
Current Progress and Future Perspectives on the Use of Bacillus clausii

 ,
,
Abstract
:1. Introduction
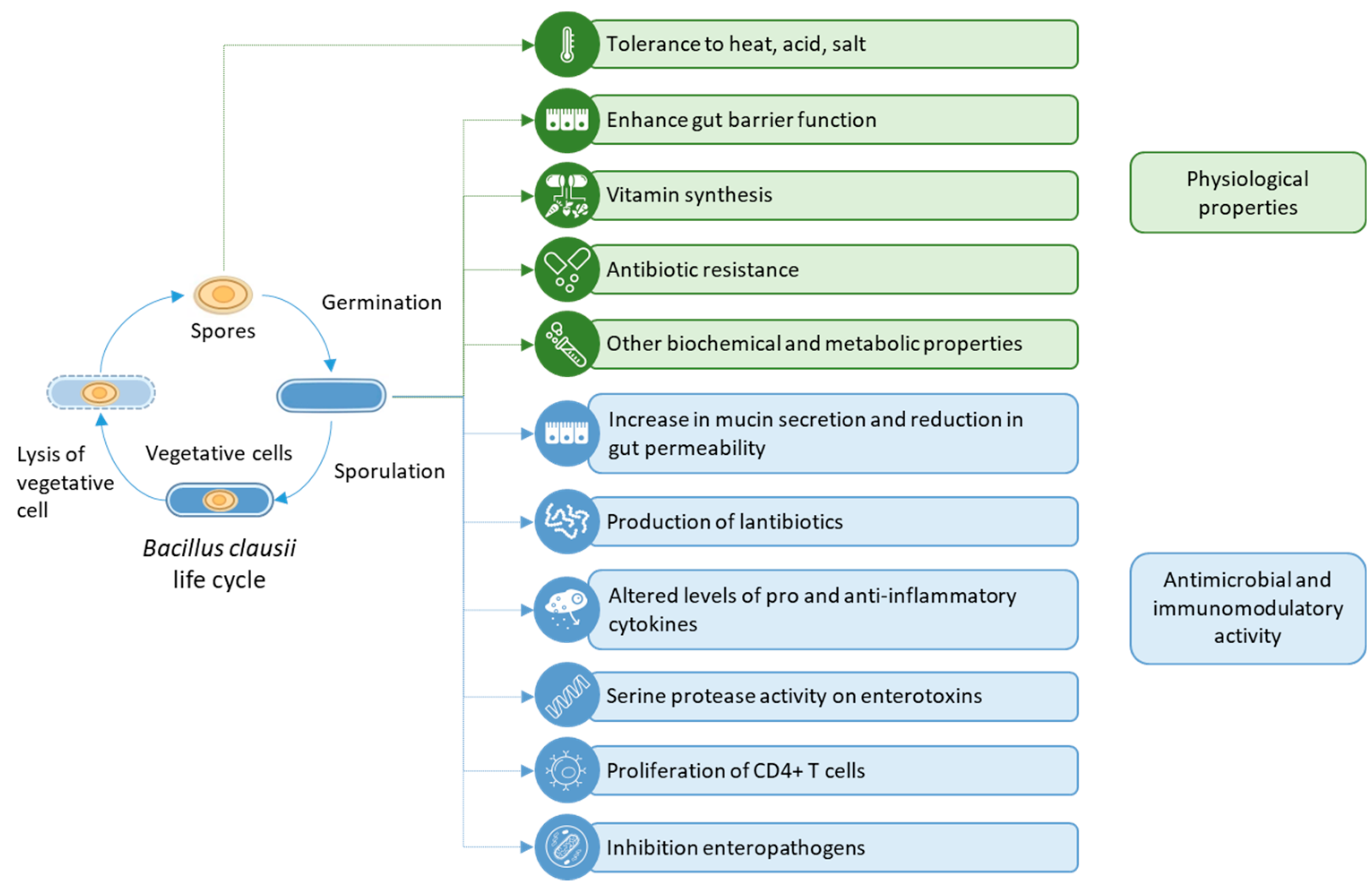
2. Physiological Properties of Bacillus clausii
2.1. Tolerance to Heat, Acid, and Salt
2.2. Vitamin Synthesis
2.3. Antibiotic Resistance

{kind=link}
| Inhibition Zone Diameter (mm) | ||||
|---|---|---|---|---|
| Antibiotic | B. clausii O/C | B. clausii SIN | B. clausii N/R | B. clausii T |
| Oxacillin | 8 | 0 | 0 | 9 ± 1.1 |
| Cefuroxime | 10 ± 0.7 | 0 | 0 | 12 ± 0.8 |
| Cefepime | 8 ± 1 | 0 | 0 | 11 ± 0.5 |
| Streptomycin | 28 ± 0.4 | 0 | 26 ± 0.6 | 30 ± 0.5 |
| Chloramphenicol | 0 | 16 ± 0.6 | 13 | 15 ± 0.6 |
| Rifampicin | 24 ± 0.5 | 26 ± 0.5 | 0 | 27 ± 0.6 |
| Metronidazole | 0 | 0 | 0 | 0 |
3. Preclinical Studies on the Probiotic Effects of B. clausii
3.1. Gut Immune Function
3.1.1. Enhancing Gut Barrier Function
3.1.2. Contributing to Gut Homoeostasis
3.2. Antimicrobial and Immunomodulatory Activity
3.2.1. Antimicrobial Activity
3.2.2. Immunomodulatory Activity
3.2.3. Inhibition of Enteropathogens
4. Clinical Studies on Probiotic Effects of B. clausii
5. Compositional Quality and Safety
6. Other Biochemical and Metabolic Properties
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bach, J.F. The effect of infections on susceptibility to autoimmune and allergic diseases. N. Engl. J. Med. 2002, 347, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Gagliardi, A.; Totino, V.; Cacciotti, F.; Iebba, V.; Neroni, B.; Bonfiglio, G.; Trancassini, M.; Passariello, C.; Pantanella, F.; Schippa, S. Rebuilding the Gut Microbiota Ecosystem. Int. J. Environ. Res. Public Health 2018, 15, 1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Wieers, G.; Belkhir, L.; Enaud, R.; Leclercq, S.; De Foy, P.J.M.; Dequenne, I.; de Timary, P.; Cani, P.D. How Probiotics Affect the Microbiota. Front. Cell. Infect. Microbiol. 2019, 9, 454. [Google Scholar] [CrossRef] [Green Version]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef]
- Navarro-Lopez, V.; Martinez-Andres, A.; Ramirez-Bosca, A.; Ruzafa-Costas, B.; Nunez-Delegido, E.; Carrion-Gutierrez, M.A.; Prieto-Merino, D.; Codoner-Cortes, F.; Ramon-Vidal, D.; Genoves-Martinez, S.; et al. Efficacy and Safety of Oral Administration of a Mixture of Probiotic Strains in Patients with Psoriasis: A Randomized Controlled Clinical Trial. Acta Derm. Venereol. 2019, 99, 1078–1084. [Google Scholar] [CrossRef] [Green Version]
- Michail, S.K.; Stolfi, A.; Johnson, T.; Onady, G.M. Efficacy of probiotics in the treatment of pediatric atopic dermatitis: A meta-analysis of randomized controlled trials. Ann. Allergy Asthma Immunol. 2008, 101, 508–516. [Google Scholar] [CrossRef]
- Kim, S.O.; Ah, Y.M.; Yu, Y.M.; Choi, K.H.; Shin, W.G.; Lee, J.Y. Effects of probiotics for the treatment of atopic dermatitis: A meta-analysis of randomized controlled trials. Ann. Allergy Asthma Immunol. 2014, 113, 217–226. [Google Scholar] [CrossRef]
- Leblhuber, F.; Steiner, K.; Schuetz, B.; Fuchs, D.; Gostner, J.M. Probiotic Supplementation in Patients with Alzheimer’s Dementia-An Explorative Intervention Study. Curr. Alzheimer Res. 2018, 15, 1106–1113. [Google Scholar] [CrossRef]
- Kouchaki, E.; Tamtaji, O.R.; Salami, M.; Bahmani, F.; Kakhaki, R.D.; Akbari, E.; Tajabadi-Ebrahimi, M.; Jafari, P.; Asemi, Z. Clinical and metabolic response to probiotic supplementation in patients with multiple sclerosis: A randomized, double-blind, placebo-controlled trial. Clin. Nutr. 2017, 36, 1245–1249. [Google Scholar] [CrossRef] [PubMed]
- Groeger, D.; O’Mahony, L.; Murphy, E.F.; Bourke, J.F.; Dinan, T.G.; Kiely, B.; Shanahan, F.; Quigley, E.M. Bifidobacterium infantis 35624 modulates host inflammatory processes beyond the gut. Gut Microbes 2013, 4, 325–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emre, I.E.; Eroglu, Y.; Kara, A.; Dinleyici, E.C.; Ozen, M. The effect of probiotics on prevention of upper respiratory tract infections in the paediatric community-A systematic review. Benef. Microbes 2020, 11, 201–211. [Google Scholar] [CrossRef] [PubMed]
- McFarland, L.V.; Ozen, M.; Dinleyici, E.C.; Goh, S. Comparison of pediatric and adult antibiotic-associated diarrhea and Clostridium difficile infections. World J. Gastroenterol. 2016, 22, 3078–3104. [Google Scholar] [CrossRef] [PubMed]
- Szajewska, H.; Horvath, A.; Kolodziej, M. Systematic review with meta-analysis: Saccharomyces boulardii supplementation and eradication of Helicobacter pylori infection. Aliment. Pharmacol. Ther. 2015, 41, 1237–1245. [Google Scholar] [CrossRef]
- Miller, L.E.; Lehtoranta, L.; Lehtinen, M.J. Short-term probiotic supplementation enhances cellular immune function in healthy elderly: Systematic review and meta-analysis of controlled studies. Nutr. Res. 2019, 64, 1–8. [Google Scholar] [CrossRef]
- Zimmermann, P.; Curtis, N. The influence of probiotics on vaccine responses—A systematic review. Vaccine 2018, 36, 207–213. [Google Scholar] [CrossRef]
- Lei, W.T.; Shih, P.C.; Liu, S.J.; Lin, C.Y.; Yeh, T.L. Effect of Probiotics and Prebiotics on Immune Response to Influenza Vaccination in Adults: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2017, 9, 1175. [Google Scholar] [CrossRef]
- Elshaghabee, F.M.F.; Rokana, N.; Gulhane, R.D.; Sharma, C.; Panwar, H. Bacillus As Potential Probiotics: Status, Concerns, and Future Perspectives. Front. Microbiol. 2017, 8, 1490. [Google Scholar] [CrossRef] [Green Version]
- Cutting, S.M.; Ricca, E. Bacterial spore-formers: Friends and foes. FEMS Microbiol. Lett. 2014, 358, 107–109. [Google Scholar] [CrossRef] [Green Version]
- Hong, H.A.; Duc, L.H.; Cutting, S.M. The use of bacterial spore formers as probiotics. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, M.E.; Morelli, L.; Tompkins, T.A. Sporeformers as Human Probiotics: Bacillus, Sporolactobacillus, and Brevibacillus. Compr. Rev. Food Sci. Food Saf. 2003, 2, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Hoyles, L.; Honda, H.; Logan, N.A.; Halket, G.; La Ragione, R.M.; McCartney, A.L. Recognition of greater diversity of Bacillus species and related bacteria in human faeces. Res. Microbiol. 2012, 163, 3–13. [Google Scholar] [CrossRef]
- Lopetuso, L.R.; Scaldaferri, F.; Franceschi, F.; Gasbarrini, A. Bacillus clausii and gut homeostasis: State of the art and future perspectives. Expert Rev. Gastroenterol. Hepatol. 2016, 10, 943–948. [Google Scholar] [CrossRef] [PubMed]
- Senesi, S.; Celandroni, F.; Tavanti, A.; Ghelardi, E. Molecular characterization and identification of Bacillus clausii Strains marketed for use in oral bacteriotherapy. Appl. Environ. Microbiol. 2001, 67, 834–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celandroni, F.; Vecchione, A.; Cara, A.; Mazzantini, D.; Lupetti, A.; Ghelardi, E. Identification of Bacillus species: Implication on the quality of probiotic formulations. PLoS ONE 2019, 14, e0217021. [Google Scholar] [CrossRef] [PubMed]
- Plomer, M.; Perez, M.I.; Greifenberg, D.M. Effect of Bacillus clausii Capsules in Reducing Adverse Effects Associated with Helicobacter pylori Eradication Therapy: A Randomized, Double-Blind, Controlled Trial. Infect. Dis. Ther. 2020, 9, 867–878. [Google Scholar] [CrossRef]
- Nista, E.C.; Candelli, M.; Cremonini, F.; Cazzato, I.A.; Zocco, M.A.; Franceschi, F.; Cammarota, G.; Gasbarrini, G.; Gasbarrini, A. Bacillus clausii therapy to reduce side-effects of anti-Helicobacter pylori treatment: Randomized, double-blind, placebo controlled trial. Aliment. Pharmacol. Ther. 2004, 20, 1181–1188. [Google Scholar] [CrossRef]
- Abbrescia, A.P.; Palese, L.L.; Papa, S.; Gaballo, A.; Alifano, P.; Sardanelli, A.M. Antibiotic Sensitivity of Bacillus clausii Strains in Commercial Preparation. Clin. Immunol. Endocr. Metab. Drugs 2014, 1, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Ghelardi, E.; Celandroni, F.; Salvetti, S.; Gueye, S.A.; Lupetti, A.; Senesi, S. Survival and persistence of Bacillus clausii in the human gastrointestinal tract following oral administration as spore-based probiotic formulation. J. Appl. Microbiol. 2015, 119, 552–559. [Google Scholar] [CrossRef]
- Ciffo, F. Determination of the spectrum of antibiotic resistance of the “Bacillus subtilis” strains of Enterogermina. Chemioterapia 1984, 3, 45–52. [Google Scholar] [PubMed]
- dos Santos, D.F.A.; dos Santos, S.S.F. Bacillus clausii: Review of characteristics and applications in medicine, biotechnology and the food industry. Rev. Biocienc. 2019, 25, 29–38. [Google Scholar]
- Kolacek, S.; Hojsak, I.; Berni Canani, R.; Guarino, A.; Indrio, F.; Orel, R.; Pot, B.; Shamir, R.; Szajewska, H.; Vandenplas, Y.; et al. Commercial Probiotic Products: A Call for Improved Quality Control. A Position Paper by the ESPGHAN Working Group for Probiotics and Prebiotics. J. Pediatr. Gastroenterol. Nutr. 2017, 65, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Vecchione, A.; Celandroni, F.; Mazzantini, D.; Senesi, S.; Lupetti, A.; Ghelardi, E. Compositional Quality and Potential Gastrointestinal Behavior of Probiotic Products Commercialized in Italy. Front. Med. 2018, 5, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvetti, S.; Celandroni, F.; Ghelardi, E.; Baggiani, A.; Senesi, S. Rapid determination of vitamin B2 secretion by bacteria growing on solid media. J. Appl. Microbiol. 2003, 95, 1255–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.X.; Xu, Q.M.; Yu, S.C.; Cheng, J.S.; Yuan, Y.J. Bio-removal of tetracycline antibiotics under the consortium with probiotics Bacillus clausii T and Bacillus amyloliquefaciens producing biosurfactants. Sci. Total Environ. 2020, 710, 136329. [Google Scholar] [CrossRef]
- Kong, X.X.; Jiang, J.L.; Qiao, B.; Liu, H.; Cheng, J.S.; Yuan, Y.J. The biodegradation of cefuroxime, cefotaxime and cefpirome by the synthetic consortium with probiotic Bacillus clausii and investigation of their potential biodegradation pathways. Sci. Total Environ. 2019, 651, 271–280. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations; World Health Organization. Guidelines for the Evaluation of Probiotics in Food; Food and Agriculture Organization of the United Nations: Rome, Italy; World Health Organization: Geneva, Switzerland, 2002.
- Girlich, D.; Leclercq, R.; Naas, T.; Nordmann, P. Molecular and biochemical characterization of the chromosome-encoded class A β-lactamase BCL-1 from Bacillus clausii. Antimicrob. Agents Chemother. 2007, 51, 4009–4014. [Google Scholar] [CrossRef] [Green Version]
- Bozdogan, B.; Galopin, S.; Leclercq, R. Characterization of a new erm-related macrolide resistance gene present in probiotic strains of Bacillus clausii. Appl. Environ. Microbiol. 2004, 70, 280–284. [Google Scholar] [CrossRef] [Green Version]
- Bozdogan, B.; Galopin, S.; Gerbaud, G.; Courvalin, P.; Leclercq, R. Chromosomal aadD2 encodes an aminoglycoside nucleotidyltransferase in Bacillus clausii. Antimicrob. Agents Chemother. 2003, 47, 1343–1346. [Google Scholar] [CrossRef] [Green Version]
- Galopin, S.; Cattoir, V.; Leclercq, R. A chromosomal chloramphenicol acetyltransferase determinant from a probiotic strain of Bacillus clausii. FEMS Microbiol. Lett. 2009, 296, 185–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatri, I.; Sharma, G.; Subramanian, S. Composite genome sequence of Bacillus clausii, a probiotic commercially available as Enterogermina®, and insights into its probiotic properties. BMC Microbiol. 2019, 19, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapse, N.G.; Engineer, A.S.; Gowdaman, V.; Wagh, S.; Dhakephalkar, P.K. Genome Profiling for Health Promoting and Disease Preventing Traits Unraveled Probiotic Potential of Bacillus clausii B106. Microbiol. Biotechnol. Lett. 2018, 46, 334–345. [Google Scholar] [CrossRef]
- Upadrasta, A.; Pitta, S.; Madempudi, R.S. Draft Genome Sequence of Bacillus clausii UBBC07, a Spore-Forming Probiotic Strain. Genome Announc. 2016, 4, e00235-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higuchi, Y.; Mori, K.; Suyama, A.; Huang, Y.; Tashiro, K.; Kuhara, S.; Takegawa, K. Draft Genome Sequence of Bacillus clausii AKU0647, a Strain That Produces Endo-β-N-Acetylglucosaminidase A. Genome Announc. 2016, 4, e00310-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahire, J.J.; Kashikar, M.S.; Madempudi, R.S. Survival and Germination of Bacillus clausii UBBC07 Spores in in vitro Human Gastrointestinal Tract Simulation Model and Evaluation of Clausin Production. Front. Microbiol. 2020, 11, 1010. [Google Scholar] [CrossRef] [PubMed]
- Paparo, L.; Tripodi, L.; Bruno, C.; Pisapia, L.; Damiano, C.; Pastore, L.; Canani, R.B. Protective action of Bacillus clausii probiotic strains in an in vitro model of Rotavirus infection. Sci. Rep. 2020, 10, 12636. [Google Scholar] [CrossRef]
- Di Caro, S.; Tao, H.; Grillo, A.; Franceschi, F.; Elia, C.; Zocco, M.A.; Gasbarrini, G.; Sepulveda, A.R.; Gasbarrini, A. Bacillus clausii effect on gene expression pattern in small bowel mucosa using DNA microarray analysis. Eur. J. Gastroenterol. Hepatol. 2005, 17, 951–960. [Google Scholar] [CrossRef]
- Riquelme, E.; Zhang, Y.; Zhang, L.; Montiel, M.; Zoltan, M.; Dong, W.; Quesada, P.; Sahin, I.; Chandra, V.; Lucas, A.S.; et al. Tumor Microbiome Diversity and Composition Influence Pancreatic Cancer Outcomes. Cell 2019, 178, 795–806.e12. [Google Scholar] [CrossRef]
- Duysburgh, C.; Van den Abbeele, P.; Krishnan, K.; Bayne, T.F.; Marzorati, M. A synbiotic concept containing spore-forming Bacillus strains and a prebiotic fiber blend consistently enhanced metabolic activity by modulation of the gut microbiome in vitro. Int. J. Pharm. X 2019, 1, 100021. [Google Scholar] [CrossRef]
- Patel, C.; Patel, P.; Acharya, S. Therapeutic Prospective of a Spore-Forming Probiotic-Bacillus clausii UBBC07 Against Acetaminophen-Induced Uremia in Rats. Probiotics Antimicrob. Proteins 2020, 12, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Rochin-Medina, J.J.; Ramirez-Medina, H.K.; Rangel-Peraza, J.G.; Pineda-Hidalgo, K.V.; Iribe-Arellano, P. Use of whey as a culture medium for Bacillus clausii for the production of protein hydrolysates with antimicrobial and antioxidant activity. Food Sci. Technol. Int. 2018, 24, 35–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouhss, A.; Al-Dabbagh, B.; Vincent, M.; Odaert, B.; Aumont-Nicaise, M.; Bressolier, P.; Desmadril, M.; Mengin-Lecreulx, D.; Urdaci, M.C.; Gallay, J. Specific interactions of clausin, a new lantibiotic, with lipid precursors of the bacterial cell wall. Biophys. J. 2009, 97, 1390–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.; Jung, A.-Y.; Chang, C.-S.; Kim, Y.H. Bacillus clausii, a Foreshore-Derived Probiotic, Attenuates Allergic Airway Inflammation through Downregulation of Hypoxia Signaling. J. Rhinol. 2020, 27, 108–116. [Google Scholar] [CrossRef]
- Pradhan, B.; Guha, D.; Ray, P.; Das, D.; Aich, P. Comparative Analysis of the Effects of Two Probiotic Bacterial Strains on Metabolism and Innate Immunity in the RAW 264.7 Murine Macrophage Cell Line. Probiotics Antimicrob. Proteins 2016, 8, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, B.; Guha, D.; Naik, A.K.; Banerjee, A.; Tambat, S.; Chawla, S.; Senapati, S.; Aich, P. Probiotics L. acidophilus and B. clausii Modulate Gut Microbiota in Th1- and Th2-Biased Mice to Ameliorate Salmonella Typhimurium-Induced Diarrhea. Probiotics Antimicrob. Proteins 2019, 11, 887–904. [Google Scholar] [CrossRef]
- Ripert, G.; Racedo, S.M.; Elie, A.M.; Jacquot, C.; Bressollier, P.; Urdaci, M.C. Secreted Compounds of the Probiotic Bacillus clausii Strain O/C Inhibit the Cytotoxic Effects Induced by Clostridium difficile and Bacillus cereus Toxins. Antimicrob. Agents Chemother. 2016, 60, 3445–3454. [Google Scholar] [CrossRef] [Green Version]
- Urdaci, M.C.; Bressollier, P.; Pinchuk, I. Bacillus clausii probiotic strains: Antimicrobial and immunomodulatory activities. J. Clin. Gastroenterol. 2004, 38, S86–S90. [Google Scholar] [CrossRef] [Green Version]
- Villeger, R.; Saad, N.; Grenier, K.; Falourd, X.; Foucat, L.; Urdaci, M.C.; Bressollier, P.; Ouk, T.S. Characterization of lipoteichoic acid structures from three probiotic Bacillus strains: Involvement of D-alanine in their biological activity. Antonie Van Leeuwenhoek 2014, 106, 693–706. [Google Scholar] [CrossRef] [Green Version]
- Scaldaferri, F.G.C.; Mora, V.; Petito, V.; Lopetuso, L.R.; Puca, P.; Ianiro, G.; Napolitano, D.; Quaranta, G.; Masucci, L.; Sanguinetti, M.; et al. Bacillus Clausii (O/C, SIN, N/R, T) Improves Acute Mild Colitis in Mice while in vivo Modulating Gut Microbiota. Ann. Gastroenterol. Dig. Syst. 2018, 4, 1035. [Google Scholar]
- Cruz, C.S. Estudo do Potencial Imunomodulatório do Bacillus Clausii (O/C, N/R, T e SIN) Sobre a Esquistossomose Mansoni Experimental; Universidade Federal de Pernambuco: Recife, Brazil, 2020. [Google Scholar]
- Yu, M.G.T.R.; Tuano, D.F.; Tud, R.M.; Umali, A.; Umandap, C.H.; Ver, M.L.; Villalobos, R.E.; Villanueva, A.P.; Villarante, K.L.; Villasenor, L.; et al. Histomorphologic Effects of Bacillus clausii spores in Enteropathogenic E. coli O127:H21-infected Mice: A Pilot Study. Philipp. J. Intern. Med. 2016, 54, wpr-633362. [Google Scholar]
- Montassier, E.; Gastinne, T.; Vangay, P.; Al-Ghalith, G.A.; Bruley des Varannes, S.; Massart, S.; Moreau, P.; Potel, G.; de La Cochetiere, M.F.; Batard, E.; et al. Chemotherapy-driven dysbiosis in the intestinal microbiome. Aliment. Pharmacol. Ther. 2015, 42, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Miranda Alatriste, P.V.; Urbina Arronte, R.; Gomez Espinosa, C.O.; Espinosa Cuevas Mde, L. Effect of probiotics on human blood urea levels in patients with chronic renal failure. Nutr. Hosp. 2014, 29, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Kuipers, O.P. Identification and classification of known and putative antimicrobial compounds produced by a wide variety of Bacillales species. BMC Genom. 2016, 17, 882. [Google Scholar] [CrossRef] [Green Version]
- Abriouel, H.; Franz, C.M.; Ben Omar, N.; Galvez, A. Diversity and applications of Bacillus bacteriocins. FEMS Microbiol. Rev. 2011, 35, 201–232. [Google Scholar] [CrossRef] [Green Version]
- Rochin-Medina, J.J.; Ramirez, K.; Rangel-Peraza, J.G.; Bustos-Terrones, Y.A. Increase of content and bioactivity of total phenolic compounds from spent coffee grounds through solid state fermentation by Bacillus clausii. J. Food Sci. Technol. 2018, 55, 915–923. [Google Scholar] [CrossRef]
- Bressollier, P.; Brugo, M.A.; Robineau, P.; Schmitter, J.M.; Sofeir, M.; Urdaci, M.C.; Verneuil, B. Peptide Compound with Biological Activity, Its Preparation and Its Application. U.S. Patent No. 8,691,773, 6 April 2007. [Google Scholar]
- Holgate, S.T. Pathophysiology of asthma: What has our current understanding taught us about new therapeutic approaches? J. Allergy Clin. Immunol. 2011, 128, 495–505. [Google Scholar] [CrossRef]
- Saglani, S.; Lloyd, C.M. Novel concepts in airway inflammation and remodelling in asthma. Eur. Respir. J. 2015, 46, 1796–1804. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, H.; Zhao, J. Macrophage Polarization Induced by Probiotic Bacteria: A Concise Review. Probiotics Antimicrob. Proteins 2019, 12, 798–808. [Google Scholar] [CrossRef]
- Boros, É.; Prontvai, B.; Kellermayer, Z.; Balogh, P.; Sarlós, P.; Vincze, Á.; Varga, C.; Maróti, Z.; Bálint, B.; Nagy, I. Transcriptome Based Profiling of the Immune Cell Gene Signature in Rat Experimental Colitis and Human IBD Tissue Samples. Biomolecules 2020, 10, 974. [Google Scholar] [CrossRef]
- Pavlidis, S.; Monast, C.; Loza, M.J.; Branigan, P.; Chung, K.F.; Adcock, I.M.; Guo, Y.; Rowe, A.; Baribaud, F. I_MDS: An inflammatory bowel disease molecular activity score to classify patients with differing disease-driving pathways and therapeutic response to anti-TNF treatment. PLoS Comput. Biol. 2019, 15, e1006951. [Google Scholar] [CrossRef] [PubMed]
- CDC. About Schistosomiasis. Available online: https://www.cdc.gov/parasites/schistosomiasis/gen_info/faqs.html (accessed on 19 October 2021).
- de Castro, J.A.; Guno, M.J.V.; Perez, M.O. Bacillus clausii as adjunctive treatment for acute community-acquired diarrhea among Filipino children: A large-scale, multicenter, open-label study (CODDLE). Trop. Dis. Travel Med. Vaccines 2019, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Ianiro, G.; Rizzatti, G.; Plomer, M.; Lopetuso, L.; Scaldaferri, F.; Franceschi, F.; Cammarota, G.; Gasbarrini, A. Bacillus clausii for the Treatment of Acute Diarrhea in Children: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2018, 10, 1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tewari, V.V.; Dubey, S.K.; Gupta, G. Bacillus clausii for Prevention of Late-onset Sepsis in Preterm Infants: A Randomized Controlled Trial. J. Trop. Pediatr. 2015, 61, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Marseglia, G.L.; Tosca, M.; Cirillo, I.; Licari, A.; Leone, M.; Marseglia, A.; Castellazzi, A.M.; Ciprandi, G. Efficacy of Bacillus clausii spores in the prevention of recurrent respiratory infections in children: A pilot study. Ther. Clin. Risk Manag. 2007, 3, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Ciprandi, G.; Tosca, M.A.; Milanese, M.; Caligo, G.; Ricca, V. Cytokines evaluation in nasal lavage of allergic children after Bacillus clausii administration: A pilot study. Pediatr. Allergy Immunol. 2004, 15, 148–151. [Google Scholar] [CrossRef]
- Ciprandi, G.; Vizzaccaro, A.; Cirillo, I.; Tosca, M.A. Bacillus clausii effects in children with allergic rhinitis. Allergy 2005, 60, 702–703. [Google Scholar] [CrossRef]
- Sudha, M.R.; Jayanthi, N.; Pandey, D.C.; Verma, A.K. Bacillus clausii UBBC-07 reduces severity of diarrhoea in children under 5 years of age: A double blind placebo controlled study. Benef. Microbes 2019, 10, 149–154. [Google Scholar] [CrossRef]
- Smiyan, O.I.; Smiian-Horbunova, K.O.; Bynda, T.P.; Loboda, A.M.; Popov, S.V.; Vysotsky, I.Y.; Moshchych, O.P.; Vasylieva, O.G.; Manko, Y.A.; Ovsianko, O.L.; et al. Optimization of the Treatment of Rotavirus Infection in Children by Using Bacillus Clausii. Wiadomości Lek. 2019, 72, 1320–1323. [Google Scholar] [CrossRef]
- Smiian, K.S.O.; Bynda, T.; Loboda, A. Bacillus clausii in treatment of rotavirus infection in children. In Proceedings of the International Scientific Conference on Medicine, Riga, Latvia, 20 March 2020; p. 225. [Google Scholar]
- McFarlin, B.K.; Henning, A.L.; Bowman, E.M.; Gary, M.A.; Carbajal, K.M. Oral spore-based probiotic supplementation was associated with reduced incidence of post-prandial dietary endotoxin, triglycerides, and disease risk biomarkers. World J. Gastrointest. Pathophysiol. 2017, 8, 117–126. [Google Scholar] [CrossRef]
- Nirmala, M.; Smitha, S.G.; Kamath, G.J. A Study to Assess The Efficacy of Local Application of Oral Probiotic in Treating Recurrent Aphthous Ulcer and Oral Candidiasis. Indian J. Otolaryngol. Head Neck Surg. 2019, 71, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Navarra, P.; Milleri, S.; Perez, I.M.; Uboldi, M.C.; Pellegrino, P.; De Fer, B.B.; Morelli, L. Kinetics of Intestinal Presence of Spores Following Oral Administration of Bacillus clausii Formulations: Three Single-Centre, Crossover, Randomised, Open-Label Studies. Eur. J. Drug Metab. Pharmacokinet. 2021, 46, 375–384. [Google Scholar] [CrossRef] [PubMed]
- De Castro, J.A.; Kesavelu, D.; Lahiri, K.R.; Chaijitraruch, N.; Chongsrisawat, V.; Jog, P.P.; Liaw, Y.H.; Nguyen, G.K.; Nguyen, T.V.H.; Pai, U.A.; et al. Recommendations for the adjuvant use of the poly-antibiotic-resistant probiotic Bacillus clausii (O/C, SIN, N/R, T) in acute, chronic, and antibiotic-associated diarrhea in children: Consensus from Asian experts. Trop. Dis. Travel Med. Vaccines 2020, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Sudha, M.R.; Bhonagiri, S.; Kumar, M.A. Efficacy of Bacillus clausii strain UBBC-07 in the treatment of patients suffering from acute diarrhoea. Benef. Microbes 2013, 4, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Soman, R.J.; Swamy, M.V. A prospective, randomized, double-blind, placebo-controlled, parallel-group study to evaluate the efficacy and safety of SNZ TriBac, a three-strain Bacillus probiotic blend for undiagnosed gastrointestinal discomfort. Int. J. Color. Dis. 2019, 34, 1971–1978. [Google Scholar] [CrossRef]
- Hungin, A.P.; Mulligan, C.; Pot, B.; Whorwell, P.; Agreus, L.; Fracasso, P.; Lionis, C.; Mendive, J.; de Foy, J.M.P.; Rubin, G.; et al. Systematic review: Probiotics in the management of lower gastrointestinal symptoms in clinical practice-an evidence-based international guide. Aliment. Pharmacol. Ther. 2013, 38, 864–886. [Google Scholar] [CrossRef] [Green Version]
- Catinean, A.; Neag, A.M.; Nita, A.; Buzea, M.; Buzoianu, A.D. Bacillus spp. Spores-A Promising Treatment Option for Patients with Irritable Bowel Syndrome. Nutrients 2019, 11, 1968. [Google Scholar] [CrossRef] [Green Version]
- Lahiri, K.J.K.; Gahlowt, P.; Najmuddin, F. Bacillus clausii as an adjuvant therapy in acute childhood diarrhea. IOSR J. Dent. Med. Sci. 2015, 14, 74–76. [Google Scholar] [CrossRef]
- Bueno, C.P.A.R.F.R.V.; Sanchez, A.C.; Reyes, M.L.U. Effectiveness of Bacillus clausii as adjutant treatment for paediatric irritable bowel syndrome (IBS). In Proceedings of the 6th World Congress of PGHAN: Abstracts, Vienna, Austria, 2–5 June 2021; pp. 1–1313. [Google Scholar]
- Guarner, F.; Khan, A.G.; Garisch, J.; Eliakim, R.; Gangl, A.; Thomson, A.; Krabshuis, J.; Lemair, T.; Kaufmann, P.; De Paula, J.A.; et al. World gastroenterology organisation global guidelines: Probiotics and prebiotics october 2011. J. Clin. Gastroenterol. 2012, 46, 468–481. [Google Scholar] [CrossRef]
- Ciprandi, G.; Vizzaccaro, A.; Cirillo, I.; Tosca, M.A. Bacillus clausii exerts immuno-modulatory activity in allergic subjects: A pilot study. Eur. Ann. Allergy Clin. Immunol. 2005, 37, 129–134. [Google Scholar]
- Cheng, B.; Zeng, X.; Liu, S.; Zou, J.; Wang, Y. The efficacy of probiotics in management of recurrent aphthous stomatitis: A systematic review and meta-analysis. Sci. Rep. 2020, 10, 21181. [Google Scholar] [CrossRef] [PubMed]
- Mundula, T.; Ricci, F.; Barbetta, B.; Baccini, M.; Amedei, A. Effect of Probiotics on Oral Candidiasis: A Systematic Review and Meta-Analysis. Nutrients 2019, 11, 2449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzantini, D.; Calvigioni, M.; Celandroni, F.; Lupetti, A.; Ghelardi, E. Spotlight on the Compositional Quality of Probiotic Formulations Marketed Worldwide. Front. Microbiol. 2021, 12, 693973. [Google Scholar] [CrossRef] [PubMed]
- Patrone, V.; Molinari, P.; Morelli, L. Microbiological and molecular characterization of commercially available probiotics containing Bacillus clausii from India and Pakistan. Int. J. Food Microbiol. 2016, 237, 92–97. [Google Scholar] [CrossRef]
- Morelli, L.; Pellegrino, P. A critical evaluation of the factors affecting the survival and persistence of beneficial bacteria in healthy adults. Benef. Microbes 2021, 12, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Princess, I.; Natarajan, T.; Ghosh, S. When good bacteria behave badly: A case report of Bacillus clausii sepsis in an immunocompetant adult. Access Microbiol. 2020, 2, e000097. [Google Scholar] [CrossRef]
- Joshi, S.; Udani, S.; Sen, S.; Kirolikar, S.; Shetty, A. Bacillus Clausii Septicemia in a Pediatric Patient After Treatment With Probiotics. Pediatr. Infect. Dis. J. 2019, 38, e228–e230. [Google Scholar] [CrossRef]
- Lakshmi, S.G.; Jayanthi, N.; Saravanan, M.; Ratna, M.S. Safety assesment of Bacillus clausii UBBC07, a spore forming probiotic. Toxicol. Rep. 2017, 4, 62–71. [Google Scholar] [CrossRef]
- European Food Safety Authority. Qualified Presumption of Safety (QPS). Available online: https://www.efsa.europa.eu/en/topics/topic/qualified-presumption-safety-qps (accessed on 6 May 2022).
- United States Food and Drug Administration. GRN No. 971. Bacillus clausii Strain 088AE Spore Preparation. Available online: https://www.cfsanappsexternal.fda.gov/scripts/fdcc/?set=GRASNotices&id=971&sort=GRN_No&order=DESC&startrow=1&type=basic&search=clausii (accessed on 23 May 2022).
- Brander, S.; Mikkelsen, J.D.; Kepp, K.P. Characterization of an alkali- and halide-resistant laccase expressed in E. coli: CotA from Bacillus clausii. PLoS ONE 2014, 9, e99402. [Google Scholar] [CrossRef] [Green Version]
- Ihssen, J.; Reiss, R.; Luchsinger, R.; Thony-Meyer, L.; Richter, M. Biochemical properties and yields of diverse bacterial laccase-like multicopper oxidases expressed in Escherichia coli. Sci. Rep. 2015, 5, 10465. [Google Scholar] [CrossRef] [Green Version]
- Paduch, R.; Kandefer-Szerszeń, M.; Trytek, M.; Fiedurek, J. Terpenes: Substances useful in human healthcare. Arch. Immunol. Ther. Exp. 2007, 55, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Ueda, D.; Yamaga, H.; Murakami, M.; Totsuka, Y.; Shinada, T.; Sato, T. Biosynthesis of Sesterterpenes, Head-to-Tail Triterpenes, and Sesquarterpenes in Bacillus clausii: Identification of Multifunctional Enzymes and Analysis of Isoprenoid Metabolites. ChemBioChem 2015, 16, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Yamaga, H.; Kashima, S.; Murata, Y.; Shinada, T.; Nakano, C.; Hoshino, T. Identification of novel sesterterpene/triterpene synthase from Bacillus clausii. Chembiochem 2013, 14, 822–825. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, V.; Vashisht, D.; Cletus, J.; Sakthivel, N. Plant β-1,3-glucanases: Their biological functions and transgenic expression against phytopathogenic fungi. Biotechnol. Lett. 2012, 34, 1983–1990. [Google Scholar] [CrossRef]
- Miyanishi, N.; Iwamoto, Y.; Watanabe, E.; Odaz, T. Induction of TNF-α production from human peripheral blood monocytes with β-1,3-glucan oligomer prepared from laminarin with β-1,3-glucanase from Bacillus clausii NM-1. J. Biosci. Bioeng. 2003, 95, 192–195. [Google Scholar] [CrossRef]
- Miyanishi, N.; Hamada, N.; Kobayashi, T.; Imada, C.; Watanabe, E. Purification and characterization of a novel extracellular β-1,3-glucanase produced by Bacillus clausii NM-1 isolated from ezo abalone Haliotis discus hannai. J. Biosci. Bioeng. 2003, 95, 45–51. [Google Scholar] [CrossRef]
- Wang, M.; Fu, J.; Zhang, X.; Chen, T. Metabolic engineering of Bacillus subtilis for enhanced production of acetoin. Biotechnol. Lett. 2012, 34, 1877–1885. [Google Scholar] [CrossRef]
- Muschallik, L.; Molinnus, D.; Bongaerts, J.; Pohl, M.; Wagner, T.; Schoning, M.J.; Siegert, P.; Selmer, T. (R,R)-Butane-2,3-diol dehydrogenase from Bacillus clausii DSM 8716(T): Cloning and expression of the bdhA-gene, and initial characterization of enzyme. J. Biotechnol. 2017, 258, 41–50. [Google Scholar] [CrossRef]
- Davis, M.E.; Brewster, M.E. Cyclodextrin-based pharmaceutics: Past, present and future. Nat. Rev. Drug Discov. 2004, 3, 1023–1035. [Google Scholar] [CrossRef]
- Alves-Prado, H.F.; Carneiro, A.A.; Pavezzi, F.C.; Gomes, E.; Boscolo, M.; Franco, C.M.; da Silva, R. Production of cyclodextrins by CGTase from Bacillus clausii using different starches as substrates. Appl. Biochem. Biotechnol. 2008, 146, 3–13. [Google Scholar] [CrossRef]
- Alves-Prado, H.F.; Bocchini, D.A.; Gomes, E.; Baida, L.C.; Contiero, J.; Roberto, I.C.; Da Silva, R. Optimization of cyclodextrin glucanotransferase production from Bacillus clausii E16 in submerged fermentation using response surface methodology. Appl. Biochem. Biotechnol. 2007, 137–140, 27–40. [Google Scholar] [CrossRef]
- Abbrescia, A.; Martino, P.L.; Panelli, D.; Sardanelli, A.M.; Papa, S.; Alifano, P.; Palese, L.L.; Gaballo, A. The respiratory chains of four strains of the alkaliphilic Bacillus clausii. FEBS Open Bio 2014, 4, 714–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lippolis, R.; Siciliano, R.A.; Mazzeo, M.F.; Abbrescia, A.; Gnoni, A.; Sardanelli, A.M.; Papa, S. Comparative secretome analysis of four isogenic Bacillus clausii probiotic strains. Proteome Sci. 2013, 11, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]

| Mechanism of Action | Tested Strain | Host Environment | Effect of Probiotic | Reference |
|---|---|---|---|---|
| Enhancing gut immune function | B. clausii (O/C, N/R, SIN, and T) live cells | Caco-2 cell line | Production of antimicrobial peptides, mucin, and tight junction proteins; increase of cell proliferation; release of pro-inflammatory cytokines | [48] |
| B. clausii (O/C, N/R, SIN, and T) live cells | Duodenal cells of esophagitis patients | Modulation of gene expression related to immunity, cell growth and death, cell signaling, and cell adhesion | [49] | |
| B. clausii as part of gut community in fecal microbiota transfer | Mice with pancreatic cancer | Alteration of tumor microbiome composition, tumor growth, and immune infiltration | [50] | |
| B. clausii SC-109 spores as part of a synbiotic formulation | Simulator of Human Intestinal Microbial Ecosystem (SHIME®) | Increased production of butyrate, alteration of gut microbiota | [51] | |
| B. clausii UBBC07 spores | Rat model of uremia | Reduction of acetaminophen-induced nephrotoxicity | [52] | |
| Antimicrobial and immunomodulatory activity | B. clausii Sinuberase® live cells | In vitro fermentation | Production of antimicrobial peptides | [53] |
| B. clausii UBBC07 live cells | SHIME® | Production of the lantibiotic clausin | [47] | |
| B. clausii O/C | In vitro culture medium | Production of the lantibiotic clausin | [54] | |
| Live cells of B. clausii isolate #KCTC 10,277 BP from tidal mudflats of the Korean Yellow Sea | Mouse model of allergic asthma | Reduction of inflammation | [55] | |
| B. clausii MTCC-8326 live cells | RAW264.7 murine macrophage cell line | Balance expression of pro- and anti-inflammatory cytokines, protection from S. typhimurium infections and related toxicity | [56,57] | |
| B. clausii O/C live cells | Caco-2 cell line | Protection from cytotoxic effects of Clostridium difficile and Bacillus cereus | [58] | |
| B. clausii (O/C, N/R, SIN, and T) live cells | Swiss murine peritoneal cells | Increased expression of pro-inflammatory cytokines and stimulation of nitrite production and proliferation of CD4+ T cells | [59] | |
| B. clausii O/C live cells | RAW 264.7 murine macrophage cell line | Induction of nitric oxide production | [60] | |
| B. clausii (O/C, N/R, SIN, and T) spores | Mouse model of ulcerative colitis | Slight improvement in symptoms of mild colitis | [61] | |
| B. clausii (O/C, N/R, SIN, and T) spores | Mouse model of schistosomiasis | Reduction of parasitic load and egg load, reduction of inflammation | [62] | |
| B. clausii (O/C, N/R, SIN, and T) spores | Mice with enteropathogenic E. coli infection | Reduction in intestinal lesions, debris and immune cell infiltration, increase in mucus-secreting goblet cells | [63] |
| Strain (Dose) | Study Design | Disease | Efficacy | Reference |
|---|---|---|---|---|
| O/C, N/R, SIN, T (2 × 109 to 4 × 109 CFU/day) | Prospective, open-label, multi-center, observational study; n = 3178 | Acute pediatric diarrhea | Reduced duration of diarrhea | [76] |
| O/C, SIN, N/R, T (2 × 109 to 4 × 109 CFU/day) | Meta-analysis; n = 898 from 6 studies | Rotavirus infection | Reduced frequency and duration of diarrhea Shortened hospital stay | [77] |
| O/C, SIN, N/R, T (6 × 109 spores/day) | Single-center, double blind, randomized, placebo-controlled prospective study; n = 120 Randomized, double blind, single-center, placebo-controlled, parallel group, phase 3b study; n = 130 | Helicobacter pylori treatment | Reduced nausea, diarrhea, and epigastric pain Fewer days of diarrhea Lower incidence of diarrhea | [27,28] |
| O/C, SIN, N/R, T | Randomized, double-blind, placebo-controlled trial; n = 244 | Necrotizing enterocolitis and late-onset sepsis | Faster attainment of full feeds | [78] |
| O/C, SIN, N/R, T | Randomized, single-blind, multi-center, two arm parallel group study; n = 80 | Upper respiratory tract infections | Fewer and shorter duration of infections | [79] |
| O/C, SIN, N/R, T | Single-blind, non-controlled study; n = 10 | Allergic rhinitis | Reduction in pro-inflammatory cytokines, higher levels of anti-inflammatory cytokines | [80,81] |
| UBBC07 (4 × 109 spores/day) | Randomized, double-blind, placebo-controlled trial; n = 153 | Acute pediatric diarrhea | Reduced stool frequency and duration of diarrhea | [82] |
| Unknown | Observational study; n = 65 | Rotavirus infection | Normalization of IgA and IgM to pre-infection levels Reduction in general weakness, swelling, and/or abdominal pain, fever, vomiting, and duration of diarrhea | [83,84] |
| Mix of Bacillus species, strain of B. clausii unavailable | Randomized, double-blind, placebo-controlled study with pre-screening for responders; n = 28 | Endotoxemia | Reduction in serum endotoxin and serum triglycerides, reduction in levels of pro-inflammatory markers | [85] |
| Unknown | Randomized controlled study; n = 80 | Recurrent aphthous stomatitis | Reduction in erythema, pain, burning sensation, and oral thrush | [86] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghelardi, E.; Abreu y Abreu, A.T.; Marzet, C.B.; Álvarez Calatayud, G.; Perez, M., III; Moschione Castro, A.P. Current Progress and Future Perspectives on the Use of Bacillus clausii. Microorganisms 2022, 10, 1246. https://doi.org/10.3390/microorganisms10061246
Ghelardi E, Abreu y Abreu AT, Marzet CB, Álvarez Calatayud G, Perez M III, Moschione Castro AP. Current Progress and Future Perspectives on the Use of Bacillus clausii. Microorganisms. 2022; 10(6):1246. https://doi.org/10.3390/microorganisms10061246
Chicago/Turabian StyleGhelardi, Emilia, Ana Teresa Abreu y Abreu, Christian Boggio Marzet, Guillermo Álvarez Calatayud, Marcos Perez, III, and Ana Paula Moschione Castro. 2022. "Current Progress and Future Perspectives on the Use of Bacillus clausii" Microorganisms 10, no. 6: 1246. https://doi.org/10.3390/microorganisms10061246
APA StyleGhelardi, E., Abreu y Abreu, A. T., Marzet, C. B., Álvarez Calatayud, G., Perez, M., III, & Moschione Castro, A. P. (2022). Current Progress and Future Perspectives on the Use of Bacillus clausii. Microorganisms, 10(6), 1246. https://doi.org/10.3390/microorganisms10061246