose, and triglycerides significantly increased, and it was verified that the protein content level was restored after feeding sterile R. ferrugineus Olivier and Lactococcus lactis. Enterobacter cloacae significantly increases its hemolymph triglyceride and glucose content [25]. The changes in host behavior induced by these gut bacteria may reflect the metabolism of the gut microbiota and the nutritional needs of the host [26]. The above research supplies fundamental data for the breeding of insects and the functional research of intestinal microorganisms (Table 1).
2.2. Immune Defense
Insects are constantly in contact with pathogenic microorganisms such as viruses, fungi, and bacteria. Insects primarily rely on insect intestinal epithelial cells to generate reactive oxygen species (ROS) and antimicrobial peptides (AMPs) to resist the invasion of pathogenic microorganisms and preserve intestinal microbial homeostasis [27,28]. Recent research has shown that the insect gut microbiota plays a role in helping the host resist infection by foreign pathogens. Shao et al. found that Enterococcus mundtii can stably secrete a class IIa bacteriocin (mundticin KS) in the intestine of Spodoptera littoralis against invading bacteria to maintain the balance of the host gut microbial community [29]. Akbar et al. inoculated RPMI 1640 with intestinal bacteria isolated from cockroaches. Cockroach intestinal flora secretions can effectively inhibit Gram-positive (methicillin-resistant Staphylococcus aureus, Streptococcus pyogenes, Bacillus cereus) and Gram-negative (Escherichia coli K1, Salmonella enterica, etc.) [30]. Knight et al. isolated antifungal cyclic lipopeptides (bacteromycin F and fengycin) from the metabolites of Bacillus subtilis, which have inhibitory effects on a variety of fungi such as Alternaria alternata, Aspergillus niger, and Cladosporium sp. [31]. The above studies reveal that the metabolites of insect gut microbiota play an important role in maintaining gut microbial homeostasis and resisting the invasion of pathogenic microorganisms.
Recent studies have shown that gut commensals are important participants in the host immune system, and there are diverse interactions between gut commensals and the host immune system [32]. Pandey et al. analyzed the possible links between the gut microbial dynamics of the model organism Spodoptera litura and stress-inducing factors and found that Pseudomonas and Enterobacter are related to the inflammatory effects of insects. Acinetobacter, on the other hand, promotes larval fitness and decreases the inflammatory effects generated by dextran sulfate sodium [33]. Krams et al. demonstrated that Enterococcus was the main genus in the midgut of G. mellonella larvae, and the number of Enterococcus positively correlated with antimicrobial peptide-related genes (such as Gallerimycin, Gloverin, 6-tox, Cecropin-D, and Galiomicin) [1]. In addition, the R. ferrugineus Olivier larvae had higher antibacterial activity and phenoloxidase activity, while the immune-related genes and survival rate of sterile larvae were significantly down-regulated. The reintroduction of sterile R. ferrugineus Olivier larvae into the gut microbiota enhanced their immunity and survival [34]. Gao et al. revealed that the gut PGRP-LA gene of Anopheles stephensi could regulate immune responses by sensing the dynamics of the gut microbiota [35]. Park et al. demonstrated that the gelatinase secreted by Enterococcus faecalis in the intestinal tract of Galleria mellonella larvae can degrade antimicrobial peptides (Gm cecropin) in the hemolymph and damage the immune system [36]. The above studies have confirmed that intestinal commensal flora has a stimulating effect on host immunity. The research on the interaction mechanism between gut microbiota and insect immunity will help provide new strategies for pest management.
2.3. Antioxidation Function
Under normal circumstances, the normal metabolism of oxygen in the insect host will generate reactive oxygen and free radicals, but when they accumulate too much, they will destroy cells and endanger the health of the insect. Therefore, the elimination of excessive oxidation and free radicals can prevent related diseases. Previous studies indicated that different gut probiotics can exert their antioxidant power in different ways to maintain host health [37]. Saeedi et al. found that Lactobacillus can activate liver Nrf2 in Drosophila and mice by producing 5-methoxyindoleacetic acid, consequently achieving the “distant control” of oxidative stress in the liver and thus protecting the liver from oxidative damage induced by acetaminophen overdose and acute ethanol poisoning [38]. Elzeini et al. found that EPS (extracellular polysaccharide) generated by Enterococcus faecalis-HBE1, Lactobacillus brevis-HBE2, E. faecalis-HBE3 and E. faecalis-HBE4 isolated in the intestinal tract of Apis mellifera L. has antioxidant activity to scavenge DPPH free radicals [39]. Barretto et al. found that staphyloxanthin pigment generated by Staphylococcus gallinarum KX912244 isolated in the intestinal tract of B. mori can inhibit S. aureus, E. coli, and Candida albicans and scavenge DPPH free radicals [40]. Although the active metabolites of probiotics have better antioxidant functions, further investigation is essential to explore the mechanism of action.
2.4. Enhance Host Drug Resistance
Due to the excessive and irregular use of insecticides, insects have evolved drug resistance, which leads to non-target insect death and environmental pollution [41]. Insect drug resistance mechanisms are dominated by metabolic resistance and target resistance, and these mechanisms are primarily caused by the evolution of the host genome [41]. Studies have found that certain gut symbionts can degrade chemical pesticides, which in turn affects host resistance to pesticides. Chen et al. found that Stegotrophomonas of B. mori can increase the content of essential amino acids in the gut so that the larvae can more effectively avoid the effects of chlorpyrifos [42]. Trinder et al. found that supplementation of Lactobacillus rhamnosus in the Drosophila melanogaster diet could reduce the toxicity of organophosphorus pesticides [43]. Danilenko et al. described that Lactobacillus species of L. plantarum (strains H28, H24, KX519413, KX519414, LP8, LP25, LP86, LP95, and LP100) and Lactobacillus kunkeei were characterized by an increased antioxidant potential that protects against different pesticides in the gut of honey bees [44]. Sato et al. found that the intestinal symbiotic bacteria of Riptortus pedestris can degrade the organophosphorus insecticide into 3-methyl-4-nitrophenol, which is bactericidal but not insecticidal [45]. Itoh et al. found that the bed bug R. pedestris can acquire resistance to the insecticide phosphine (MEP) by acquiring MEP-degrading Burkholderia from the environment [46]. This indicates that the gut microbiota plays an important role in host drug resistance and that part of the gut microbiota can defend the host from chemical pesticides.
3. Factors Affecting the Gut Microbiome of Insects
3.1. Effects of Feed, Antibiotics and Culture Temperature on Insect Gut Microbes
The study finds food, antibiotics, and temperature have important effects on insect gut microbial diversity [47,48,49]. High-throughput sequencing technology was used to study the effects of Spodoptera frugiperda feeding on maize, wild oats, oilseed rape, and pepper on host gut microbial community structure and diversity. The results revealed that the gut microbial diversity of insects fed rapeseed was the highest and the gut microbial diversity of insects fed wild oats was the lowest, while the gut microbial diversity of insects fed maize without a seed coating agent was significantly higher than that with such an agent [50]. Liang et al. found that the midgut microbes of the 5th instar Helicoverpa armigera larvae feeding on lettuce leaves were significantly different from those of common silkworms at the genus level. The bacteria of the genera Acinetobacter and Anaerofilum are the main bacteria, while Bacillus and Arcobacter are the main bacteria in the normally fed H. armigera [51]. Priya et al. found that bacterial diversity varied widely between H. armigera from different host plants and the same host plant from different locations. Compared with insects that feed on crops, insects fed on artificial diets have significantly fewer gut microbial species [52]. Thakur et al. analyzed the effects of adding streptomycin to artificial diets on the survival and fitness of Spodoptera litura (Lepidoptera: Spodoptera) and its gut microbial diversity. Changes in microbial diversity were found in the guts of the larvae, with the larvae growing faster compared to the treatment group without antibiotics. The total activity of various digestive enzymes increased, and the activity of detoxification enzymes decreased significantly [48]. Wang et al. found that the overwintering stage of Dendroctonus armandi led to changes in the intestinal flora. The Proteobacteria (mainly γ-Proteobacteria) become the main phylum in the larva gut, followed by Actinobacteria and Firmicutes [53]. In addition, recent studies have found that the survival of Nezara viridula depends on intestinal symbiotic bacteria. An increase in temperature may result in a decrease in intestinal symbiotic bacteria content and host survival, and the antibiotic-treated animals also induced the same results [54]. Hence, studying the influence of the external environment on the microorganism diversity of the insect intestinal tract can be of positive significance for us to screen beneficial microorganisms and manage the rise and fall of pest populations [55].
3.2. Effects of Sex and Developmental Stage on Insect Gut Microbes
The diversity and proportion of insect gut microbes differed by sex and developmental period. The intestinal tract of male and female larvae of the 5th instar of B. mori was dominated by Enterococcus, Delftia, Pelomonas, Ralstonia, and Staphylococcus. The abundance of Enterococcus was significantly lower in female larvae than in male larvae, while the abundances of Delftia, Aurantimonas, and Staphylococcus were significantly increased [56]. Chen et al. reported that the dominant phyla Proteobacteria, Firmicutes, Actinobacteria, and Bacteroidetes were detected in the whole life history of silkworms and found that these four phyla were also present in the mulberry-eating larvae of Acronicta major and Diaphania pyloalis. The microbial community changes significantly between the early and late larvae of B. mori, consistent with host developmental changes [57]. The eggs of Spodoptera exigua are rich in Enterococcus, Pseudomonas, and Asaia, while Methylobacter and Halomonas are dominant in newly hatched larvae, and Enterococcus dominates in 3rd and 5th instar larvae. The pupal stage has the highest microbial diversity. There were no significant differences between newly hatched male and female S. exigua larvae [58]. In addition, bacteria and fungi showed dynamic changes at various developmental stages of the brown planthopper. The predominant fungal genus in the nymphal and adult stages was Wallemia, and the abundance of Acinetobacter in the egg stage was significantly reduced. The microbial community composition in female and male brown planthoppers is different and sex-dependent [59]. The above results reveal that there are different dominant flora in different developmental stages and sexes of insects, which may play an important role in different developmental stages and sexes, but the strains with specific functions need further analysis and experimental proof.
3.3. Effects of Pesticides on Insect Gut Microbes
Extensive use of pesticides will affect insects’ food digestion, nutrient absorption, metabolism, immune response, defense against pathogenic bacteria invasion, and microbial homeostasis [56,60,61]. Li et al. found that the activity and expression of enzymes related to nutrient metabolism in the midgut were unregulated, and the growth of B. mori was slow after feeding on phoxim [62]. Sun et al. studied the effects of microbial pesticides and camptothecin on the mortality of two lepidopteran insects, Trichoplusia ni and S. exigua. The bioassay results demonstrated that camptothecin significantly enhanced the toxicity of B. thuringiensis to S. exigua and T. ni, as well as to Autographa californica nucleopolyhedrovirus (AcMNPV) and S. exigua nuclear polyhedrosis virus (SeMNPV). It is speculated that camptothecin can affect the permeability of the peritrophic membrane to increase its toxicity [63]. Wei et al. studied the synergistic interaction mode of gut microbiota and B. bassiana in mosquitoes and confirmed that fungi can reduce the abundance of gut microbiota and the number of probiotics. Fungi can significantly increase the abundance of the opportunistic pathogen Serratia marcescens, which overgrows in the midgut and transfers to the hemocoel, causing systemic infections and accelerating mosquito death [60]. Kumar et al. found that the proportion of Proteobacteria increased at 48 h and 96 h but decreased after 144 h after B. mori infection with BmBDV. At the genus level, the proportion of Enterococcus increased gradually after BmBDV infection of B. mori, but the proportion of Incertae sedis increased at 96 h, while the proportion of Lactococcus decreased at 96 h. Enterococcus abundance was positively correlated with the expression levels of spatzle-1, PGRP-LE, and PGRP-LB genes, indicating that the increased abundance of Enterococcus activates the Toll and IMD immune pathways [64]. Motta et al. found that glyphosate consumption by honeybees under laboratory or field conditions disrupts the host gut microbiota and affects health [65]. Dai et al. tested the effect of glyphosate on bacterial diversity in the midgut of Italian honeybees in the laboratory. After treatment with 20 mg/L glyphosate, the species diversity and richness in the intestinal tract of honeybees changed significantly, and the survival rate of honeybees decreased [66]. Zhu et al. found that after Apis mellifera L. ingested the neonicotinoid insecticide nitenpyram, metabolism, detoxification, and immune-related genes were significantly changed, resulting in an intestinal flora imbalance, which further reduced food consumption and the survival of honey bees [67].
4. Negative Effects of Gut Microbes on the Host
Under normal circumstances, the gut microbial community is beneficial to the host. However, the consumption of pathogenic microorganisms, pesticides, and antibiotics by insects will lead to disturbance of the insect gut microbiome and endanger insect health [60,68,69,70]. Gut microbial disturbances refer to changes in the gut community structure far exceeding normal levels that put the host in an unfavorable state. Li et al. found that after Plutella xylostella fed on Bt Cry1Ac, gut microbial diversity was significantly reduced, while the relative expression of bacterial load and immune genes was significantly up-regulated, and insect death was accelerated [71]. Tan et al. found that co-infection of Paranosema locustae and B. bassiana with Locusta migratoria altered gut microbiota homeostasis and accelerated death [72]. Previous studies have shown that phoxim exposure to B. mori affected gut bacterial community composition and function [69]. Li et al. found that vaccination with sublethal doses of acetamiprid activates the Duox-ROS system and induces ROS accumulation, leading to intestinal dysbiosis and the translocation of Pseudomonas and Staphylococcus to the hemolymph. Acetamiprid-treated B. mori larvae inoculated with E. cloacae significantly reduced survival and body weight [73]. The expression of peritrophic membrane protein-related genes BmCBP-02, BmPM-41, BmPM-43, and BmCDA7, as well as the toll signaling pathway-related genes Bmcactus, Bmspatzle, and Bmrel, was significantly down-regulated in the gut of B. mori following exposure to phoxim, leading to B. mori’s susceptibility to E. cloacae [74]. Silva et al. found that G. mellonella larval mortality and immune responses were significantly increased when inoculated with sub-inhibitory concentrations of nisin and S. enterica [75].
In addition, antibiotics are frequently used to prevent bacterial infection of larval honeybees, but prolonged use of antibiotics leads to disruption of gut microbial homeostasis, increasing susceptibility to opportunistic pathogens and affecting honeybee health [76]. Meyel et al. found that after feeding on rifampicin by European earwig larvae, the homeostasis of the host’s gut microbes was disrupted, and the weight of eggs and larvae decreased [77]. Powell et al. found that Apis mellifera ingested tylosin tartrate, which significantly increased their susceptibility to S. marcescens [78]. Schretter et al. found that Drosophila feeding on antibiotics (sterile Drosophila larvae) leads to hyperactive motor behaviors. After supplementation with Lactobacillus brevis, the motor behavior of Drosophila was improved [79]. These results suggest that intestinal microbial homeostasis is crucial to host health.
Some gut commensal bacteria may be transformed into pathogenic bacteria in the presence of dysregulated insect gut microbial homeostasis, such as S. marcescens, Pseudomonas protegens Ramette, and B. cereus, which can elicit host immune responses and disrupt gut microbial homeostasis, and thus pose a serious threat to insect health [80,81,82]. S. marcescens, a symbiotic bacteria in the mosquito gut, secretes a secreted protein called SmEnhancin to facilitate arbovirus infection [75,83]. Johnson et al. found that Drosophila significantly increased the host’s susceptibility to B. bassiana after feeding on Pseudomonas protegens Ramette. After S. exigua fed on Bt GS57, gut microbial diversity was significantly reduced. Bt GS57 accelerates insect death when complexed with the gut commensal B. cereus [81]. To sum up, the interaction between conditioned pathogenic bacteria and insect intestinal microorganisms will affect their insecticidal activity in insects, which provides a new research direction into the pathogenic mechanisms of fungi/bacteria in insects.
5. Detection Methods of Insect Gut Microbes
5.1. Traditional Identification Methods
The traditional identification methods of gut microbes refer to methods that utilize morphological, physiological, and biochemical characteristics as evaluation criteria. For a long time, the identification of gut microbes has always followed the identification of species by purification culture, morphological, physiological, and biochemical characteristics [84,85,86] (Table 2). At present, the traditional culture detection method needs to inoculate the isolated microorganisms in the NA medium, LB medium, BHI medium, KIA medium, LIA medium, and TSI medium. Due to the limitations of medium components and culture conditions, many insect gut bacteria cannot be cultured [87,88]. Anjum et al. used biochemical analysis, 16S rDNA sequencing, and bioinformatics to identify 150 aerobic or facultative anaerobic bacteria from the guts of 45 worker bees. It was found that there are mainly Staphylococcus, Bacillus, Enterococcus, Corynebacterium, and Micrococcus in the intestinal tract of honeybees. The isolated bacteria are resistant to acidic environments and ferment sugars, which are beneficial to the survival of bees [89]. Khan et al. used high-throughput sequencing and found that the gut of honeybees contained (Firmicutes, Proteobacteria, Actinobacteria, Flavobacteriia, and Mollicutes) bacteria and fungi (Dothideomycetes, Eurotiomycetes, Mucormycotina, Saccharomycetes, Zygomycetes, Yeasts, and Molds) [8]. Broderick et al., using traditional culture methods, found that culturable microorganisms in the gut of Lymantria dispar larvae accounted for more than 40% of the bacteria identified by sequencing [9]. Therefore, the types of gut bacteria identified by traditional culture methods are very limited, which has great limitations for revealing the diversity and composition of insect gut microbes.
5.2. Electrophoresis
With the development of DNA labeling technology, many new molecular markers have been used for the classification and identification of bacteria, such as restriction/amplified fragment length polymorphism (RFLP/AFLP), denatured gradient gel electrophoresis (DGGE), amplified rDNA restriction analysis (ARDRA), and pulsed-field gel electrophoresis (PFGE). Both RFLP and ARDRA require the use of endonucleases to digest specific PCR amplification products and then obtain fragments of different numbers and lengths to distinguish bacteria. This method is suitable for strains that have been successfully isolated and can determine whether there are differences in the genus of a large number of strains [90,91] (Table 2). He et al. identified 12 and 11 species in the Camponotus midgut, respectively, utilizing traditional culture methods and 16S rRNA-RFLP technology [92]. Dec et al. used 16S-ARDRA and MALDI-TOF technology to isolate 80 Lactobacillus strains from the cloaca of chicken, goose, and turkey, reaching the species level [93]. AFLP generally requires a pair of endonucleases to digest genomic DNA and then add artificial linkers at both ends of the digested product. Using this as a DNA template, specific primers are used to amplify the DNA. Identification of bacteria by analysis of the different DNA fragments generated by gel electrophoresis [94]. Lindstedt et al. found that AFLP had a better discriminative ability in identifying C. jejuni strains than PFGE and PCR-RFLP [95]. DGGE is a technology that separates DNA fragments of the same size with different base compositions, and the melting behavior of the deforming agent at different concentrations is different, resulting in different mobility, so as to separate them [96]. Lin et al. used PCR-DGGE technology to successfully obtain 15 different bands in the study of the microbial diversity in the gut of the diamondback moth P. xylostella. Phylogenetic tree analysis revealed that the dominant bacteria in the gut of 4th instar P. xylostella larvae belonged to Actinobacteri, Proteobacteria, and Firmicutes [97].
5.3. Molecular Detection Method Based on 16S rRNA Gene
At present, microorganisms can be more accurately distinguished according to the variable region in the bacterial 16S rRNA gene sequence, fungal 18S rRNA, or internal transcribed spacer (ITS) sequence. Among them, the 16S rRNA gene is a DNA sequence corresponding to the ribosome 30S small subunit in prokaryotic cells, which is present in all bacterial genomes. The 16S rRNA gene is highly conserved and specific in all bacteria, and universal primers can be used to identify the species of microorganisms. Specific primers or probes can also be designed based on the variable region of the 16S rRNA gene to identify specific strains (Table 2). Snyman et al. collected 78 bacterial strains from the midgut of Busseola fusca larvae from 30 sites in South African maize production areas and used 16S rRNA gene sequencing to identify them. The results revealed that the midgut of B. fusca larvae mainly contained three phyla, Proteobacteria, Actinobacteria, and Firmicutes, and 20 species from 15 genera, including Bacillus, Enterococcus, and Klebsiella [98]. Yadav et al. collected Aedes larvae and pupae in India and used 16S1/16S2 primers to identify the isolated gut bacteria. Twenty-four strains of Aedes larval gut bacteria were identified as belonging to four phyla: Proteobacteria, Firmicutes, Bacteroidetes, and Actinobacteria [99]. Based on the 16S rRNA V4 hypervariable region, the 515F/806R primers were used to identify the secondary commensal bacteria Hamiltonella in the intestinal tract of Bemisia tabaci MED [100]. When the Ham-F/R primers were used to detect the intestinal bacteria of B. tabaci by ordinary PCR, the secondary symbiotic bacteria Hamiltonella was detected in vivo [101,102]. However, Su et al. used 341F/805R primers to molecularly identify the intestinal microbes in B. tabaci MEAM1 but did not discover Hamiltonella in its body [103]. However, 16S rRNA amplicon sequencing analysis is usually limited to relative community insights that do not even represent relative abundances well due to extraction and amplification biases. Recent method additions involve the addition of internal and external standards to account for this shortcoming. Therefore, based on the 16S rRNA gene sequence, the preference of the designed primers may lead to different abundances in some bacteria.
5.4. qRT-PCR
qRT-PCR is based on the continuous accumulation of PCR reaction products, and the amount of fluorescence increases proportionally. The amount of nucleic acid is determined by detecting the amount of fluorescence in the sample. Regarding qRT-PCR, I consider that the inclusion of controls and independent confirmation of species/strain identification is of the highest importance, e.g., by (high-resolution) melting-curve analysis and sequencing of PCR products. Quantification beyond presence/absence requires standards for each species/strain, but a further improvement is represented by digital PCRs, which really quantify the presence/absence of template molecules (Table 2). Wei et al. used qRT-PCR technology to discover that the relative expression level of S. marcescens in the gut of Anopheles was significantly up-regulated after Duox gene silencing [60]. Tong et al. used qRT-PCR technology to detect Bacteroides fragilis in clinical specimens. The results demonstrated that the target bacteria were detected by qRT-PCR from 132 samples (33%), which was much higher than the target bacteria detected by the culture method from 31 samples (8%) [104]. Sedgley et al. used qRT-PCR to detect E. faecalis in the provided samples. The results revealed that qRT-PCR detected the target bacteria from five samples (17%), which was much higher than the target bacteria detected from two samples (7%) in the parallel culture experiment [105]. This indicates that qRT-PCR is a rapid and sensitive detection method for identifying bacterial species.

{kind=link}
Table 2.
Detection methods, advantages and disadvantages, and applications of intestinal microorganisms.
Table 2.
Detection methods, advantages and disadvantages, and applications of intestinal microorganisms.
| Techniques | Advantages | Disadvantages | Application | References |
|---|---|---|---|---|
| Traditional culture method | Low-cost. | Many bacteria cannot be cultured because of the limitations of culture medium and culture condition. | Staphylococcus, Bacillus, Enterococcus, Corynebacterium, Micrococcus Dothideomycetes, Eurotiomycetes, Mucormycotina, Saccharomycetes, Zygomycetes, Yeasts, Molds | [8,87,88,89] |
| Electrophoresis | Rapid analysis of a large number of specimens. | Suitable for small DNA fragments, and can only reflect the dominant genus. | Lactobacillus, Campylobacter jejuni | [93,95,97] |
| Molecular detection method based on 16S rRNA gene | Generally accurate to the level of genus, a few can be identified to species. For some bacteria, the similarity of sequences in hypervariable regions is very high. | Other methods and techniques are needed to identify species with small interspecific differences. | Bacillus, Enterococcus, Klebsiella, Hamiltonella | [98,100,101,102] |
| qRT-PCR | The method is simple, rapid, quantifiable, repeatable, sensitive, and specific and can be used to detect live and dead bacteria. | High cost, strict requirements for the operation of laboratory personnel, the need for specific instruments and reagents. | Serratia marcescens, Bacteroides fragilis, Enterococcus faecalis | [60,104,105] |
| Metagenomics | Metagenomics overcomes the technical limitations of traditional pure culture methods and can provide information about low abundance or even trace microorganisms in the environment, which can more accurately reflect the true state of microbial survival. | The sequencing data are large, the price is much more expensive than 16S sequencing, and the computing resources required for subsequent data processing are high. | Major Phylum: Firmicutes, Proteobacteria, Elusimicrobia, Mycobacterium, Bacteroidetes Major Genus: Enterococcus, Pantoea, Acinetobacter, Enterobacteriales, Lactobacillales | [106,107,108,109,110] |
5.5. Metagenomics
Metagenomic technology refers to a new technology for studying the structure and function of the microbial community in a sample. Metagenomics can be challenged by biases during extraction, amplification, and subsequent assembly steps (e.g., Gram-positive bacteria with a robust cell wall and species/strains with extremely high/low GC content) (Table 2). The research found that the intestinal microbes of Termites are dominated by Firmicutes, Mycobacterium, and Bacteroidetes, and Enterobacteriaceae and Bacteroidetes are the dominant genera [106]. Firmicutes and Proteobacteria are the dominant phyla in the intestinal microbes of the Lepidopteran insects S. exigua and S. litura. S. exigua is dominated by Enterococcus of the family Enterococcus [107,109]. S. litura is dominated by Pantoea and Acinetobacter [110]. Insect gut microbes were significantly different from other Lepidopteran insects at the genus level. Therefore, differences at the genus level can be used as an important indicator for evaluating microbial diversity.
6. Application of Insect Intestinal Microflora in Pest Management
Commensal bacteria in the insect gut play an important role in enhancing host energy metabolism, immune defense, and resistance to infection by pathogenic microorganisms [14,27,111]. Therefore, in recent years, more and more studies have focused on the function of intestinal flora. It is reported that B. thuringiensis kurstaki can induce the mortality of Lymantri dispar, which depends on the intestinal bacteria of the host [112]. Paramasiva et al. found that the elimination of intestinal flora in H. armigera affected the sensitivity of B. thuringiensis to H. armigera [113]. However, the insecticidal activity of the B. thuringiensis HD-73 strain against Manduca sexta and the insecticidal activity of diverse doses of B. thuringiensis HD-1 and HD-73 strains against P. xylostella larvae indicate that the presence of gut microbes is not essential [114]. Since then, various methods have been used to demonstrate the role of insect gut microbes in the pathogenicity of B. thuringiensis, but until now, it has been hard to reach a consistent conclusion.
Insect intestinal symbiotic bacteria have been isolated and purified from different insects, and the functions of a few isolated bacteria have been reported. In mosquitoes, the intestinal symbiotic bacteria S. marcescens can significantly promote the tolerance of mosquitoes to arboviruses by secreting toxin proteins [60]. Wei et al. also confirmed that the interaction between B. bassiana and mosquito intestinal microbes can accelerate the death of mosquitoes [60]. Ren et al. found that antibiotics secreted by B. cereus can decrease the number of symbiotic intestinal flora in the co-infection experiment of B. cereus and B. thuringiensis and promote the infection of B. thuringiensis [115]. Therefore, research on the function of conditionally pathogenic bacteria in the insect intestine may be an important research direction to increase the insecticidal activity of B. thuringiensis.
In addition, B. thuringiensis can generate Cry toxin protein with insecticidal activity or δ-endotoxin, which, when combined with receptors on intestinal epithelial cells, can induce intestinal epithelial cells to form holes, and intestinal microorganisms will penetrate the blood cavity [116,117,118]. It is reported that Enterococcus and Bacillus, as the dominant bacteria in the gut of S. exigua, are easily able to cause sepsis when they escape to the blood cavity [119]. Mason et al. found that after the larvae of the M. sexta were infected with B. thuringiensis, Enterococcus faecalis would translocate to the blood cavity and induce sepsis, increasing the sensitivity of the M. sexta to B. thuringiensis [120]. These results indicate that the translocation of intestinal symbiotic bacteria to the hemocoel is an important factor causing insect death.
7. Conclusions
Insect guts are inhabited by a large number of microorganisms and are rich in microbial resources. The gut microbiota is essential for host nutrient metabolism, immune defense, and drug resistance, and trials using insects as models have revealed that changes in microbiota structure can affect host health (Figure 1). Intestinal microbes also include viruses, bacteriophages, eukaryotic microbes, etc. The functional studies of these microbes are not yet perfect, and further in-depth research is needed. In addition, the details of the interaction between the insect immune system and the gut microbiota still need to be further explored.
At present, important progress has been made in the study of the taxonomic composition of insect intestinal microorganisms and their effects on growth and development. However, there are abundant microorganisms on the surface of insect skin, feces, and living environments, which can also affect the health of insects. The relationship between insect growth environment and host intestinal flora requires further exploration. Therefore, the research in this field will assist us in better understanding how to control intestinal flora according to the living environment, diet, and physiological changes of insects and will further promote our understanding of insect intestinal microbial functions, thus providing new opportunities for enhancing the overall health of beneficial insects and the management of harmful insects.
Author Contributions
Y.L. writing—original draft; Y.L., L.C. and K.X. investigation, resources; S.Z. writing—review and editing; F.G. and Y.F. conceptualization, supervision, writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financially supported by the Natural Science Foundation of China (22078171), S&T Program of Hebei (21374301D) and the Scientific Research Foundation of Tangshan Normal University (2023B08).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful to all lab members for helpful discussions and suggestions in this review writing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Krams, I.A.; Kecko, S.; Joers, P.; Trakimas, G.; Elferts, D.; Krams, R.; Luoto, S.; Rantala, M.J.; Inashkina, I.; Gudra, D.; et al. Microbiome symbionts and diet diversity incur costs on the immune system of insect larvae. J. Exp. Biol. 2017, 220, 4204–4212. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, F.; Lu, X. Diversity and functional roles of the gut microbiota in lepidopteran insects. Microorganisms. 2022, 10, 1234. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Kikuchi, Y. Impact of the insect gut microbiota on ecology, evolution, and industry. Curr. Opin. Insect Sci. 2020, 41, 33–39. [Google Scholar] [CrossRef]
- Lee, W.J.; Hase, K. Gut microbiota-generated metabolites in animal health and disease. Nat. Chem. Biol. 2014, 10, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Whitten, M.M.; Facey, P.D.; Del, S.R.; Fernandez-Martinez, L.T.; Evans, M.C.; Mitchell, J.J.; Bodger, O.G.; Dyson, P.J. Symbiont-mediated RNA interference in insects. Proc. Biol. Sci. 2016, 283, 20160042. [Google Scholar] [CrossRef]
- Blow, F.; Bueno, E.; Clark, N.; Zhu, D.T.; Chung, S.H.; Gullert, S.; Schmitz, R.A.; Douglas, A.E. B-vitamin nutrition in the pea aphid-Buchnera symbiosis. J. Insect Physiol. 2020, 126, 104092. [Google Scholar] [CrossRef]
- Wier, A.; Dolan, M.; Grimaldi, D.; Guerrero, R.; Wagensberg, J.; Margulis, L. Spirochete and protist symbionts of a termite (Mastotermes electrodominicus) in Miocene amber. Proc. Natl. Acad. Sci. USA 2002, 99, 1410–1413. [Google Scholar] [CrossRef]
- Khan, K.A.; Al-Ghamdi, A.A.; Ghramh, H.A.; Ansari, M.J.; Ali, H.; Alamri, S.A.; Al-Kahtani, S.N.; Adgaba, N.; Qasim, M.; Hafeez, M. Structural diversity and functional variability of gut microbial communities associated with honey bees. Microb. Pathog. 2020, 138, 103793. [Google Scholar] [CrossRef]
- Broderick, N.A.; Raffa, K.F.; Goodman, R.M.; Handelsman, J. Census of the bacterial community of the gypsy moth larval midgut by using culturing and culture-independent methods. Appl. Environ. Microbiol. 2004, 70, 293–300. [Google Scholar] [CrossRef]
- Settanni, L.; Guarcello, R.; Gaglio, R.; Francesca, N.; Aleo, A.; Felis, G.E.; Moschetti, G. Production, stability, gene sequencing and in situ anti-Listeria activity of mundticin KS expressed by three Enterococcus mundtii strains. Food Control. 2014, 35, 311–322. [Google Scholar] [CrossRef]
- Jing, T.Z.; Qi, F.H.; Wang, Z.Y. Most dominant roles of insect gut bacteria: Digestion, detoxification, or essential nutrient provision? Microbiome 2020, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Yang, L.; Pang, X.; Zhang, R.; Zhu, Y.; Wang, P.; Gao, G.; Cheng, G. A Mesh-Duox pathway regulates homeostasis in the insect gut. Nat. Microbiol. 2017, 2, 17020. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Sanders, J.G.; Lukasik, P.; D’Amelio, C.L.; Millar, J.S.; Vann, D.R.; Lan, Y.; Newton, J.A.; Schotanus, M.; Kronauer, D.; et al. Herbivorous turtle ants obtain essential nutrients from a conserved nitrogen-recycling gut microbiome. Nat. Commun. 2018, 9, 964. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.A.; Vennison, S.J.; Sankar, S.G.; Prabhu, D.I.; Vasan, P.T.; Raghuraman, T.; Geoffrey, C.J.; Vendan, S.E. Isolation and characterization of bacteria from the gut of Bombyx mori that degrade cellulose, xylan, pectin and starch and their impact on digestion. J. Insect Sci. 2010, 10, 107. [Google Scholar] [CrossRef]
- Su, L.; Liu, H.; Li, Y.; Zhang, H.; Chen, M.; Gao, X.; Wang, F.; Song, A. Cellulolytic activity and structure of symbiotic bacteria in locust guts. Genet. Mol. Res. 2014, 13, 7926–7936. [Google Scholar] [CrossRef]
- Guégan, M.; Tran Van, V.; Martin, E.; Minard, G.; Tran, F.H.; Fel, B.; Hay, A.E.; Simon, L.; Barakat, M.; Potier, P.; et al. Who is eating fructose within the Aedes albopictus gut microbiota? Environ. Microbiol. 2020, 22, 1193–1206. [Google Scholar] [CrossRef]
- Tokuda, G.; Lo, N.; Watanabe, H. Marked variations in patterns of cellulase activity against crystalline-vs. carboxymethyl-cellulose in the digestive systems of diverse, wood-feeding termites. Physiol. Entomol. 2005, 30, 372–380. [Google Scholar] [CrossRef]
- Dantur, K.I.; Enrique, R.; Welin, B.; Castagnaro, A.P. Isolation of cellulolytic bacteria from the intestine of Diatraea saccharalis larvae and evaluation of their capacity to degrade sugarcane biomass. AMB Express 2015, 5, 15. [Google Scholar] [CrossRef]
- Zheng, H.; Powell, J.E.; Steele, M.I.; Dietrich, C.; Moran, N.A. Honeybee gut microbiota promotes host weight gain via bacterial metabolism and hormonal signaling. Proc. Natl. Acad. Sci. USA 2017, 114, 4775–4780. [Google Scholar] [CrossRef]
- Engel, P.; Moran, N.A. The gut microbiota of insects—diversity in structure and function. Fems Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef]
- Duplais, C.; Sarou-Kanian, V.; Massiot, D.; Hassan, A.; Perrone, B.; Estevez, Y.; Wertz, J.T.; Martineau, E.; Farjon, J.; Giraudeau, P.; et al. Gut bacteria are essential for normal cuticle development in herbivorous turtle ants. Nat. Commun. 2021, 12, 676. [Google Scholar] [CrossRef] [PubMed]
- Ludington, W.B.; Ja, W.W. Drosophila as a model for the gut microbiome. PLoS Pathog. 2020, 16, e1008398. [Google Scholar] [CrossRef]
- Wong, A.C.; Wang, Q.P.; Morimoto, J.; Senior, A.M.; Lihoreau, M.; Neely, G.G.; Simpson, S.J.; Ponton, F. Gut microbiota modifies olfactory-guided microbial preferences and foraging decisions in Drosophila. Curr. Biol. 2017, 27, 2397–2404. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Keesey, I.W.; Hansson, B.S.; Knaden, M. Gut microbiota affects development and olfactory behavior in Drosophila melanogaster. J. Exp. Biol. 2019, 222, jeb192500. [Google Scholar] [CrossRef] [PubMed]
- Habineza, P.; Muhammad, A.; Ji, T.; Xiao, R.; Yin, X.; Hou, Y.; Shi, Z. The promoting effect of gut microbiota on growth and development of red palm weevil, Rhynchophorus ferrugineus (Olivier) (Coleoptera: Dryophthoridae) by modulating its nutritional metabolism. Front. Microbiol. 2019, 10, 1212. [Google Scholar] [CrossRef] [PubMed]
- Ben-Yosef, M.; Pasternak, Z.; Jurkevitch, E.; Yuval, B. Symbiotic bacteria enable olive flies (Bactrocera oleae) to exploit intractable sources of nitrogen. J. Evol. Biol. 2014, 27, 2695–2705. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.W.; Lu, Z.Q. Midgut immune responses induced by bacterial infection in the silkworm, Bombyx mori. J. Zhejiang Univ. Sci. B 2015, 16, 875–882. [Google Scholar] [CrossRef]
- Zeng, T.; Jaffar, S.; Xu, Y.; Qi, Y. The intestinal immune defense system in insects. Int. J. Mol. Sci. 2022, 23, 15132. [Google Scholar] [CrossRef]
- Shao, Y.; Chen, B.; Sun, C.; Ishida, K.; Hertweck, C.; Boland, W. Symbiont-derived antimicrobials contribute to the control of the Lepidopteran gut microbiota. Cell Chem. Biol. 2017, 24, 66–75. [Google Scholar] [CrossRef]
- Akbar, N.; Siddiqui, R.; Iqbal, M.; Sagathevan, K.; Khan, N.A. Gut bacteria of cockroaches are a potential source of antibacterial compound(s). Lett. Appl. Microbiol. 2018, 66, 416–426. [Google Scholar] [CrossRef]
- Knight, C.A.; Bowman, M.J.; Frederick, L.; Day, A.; Lee, C.; Dunlap, C.A. The first report of antifungal lipopeptide production by a Bacillus subtilis subsp. inaquosorum strain. Microbiol. Res. 2018, 216, 40–46. [Google Scholar] [CrossRef]
- Douglas, A.E. The molecular basis of bacterial-insect symbiosis. J. Mol. Biol. 2014, 426, 3830–3837. [Google Scholar] [CrossRef]
- Pandey, N.; Rajagopal, R. Tissue damage induced midgut stem cell proliferation and microbial dysbiosis in Spodoptera litura. FEMS Microbiol. Ecol. 2017, 93, fix132. [Google Scholar] [CrossRef]
- Muhammad, A.; Habineza, P.; Ji, T.; Hou, Y.; Shi, Z. Intestinal microbiota confer protection by priming the immune system of red palm weevil Rhynchophorus ferrugineus Olivier (Coleoptera: Dryophthoridae). Front. Physiol. 2019, 10, 1303. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Song, X.; Wang, J. Gut microbiota is essential in PGRP-LA regulated immune protection against Plasmodium berghei infection. Parasites Vectors. 2020, 13, 3. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Kim, K.M.; Lee, J.H.; Seo, S.J.; Lee, I.H. Extracellular gelatinase of Enterococcus faecalis destroys a defense system in insect hemolymph and human serum. Infect. Immun. 2007, 75, 1861–1869. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant properties of probiotic bacteria. Nutrients. 2017, 9, 521. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, B.J.; Liu, K.H.; Owens, J.A.; Hunter-Chang, S.; Camacho, M.C.; Eboka, R.U.; Chandrasekharan, B.; Baker, N.F.; Darby, T.M.; Robinson, B.S.; et al. Gut-resident Lactobacilli activate hepatic Nrf2 and protect against oxidative liver injury. Cell. Metab. 2020, 31, 956–968. [Google Scholar] [CrossRef] [PubMed]
- Elzeini, H.M.; Ali, A.; Nasr, N.F.; Hassan, M.; Hassan, A.; Elenany, Y.E. Probiotic capability of novel lactic acid bacteria isolated from worker honey bees gut microbiota. FEMS Microbiol. Lett. 2021, 368, fnab030. [Google Scholar] [CrossRef]
- Barretto, D.A.; Vootla, S.K. In vitro anticancer activity of Staphyloxanthin pigment extracted from Staphylococcus gallinarum KX912244, a gut microbe of Bombyx mori. Indian J. Microbiol. 2018, 58, 146–158. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Hayatsu, M.; Hosokawa, T.; Nagayama, A.; Tago, K.; Fukatsu, T. Symbiont-mediated insecticide resistance. Proc. Natl. Acad. Sci. USA. 2012, 109, 8618–8622. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, N.; Xie, S.; Zhang, X.; He, J.; Muhammad, A.; Sun, C.; Lu, X.; Shao, Y. Gut bacteria of the silkworm Bombyx mori facilitate host resistance against the toxic effects of organophosphate insecticides. Environ. Int. 2020, 143, 105886. [Google Scholar] [CrossRef] [PubMed]
- Trinder, M.; Mcdowell, T.W.; Daisley, B.A.; Ali, S.N.; Leong, H.S.; Sumarah, M.W.; Reid, G. Probiotic Lactobacillus rhamnosus reduces organophosphate pesticide absorption and toxicity to Drosophila melanogaster. Appl. Environ. Microbiol. 2016, 82, 6204–6213. [Google Scholar] [CrossRef]
- Danilenko, V.N.; Devyatkin, A.V.; Marsova, M.V.; Shibilova, M.U.; Ilyasov, R.A.; Shmyrev, V.I. Common inflammatory mechanisms in COVID-19 and parkinson’s diseases: The Role of microbiome, pharmabiotics and postbiotics in their prevention. J. Iamm. Res. 2021, 14, 6349–6381. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Jang, S.; Takeshita, K.; Itoh, H.; Koike, H.; Tago, K.; Hayatsu, M.; Hori, T.; Kikuchi, Y. Insecticide resistance by a host-symbiont reciprocal detoxification. Nat. Commun. 2021, 12, 6432. [Google Scholar] [CrossRef]
- Itoh, H.; Hori, T.; Sato, Y.; Nagayama, A.; Tago, K.; Hayatsu, M.; Kikuchi, Y. Infection dynamics of insecticide-degrading symbionts from soil to insects in response to insecticide spraying. ISME J. 2018, 12, 909–920. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.A.; Moreau, C.S.; Goldman-Huertas, B.; Fujiwara, M.; Lohman, D.J.; Pierce, N.E. Bacterial gut symbionts are tightly linked with the evolution of herbivory in ants. Proc. Natl. Acad. Sci. USA 2009, 106, 21236–21241. [Google Scholar] [CrossRef]
- Thakur, A.; Dhammi, P.; Saini, H.S.; Kaur, S. Effect of antibiotic on survival and development of Spodoptera litura (Lepidoptera: Noctuidae) and its gut microbial diversity. Bull. Entomol. Res. 2016, 106, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Raza, M.F.; Wang, Y.; Cai, Z.; Bai, S.; Yao, Z.; Awan, U.A.; Zhang, Z.; Zheng, W.; Zhang, H. Gut microbiota promotes host resistance to low-temperature stress by stimulating its arginine and proline metabolism pathway in adult Bactrocera dorsalis. PLoS Pathog. 2020, 16, e1008441. [Google Scholar] [CrossRef]
- Lv, D.; Liu, X.; Dong, Y.; Yan, Z.; Zhang, X.; Wang, P.; Yuan, X.; Li, Y. Comparison of Gut Bacterial Communities of Fall Armyworm (Spodoptera frugiperda) Reared on Different Host Plants. Int. J. Mol. Sci. 2021, 22, 11266. [Google Scholar] [CrossRef]
- Liang, X.; Fu, Y.; Tong, L.; Liu, H. Microbial shifts of the silkworm larval gut in response to lettuce leaf feeding. Appl. Microbiol. Biotechnol. 2014, 98, 3769–3776. [Google Scholar] [CrossRef]
- Priya, N.G.; Ojha, A.; Kajla, M.K.; Raj, A.; Rajagopal, R. Host plant induced variation in gut bacteria of Helicoverpa armigera. PLoS ONE 2012, 7, e30768. [Google Scholar]
- Wang, J.; Chen, H.; Tang, M. Community structure of gut bacteria of Dendroctonus armandi (Coleoptera: Curculionidae: Scolytinae) larvae during overwintering stage. Sci. Rep. 2017, 7, 14242. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Tada, A.; Musolin, D.L.; Hari, N.; Hosokawa, T.; Fujisaki, K.; Fukatsu, T. Collapse of insect gut symbiosis under simulated climate change. Mbio 2016, 7, e01578-16. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Nair, S. Dynamics of insect-microbiome interaction influence host and microbial symbiont. Front. Microbiol. 2020, 11, 1357. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Lu, Y.; Zhang, H.; Kumar, D.; Liu, B.; Gong, Y.; Zhu, M.; Zhu, L.; Liang, Z.; Kuang, S.; et al. Effects of BmCPV infection on silkworm Bombyx mori intestinal bacteria. PLoS ONE 2016, 11, e146313. [Google Scholar] [CrossRef]
- Chen, B.; Du, K.; Sun, C.; Vimalanathan, A.; Liang, X.; Li, Y.; Wang, B.; Lu, X.; Li, L.; Shao, Y. Gut bacterial and fungal communities of the domesticated silkworm (Bombyx mori) and wild mulberry-feeding relatives. ISME J. 2018, 12, 2252–2262. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Li, W.; Luo, J.; Zhang, L.; Ji, J.; Zhu, X.; Wang, L.; Zhang, S.; Cui, J. Biodiversity of the microbiota in Spodoptera exigua (Lepidoptera: Noctuidae). J. Appl. Microbiol. 2019, 126, 1199–1208. [Google Scholar] [CrossRef]
- Wang, Z.L.; Wang, T.Z.; Zhu, H.F.; Pan, H.B.; Yu, X.P. Diversity and dynamics of microbial communities in brown planthopper at different developmental stages revealed by high-throughput amplicon sequencing. Insect Sci. 2020, 27, 883–894. [Google Scholar] [CrossRef]
- Wei, G.; Lai, Y.; Wang, G.; Chen, H.; Li, F.; Wang, S. Insect pathogenic fungus interacts with the gut microbiota to accelerate mosquito mortality. Proc. Natl. Acad. Sci. USA 2017, 114, 5994–5999. [Google Scholar] [CrossRef]
- Gu, Z.; Li, F.; Hu, J.; Ding, C.; Wang, C.; Tian, J.; Xue, B.; Xu, K.; Shen, W.; Li, B. Sublethal dose of phoxim and Bombyx mori nucleopolyhedrovirus interact to elevate silkworm mortality. Pest Manag. Sci. 2017, 73, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Hu, J.; Tian, J.; Xu, K.; Ni, M.; Wang, B.; Shen, W.; Li, B. Effects of phoxim on nutrient metabolism and insulin signaling pathway in silkworm midgut. Chemosphere 2016, 146, 478–485. [Google Scholar] [CrossRef]
- Sun, S.; Cheng, Z.; Fan, J.; Cheng, X.; Pang, Y. The utility of camptothecin as a synergist of Bacillus thuringiensis var. kurstaki and nucleopolyhedroviruses against Trichoplusia ni and Spodoptera exigua. J. Econ. Entomol. 2012, 105, 1164–1170. [Google Scholar] [CrossRef]
- Kumar, D.; Sun, Z.; Cao, G.; Xue, R.; Hu, X.; Gong, C. Bombyx mori bidensovirus infection alters the intestinal microflora of fifth instar silkworm (Bombyx mori) larvae. J. Invertebr. Pathol. 2019, 163, 48–63. [Google Scholar] [CrossRef]
- Motta, E.; Mak, M.; De Jong, T.K.; Powell, J.E.; O’Donnell, A.; Suhr, K.J.; Riddington, I.M.; Moran, N.A. Oral or topical exposure to glyphosate in herbicide formulation impacts the gut microbiota and survival rates of Honey Bees. Appl. Environ. Microbiol. 2020, 86, e01150-20. [Google Scholar] [CrossRef] [PubMed]
- Dai, P.; Yan, Z.; Ma, S.; Yang, Y.; Wang, Q.; Hou, C.; Wu, Y.; Liu, Y.; Diao, Q. The herbicide glyphosate negatively affects midgut bacterial communities and survival of honey bee during larvae reared in vitro. J. Agric. Food Chem. 2018, 66, 7786–7793. [Google Scholar] [CrossRef]
- Zhu, L.; Qi, S.; Xue, X.; Niu, X.; Wu, L. Nitenpyram disturbs gut microbiota and influences metabolic homeostasis and immunity in honey bee (Apis mellifera L.). Environ. Pollut. 2020, 258, 113671. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.R.; Liu, Q.Z. Evaluation and entomopathogenicity of gut bacteria associated with dauer juveniles of Oscheius chongmingensis (Nematoda: Rhabditidae). Microbiol. Open 2019, 8, e823. [Google Scholar] [CrossRef]
- Li, F.; Li, M.; Mao, T.; Wang, H.; Chen, J.; Lu, Z.; Qu, J.; Fang, Y.; Gu, Z.; Li, B. Effects of phoxim exposure on gut microbial composition in the silkworm, Bombyx mori. Ecotox. Environ. Saf. 2020, 189, 110011. [Google Scholar] [CrossRef]
- Raymann, K.; Bobay, L.M.; Moran, N.A. Antibiotics reduce genetic diversity of core species in the honeybee gut microbiome. Mol. Ecol. 2018, 27, 2057–2066. [Google Scholar] [CrossRef]
- Li, S.; Xu, X.; De Mandal, S.; Shakeel, M.; Hua, Y.; Shoukat, R.F.; Fu, D.; Jin, F. Gut microbiota mediate Plutella xylostella susceptibility to Bt Cry1Ac protoxin is associated with host immune response. Environ. Pollut. 2021, 271, 116271. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.Q.; Yin, Y.; Cao, K.L.; Zhao, X.X.; Wang, X.Y.; Zhang, Y.X.; Shi, W.P. Effects of a combined infection with Paranosema locustae and Beauveria bassiana on Locusta migratoria and its gut microflora. Insect Sci. 2021, 28, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, M.; Zhu, Q.; Mao, T.; Dai, M.; Ye, W.; Bian, D.; Su, W.; Feng, P.; Ren, Y.; et al. Imbalance of intestinal microbial homeostasis caused by acetamiprid is detrimental to resistance to pathogenic bacteria in Bombyx mori. Environ. Pollut. 2021, 289, 117866. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, M.; Wang, H.; Mao, T.; Chen, J.; Lu, Z.; Qu, J.; Fang, Y.; Li, B. Effects of phoxim pesticide on the immune system of silkworm midgut. Pest. Biochem. Physiol. 2020, 164, 58–64. [Google Scholar] [CrossRef]
- Silva, F.; Fernandes, K.M.; Freitas, L.L.; Cascardo, R.S.; Bernardes, R.C.; Oliveira, L.L.; Martins, G.F.; Vanetti, M. Effects of sub-lethal doses of nisin on the virulence of Salmonella enterica in Galleria mellonella larvae. Res. Microbiol. 2021, 172, 103836. [Google Scholar] [CrossRef]
- Raymann, K.; Shaffer, Z.; Moran, N.A. Antibiotic exposure perturbs the gut microbiota and elevates mortality in honeybees. PLoS Biol. 2017, 15, e2001861. [Google Scholar] [CrossRef]
- Van Meyel, S.; Devers, S.; Dupont, S.; Dedeine, F.; Meunier, J. Alteration of gut microbiota with a broad-spectrum antibiotic does not impair maternal care in the European earwig. J. Evol. Biol. 2021, 34, 1034–1045. [Google Scholar] [CrossRef]
- Powell, J.E.; Carver, Z.; Leonard, S.P.; Moran, N.A. Field-realistic tylosin exposure impacts honey bee microbiota and pathogen susceptibility, which is ameliorated by native gut probiotics. Microbiol. Spectr. 2021, 9, e10321. [Google Scholar] [CrossRef]
- Schretter, C.E.; Vielmetter, J.; Bartos, I.; Marka, Z.; Marka, S.; Argade, S.; Mazmanian, S.K. A gut microbial factor modulates locomotor behaviour in Drosophila. Nature 2018, 563, 402–406. [Google Scholar] [CrossRef]
- Stathopoulos, S.; Neafsey, D.E.; Lawniczak, M.K.; Muskavitch, M.A.; Christophides, G.K. Genetic dissection of Anopheles gambiae gut epithelial responses to Serratia marcescens. PLoS Pathog. 2014, 10, e1003897. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, D.; Wu, H.; Ji, Y.; Liu, Z.; Guo, X.; Guo, W.; Bi, Y. Bt GS57 interaction with gut microbiota accelerates Spodoptera exigua mortality. Front. Microbiol. 2022, 13, 835227. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.M.; Weeks, E.; Lovullo, E.D.; Shirk, P.D.; Geden, C.J. Mortality effects of three bacterial pathogens and Beauveria bassiana when topically applied or injected into house flies (Diptera: Muscidae). J. Med. Entomol. 2019, 56, 774–783. [Google Scholar] [CrossRef]
- Wu, P.; Sun, P.; Nie, K.; Zhu, Y.; Shi, M.; Xiao, C.; Liu, H.; Liu, Q.; Zhao, T.; Chen, X.; et al. A gut commensal bacterium promotes mosquito permissiveness to arboviruses. Cell Host Microbe 2019, 25, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Eski, A.; Demir, İ.; Güllü, M.; Demirbağ, Z. Biodiversity and pathogenicity of bacteria associated with the gut microbiota of beet armyworm, Spodoptera exigua Hübner (Lepidoptera: Noctuidae). Microb. Pathog. 2018, 121, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Secil, E.S.; Sevim, A.; Demirbag, Z.; Demir, I. Isolation, characterization and virulence of bacteria from Ostrinia nubilalis (Lepidoptera: Pyralidae). Biologia 2012, 67, 767–776. [Google Scholar] [CrossRef]
- Galeano-Castaneda, Y.; Urrea-Aguirre, P.; Piedrahita, S.; Bascunan, P.; Correa, M.M. Composition and structure of the culturable gut bacterial communities in Anopheles albimanus from Colombia. PLoS ONE 2019, 14, e225833. [Google Scholar] [CrossRef] [PubMed]
- Pace, N.R. A molecular view of microbial diversity and the biosphere. Science 1997, 276, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Hugenholtz, P.; Goebel, B.M.; Pace, N.R. Impact of culture-independent studies on the emerging phylogenetic view of bacterial diversity. J. Bacteriol. 1998, 180, 4765–4774. [Google Scholar] [CrossRef]
- Anjum, S.I.; Shah, A.H.; Aurongzeb, M.; Kori, J.; Azim, M.K.; Ansari, M.J.; Bin, L. Characterization of gut bacterial flora of Apis mellifera from north-west Pakistan. Saudi J. Biol. Sci. 2018, 25, 388–392. [Google Scholar] [CrossRef]
- Sklarz, M.Y.; Angel, R.; Gillor, O.; Soares, M.I. Evaluating amplified rDNA restriction analysis assay for identification of bacterial communities. Antonie Leeuwenhoek 2009, 96, 659–664. [Google Scholar] [CrossRef]
- Tabar, E.A.; Staji, H.; Mahdavi, A. Comparative restriction enzyme mapping of Campylobacter jejuni isolates from turkeys and broilers based on flaA flagellar gene using HpyF3I endonuclease. Folia Microbiol. 2019, 64, 189–195. [Google Scholar] [CrossRef]
- He, H.; Wei, C.; Wheeler, D.E. The gut bacterial communities associated with lab-raised and field-collected ants of Camponotus fragilis (Formicidae: Formicinae). Curr. Microbiol. 2014, 69, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Dec, M.; Puchalski, A.; Urban-Chmiel, R.; Wernicki, A. 16S-ARDRA and MALDI-TOF mass spectrometry as tools for identification of Lactobacillus bacteria isolated from poultry. BMC Microbiol. 2016, 16, 105. [Google Scholar] [CrossRef] [PubMed]
- van Burgh, S.; Maghdid, D.M.; Ganjo, A.R.; Mansoor, I.Y.; Kok, D.J.; Fatah, M.H.; Alnakshabandi, A.A.; Asad, D.; Hammerum, A.M.; Ng, K.; et al. PME and other ESBL-positive multiresistant Pseudomonas aeruginosa isolated from hospitalized patients in the region of kurdistan, Iraq. Microb. Drug Resist. 2019, 25, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Lindstedt, B.A.; Heir, E.; Vardund, T.; Melby, K.K.; Kapperud, G. Comparative fingerprinting analysis of Campylobacter jejuni subsp. jejuni strains by amplified-fragment length polymorphism genotyping. J. Clin. Microbiol. 2000, 38, 3379–3387. [Google Scholar] [CrossRef] [PubMed]
- Al-Soud, W.A.; Bennedsen, M.; On, S.; Ouis, I.S.; Vandamme, P.; Nilsson, H.O.; Ljungh, A.; Wadstrom, T. Assessment of PCR-DGGE for the identification of diverse Helicobacter species, and application to faecal samples from zoo animals to determine Helicobacter prevalence. J. Med. Microbiol. 2003, 52, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.L.; Pan, Q.J.; Tian, H.G.; Douglas, A.E.; Liu, T.X. Bacteria abundance and diversity of different life stages of Plutella xylostella (Lepidoptera: Plutellidae), revealed by bacteria culture-dependent and PCR-DGGE methods. Insect Sci. 2015, 22, 375–385. [Google Scholar] [CrossRef]
- Snyman, M.; Gupta, A.K.; Bezuidenhout, C.C.; Claassens, S.; van den Berg, J. Gut microbiota of Busseola fusca (Lepidoptera: Noctuidae). World J. Microbiol. Biotechnol. 2016, 32, 115. [Google Scholar] [CrossRef]
- Yadav, K.K.; Bora, A.; Datta, S.; Chandel, K.; Gogoi, H.K.; Prasad, G.B.; Veer, V. Molecular characterization of midgut microbiota of Aedes albopictus and Aedes aegypti from Arunachal Pradesh, India. Parasites Vectors 2015, 8, 641. [Google Scholar] [CrossRef]
- Lv, Z.H.; Wei, X.Y.; Tao, Y.L.; Chu, D. Differential susceptibility of whitefly-associated bacteria to antibiotic as revealed by metagenomics analysis. Infect. Genet. Evol. 2018, 63, 24–29. [Google Scholar] [CrossRef]
- Chiel, E.; Gottlieb, Y.; Zchori-Fein, E.; Mozes-Daube, N.; Katzir, N.; Inbar, M.; Ghanim, M. Biotype-dependent secondary symbiont communities in sympatric populations of Bemisia tabaci. Bull. Entomol. Res. 2007, 97, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.; Gao, C.S.; De Barro, P.; Zhang, Y.J.; Wan, F.H.; Khan, I.A. Further insights into the strange role of bacterial endosymbionts in whitefly, Bemisia tabaci: Comparison of secondary symbionts from biotypes B and Q in China. Bull. Entomol. Res. 2011, 101, 477–486. [Google Scholar] [CrossRef]
- Su, M.M.; Guo, L.; Tao, Y.L.; Zhang, Y.J.; Wan, F.H.; Chu, D. Effects of host plant factors on the bacterial communities associated with two whitefly sibling species. PLoS ONE 2016, 11, e152183. [Google Scholar] [CrossRef]
- Tong, J.; Liu, C.; Summanen, P.; Xu, H.; Finegold, S.M. Application of quantitative real-time PCR for rapid identification of Bacteroides fragilis group and related organisms in human wound samples. Anaerobe 2011, 17, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Sedgley, C.M.; Nagel, A.C.; Shelburne, C.E.; Clewell, D.B.; Appelbe, O.; Molander, A. Quantitative real-time PCR detection of oral Enterococcus faecalis in humans. Arch. Oral Biol. 2005, 50, 575–583. [Google Scholar] [CrossRef]
- Telmadarrehei, T.; Tang, J.D.; Raji, O.; Rezazadeh, A.; Narayanan, L.; Shmulsky, R.; Jeremic, D. A Study of the gut bacterial community of Reticulitermes virginicus exposed to chitosan treatment. Insects 2020, 11, 681. [Google Scholar] [CrossRef] [PubMed]
- Bapatla, K.G.; Singh, A.; Yeddula, S.; Patil, R.H. Annotation of gut bacterial taxonomic and functional diversity in Spodoptera litura and Spilosoma obliqua. J. Basic Microbiol. 2018, 58, 217–226. [Google Scholar] [CrossRef]
- Xia, X.; Lan, B.; Tao, X.; Lin, J.; You, M. Characterization of Spodoptera litura Gut Bacteria and Their Role in Feeding and Growth of the Host. Front. Microbiol. 2020, 11, 1492. [Google Scholar] [CrossRef]
- Gao, X.; Li, W.; Luo, J.; Zhang, L.; Ji, J.; Zhu, X.; Wang, L.; Zhang, K.; Zhang, S.; Cui, J. DNA sequencing reveals bacterial communities in midgut and other parts of the larvae of Spodoptera exigua Hubner (Lepidoptera: Noctuidae). FEMS Microbiol. Lett. 2020, 367, fnaa002. [Google Scholar] [CrossRef]
- Chen, B.; Teh, B.S.; Sun, C.; Hu, S.; Lu, X.; Boland, W.; Shao, Y. Biodiversity and activity of the gut microbiota across the life history of the insect herbivore Spodoptera littoralis. Sci. Rep. 2016, 6, 29505. [Google Scholar] [CrossRef]
- Shan, Y.; Shu, C.; Crickmore, N.; Liu, C.; Xiang, W.; Song, F.; Zhang, J. Cultivable gut bacteria of scarabs (Coleoptera: Scarabaeidae) inhibit Bacillus thuringiensis multiplication. Environ. Entomol. 2014, 43, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Broderick, N.A.; Raffa, K.F.; Handelsman, J. Midgut bacteria required for Bacillus thuringiensis insecticidal activity. Proc. Natl. Acad. Sci. USA 2006, 103, 15196–15199. [Google Scholar] [CrossRef] [PubMed]
- Paramasiva, I.; Sharma, H.C.; Krishnayya, P.V. Antibiotics influence the toxicity of the delta endotoxins of Bacillus thuringiensis towards the cotton bollworm, Helicoverpa armigera. BMC Microbiol. 2014, 14, 200. [Google Scholar] [CrossRef] [PubMed]
- Raymond, B.; Johnston, P.R.; Wright, D.J.; Ellis, R.J.; Crickmore, N.; Bonsall, M.B. A mid-gut microbiota is not required for the pathogenicity of Bacillus thuringiensis to diamondback moth larvae. Environ. Microbiol. 2009, 11, 2556–2563. [Google Scholar] [CrossRef]
- Ren, X.L.; Chen, R.R.; Zhang, Y.; Ma, Y.; Cui, J.J.; Han, Z.J.; Mu, L.L.; Li, G.Q. A Spodoptera exigua cadherin serves as a putative receptor for Bacillus thuringiensis Cry1Ca toxin and shows differential enhancement of Cry1Ca and Cry1Ac toxicity. Appl. Environ. Microbiol. 2013, 79, 5576–5583. [Google Scholar] [CrossRef]
- Lucena, W.A.; Pelegrini, P.B.; Martins-De-Sa, D.; Fonseca, F.C.; Gomes, J.J.; de Macedo, L.L.; Da, S.M.; Oliveira, R.S.; Grossi-De-Sa, M.F. Molecular approaches to improve the insecticidal activity of Bacillus thuringiensis Cry toxins. Toxins 2014, 6, 2393–2423. [Google Scholar] [CrossRef]
- Pigott, C.R.; Ellar, D.J. Role of receptors in Bacillus thuringiensis crystal toxin activity. Microbiol. Mol. Biol. Rev. 2007, 71, 255–281. [Google Scholar] [CrossRef]
- Broderick, N.A.; Raffa, K.F.; Handelsman, J. Chemical modulators of the innate immune response alter gypsy moth larval susceptibility to Bacillus thuringiensis. BMC Microbiol. 2010, 10, 129. [Google Scholar] [CrossRef]
- Caccia, S.; Di Lelio, I.; La Storia, A.; Marinelli, A.; Varricchio, P.; Franzetti, E.; Banyuls, N.; Tettamanti, G.; Casartelli, M.; Giordana, B.; et al. Midgut microbiota and host immunocompetence underlie Bacillus thuringiensis killing mechanism. Proc. Natl. Acad. Sci. USA 2016, 113, 9486–9491. [Google Scholar] [CrossRef]
- Mason, K.L.; Stepien, T.A.; Blum, J.E.; Holt, J.F.; Labbe, N.H.; Rush, J.S.; Raffa, K.F.; Handelsman, J. From commensal to pathogen: Translocation of Enterococcus faecalis from the midgut to the hemocoel of Manduca sexta. mBio 2011, 2, e11–e65. [Google Scholar] [CrossRef]
Figure 1.
The main function of gut microbiota in insects.
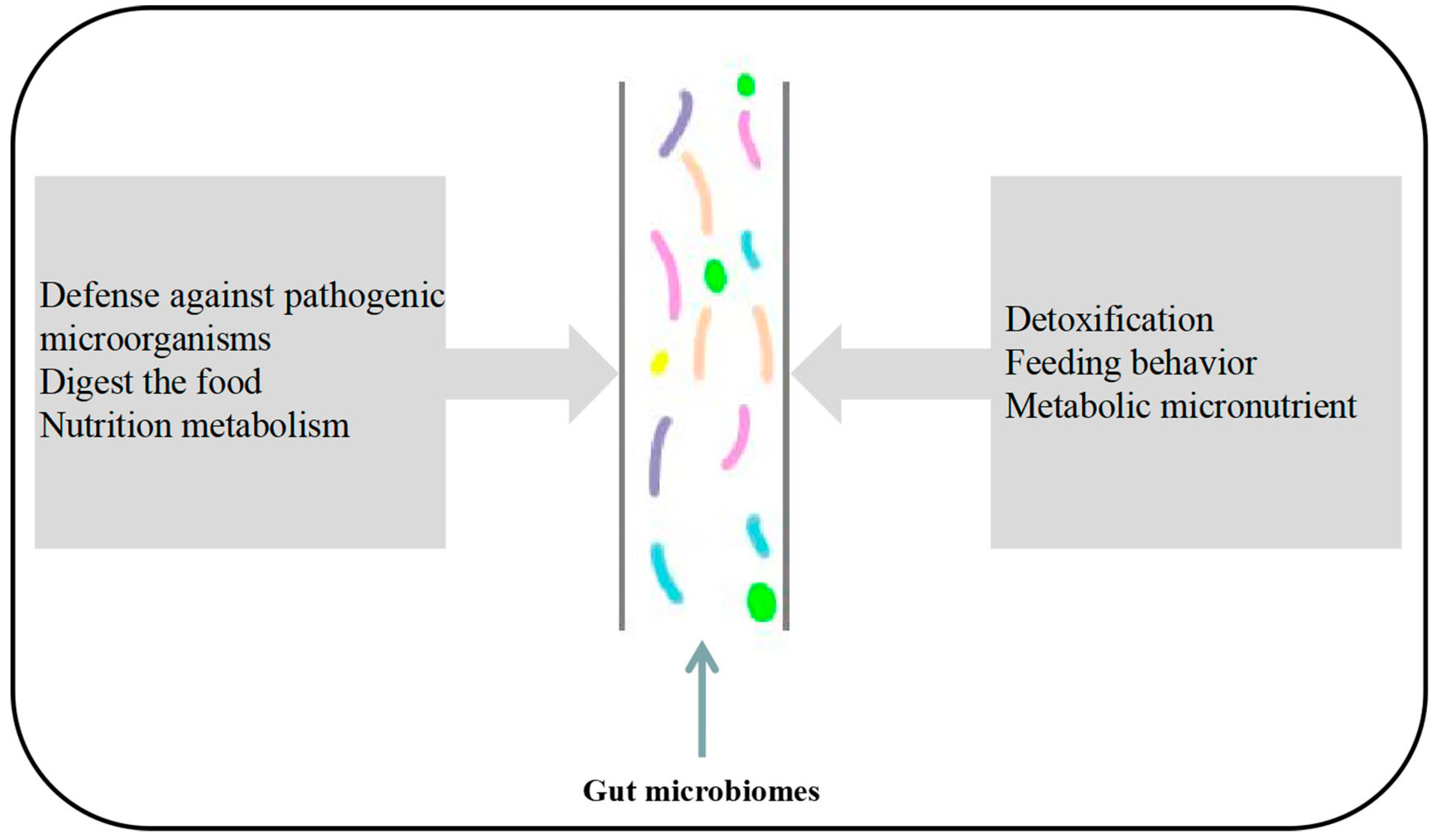
Table 1.
The role of insect gut microbes in nutrient metabolism.
| Insect | Bacteria Genera | Function | References |
|---|---|---|---|
| Bombyx mori | Pseudomonas, Klebsiella pneumoniae, Clostridium flexneri | It secretes cellulase, which breaks down carbohydrates to provide energy for the host. | [14] |
| Locusta migratoria manilensis | Klebsiella pneumoniae | It breaks down grass into carbohydrates, amino acids and sugars, providing energy to the host. | [15] |
| Aedes albopictus | Lelliottia, Cladosporium, Aspergillus, Ampullimonas, Cyberlindnera | It can digest food and provide nutrients for insects. | [16] |
| Herbivorous turtle ants Cephalotes | Burkholderiales, Opitutales | It can participate in nitrogen recycling to provide amino acids for the host. | [13] |
| Acyrthosiphon pisum | Buchnera aphidicola | It can provide the host with vitamins B2 and B5. | [6] |
| Honeybee | Gilliamella apicola and Lactobacillus sp. | It produces SCFAs that promote host growth. | [19] |
| Drosophila | Acetobacter pomorum, Lactobacillus plantarum, Saccharomyces cerevisiae, Lactobacillus plantarum, Acetobacter malorum | It can influence the feeding behavior of the host. | [23] |
| Rhynchophorus ferrugineus Olivier | Lactococcus lactis, Enterobacter cloacae | It can affect protein, glucose, and triglyceride levels in the host’s hemolymph. | [25] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, Y.; Chang, L.; Xu, K.; Zhang, S.; Gao, F.; Fan, Y. Research Progresses on the Function and Detection Methods of Insect Gut Microbes. Microorganisms 2023, 11, 1208. https://doi.org/10.3390/microorganisms11051208
AMA Style
Li Y, Chang L, Xu K, Zhang S, Gao F, Fan Y. Research Progresses on the Function and Detection Methods of Insect Gut Microbes. Microorganisms. 2023; 11(5):1208. https://doi.org/10.3390/microorganisms11051208
Chicago/Turabian StyleLi, Yazi, Liyun Chang, Ke Xu, Shuhong Zhang, Fengju Gao, and Yongshan Fan. 2023. "Research Progresses on the Function and Detection Methods of Insect Gut Microbes" Microorganisms 11, no. 5: 1208. https://doi.org/10.3390/microorganisms11051208
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.