
The Role of Hydrogen Sulfide in Plant Roots during Development and in Response to Abiotic Stress

1
College of Life Science, Henan Agricultural University, Zhengzhou 450002, China
2
State Key Laboratory of Crop Biology, Shandong Agricultural University, Taian 271018, China
3
College of Agronomy, Henan Agricultural University, Zhengzhou 450002, China
4
State Key Laboratory of Wheat and Maize Crop Science, Henan Agricultural University,
Zhengzhou 450002, China
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(3), 1024; https://doi.org/10.3390/ijms23031024
Submission received: 27 December 2021
/
Revised: 16 January 2022
/
Accepted: 17 January 2022
/
Published: 18 January 2022
(This article belongs to the Special Issue Hydrogen Sulfide and Reactive Oxygen Species, Antioxidant Defense, Abiotic Stress Tolerance Mechanisms in Plants)
Abstract
:Hydrogen sulfide (H2S) is regarded as a “New Warrior” for managing plant stress. It also plays an important role in plant growth and development. The regulation of root system architecture (RSA) by H2S has been widely recognized. Plants are dependent on the RSA to meet their water and nutritional requirements. They are also partially dependent on the RSA for adapting to environment change. Therefore, a good understanding of how H2S affects the RSA could lead to improvements in both crop function and resistance to environmental change. In this review, we summarized the regulating effects of H2S on the RSA in terms of primary root growth, lateral and adventitious root formation, root hair development, and the formation of nodules. We also discussed the genes involved in the regulation of the RSA by H2S, and the relationships with other signal pathways. In addition, we discussed how H2S regulates root growth in response to abiotic stress. This review could provide a comprehensive understanding of the role of H2S in roots during development and under abiotic stress.
1. Introduction
The root system is an important vegetative organ of plants. In terrestrial environments, the root system provides structural support, uptakes water and nutrition from soil, and is where some amino acids, endogenous hormones, and other substances are synthesized. The growth and development of the root system largely determines the water and nutrient absorption efficiency of plants. With the improvement of genomics, genetics, molecular biology, and other research methods, as well as the generation of a mutant library related to root development, more and more functional genes and regulatory genes that affect root growth have been identified [1,2]. In addition to being regulated by internal genes, the physical environment can also have a regulatory effect on the growth and development of plant roots, such as the soil temperature, moisture, nutrients, and pH [3,4,5,6,7]. Plant signal molecules act as a bridge between the physical environment and root-growth-related genes, and hence determine how plants respond to environmental stress and changes. The plant signal molecules regulate the expression of root-growth-related genes through the transmission and transduction of environmental signals, which can lead to changes in the root system architecture (RSA) (including primary growth, the formation of lateral or adventitious roots, and the distribution and length of root hairs). These signal molecules include plant hormones, nitric oxide (NO), carbon monoxide (CO), reactive oxygen species (ROS), and hydrogen sulfide (H2S). The importance of plant hormones, NO, and ROS to root growth and development has been reviewed in many articles [8,9,10,11,12,13,14]. Hydrogen sulfide plays an important role in plant growth and development, and responds to various environmental stresses. In the past ten years, more than 2000 literatures have reported and discussed the impact of H2S on plant physiology. The importance of H2S for regulating plant responses to abiotic stresses (such as drought, salt, heat, and heavy metals) [15,16,17] and the effects on stomatal movement, seed germination, leaf senescence, and fruit ripening [18,19] have been extensively studied and discussed. In this review, we focused on the role of H2S in root growth and development. The effects of H2S on primary root growth, lateral and adventitious root formation, root hair development, and nodules were summarized here. As H2S can alleviate the negative impacts of abiotic stress on plant root growth, we also reviewed how H2S regulates root growth in response to abiotic stress.
2. The Role of Hydrogen Sulfide during Root Development
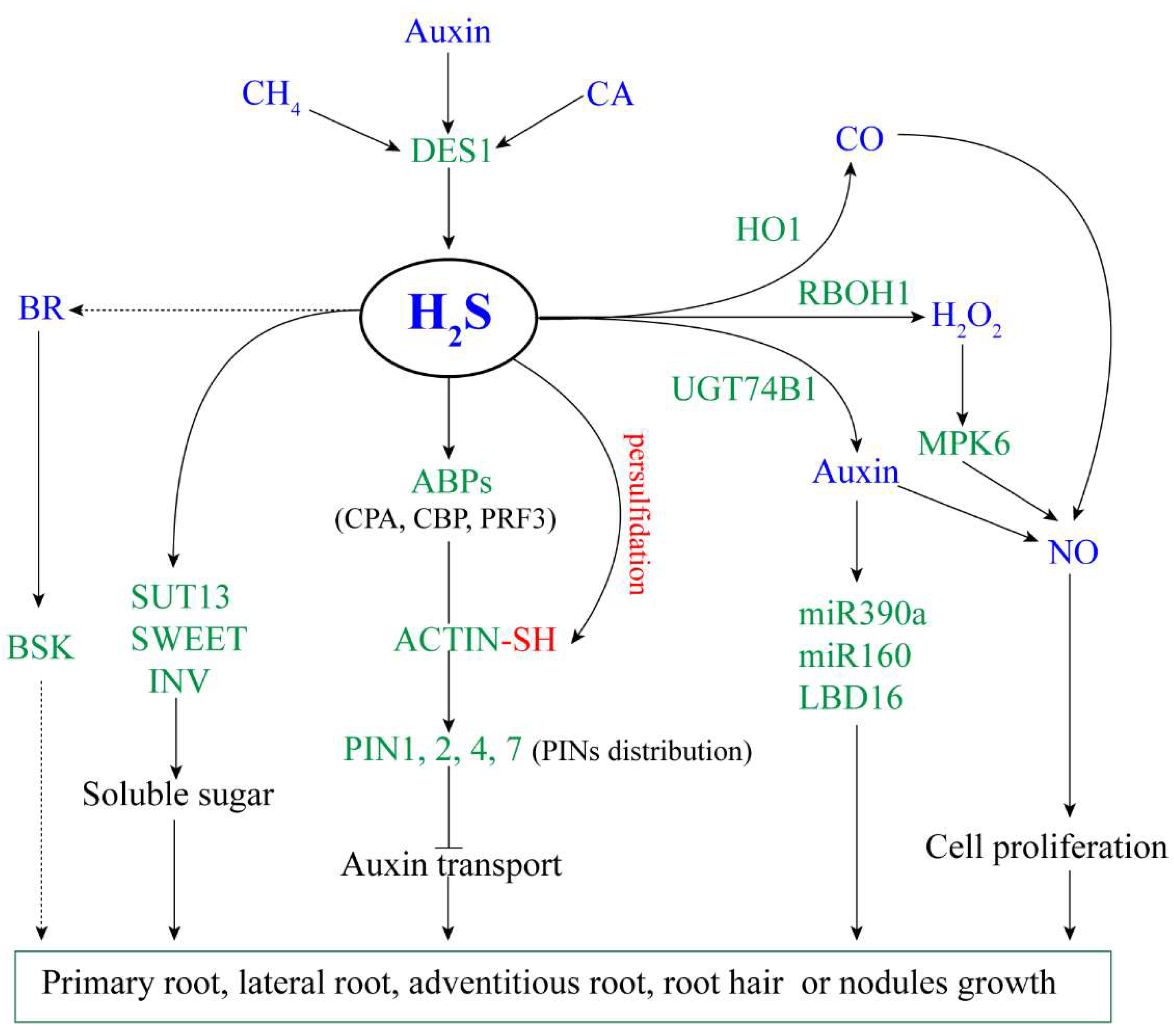
The literature on the regulation of H2S on root growth and development were shown in Table 1. According to these studies, we summarized a model for the regulatory mechanism of H2S on root growth and development (Figure 1).
2.1. Hydrogen Sulfide Regulates the Formation and Growth of Lateral and Adventitious Roots
Hydrogen sulfide has a concentration-dependent effect on the regulation of root growth. In tomato, 0.01–1.0 mM sodium hydrosulfide (NaHS) (a H2S donor) can significantly promote the initiation and length of lateral roots (LRs) and can increase the number and density of LRs. However, a high concentration of NaHS (10 mM) inhibits the LRs’ growth [23]. A similar phenomenon was found in the mangrove plant Kandelia obovata, where a concentration of 0.01–1.0 mM NaHS led to a notable increase in the length and total surface area of LRs [27]. H2S was essential for the formation of pepper LRs, where a concentration of 0.5–8.0 mM NaHS significantly increased the number of LRs. In contrast, different concentrations of the H2S scavenger hypotaurine (HT) markedly inhibited the formation of LRs [24]. In peach, H2S had a notable effect on the formation of LRs, with a concentration of 0.2 mM NaHS leading to a significant increase in the number of LRs [33]. Our previous research also found that H2S promoted the growth and development of lateral roots in wheat, with a concentration of 0.4 mM NaHS resulting in an increase in the number, density, and length of LRs [35]. However, in Arabidopsis, the effects of H2S on lateral roots were slightly different. H2S can promote the occurrence of LRs, but inhibits the LRs’ length [32], which may be related to the concentration of NaHS used in the treatment.
The effect of H2S on the formation and growth of adventitious roots is the same as that for lateral roots. The application of the appropriate concentration of H2S promoted the number and length of adventitious roots in sweet potato [20]. In addition, the same result was obtained for excised willow, soybeans [20], and cucumber [21,22].
2.2. Hydrogen Sulfide Inhibits the Growth of Primary Roots and Root Hair
Several studies have shown that H2S has a toxic effect on the growth of primary roots. The concentration of exogenous NaHS used in treatments was negatively correlated with the rate of growth (length) of primary roots [28,29,32]. This inhibitory effect of H2S on primary roots may be related to the reduction in the meristem cell division potential, as the length of the root meristematic zones were reduced when treated with NaHS [28]. Although H2S inhibited the length of root meristematic zones, Li et al. [29] found that the length from the root apex to a root hair for the seedlings was promoted by H2S. This may be due to the inhibitory effect of H2S on root hairs. H2S is known to inhibit the initiation of root hair; that is, the starting site of the root hair zones may be further away from the root apex [29], resulting in a longer distance from the root hair to root apex, even when H2S inhibits the meristem zones. In addition, H2S not only inhibited the initiation of root hair growth, but also significantly decreased the root hair length and density [29].
2.3. Hydrogen Sulfide Promotes the Formation of Root Nodules and Nitrogen Fixation
Root nodules are a special organ formed by symbiosis between leguminous plants (Fabaceae) and rhizobia. The formation and growth of the nodules are strictly controlled by plant hormones [36]. As a recognized signal molecule that interacts with plant hormones to regulate plant growth and development, H2S is known to influence the growth of root nodules [30,31]. Endogenous H2S production in both young soybean nodules (14 days post-inoculation [DPI] with the Sinorhizobium fredii Q8 strain) and mature nodules (28 DPI) can be detected by fluorescent probes SF7-A, whereas no significant fluorescence was observed in the nascent soybean nodules (7 DPI). This suggested that H2S may mediate the growth of root nodules [31]. Indeed, the application of NaHS significantly increased the number of soybean nodules and enhanced nitrogenase (Nase) activity after 7 DPI and 24 DPI, respectively. In addition, H2S was found to affect rhizobial infection, where a greater abundance of developing infection threads and cortex infection threads was found in NaHS-treated soybean roots than those in untreated controls at 5 DPI and 7 DPI, respectively [30]. On the contrary, an endogenous H2S production deficit rhizobia mutant ΔCSE (cystathionine γ-lyase) symbiosis with soybean roots significantly reduced the nitrogenase activity and H2S content in nodule cells. Moreover, higher contents of H2O2 (hydrogen peroxide), MDA (malondialdehyde), and protein carbonyl were observed in ΔCSE root nodules; that is, the H2S-induced nitrogen-fixation ability of root nodules may be related to its regulation of the antioxidant system that protects nodule cells from oxidative damage [31]. These studies suggested that H2S might have a positive effect on the soybean–rhizobium symbiosis system and may enhance nitrogen fixation.
2.4. Hydrogen Sulfide Interacts with Other Signaling Molecules to Regulate Root Development
2.4.1. Auxin
The inhibition of primary root growth by H2S, and the promotion of lateral and adventitious root formation was consistent with the known effects of auxin on root development. It is not difficult to associate H2S and auxin signaling to RSA. The change in the endogenous IAA (indole acetic acid) content was similar to that reported for H2S, but with different time-courses in sweet potato explants. The increase in the H2S content during the formation of sweet potato adventitious roots preceded changes to the IAA content [20]. The research of Wu et al. (2021) [33] on peach roots also obtained similar results: NaHS induced a significant increase in the endogenous H2S content in roots at 1 DAT (days after treatment), while it increased the concentration of endogenous auxin in roots by 44.50% at 5 DAT. Moreover, it was found that treatment with NaHS significantly increased the production of IAA, and that N-1-naphthylphthalamic acid (an IAA transport inhibitor, NPA) weakened the effect of H2S on the number of adventitious roots in sweet potato, soybean, and willow [20]. These results showed that IAA may be located downstream of H2S in order to mediate root development. However, the results in tomato indicated H2S might partially act as a downstream component of the auxin signaling to trigger lateral root formation [23]. The depletion of auxin down-regulated the transcription of SlDES1 (L-cysteine desulfhydrase 1, a H2S synthesis gene), DES activity, and endogenous H2S contents in tomato roots, and the inhibitory effect of NPA on lateral root formation was offset by NaHS, whereas the inhibition of lateral root formation by HT was not reversed by naphthalene acetic acid (NAA) [23]. In addition, H2S not only induced auxin synthesis, but also affected the auxin response and transport. After the application of NaHS, the expression of the indicator of the auxin response DR5::GUS (synthetic auxin-responsive promoter::β-glucuronidase) was attenuated in the quiescent center (QC), columella initial cells, and mature columella cells of the root apex, and was concentrated to the QC [32]. The movement of auxin in the root acropetal and basipetal was reduced by an increase in the NaHS concentration, which implied that an increase in H2S levels reduces the IAA transport capacity. Further research showed that the inhibition of IAA transport by H2S was related to the polar subcellular localization of PIN proteins (PIN1, PIN2, PIN4, and PIN7) [32].
2.4.2. Reactive Oxygen Species
High concentrations of ROS (reactive oxygen species) often cause oxidative damage to plants, but low concentrations of ROS are necessary for signaling to maintain plant growth and development. The ROS-related regulation of root development has been reported for Arabidopsis [37], tomato [38], maize [39,40], and sweet potato [41]. The relationship between ROS and H2S for the regulation of root growth was also discussed in several studies [25,28,34]. These studies found that ROS signaling might be downstream of H2S to mediate RSA. For example, H2S could induce the expression of RBOH1 (respiratory burst oxidase 1) in tomato roots and could enhance the accumulation of H2O2, thereby promoting lateral root formation. These H2S-related effects on lateral roots were destroyed by DMTU (dimethylthiourea, a H2O2 scavenger) and DPI (diphenylene idonium, an inhibitor of NADPH oxidase) [25]. The inhibitory effect of H2S on primary root growth depended on the ROS pathway, as the relative root growth in rbohF and rbohD/F was higher than that in WT for the NaHS treatment, which meant that respiratory burst oxidase homolog mutants (rboh) were less sensitive to treatment with NaHS [28]. The promoting effect of H2S on strawberry roots during plug transplant production could also be attributed (in part) to the elevated H2O2 [34].
2.4.3. Nitric Oxide and Carbon Monoxide
Nitric oxide (NO), carbon monoxide (CO), and H2S are the three gas signal molecules in organisms. NO and CO also participate in root growth and development [42,43,44,45,46,47]. Therefore, the relationship between H2S and NO or CO has attracted attention in the regulation of RSA. The H2S-mediated adventitious root formation was alleviated by 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (cPTIO, an NO scavenger) in sweet potato, willow, and soybean [20]. The toxic effect of H2S on the primary root of Arabidopsis was reduced in NO synthase mutants (nia1/2 and noa1), or when treated with cPTIO and NG-nitro-L-Arg-methyl ester (L-NAME, NO synthesis inhibitor) [28]. These results indicated that H2S acts upstream of NO signal transduction pathways when regulating adventitious root formation and primary root growth. From the results reported by Lin et al. (2012) [21], it is known that haem oxygenase-1/carbon monoxide (HO-1/CO) also acts as a downstream signal system during H2S-induced adventitious root formation. NaHS up-regulated HO1 gene expression and promoted HO1 protein accumulation, thereby increasing the number of cucumber adventitious roots. These phenomena were inhibited by ZnPPIX (zinc protoporphyrin IX, an inhibitor of HO-1), whereas the removal of H2S by HT did not affect the CO-induced adventitious rooting.
2.4.4. Brassinosteroid, Methane, and Cinnamaldehyde
Brassinosteroid (BR) contributes to the maintenance of root meristems, root cell elongation, lateral root development, root hair formation, and rhizosphere symbiosis [48,49,50,51]. At present, there is no direct evidence that H2S interacts with BR to regulate root development, but a recent proteomic analysis in Kandelia obovata has shown that H2S induced the accumulation of the BR-positive regulator protein BSK [27]. An RNA-seq analysis also showed that differentially expressed genes (DEGs) in peach roots, regulated by H2S, were significantly enriched in the “Brassinosteroid biosynthesis” pathway [33]. These results implied that H2S-mediated RSA might depend on the BR signal pathway.
Methane (CH4) plays an important role in the response to abiotic stress (such as heavy metal, salinity, and osmotic stress) [52]. In recent years, the role of CH4 in the formation of lateral and adventitious roots has been elucidated [22,26,53,54,55,56]. Both NO and CO signaling pathways were involved in CH4-induced adventitious root formation in cucumber [53,54]. Hydrogen peroxide (H2O2) signaling is also known to mediate the effects of CH4 on tomato lateral root formation [56]. As expected, H2S was confirmed to be located downstream of CH4 in order to regulate adventitious and lateral root formation in both cucumber and tomato. Methane induced the DES enzyme activity and promoted the production of endogenous H2S. These methane-related effects on the adventitious roots of cucumber were blocked by HT [22]. The same results were reported for the relationship between CH4 and H2S on the formation of lateral roots in tomato [26].
Cinnamaldehyde (CA) is a natural plant essential oil with antibacterial properties. It is widely used as a food additive and in medicines [57]. Recently, CA has also been used as a biological agent for plant disease resistance. For example, CA showed significant antibacterial activity against Pseudomonas syringae pv. actinidiae, which causes bacterial canker disease in kiwifruit [58]. Cinnamaldehyde reduced the number of Meloidogyne incognita galls and eggs on the roots of soybean plants to approximately 14% and 7%, respectively [59]. In addition, CA was found to play an important role in root development, as it markedly induced the formation of lateral roots in pepper, but without any inhibitory effect on primary root growth. Further study showed that H2S participated in this regulation process. Cinnamaldehyde increased the DES activity and promoted endogenous H2S production, thereby increasing the number of lateral roots. However, treatment with HT counteracted the effect of CA on endogenous H2S and lateral roots [24].
2.5. The Genes Involved in Hydrogen Sulfide-Mediated Root Development
Root system architecture is continuously adjusted in response to changes in various endogenous and exogenous factors (such as plant hormones, light, nutrition, and water). The regulation of these factors on root development involves many genes, including genes related to auxin synthesis, transport, and response, and genes related to cytokinin, abscisic acid (ABA), nitrate sensing and transport, and photoreceptors. The roles of these genes in plant growth and development were reviewed by Satbhai et al. (2015) [60]. In addition, many miRNAs are also involved in root development and architecture [2]. It is therefore important for researchers to have a clear understanding of which genes are involved in H2S signaling, and hence the regulation of root development. We have carried out a detailed discussion and summary of gene regulation below.
2.5.1. Genes Associated with the Auxin Signaling Pathway
The RNA-seq results for peach roots showed that 963 and 1113 DEGs were detected after H2S treatments for 1 day and 5 days, respectively [33]. These DEGs were significantly enriched in the “Glutathione metabolism”, “Plant-pathogen interaction”, “Plant hormone signal transduction”, “Brassinosteroid biosynthesis”, and “Cyanoamino acid metabolism” pathways. In particular, the pathway for “Plant hormone signal transduction” was significantly enriched when treated with H2S for 1 day and 5 days. A significant proportion (73.68%) of the genes associated with this pathway were related to auxin. More specifically, there were 2, 7, and 17 genes involved in auxin biosynthesis, transport, and signal transduction, respectively. These auxin-related genes included UGT74B1, TAA1, PINs, ABCBs, ARFs, Aux/IAAs, GH3, and SAUR. The auxin-synthesis-related gene UGT74B1 was up-regulated 1.95-fold when subjected to the H2S treatment. This might explain the H2S-induced increase in the root auxin content [20,33]. PINs exhibited different expression patterns over time under the NaHS treatment. After treatment with NaHS, PIN1 was up-regulated during 3 to 6 h and recovered to the control levels by 6 h, and the expression of PIN2 and PIN7 increased during 3 to 6 h, whereas it decreased in 12 or 24 h. On the contrary, the expression of PIN4 decreased after being treated with NaHS for 3 to 12 h, but recovered by 24 h. Although H2S had different effects on the expression of the PIN genes, its effect on the subcellular distribution of the PIN proteins was consistent. H2S disrupted the polar distribution of the PIN proteins (PIN1, PIN2, PIN4, and PIN7) on the plasma membrane in the root epidermal cells, and a large amount of PIN::GFP signals were found to dissociate from the plasma membrane upon cytoplasmic entry. Therefore, H2S inhibited auxin transport through its effect on the polarity distribution of PIN proteins, thus promoting the initiation of lateral roots [32]. It has been noted that the location of PIN proteins on the membrane was affected by F-actin [61,62], while H2S significantly reduced the occupancy rate of F-actin bundles in each cell. This led to the disappearance of thick actin cables [32]. This implied that the influence of H2S on the distribution of PIN proteins depended on the actin cytoskeleton, which is directly controlled by different ABPs (actin-binding proteins) [63]. Therefore, the expression of ABPs (CPA, CBP, and PRF3) was found to be up-regulated by H2S, whereas the effects of H2S on the percentage occupancy of the F-actin bundles was partially removed in the cpa, cbp, and prf3 mutants [32]. In addition, some auxin signal transduction genes were found to be regulated by H2S during root development. CsAux22D-like and CsAux22B-like were up-regulated by H2S during the formation of cucumber adventitious roots [22]. Hydrogen sulfide induced miR390a and miR160, and thus inhibited the expression of their target genes ARF4 and ARF16 in both tomato and Arabidopsis roots [25,26]. AtGATA23 and AtLBD16 were down-regulated in the Atdes1 mutant compared to WT, whereas AtGH3.1 and AtIAA28 were up-regulated in the Atdes1 mutant [26].
2.5.2. Genes Associated with Cell Proliferation
Cell proliferation is the basis for root growth and development, so the expression of cell-proliferation-related genes is very important during root growth. In the tomato root, H2S up-regulated SlCDKA;1, SlCYCA2;1, and AtCYCA2;3, but down-regulated SlKRP2 and AtKRP2 [25,26]. These genes are involved in the cell cycle. DNAJ-1, a gene phase that specifically regulates the G2/M cell cycle, was significantly induced by H2S in cucumber roots [21,22]. In addition, the expression of CsCDC6 (a cell-division-related gene) also increased in response to the NaHS treatment [22]. Interestingly, these cell proliferation-related genes also responded to auxin, CO, and CH4, which are closely related to the H2S signaling pathway. From the results of the RNA-seq work on peach roots, researchers identified that three cyclin genes and thirteen cell wall formation and remodeling-related genes were regulated by H2S [33]. All three cyclin genes (LOC109950471, LOC18790988, and LOC18784990) were up-regulated by H2S. In contrast, the cell wall formation and remodeling-related genes showed different patterns of expression in response to the H2S treatment [33].
2.5.3. Transcription Factors and Protein Kinases
Both transcription factors (TFs) and protein kinases are regulatory genes that mediate plant growth and development. Wu et al. (2021) [33] found that 36 transcription factors in peach roots were regulated by H2S, including LBD, MYB, and the AP2/ERF family. The overexpression of the peach PpLBD16, which was induced by H2S, significantly increased the number of lateral roots in Arabidopsis, whereas the Arabidopsis mutant ldb16 and ldb18 showed a decrease in the number of lateral roots [64]. These results strengthened our understanding of LBD-mediated lateral root growth. Interestingly, LBDs (such as AtLBD16, AtLBD18, and AtLBD29) have been shown to be directly regulated by ARFs when regulating the formation of lateral roots [65,66], which implies that H2S may interact with the auxin signaling pathway to regulate the growth of lateral roots, partly dependent on LBD genes. In Kandelia obovata roots, other TFs were also found to respond to H2S, such as trihelix transcription factor GT-3b (GT-3B), the zinc finger CCCH domain-containing protein 14 (ZC3H14), and the MADS-box transcription factor [27].
Previous studies have shown that several protein kinases respond to H2S during root development. The calmodulin kinases CsCDPK1 and CsCDPK5 were up-regulated by H2S in cucumber roots [21]. MPK6 was involved in H2S-inhibited primary root growth. When subjected to the NaHS treatment, the root length of the mutant mpk6 was significantly longer than that for WT. Moreover, MPK6 was shown to function downstream of H2S-induced ROS and upstream of NO [35]. In addition, in peach roots, the DEGs in the H2S treatment for five days were significantly enriched in the mitogen-activated protein kinase (MAPK) signaling pathway, relative to the control group [33]. These results suggested that CDPK and MAPK may play an important role in H2S-regulated root development.
2.5.4. Genes Associated with Carbohydrate Metabolism
Hu et al. (2020) [34] reported that H2S induced the accumulation of soluble sugar in strawberry roots during plug production. Subsequently, the transcriptome and proteome data showed that the H2S-regulated genes in roots were significantly enriched in “Starch and sucrose metabolism” [27,33]. These data indicated that soluble sugar was either directly or indirectly involved with H2S-regulated root development. The sucrose transport protein SUT13, bidirectional sugar transporter SWEET, and invertase (INV) were found to be up-regulated by H2S in Kandelia obovata roots, which led researchers to speculate that H2S may facilitate sucrose transport and promote the hydrolysis of sucrose to provide metabolites and energy for root growth.
3. The Role of Hydrogen Sulfide in Roots Exposed to Abiotic Stress
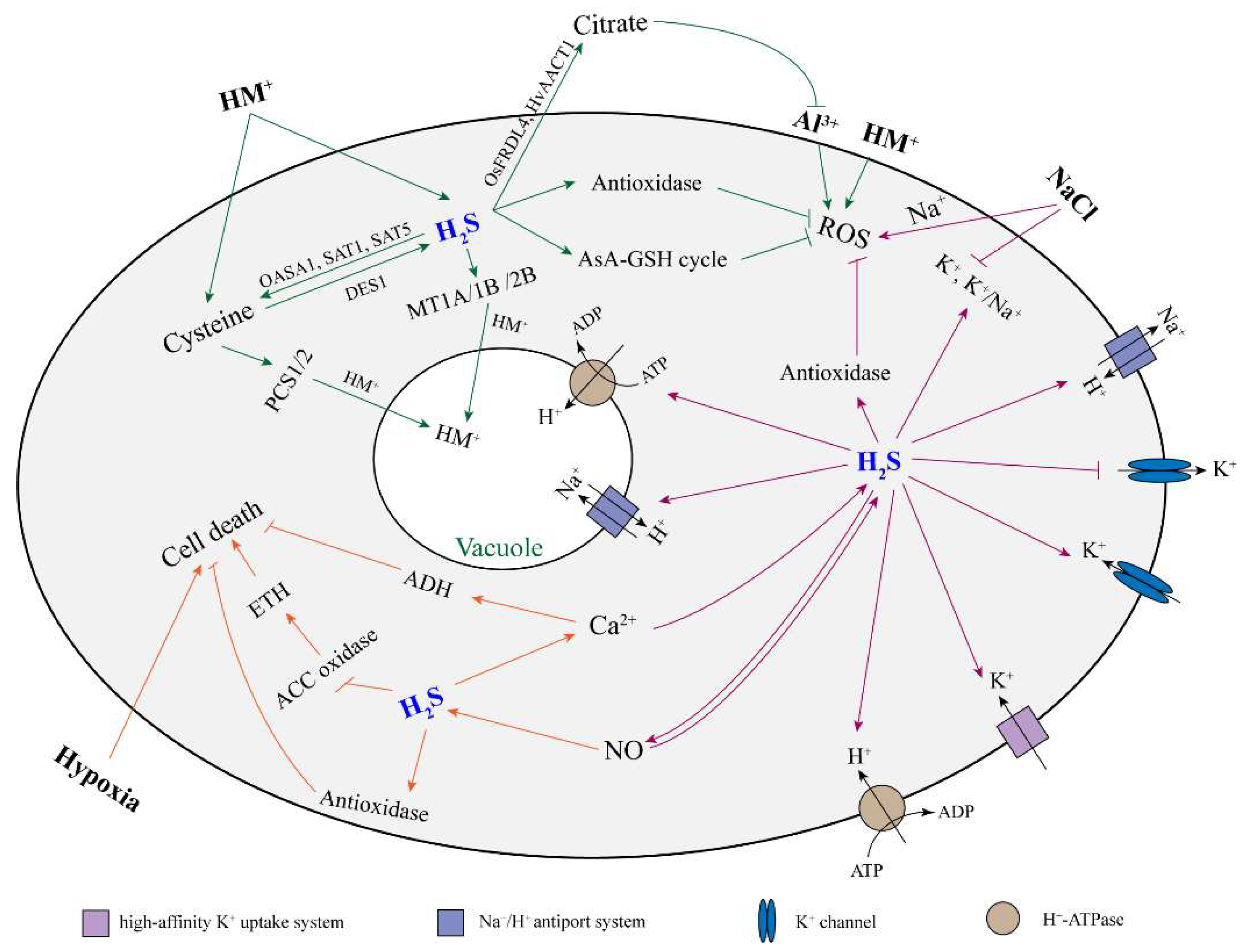
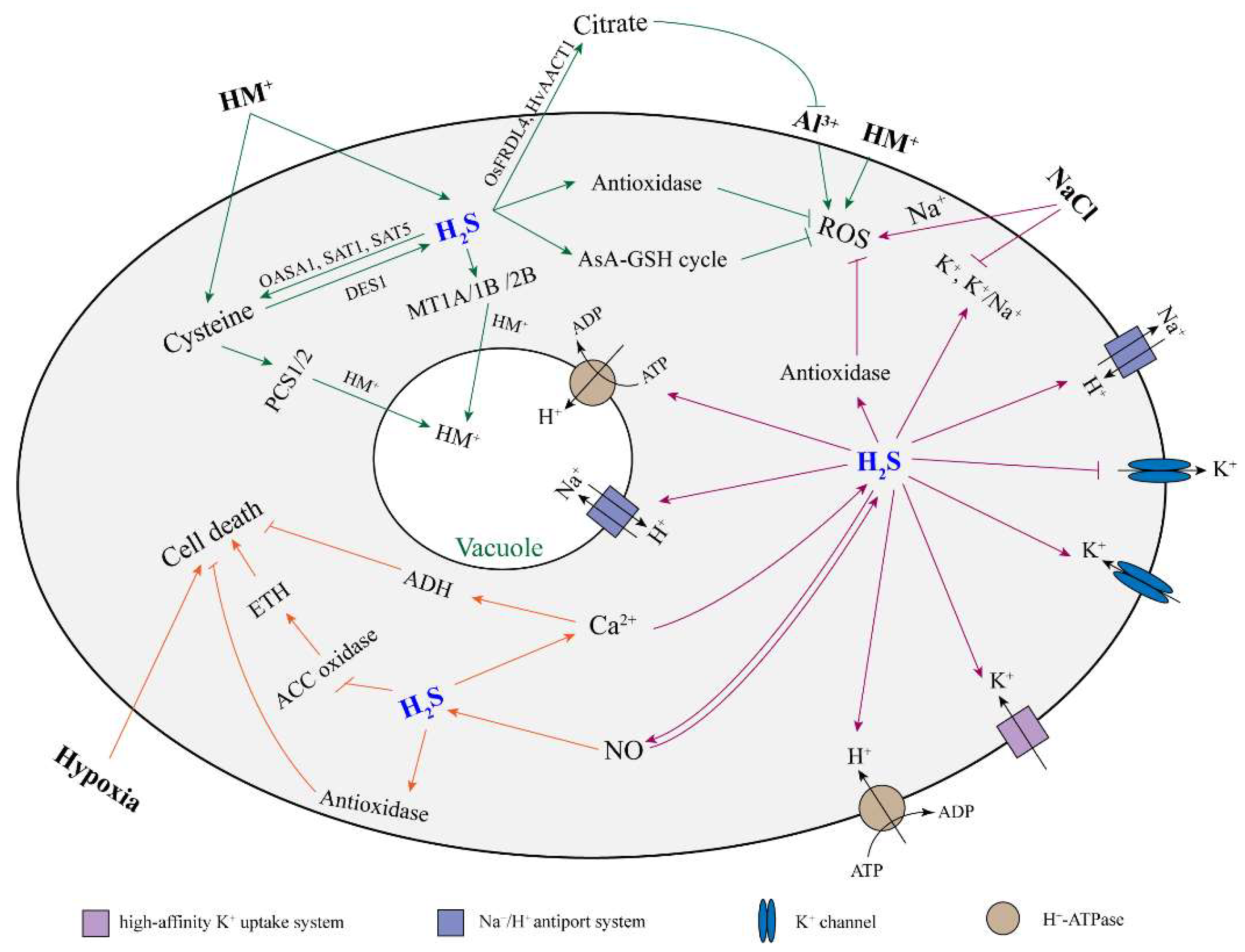
Abiotic stress often stimulates oxidative damage by generating ROS, which leads to the inhibition of plant growth and even death. Plant root growth is sensitive to abiotic stress factors in the soil, such as heavy metals (HMs), aluminum, salinity, and hypoxia. It has been reported that H2S could alleviate the inhibitory effect of abiotic stress on root growth in many plants (Table 2). Here, we discussed and reviewed the role and mechanism of H2S on root growth when exposed to an abiotic stress (Figure 2).
3.1. Heavy Metals
Cadmium (Cd) is regarded as the most toxic of the heavy metals (HMs) for plants. The function of H2S in plants subject to Cd-related stress has been extensively studied. Hydrogen sulfide could alleviate the Cd-induced inhibition of root growth in Arabidopsis [69], Medicago sativa [67,77], Brassica rapa [68,70,76], Setaria italica [71], Cucumis sativus [72,75], Solanum lycopersicum [74], and Hordeum vulgare [73]. Furthermore, the suppression of plant root growth caused by other heavy metals, such as chromium (Cr), lead (Pb), mercury (Hg), and nickel (Ni), could also be relieved when exposed to H2S [73,78,79,82,83,84,92,93].
When plants are exposed to HM-related stress, they first reduce the absorption of the HMs, or translocate the HMs to vacuoles to reduce oxidative damage to cells. As expected, the alleviation of the HM-related stress by H2S is partly dependent on these pathways. The Cd content in NaHS-pretreated root tissues was 33–37% lower than that for untreated Cd-stressed plant samples [67]. Pretreatment with NaHS markedly reduced the Pb content in maize roots [83]. The Ni content declined in the NaHS+Ni treatment of zucchini roots in comparison to the alone treatment of Ni-stressed [84]. Hydrogen sulfide also had a significant inhibitory effect on the absorption of Cr in cauliflower roots, stems, leaves, and flowers [79].
One of the most significant effects of HMs on plants is the production of large amounts of ROS, which, in turn, leads to oxidative damage to cells. The role of H2S in oxidative stress has been a major research focus for many years. Not surprisingly, the antioxidant function of H2S plays an important role in the response of plants to exposure to HM stress. When subjected to Cd-stress, treatment with NaHS reduced the accumulation of ROS and lipid peroxidation in Brassica rapa and barley [68,73]. The antioxidant function of H2S also had an effective response to Cr, Pb, and Hg-induced stress [78,79,82,83,93]. Generally, H2S reduced oxidative damage to cells, mainly by inducing antioxidant-related enzyme activity. The NaHS pretreatment significantly increased the activity of SOD (superoxide dismutase), APX (ascorbate peroxidase), CAT (catalase), and POD (peroxidase) in Cr-stressed cauliflower [79]. In addition, the positive effect of H2S on anti-oxidation was attributed to glutathione (GSH) homeostasis. The treatment with Cd significantly decreased the content of GSH and homoglutathione (hGSH) and increased the content of GSSG (oxidized GSH) and hGSSGh (oxidized hGSH), whereas the decreased ratio of hGSH/hGSSGh and GSH/GSSG in alfalfa seedlings was obviously inhibited by H2S [77]. In the presence of HT, the activities of the AsA-GSH-cycle-related enzymes (ascorbate peroxidase APX, monodehydroascorbate reductase MDHAR, dehydroascorbate reductase DHAR, and glutathione reductase GR) were reduced compared to the activities in the untreated Cd-stressed group, and thus the ratio of AsA/DHA (ascorbate/ dehydroascorbate) and GSH/GSSG declined [74].
In plants, H2S is mainly produced from cysteine desulfhydrylase (CDes) catalyzing the degradation of cysteine (Cys). Like H2S, when exposed to HM stress, the endogenous Cys content increased in Arabidopsis roots. The addition of exogenous Cys can significantly alleviate the inhibitory effect of HMs on primary growth. Cys synthesis could be induced by H2S by up-regulating the expression of the Cys-generation-related genes OASTLa, SAT1, and SAT5. Subsequently, Cys promoted GSH accumulation and induced the expression of phytochelatin (PC) genes (PCS1 and PCS2) counteracting Cd2+ and Cr6+ toxicity, and H2S could up-regulate the metallothionein gene (MT1A, MT1B, MT2B) to alleviate the Cd2+ and Cr6+ toxicity [69,78]. These results indicated that the H2S-Cys cycle system played a key role in plant responses to HM-related stress.
Hydrogen sulfide interacted with other signaling molecules to regulate heavy-metal-induced oxidative damage to cells. For example, the accumulation of NO was enhanced by NaHS treatment during exposure to Cd-induced stress. Hydrogen sulfide reduced Cd-induced oxidative damage in alfalfa roots, which was reversed by cPTIO [67]. MeJA enhanced Cd tolerance and alleviated growth inhibition in foxtail millet, whereas these effects were weakened by HT [71]. Similarly, the effects of CH4 on redox imbalance and cell death in alfalfa roots subjected to Cd stress was dependent on the induction of H2S metabolism. Treatment with either HT or PAG (propargylglycine, a H2S biosynthesis inhibitor) reduced the alleviating effects of CH4 on Cd-stressed plants [77].
3.2. Aluminum
Like heavy metal stress, excessive aluminum (Al) can also cause a large amount of ROS production in cells, resulting in oxidative damage and even cell death. Several studies have reported that H2S can weaken the inhibitory effect of aluminum toxicity on plant root growth by inhibiting Al3+ absorption and enhancing the antioxidant system. For example, both in barley and rice, the Al content in the leaves and roots of Al-stressed plants treated with NaHS was much lower than for untreated plants [80,81,94]. Further, the inhibitory effect of H2S on the absorption of Al3+ may be related to an increase in the secretion of citrate. In rice, the expression of OsFRDL4, a gene that regulates the efflux of citrate, was significantly up-regulated by NaHS treatments in Al-stressed conditions, and a simultaneous increase in citrate secretion from roots was found in the NaHS-pretreated group compared with the untreated Al-stressed plants [81]. Similarly, research on barley has shown that H2S could promote citrate secretion in roots and could up-regulate the expression of the citrate transporter gene (HvAACT1) when the plants were subjected to Al stress [80]. The increase in the rate of citrate secretion reduced the deposition of Al on the surface of roots. Therefore, the promotion of citrate secretion by H2S could lead to a reduction in the Al content of roots. Moreover, H2S-induced antioxidant-related enzyme activity also contributed to the mitigation of aluminum toxicity. The NaHS pretreatment significantly increased SOD, APX, CAT, and POD activity in Al-stressed rice [81]. Similarly, H2S was found to enhance SOD, CAT, POD, and GR activity in roots under Al stress in barley [94].
3.3. Salinity
According to previous studies, the inhibition of root growth by salinity (NaCl) stress could be attributed (at least in part) to a decrease in K+ concentrations and the K+/Na+ ratio in the cytoplasm. This would have disrupted ion homeostasis and hence caused cell death [95]. It displayed a net K+ efflux after exposure to NaCl, whereas NaHS could restrict the NaCl-induced K+ efflux in both salt-tolerant or salt-sensitive grape roots. Furthermore, H2S promoted Na+ efflux and the influx of H+ by up-regulating the Na+/H+ antiport system to maintain the plasma membrane (PM) polarity, thereby reducing the K+ loss by inhibiting PM depolarization-activated K+ channels [88]. K+ and Na+ homeostasis was an important adaptation by plants to salt stress. Researchers found that H2S significantly reduced the Na+ content and Na+/ K+ ratio in wheat roots. The H2S facilitated the exclusion of Na+ and absorption of K+ by regulating selective absorption and transport of K+ over Na+ [87]. The content of H2S in a Brassica napus hybrid was more than that of the two parents. When exposed to salt stress, the expression of NHX1 (Na+/H+ antiporter), AKT1 (inward-rectifying potassium channel), and HAK5 (potassium transporter) was significantly higher in the hybrid, in which, the Na+ content and Na+/ K+ ratio was reduced, and the K+ content increased. The hybrid, therefore, had a higher salt tolerance than the parents. However, these beneficial effects in the hybrid were eliminated by HT and PAG [96]. These results indicated that H2S improved the salt tolerance of plants by maintaining Na+ and K+ homeostasis. Other studies have shown that the regulation of Na+ and K+ homeostasis in salt-stressed plants by H2S involved the Ca2+ and NO signal pathways. Ca2+ and H2S had a synergistic effect on the induction of the Na+/H+ antiport system in mung bean roots. In contrast, the HT treatment negated the beneficial effects of Ca2+ on salt stress. Furthermore, a supplementation of Ca2+ could enhance H2S biosynthesis through promoting a cysteine pool. This implied the downstream functioning of H2S during the Ca2+-mediated regulation of plant adaptive responses to NaCl stress [97]. Both NO and H2S could increase the K+/Na+ ratio in alfalfa roots, whereas the treatment with cPTIO reduced the H2S-induced K+/Na+ ratio and antioxidant capacity of H2S [85]. When barley roots were exposed to salt stress, H2S could decrease the net K+ efflux by increasing the transcriptional expression of HvAKT1 (inward-rectifying potassium channel) and HvHAK4 (a high-affinity K+ uptake system), promote Na+ export by increasing the expression of PM H+-ATPase (HvHA1) and Na+/H+ antiporter (HvSOS1), and transfer excess Na+ into vacuoles by increasing the gene expression of vacuolar Na+/H+ antiporter (HvVNHX2), H+-ATPase subunit β (HvVHA-β), and the accumulating vacuolar Na+/H+ antiporter (NHE1) protein. However, these effects induced by H2S were quenched by the addition of cPTIO [86]. These results mean that the H2S is upstream of NO in order to maintain ion homeostasis and improve salt tolerance. However, Da Silva et al. (2018) [89] proposed that H2S may act downstream of NO in the mitigation of salt-induced oxidative stress. Researchers found that, after treatment with NaCl, the accumulation of H2S in tomato roots occurred later than the accumulation of NO, and that NO could increase the expression of the H2S synthesis gene (L-DES) and H2S production, whereas H2S could not induce the accumulation of NO. H2S and NO have shown complex interactions when regulating other physiological processes [98]. Therefore, the relationship between H2S and NO in plant root growth regulation under salt stress needs more research and discussion.
3.4. Hypoxia
Hypoxia leads to root cell death. However, H2S could reduce the rate of root tip cell death by inducing antioxidant enzyme activity and by inhibiting ACC oxidase (ACO) activity and ethylene production [90]. H2S also promoted endogenous Ca2+ accumulation and the Ca2+-dependent activity of alcohol dehydrogenase (ADH). It therefore improved the antioxidant defensive capabilities of the plants, and thus increased the rate of maize root tip cell survival in hypoxic conditions [91]. Subsequent studies have shown that the regulation of root tip cell death by H2S mediated the NO signal pathway. The NO-induced tolerance of hypoxia was enhanced by the application of NaHS, but was eliminated by HT [91].
4. Conclusions and Future Prospects
The effects of H2S on plant root growth and development have been widely recognized. In this review, we summarized the regulatory effects of H2S on lateral roots, adventitious roots, primary roots, root hairs, and root nodules. The mitigation effect of H2S on root growth under abiotic stress was also discussed here. Hydrogen sulfide interacts with a variety of other signals to regulate root growth. These signals mainly included auxin, NO, CO, ROS, and CH4. In addition, there are many genes involved in H2S-regulated root growth. However, there are still many issues that need to be clarified to explain how H2S regulates root growth. For example, H2S interacts with other signal molecules to regulate root growth, so finding the key genes that connect H2S and other signal molecules is crucial for understanding the complex interactions between H2S and signal molecules. Previous studies have found that many genes contribute to the regulation of H2S during root growth and development. A great number of genes involved in the regulation of H2S on root systems were identified through transcriptome and proteome, but the involvement of these genes was based on the effects of H2S on their expression. The importance of these genes to the H2S-regulated root growth pathway requires further functional verification. Finally, in recent years, studies have found that H2S could directly regulate the S-sulfhydration of proteins by converting Cys-SH to Cys-SSH. This affected the activity of proteins, and, thus, mediated plant growth and development and responses to stresses [99,100,101,102,103]. The ACTIN2 protein, associated with the development of root hair, has been found to be S-sulfhydrated at Cys-287 by H2S, thereby mediating H2S-regulated root hair growth [29]. This implied that there might be more proteins involved in root development that are S-sulfhydrated by H2S that still need to be identified.
Author Contributions
H.L. and C.W. conceived and planned this review paper. H.L. and H.C. prepared and drafted the manuscript. L.C. and C.W. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the State Key Laboratory of Crop Biology Open Fund (2021KF02) and Natural Science Foundation of Henan Province (212300410352).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Motte, H.; Vanneste, S.; Beeckman, T. Molecular and environmental regulation of root development. Annu. Rev. Plant Biol. 2019, 70, 465–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, G.A.; Declerck, M.; Sorin, C.; Hartmann, C.; Crespi, M.; Lelandais-Brière, C. MicroRNAs as regulators of root development and architecture. Plant Mol. Biol. 2011, 77, 47–58. [Google Scholar] [CrossRef]
- Karlova, R.; Boer, D.; Hayes, S.; Testerink, C. Root plasticity under abiotic stress. Plant Physiol. 2021, 187, 1057–1070. [Google Scholar] [CrossRef]
- Balliu, A.; Zheng, Y.; Sallaku, G.; Fernández, J.A.; Gruda, N.S.; Tuzel, Y. Environmental and cultivation factors affect the morphology, architecture and performance of root systems in soilless grown plants. Horticulturae 2021, 7, 243. [Google Scholar] [CrossRef]
- Bouain, N.; Krouk, G.; Lacombe, B.; Rouached, H. Getting to the root of plant mineral nutrition: Combinatorial nutrient stresses reveal emergent properties. Trends Plant Sci. 2019, 24, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Aggarwal, P.; Robbins, N.E.; Sturrock, C.J.; Thompson, M.C.; Tan, H.Q.; Tham, C.; Duan, L.; Rodriguez, P.L.; Vernoux, T.; et al. Plant roots use a patterning mechanism to position lateral root branches toward available water. PNAS 2014, 111, 9319–9324. [Google Scholar] [CrossRef] [Green Version]
- Robbins, N.E.; Dinneny, J.R. The divining root: Moisture-driven responses of roots at the micro-and macro-scale. J. Exp. Bot. 2015, 66, 2145–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demecsová, L.; Tamás, L. Reactive oxygen species, auxin and nitric oxide in metal-stressed roots: Toxicity or defence. Biometals 2019, 32, 717–744. [Google Scholar] [CrossRef]
- Xu, P.; Zhao, P.X.; Cai, X.T.; Mao, J.L.; Miao, Z.Q.; Xiang, C.B. Integration of jasmonic acid and ethylene into auxin signaling in root development. Front. Plant Sci. 2020, 11, 271. [Google Scholar] [CrossRef]
- Hebelstrup, K.H.; Shah, J.K.; Igamberdiev, A.U. The role of nitric oxide and hemoglobin in plant development and morphogenesis. Physiol. Plant. 2013, 148, 457–469. [Google Scholar] [CrossRef]
- Lindsey, K.; Rowe, J.; Liu, J. Hormonal crosstalk for root development: A combined experimental and modeling perspective. Front. Plant Sci. 2014, 5, 116. [Google Scholar]
- Corpas, F.J. Barroso, J.B. Functions of nitric oxide (NO) in roots during development and under adverse stress conditions. Plants 2015, 4, 240–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacifici, E.; Polverari, L.; Sabatini, S. Plant hormone cross-talk: The pivot of root growth. J. Exp. Bot. 2015, 66, 1113–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukagoshi, H. Control of root growth and development by reactive oxygen species. Curr. Opin Plant Biol. 2016, 29, 57–63. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, M.; Zhou, H.; Zhao, D.; Gotor, C.; Romero, L.C.; Shen, J.; Ge, Z.; Zhang, Z.; Shen, W.; et al. Hydrogen sulfide, a signaling molecule in plant stress responses. J. Integr. Plant Biol. 2021, 63, 146–160. [Google Scholar] [CrossRef]
- Pandey, A.K.; Gautam, A. Stress responsive gene regulation in relation to hydrogen sulfide in plants under abiotic stress. Physiol. Plant. 2020, 168, 511–525. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Huo, J.; Liao, W. Hydrogen sulfide: Roles in plant abiotic stress response and crosstalk with other signals. Plant Sci. 2021, 302, 110733. [Google Scholar] [CrossRef] [PubMed]
- Arif, Y.; Hayat, S.; Yusuf, M.; Bajguz, A. Hydrogen sulfide: A versatile gaseous molecule in plants. Plant Physiol. Bioch. 2021, 158, 372–384. [Google Scholar] [CrossRef]
- Xuan, L.; Li, J.; Wang, X.; Wang, C. Crosstalk between hydrogen sulfide and other signal molecules regulates plant growth and development. Int. J. Mol. Sci. 2020, 21, 4593. [Google Scholar] [CrossRef]
- Zhang, H.; Tang, J.; Liu, X.P.; Wang, Y.; Yu, W.; Peng, W.Y.; Fang, F.; Ma, D.F.; Wei, Z.J.; Hu, L.Y. Hydrogen sulfide promotes root organogenesis in Ipomoea batatas, Salix matsudana and Glycine max. J. Integr. Plant Biol. 2009, 51, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.T.; Li, M.Y.; Cui, W.T.; Lu, W.; Shen, W.B. Haem oxygenase-1 is involved in hydrogen sulfide-induced cucumber adventitious root formation. J. Plant Growth Regul. 2012, 31, 519–528. [Google Scholar] [CrossRef]
- Kou, N.; Xiang, Z.; Cui, W.; Li, L.; Shen, W. Hydrogen sulfide acts downstream of methane to induce cucumber adventitious root development. J. Plant Physiol. 2018, 228, 113–120. [Google Scholar] [CrossRef]
- Fang, T.; Cao, Z.; Li, J.; Shen, W.; Huang, L. Auxin-induced hydrogen sulfide generation is involved in lateral root formation in tomato. Plant Physiol. Bioch. 2014, 76, 44–51. [Google Scholar] [CrossRef]
- Xue, Y.F.; Zhang, M.; Qi, Z.Q.; Li, Y.Q.; Shi, Z.; Chen, J. Cinnamaldehyde promotes root branching by regulating endogenous hydrogen sulfide. J. Sci. Food Agric. 2016, 96, 909–914. [Google Scholar] [CrossRef]
- Mei, Y.; Chen, H.; Shen, W.; Shen, W.; Huang, L. Hydrogen peroxide is involved in hydrogen sulfide-induced lateral root formation in tomato seedlings. BMC Plant Biol. 2017, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.; Zhao, Y.; Jin, X.; Wang, R.; Xu, N.; Hu, J.; Huang, L.Q.; Guan, R.Z.; Shen, W. L-Cysteine desulfhydrase-dependent hydrogen sulfide is required for methane-induced lateral root formation. Plant Mol. Biol. 2019, 99, 283–298. [Google Scholar] [CrossRef]
- Li, H.; Ghoto, K.; Wei, M.Y.; Gao, C.H.; Liu, Y.L.; Ma, D.N.; Zheng, H.L. Unraveling hydrogen sulfide-promoted lateral root development and growth in mangrove plant Kandelia obovata: Insight into regulatory mechanism by TMT-based quantitative proteomic approaches. Tree Physiol. 2021, 41, 1749–1766. [Google Scholar] [CrossRef]
- Zhang, P.; Luo, Q.; Wang, R.; Xu, J. Hydrogen sulfide toxicity inhibits primary root growth through the ROS-NO pathway. Sci Rep. 2017, 7, 868. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, S.; Wang, X.; Shi, C.; Liu, H.; Yang, J.; Shi, W.; Guo, J.; Jia, H. Hydrogen sulfide disturbs actin polymerization via S-sulfhydration resulting in stunted root hair growth. Plant Physiol. 2018, 178, 936–949. [Google Scholar] [CrossRef] [Green Version]
- Zou, H.; Zhang, N.N.; Pan, Q.; Zhang, J.H.; Chen, J.; Wei, G.H. Hydrogen sulfide promotes nodulation and nitrogen fixation in soybean–rhizobia symbiotic system. Mol. Plant Microbe. Interact. 2019, 32, 972–985. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Zhang, N.N.; Lin, X.Y.; Zhang, W.Q.; Zhang, J.H.; Chen, J.; Wei, G.H. Hydrogen sulfide is a crucial element of the antioxidant defense system in Glycine max–Sinorhizobium fredii symbiotic root nodules. Plant Soil. 2020, 449. [Google Scholar] [CrossRef]
- Jia, H.; Hu, Y.; Fan, T.; Li, J. Hydrogen sulfide modulates actin-dependent auxin transport via regulating ABPs results in changing of root development in Arabidopsis. Sci. Rep. 2015, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Du, A.; Zhang, S.; Wang, W.; Liang, J.; Peng, F.; Xiao, Y. Regulation of growth in peach roots by exogenous hydrogen sulfide based on RNA-Seq. Plant Physiol. Bioch. 2021, 159, 179–192. [Google Scholar] [CrossRef]
- Hu, J.; Li, Y.; Liu, Y.; Kang, D.I.; Wei, H.; Jeong, B.R. Hydrogen sulfide affects the root development of strawberry during plug transplant production. Agriculture. 2020, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, Y.; Zhang, L.; Zhao, H.; Li, H. Hydrogen sulphide improves iron homeostasis in wheat under iron-deficiency. Plant Sci. 2017, 5, 170. [Google Scholar]
- Bensmihen, S. Hormonal control of lateral root and nodule development in legumes. Plants 2015, 4, 523–547. [Google Scholar] [CrossRef] [PubMed]
- Dunand, C.; Crèvecoeur, M.; Penel, C. Distribution of superoxide and hydrogen peroxide in Arabidopsis root and their influence on root development: Possible interaction with peroxidases. New Phytol. 2007, 174, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Ivanchenko, M.G.; Den Os, D.; Monshausen, G.B.; Dubrovsky, J.G.; Bednářová, A.; Krishnan, N. Auxin increases the hydrogen peroxide (H2O2) concentration in tomato (Solanum lycopersicum) root tips while inhibiting root growth. Ann. Bot. 2013, 112, 1107–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liszkay, A.; van der Zalm, E.; Schopfer, P. Production of reactive oxygen intermediates (O2−, H2O2, and OH) by maize roots and their role in wall loosening and elongation growth. Plant physiol. 2004, 136, 3114–3123. [Google Scholar] [CrossRef] [Green Version]
- Voothuluru, P.; Mäkelä, P.; Zhu, J.; Yamaguchi, M.; Cho, I.J.; Oliver, M.J.; Simmonds, J.; Sharp, R.E. Apoplastic hydrogen peroxide in the growth zone of the maize primary root. Increased levels differentially modulate root elongation under well-watered and water-stressed conditions. Front. Plant Sci. 2020, 11, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, X.P.; Cheng, Y.J.; Wu, X.B.; Kwak, S.S.; Chen, W.; Eneji, A.E. Exogenous hydrogen peroxide positively influences root growth and exogenous hydrogen peroxide positively influences root growth and metabolism in leaves of sweet potato seedlings. Aust. J. Crop. Sci. 2012, 6, 1572. [Google Scholar]
- Böhm, F.M.L.Z.; Ferrarese, M.D.L.L.; Zanardo, D.I.L.; Magalhaes, J.R.; Ferrarese-Filho, O. Nitric oxide affecting root growth, lignification and related enzymes in soybean seedlings. Acta Physiol. Plant. 2010, 32, 1039–1046. [Google Scholar] [CrossRef]
- Chen, Y.H.; Chao, Y.Y.; Hsu, Y.Y.; Hong, C.Y.; Kao, C.H. Heme oxygenase is involved in nitric oxide-and auxin-induced lateral root formation in rice. Plant Cell Rep. 2012, 31, 1085–1091. [Google Scholar] [CrossRef]
- Fernández-Marcos, M.; Sanz, L.; Lorenzo, O. Nitric oxide: An emerging regulator of cell elongation during primary root growth. Plant Signal. Behav. 2012, 7, 196–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Zhang, W.; Qi, F.; Cui, W.; Xie, Y.; Shen, W. Hydrogen-rich water regulates cucumber adventitious root development in a heme oxygenase-1/carbon monoxide-dependent manner. J. Plant Physiol. 2014, 171, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sanz, L.; Albertos, P.; Mateos, I.; Sánchez-Vicente, I.; Lechón, T.; Fernández-Marcos, M.; Lorenzo, O. Nitric oxide (NO) and phytohormones crosstalk during early plant development. J. Exp. Bot. 2015, 66, 2857–2868. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, M.; Hu, L.; Liao, W.; Dawuda, M.M.; Li, C. Carbon monoxide is involved in hydrogen gas-induced adventitious root development in cucumber under simulated drought stress. Front. Plant Sci. 2017, 8, 128. [Google Scholar] [CrossRef]
- Hacham, Y.; Holland, N.; Butterfield, C.; Ubeda-Tomas, S.; Bennett, M.J.; Chory, J.; Savaldi-Goldstein, S. Brassinosteroid perception in the epidermis controls root meristem size. Development 2011, 138, 839–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.; Singh, M.; Laxmi, A. Interaction between glucose and brassinosteroid during the regulation of lateral root development in Arabidopsis. Plant Physiol. 2015, 168, 307–320. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Li, J. Brassinosteroids regulate root growth, development, and symbiosis. Mol. Plant 2016, 9, 86–100. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.H.; Breda, A.; Hardtke, C.S. Brassinosteroid signaling directs formative cell divisions and protophloem differentiation in Arabidopsis root meristems. Development 2017, 144, 272–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Wei, S.; Shen, W. The role of methane in plant physiology: A review. Plant Cell Rep. 2020, 39, 171–179. [Google Scholar] [CrossRef]
- Cui, W.; Qi, F.; Zhang, Y.; Cao, H.; Zhang, J.; Wang, R.; Shen, W. Methane-rich water induces cucumber adventitious rooting through heme oxygenase1/carbon monoxide and Ca2+ pathways. Plant Cell Rep. 2015, 34, 435–445. [Google Scholar] [CrossRef]
- Qi, F.; Xiang, Z.; Kou, N.; Cui, W.; Xu, D.; Wang, R.; Zhu, D.; Shen, W. Nitric oxide is involved in methane-induced adventitious root formation in cucumber. Physiol. Plant. 2017, 159, 366–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; He, J.; Cheng, P.; Xiang, Z.; Zhou, H.; Wang, R.; Shen, W. Methane control of adventitious rooting requires γ-glutamyl cysteine synthetase-mediated glutathione homeostasis. Plant Cell Physiol. 2019, 60, 802–815. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, Y.; Liu, F.; Wang, R.; Huang, L.; Shen, W. Hydrogen peroxide is involved in methane-induced tomato lateral root formation. Plant Cell Rep. 2019, 38, 377–389. [Google Scholar] [CrossRef]
- Vergis, J.; Gokulakrishnan, P.; Agarwal, R.K.; Kumar, A. Essential oils as natural food antimicrobial agents: A review. Crit Rev. Food Sci. Nutr. 2015, 55, 1320–1323. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.R.; Choi, M.S.; Choi, G.W.; Park, I.K.; Oh, C.S. Antibacterial activity of cinnamaldehyde and estragole extracted from plant essential oils against Pseudomonas syringae pv. actinidiae causing bacterial canker disease in kiwifruit. Plant Pathol. J. 2016, 32, 363. [Google Scholar] [CrossRef] [Green Version]
- Jardim, I.N.; Oliveira, D.F.; Silva, G.H.; Campos, V.P.; de Souza, P.E. (E)-cinnamaldehyde from the essential oil of Cinnamomum cassia controls Meloidogyne incognita in soybean plants. J. Pest. Sci. 2018, 91, 479–487. [Google Scholar] [CrossRef]
- Satbhai, S.B.; Ristova, D.; Busch, W. Underground tuning: Quantitative regulation of root growth. J. Exp. Bot. 2015, 66, 1099–1112. [Google Scholar] [CrossRef] [Green Version]
- Muday, G.K.; Murphy, A.S. An emerging model of auxin transport regulation. Plant Cell. 2002, 14, 293–299. [Google Scholar] [CrossRef]
- Dhonukshe, P.; Grigoriev, I.; Fischer, R.; Tominaga, M.; Robinson, D.G.; Hašek, J.; Paciorek, T.; Petrásek, J.; Seifertová, D.; Tejos, R.; et al. Auxin transport inhibitors impair vesicle motility and actin cytoskeleton dynamics in diverse eukaryotes. Proc. Natl Acad Sci. USA 2008, 105, 4489–4494. [Google Scholar] [CrossRef] [Green Version]
- Pollard, T.D.; Cooper, J.A. Actin, a central player in cell shape and movement. Science 2009, 326, 1208–1212. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.W.; Kim, N.Y.; Lee, D.J.; Kim, J. LBD18/ASL20 regulates lateral root formation in combination with LBD16/ASL18 downstream of ARF7 and ARF19 in Arabidopsis. Plant Physiol. 2009, 151, 1377–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okushima, Y.; Fukaki, H.; Onoda, M.; Theologis, A.; Tasaka, M. ARF7 and ARF19 regulate lateral root formation via direct activation of LBD/ASL genes in Arabidopsis. Plant Cell. 2007, 19, 118–130. [Google Scholar] [CrossRef] [Green Version]
- Porco, S.; Larrieu, A.; Du, Y.; Gaudinier, A.; Goh, T.; Swarup, K.; Swarup, R.; Kuempers, B.; Bishopp, A.; Lavenus, J.; et al. Lateral root emergence in Arabidopsis is dependent on transcription factor LBD29 regulation of auxin influx carrier LAX3. Development 2016, 143, 3340–3349. [Google Scholar]
- Li, L.; Wang, Y.; Shen, W. Roles of hydrogen sulfide and nitric oxide in the alleviation of cadmium-induced oxidative damage in alfalfa seedling roots. Biometals 2012, 25, 617–631. [Google Scholar] [CrossRef]
- Zhang, L.; Pei, Y.; Wang, H.; Jin, Z.; Liu, Z.; Qiao, Z.; Fang, H.; Zhang, Y. Hydrogen sulfide alleviates cadmium-induced cell death through restraining ROS accumulation in roots of Brassica rapa L. ssp. pekinensis. Oxid Med. Cell Longev. 2015, 2015, 804603. [Google Scholar]
- Jia, H.; Wang, X.; Dou, Y.; Liu, D.; Si, W.; Fang, H.; Zhao, C.; Chen, S.; Xi, J.; Li, J. Hydrogen sulfide-cysteine cycle system enhances cadmium tolerance through alleviating cadmium-induced oxidative stress and ion toxicity in Arabidopsis roots. Sci Rep. 2016, 6, 39702. [Google Scholar] [CrossRef]
- Lv, W.; Yang, L.; Xu, C.; Shi, Z.; Shao, J.; Xian, M.; Chen, J. Cadmium disrupts the balance between hydrogen peroxide and superoxide radical by regulating endogenous hydrogen sulfide in the root tip of Brassica rapa. Front. Plant Sci. 2017, 8, 232. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.; Zhang, Y.; Jin, Z.; Liu, Z.; Pei, Y. Role of hydrogen sulfide in the methyl jasmonate response to cadmium stress in foxtail millet. Front. Biosci. 2017, 22, 530–538. [Google Scholar]
- Kabała, K.; Zboińska, M.; Głowiak, D.; Reda, M.; Jakubowska, D.; Janicka, M. Interaction between the signaling molecules hydrogen sulfide and hydrogen peroxide and their role in vacuolar H+-ATPase regulation in cadmium-stressed cucumber roots. Physiol. Plant. 2019, 166, 688–704. [Google Scholar] [CrossRef] [Green Version]
- Fu, M.M.; Dawood, M.; Wang, N.H.; Wu, F. Exogenous hydrogen sulfide reduces cadmium uptake and alleviates cadmium toxicity in barley. Plant Growth Regul. 2019, 89, 227–237. [Google Scholar] [CrossRef]
- Alamri, S.; Kushwaha, B.K.; Singh, V.P.; Siddiqui, M.H. Dose dependent differential effects of toxic metal cadmium in tomato roots: Role of endogenous hydrogen sulfide. Ecotoxicol. Environ. Saf. 2020, 203, 110978. [Google Scholar] [CrossRef]
- Luo, S.; Tang, Z.; Yu, J.; Liao, W.; Xie, J.; Lv, J.; Feng, Z.; Dawuda, M.M. Hydrogen sulfide negatively regulates cd-induced cell death in cucumber (Cucumis sativus L) root tip cells. BMC Plant Biol. 2020, 20, 480. [Google Scholar] [CrossRef]
- Li, G.; Shah, A.A.; Khan, W.U.; Yasin, N.A.; Ahmad, A.; Abbas, M.; Ali, A.; Safdar, N. Hydrogen sulfide mitigates cadmium induced toxicity in Brassica rapa by modulating physiochemical attributes, osmolyte metabolism and antioxidative machinery. Chemosphere 2021, 263, 127999. [Google Scholar] [CrossRef]
- Yang, X.; Kong, L.; Wang, Y.; Su, J.; Shen, W. Methane control of cadmium tolerance in alfalfa roots requires hydrogen sulfide. Environ. Pollut. 2021, 284, 117123. [Google Scholar] [CrossRef]
- Fang, H.; Liu, Z.; Jin, Z.; Zhang, L.; Liu, D.; Pei, Y. An emphasis of hydrogen sulfide-cysteine cycle on enhancing the tolerance to chromium stress in Arabidopsis. Environ. Pollut. 2016, 213, 870–877. [Google Scholar] [CrossRef]
- Ahmad, R.; Ali, S.; Rizwan, M.; Dawood, M.; Farid, M.; Hussain, A.; Wijaya, L.; Alyemeni, M.; Ahmad, P. Hydrogen sulfide alleviates chromium stress on cauliflower by restricting its uptake and enhancing antioxidative system. Physiol. Plant. 2020, 168, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wang, W.H.; Wu, F.H.; You, C.Y.; Liu, T.W.; Dong, X.J.; He, J.X.; Zheng, H.L. Hydrogen sulfide alleviates aluminum toxicity in barley seedlings. Plant Soil. 2013, 362, 301–318. [Google Scholar] [CrossRef]
- Zhu, C.Q.; Zhang, J.H.; Sun, L.M.; Zhu, L.F.; Abliz, B.; Hu, W.J.; Zhong, C.; Bai, Z.G.; Sajid, H.; Cao, X.C.; et al. Hydrogen sulfide alleviates aluminum toxicity via decreasing apoplast and symplast Al contents in rice. Front. Plant Sci. 2018, 9, 294. [Google Scholar] [CrossRef] [Green Version]
- Ali, B.; Mwamba, T.M.; Gill, R.A.; Yang, C.; Ali, S.; Daud, M.K.; Wu, Y.; Zhou, W. Improvement of element uptake and antioxidative defense in Brassica napus under lead stress by application of hydrogen sulfide. Plant Growth Regul. 2014, 74, 261–273. [Google Scholar] [CrossRef]
- Zanganeh, R.; Jamei, R.; Rahmani, F. Role of salicylic acid and hydrogen sulfide in promoting lead stress tolerance and regulating free amino acid composition in Zea mays L. Acta Physiol. Plant. 2019, 41, 94. [Google Scholar] [CrossRef]
- Valivand, M.; Amooaghaie, R.; Ahadi, A. Interplay between hydrogen sulfide and calcium/calmodulin enhances systemic acquired acclimation and antioxidative defense against nickel toxicity in zucchini. Environ. Exp. Bot. 2019, 158, 40–50. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.; Cui, W.; Xu, S.; Shen, W.; Wang, R. Hydrogen sulfide enhances alfalfa (Medicago sativa) tolerance against salinity during seed germination by nitric oxide pathway. Plant Soil 2012, 351, 107–119. [Google Scholar] [CrossRef]
- Chen, J.; Wang, W.H.; Wu, F.H.; He, E.M.; Liu, X.; Shangguan, Z.P.; Zheng, H.L. Hydrogen sulfide enhances salt tolerance through nitric oxide-mediated maintenance of ion homeostasis in barley seedling roots. Sci. Rep. 2015, 5, 12516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.Q.; Bao, J.; Yuan, F.; Liang, X.; Feng, Z.T.; Wang, B.S. Exogenous hydrogen sulfide alleviates salt stress in wheat seedlings by decreasing Na+ content. Plant Growth Regul. 2016, 79, 391–399. [Google Scholar] [CrossRef]
- Zhao, N.; Zhu, H.; Zhang, H.; Sun, J.; Zhou, J.; Deng, C.; Zhang, Y. Chen, S. Hydrogen sulfide mediates K+ and Na+ homeostasis in the roots of salt-resistant and salt-sensitive poplar species subjected to NaCl stress. Front. Plant Sci. 2018, 9, 1366. [Google Scholar] [CrossRef] [Green Version]
- da-Silva, C.J.; Mollica, D.C.; Vicente, M.H.; Peres, L.E.; Modolo, L.V. NO, hydrogen sulfide does not come first during tomato response to high salinity. Nitric Oxide 2018, 76, 164–173. [Google Scholar] [CrossRef]
- Cheng, W.; Zhang, L.; Jiao, C.; Su, M.; Yang, T.; Zhou, L.; Peng, R.; Wang, R.; Wang, C. Hydrogen sulfide alleviates hypoxia-induced root tip death in Pisum sativum. Plant Physiol. Biochem. 2013, 70, 278–286. [Google Scholar] [CrossRef]
- Peng, R.; Bian, Z.; Zhou, L.; Cheng, W.; Hai, N.; Yang, C.; Yang, T.; Wang, X.; Wang, C. Hydrogen sulfide enhances nitric oxide-induced tolerance of hypoxia in maize (Zea mays L.). Plant Cell Rep. 2016, 35, 2325–2340. [Google Scholar] [CrossRef]
- Ali, S.; Farooq, M.A.; Hussain, S.; Yasmeen, T.; Abbasi, G.H.; Zhang, G. Alleviation of chromium toxicity by hydrogen sulfide in barley. Environ. Toxicol. Chem. 2013, 32, 2234–2239. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, M.; Jiang, M. Hydrogen sulfide alleviates mercury toxicity by sequestering it in roots or regulating reactive oxygen species productions in rice seedlings. Plant Physiol. Biochem. 2017, 111, 179–192. [Google Scholar] [CrossRef]
- Dawood, M.; Cao, F.; Jahangir, M.M.; Zhang, G.; Wu, F. Alleviation of aluminum toxicity by hydrogen sulfide is related to elevated ATPase, and suppressed aluminum uptake and oxidative stress in barley. J. Hazard. Mater. 2012, 209, 121–128. [Google Scholar] [CrossRef]
- Basu, S.; Kumar, A.; Benazir, I.; Kumar, G. Reassessing the role of ion homeostasis for improving salinity tolerance in crop plants. Physiol. Plant. 2021, 171, 502–519. [Google Scholar] [CrossRef]
- Cheng, P.; Zhang, Y.; Wang, J.; Guan, R.; Pu, H.; Shen, W. Importance of hydrogen sulfide as the molecular basis of heterosis in hybrid Brassica napus: A case study in salinity response. Environ. Exp. Bot. 2021, 193, 104693. [Google Scholar] [CrossRef]
- Khan, M.N.; Siddiqui, M.H.; Mukherjee, S.; Alamri, S.; Al-Amri, A.A.; Alsubaie, Q.D.; Ai-Munqedhi, B.M.A.; Ali, H.M. Calcium-hydrogen sulfide crosstalk during K+-deficient NaCl stress operates through regulation of Na+/H+ antiport and antioxidative defense system in mung bean roots. Plant Physiol. Biochem. 2021, 159, 211–225. [Google Scholar] [CrossRef]
- Mishra, V.; Singh, P.; Tripathi, D.K.; Corpas, F.J.; Singh, V.P. Nitric oxide and hydrogen sulfide: An indispensable combination for plant functioning. Trends Plant Sci. 2021, 26, 1270–1285. [Google Scholar] [CrossRef]
- Jia, H.; Chen, S.; Liu, D.; Liesche, J.; Shi, C.; Wang, J.; Ren, M.; Wang, X.; Yang, J.; Shi, W.; et al. Ethylene-induced hydrogen sulfide negatively regulates ethylene biosynthesis by persulfidation of ACO in tomato under osmotic stress. Front. Plant Sci. 2018, 9, 1517. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Zhang, J.; Zhou, M.; Zhou, H.; Cui, B.; Gotor, C.; Romero, L.C.; Fu, L.; Yang, J.; Foyer, C.H.; et al. Persulfidation-based modification of cysteine desulfhydrase and the NADPH oxidase RBOHD controls guard cell abscisic acid signaling. Plant Cell. 2020, 32, 1000–1017. [Google Scholar] [CrossRef]
- Chen, S.; Jia, H.; Wang, X.; Shi, C.; Wang, X.; Ma, P.; Wang, J.; Ren, M.; Li, J. Hydrogen sulfide positively regulates abscisic acid signaling through persulfidation of SnRK2.6 in guard cells. Mol. Plant 2020, 13, 732–744. [Google Scholar] [CrossRef]
- Li, J.; Shi, C.; Wang, X.; Liu, C.; Ding, X.; Ma, P. Hydrogen sulfide regulates the activity of antioxidant enzymes through persulfidation and improves the resistance of tomato seedling to Copper Oxide nanoparticles (CuO NPs)-induced oxidative stress. Plant Physiol. Biochem. 2020, 156, 257–266. [Google Scholar] [CrossRef]
- Aroca, A.; Zhang, J.; Xie, Y.; Romero, L.C.; Gotor, C. Hydrogen sulfide signaling in plant adaptations to adverse conditions: Molecular mechanisms. J. Exp. Bot. 2021, 72, 5893–5904. [Google Scholar] [CrossRef]
Figure 1.
The proposed model of H2S regulating plant root growth. Arrow and bar ends indicate activation and inhibitory effects, respectively. Green fonts represent genes or proteins, blue fonts represent signal molecules. H2S: hydrogen sulfide; CH4: methane; CA: cinnamaldehyde; BR: brassinosteroid; CO: carbon monoxide; NO: nitric oxide; DES1: L-cysteine desulfhydrase 1; HO1: haem oxygenase-1; RBOH1: respiratory burst oxidase 1; UGT74B1: UDP-glycosyltransferase 74B1; MPK6: mitogen-activated protein kinase 6; ABPs: actin-binding proteins; PINs: pin-formed family; SUT13: sugar transport protein 13; SWEET: bidirectional sugar transporter; INV: invertase; BSK: BR-signaling kinase; LBD16: LOB domain-containing protein 16.
Figure 1.
The proposed model of H2S regulating plant root growth. Arrow and bar ends indicate activation and inhibitory effects, respectively. Green fonts represent genes or proteins, blue fonts represent signal molecules. H2S: hydrogen sulfide; CH4: methane; CA: cinnamaldehyde; BR: brassinosteroid; CO: carbon monoxide; NO: nitric oxide; DES1: L-cysteine desulfhydrase 1; HO1: haem oxygenase-1; RBOH1: respiratory burst oxidase 1; UGT74B1: UDP-glycosyltransferase 74B1; MPK6: mitogen-activated protein kinase 6; ABPs: actin-binding proteins; PINs: pin-formed family; SUT13: sugar transport protein 13; SWEET: bidirectional sugar transporter; INV: invertase; BSK: BR-signaling kinase; LBD16: LOB domain-containing protein 16.
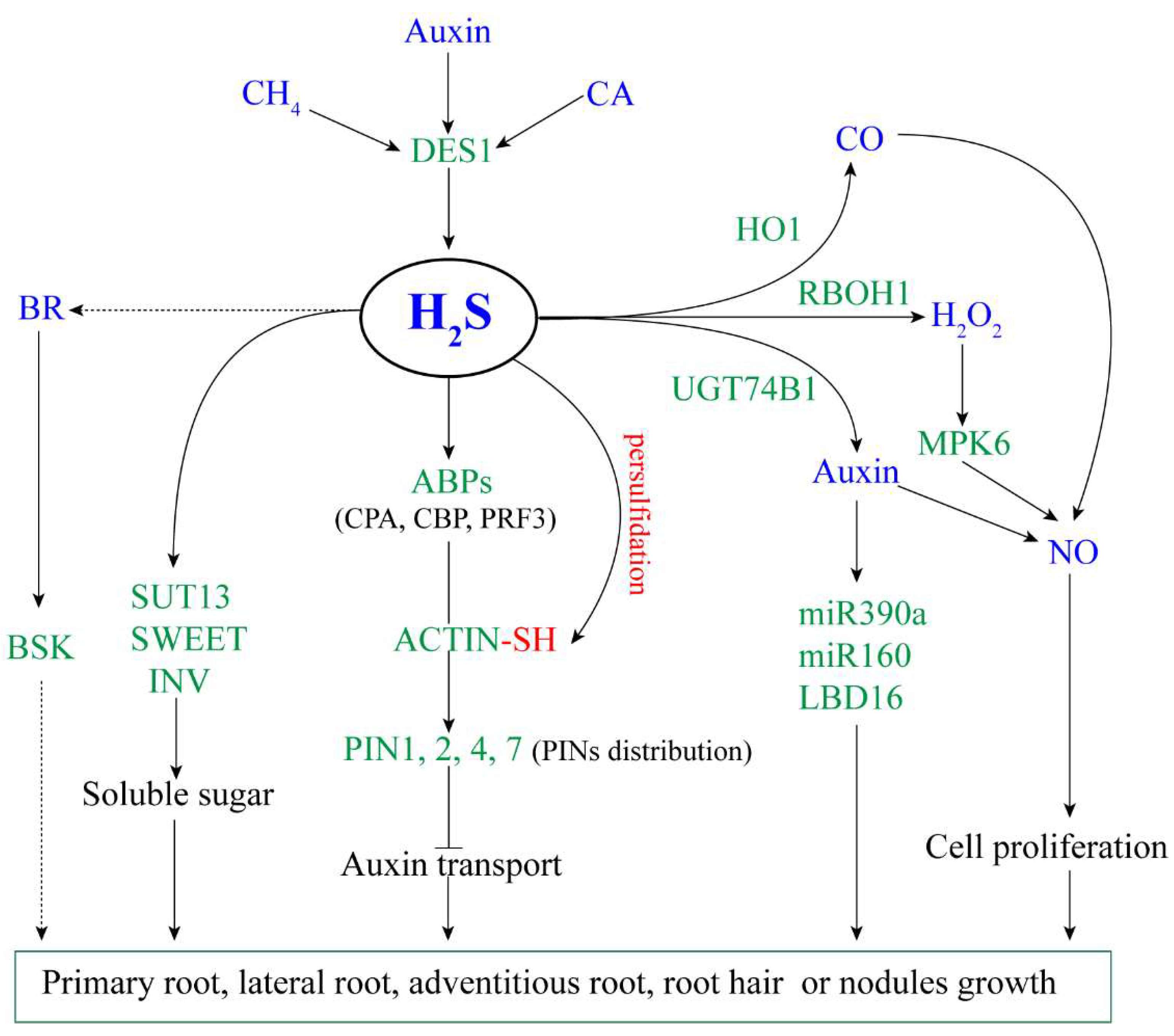
Figure 2.
The proposed model of H2S alleviating plant root cell damage under abiotic stress. Arrow and bar ends indicate activation and inhibitory effects, respectively. The green, orange, and purple arrows refer to the response of H2S in plant roots to heavy metals, hypoxia, and salt stress, respectively. H2S: hydrogen sulfide; HM: heavy metal; ROS: reactive oxygen species; NO: nitric oxide; AsA-GSH cycle: ascorbate–glutathione cycle; ETH: ethylene; ADH: alcohol dehydrogenase; DES1: L-cysteine desulfhydrase 1; OASA1: o-acetylserine lyase isoform A1; SAT1: serine acetyltransferase 1; SAT5: serine acetyltransferase 5; PCS1/5: phytochelatin synthase 1/5; MT1A/1B/2B: metallothionein 1A/1B/2B; FRDL4: citrate efflux transporter; AACT1: citrate transporter.
Figure 2.
The proposed model of H2S alleviating plant root cell damage under abiotic stress. Arrow and bar ends indicate activation and inhibitory effects, respectively. The green, orange, and purple arrows refer to the response of H2S in plant roots to heavy metals, hypoxia, and salt stress, respectively. H2S: hydrogen sulfide; HM: heavy metal; ROS: reactive oxygen species; NO: nitric oxide; AsA-GSH cycle: ascorbate–glutathione cycle; ETH: ethylene; ADH: alcohol dehydrogenase; DES1: L-cysteine desulfhydrase 1; OASA1: o-acetylserine lyase isoform A1; SAT1: serine acetyltransferase 1; SAT5: serine acetyltransferase 5; PCS1/5: phytochelatin synthase 1/5; MT1A/1B/2B: metallothionein 1A/1B/2B; FRDL4: citrate efflux transporter; AACT1: citrate transporter.

{kind=link}
{kind=link}
Table 1.
The role of hydrogen sulfide in root during development and its interaction with other signals.
Table 1.
The role of hydrogen sulfide in root during development and its interaction with other signals.
| Plant Species | Signal Involved | Root Response | Reference |
|---|---|---|---|
| Ipomoea batatas, Salix matsudana, Glycine max | Auxin and NO | Adventitious root formation | [20] |
| Cucumis sativus | HO-1/CO | Adventitious root formation | [21] |
| Cucumis sativus | Methane | Adventitious root development | [22] |
| Solanum lycopersicum | Auxin | Lateral root formation | [23] |
| Capsicum annuum | Cinnamaldehyde | Lateral root formation | [24] |
| Solanum lycopersicum | H2O2 | Lateral root formation | [25] |
| Solanum lycopersicum | Methane | Lateral root formation | [26] |
| Kandelia obovata | Brassinosteroid, carbohydrate metabolism, cellular redox homeostasis, protein metabolism, secondary metabolism, and amino acid metabolism | Lateral root development | [27] |
| Arabidopsis | ROS, NO, MPK6 | Primary root growth | [28] |
| Arabidopsis | Actin dynamics | Root hair growth | [29] |
| Glycine max | Nitrogen-fixation ability | Nodulation | [30] |
| Glycine max | Nitrogen-fixation ability | Nodulation | [31] |
| Arabidopsis | Actin-dependent auxin transport | Root development and growth | [32] |
| Prunus persica | Auxin biosynthesis, transport, and signal transduction. | Root development and growth | [33] |
| Fragaria × ananassa | H2O2 and soluble sugar accumulation | Root development and growth | [34] |
Table 2.
Hydrogen sulfide promotes root growth and its regulation mechanism under abiotic stress.
| Abiotic Stress | H2S Action | Plant Species | Reference |
|---|---|---|---|
| Cadmium | H2S improved oxidation resistance, and NO was involved in the NaHS-induced alleviation of Cd toxicity | Medicago sativa | [67] |
| H2S removed excessive ROS and reduced cell oxidative damage | Brassica rapa | [68] | |
| H2S inhibited the ROS burst, and H2S-Cys cycle system plays an important role in it | Arabidopsis | [69] | |
| H2S mediated the phytotoxicity of Cd by regulating UPB1s-modulated balance between H2O2 and O2− | Brassica rapa | [70] | |
| H2S relieved-Cd stress was involved in MeJA signal | Setaria italica | [71] | |
| H2O2 raised H2S content in root tissues independently from the desulfhydrase activity, and protected V-ATPase | Cucumis sativus | [72] | |
| H2S reduced Cd uptake/translocation and decreased MDA, H2O2, and O2− accumulation | Hordeum vulgare | [73] | |
| H2S activated glutathione biosynthetic and AsA-GSH cycle enzymes, and maintained redox status of ascorbate and glutathione | Solanum lycopersicum | [74] | |
| H2S inhibited Cd-induced cell death by reducing ROS accumulation, activating the antioxidant system, inhibiting mitochondrial Cyt c release, and reducing the opening of the MPTP | Cucumis sativus | [75] | |
| H2S improved Cd tolerance by modulating growth biomarkers and antioxidative system | Brassica rapa | [76] | |
| H2S operates downstream of CH4, enhancing tolerance against Cd stress | Medicago sativa | [77] | |
| Chromium | H2S increased Cys accumulation by up-regulating the Cys generation-related genes, enhanced glutathione generation, and activated phytochelatins (PCs) synthesis | Arabidopsis | [78] |
| H2S improved the physiological and biochemical attributes of Cr-stressed plants, and decreased Cr content in different parts of Cr-stressed plants | Brassica oleracea botrytis | [79] | |
| Aluminum | H2S protected plants against Al toxicity by inducing the activities of antioxidant enzymes, increasing citrate secretion and citrate transporter gene expression, and enhancing the expression of PM H+-ATPase. | Hordeum vulgare | [80] |
| H2S alleviated Al toxicity by decreasing the Al content in the apoplast and symplast | Oryza sativa | [81] | |
| Lead | H2S lowered the Pb concentration in roots, improved the cell structure, and presented the well-developed nucleus with continuous cell membrane | Brassica napus | [82] |
| H2S alleviated Pb toxicity by improvement of nitrate reductase activity and glutathione content and regulation of amino acids metabolism | Zea mays | [83] | |
| Nickel | H2S induced Ni tolerance that required the entry of extracellular Ca2+ into cells across the plasma membrane and the mediation of intracellular CaM | Cucurbita pepo | [84] |
| Salt | H2S enhanced plant responses against salinity stress by reducing oxidative damage, which might have a possible interaction with NO | Medicago sativa | [85] |
| H2S increased salt tolerance by maintaining Na+ and K+ ion homeostasis, which was mediated by NO signal | Hordeum vulgare | [86] | |
| H2S alleviated growth inhibition by maintaining a lower Na+ concentration under NaCl stress via the regulation of NSCCs and SOS1 pathways | Triticum aestivuml | [87] | |
| H2S up-regulated the Na+/H+ antiport system, which promoted exchange of Na+ with H+ across the PM and simultaneously restricted the channel-mediated K+ loss | Populus euphratica and Populus popularis | [88] | |
| H2S acts downstream of NO in the mitigation of NaCl-induced oxidative stress | Solanum lycopersicum | [89] | |
| Hypoxia | H2S protected root tip cell membranes from ROS damage induced by hypoxia, and stimulated a quiescence strategy through inhibiting ethylene production | Vigna radiata | [90] |
| H2S enhanced endogenous Ca2+ levels, as well as the Ca2+-dependent activity of alcohol dehydrogenase (ADH), improved the capacity for antioxidant defense, and thus increased the NO-induced hypoxia tolerance in maize | Zea mays | [91] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, H.; Chen, H.; Chen, L.; Wang, C. The Role of Hydrogen Sulfide in Plant Roots during Development and in Response to Abiotic Stress. Int. J. Mol. Sci. 2022, 23, 1024. https://doi.org/10.3390/ijms23031024
AMA Style
Li H, Chen H, Chen L, Wang C. The Role of Hydrogen Sulfide in Plant Roots during Development and in Response to Abiotic Stress. International Journal of Molecular Sciences. 2022; 23(3):1024. https://doi.org/10.3390/ijms23031024
Chicago/Turabian StyleLi, Hua, Hongyu Chen, Lulu Chen, and Chenyang Wang. 2022. "The Role of Hydrogen Sulfide in Plant Roots during Development and in Response to Abiotic Stress" International Journal of Molecular Sciences 23, no. 3: 1024. https://doi.org/10.3390/ijms23031024
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.