
miR-199a Downregulation as a Driver of the NOX4/HIF-1α/VEGF-A Pathway in Thyroid and Orbital Adipose Tissues from Graves′ Patients

 , ,
, ,
Abstract
:1. Introduction
2. Results
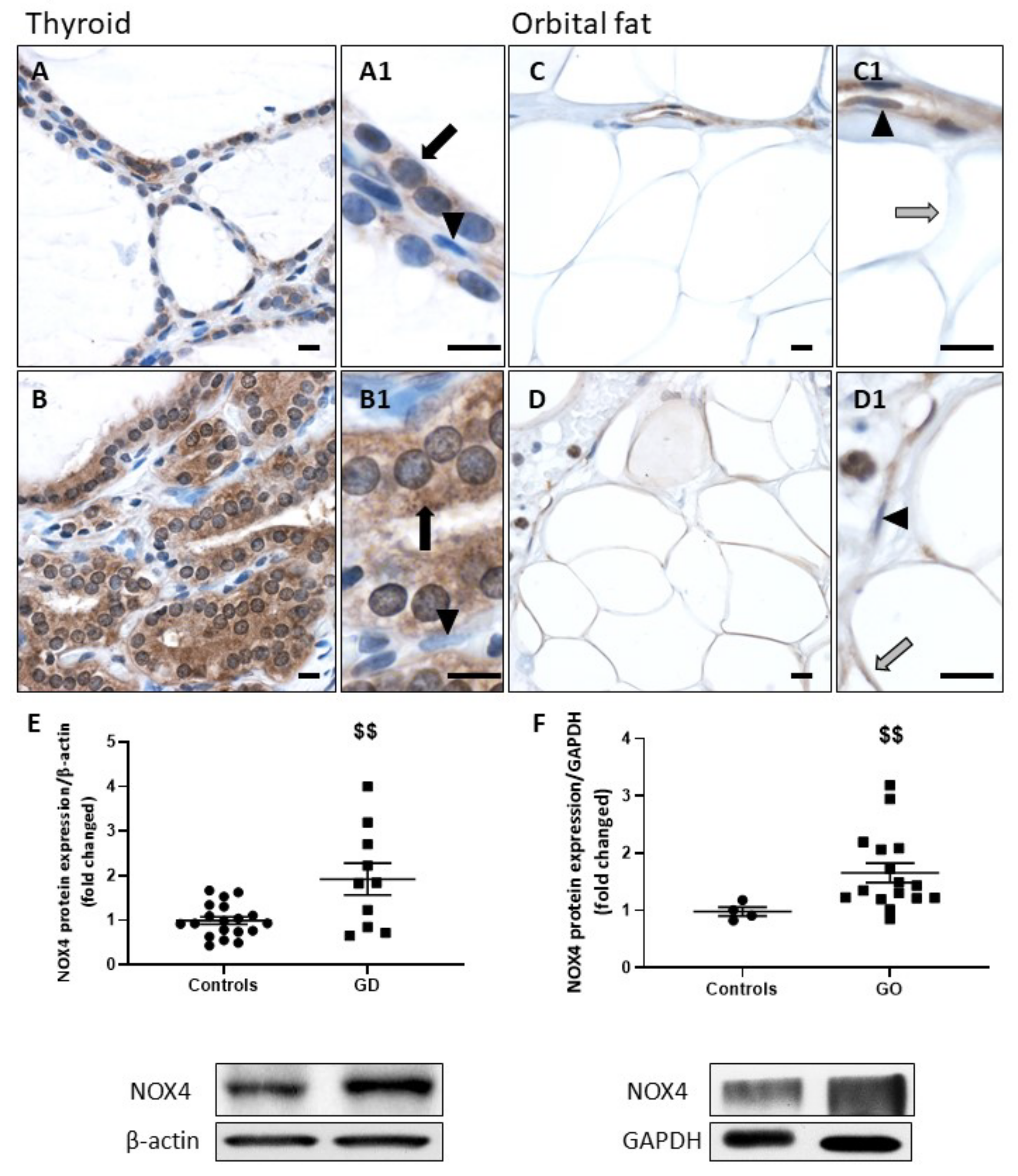
2.1. NOX4 Protein Expression Is Increased in GD and GO Tissues
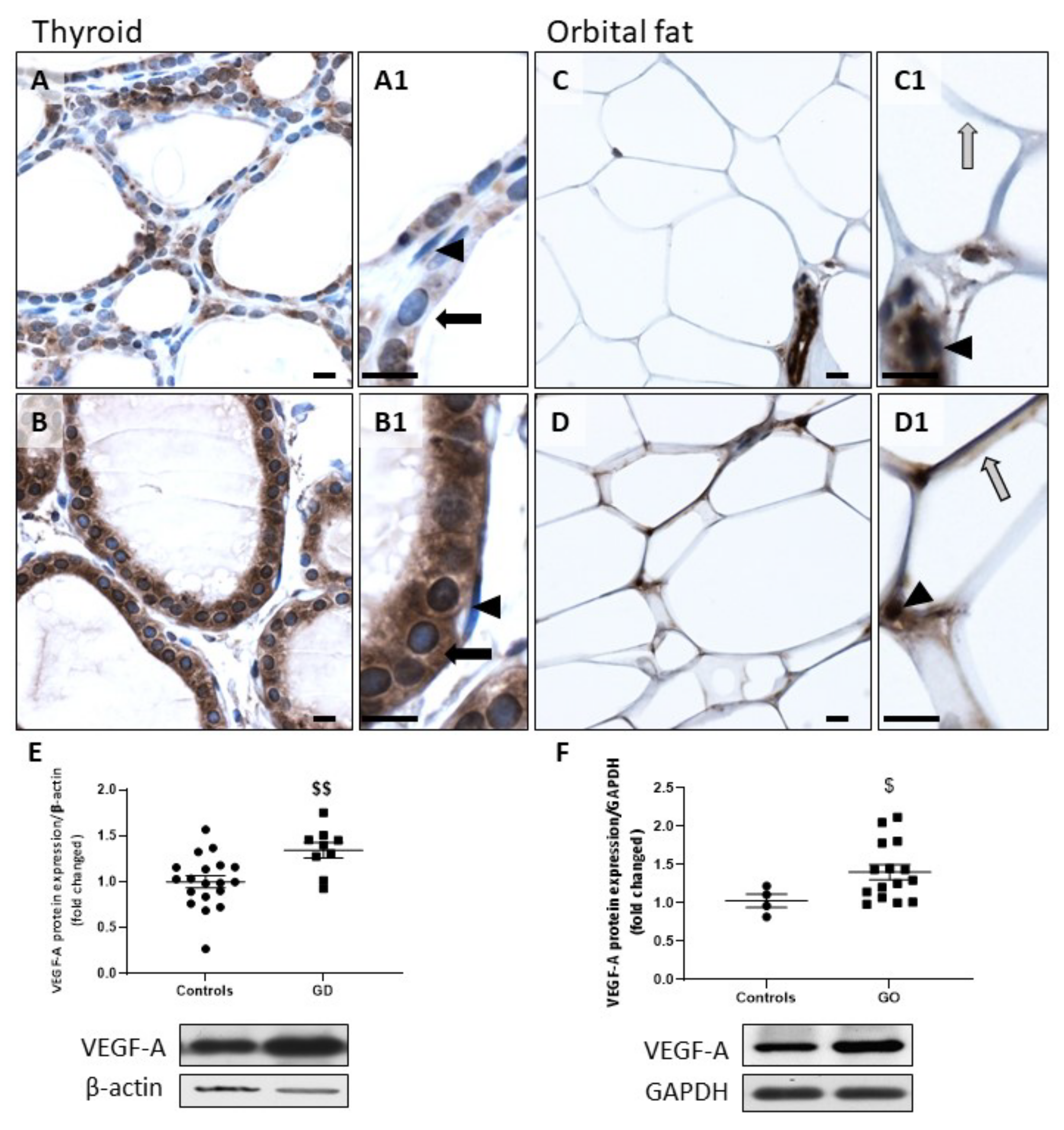
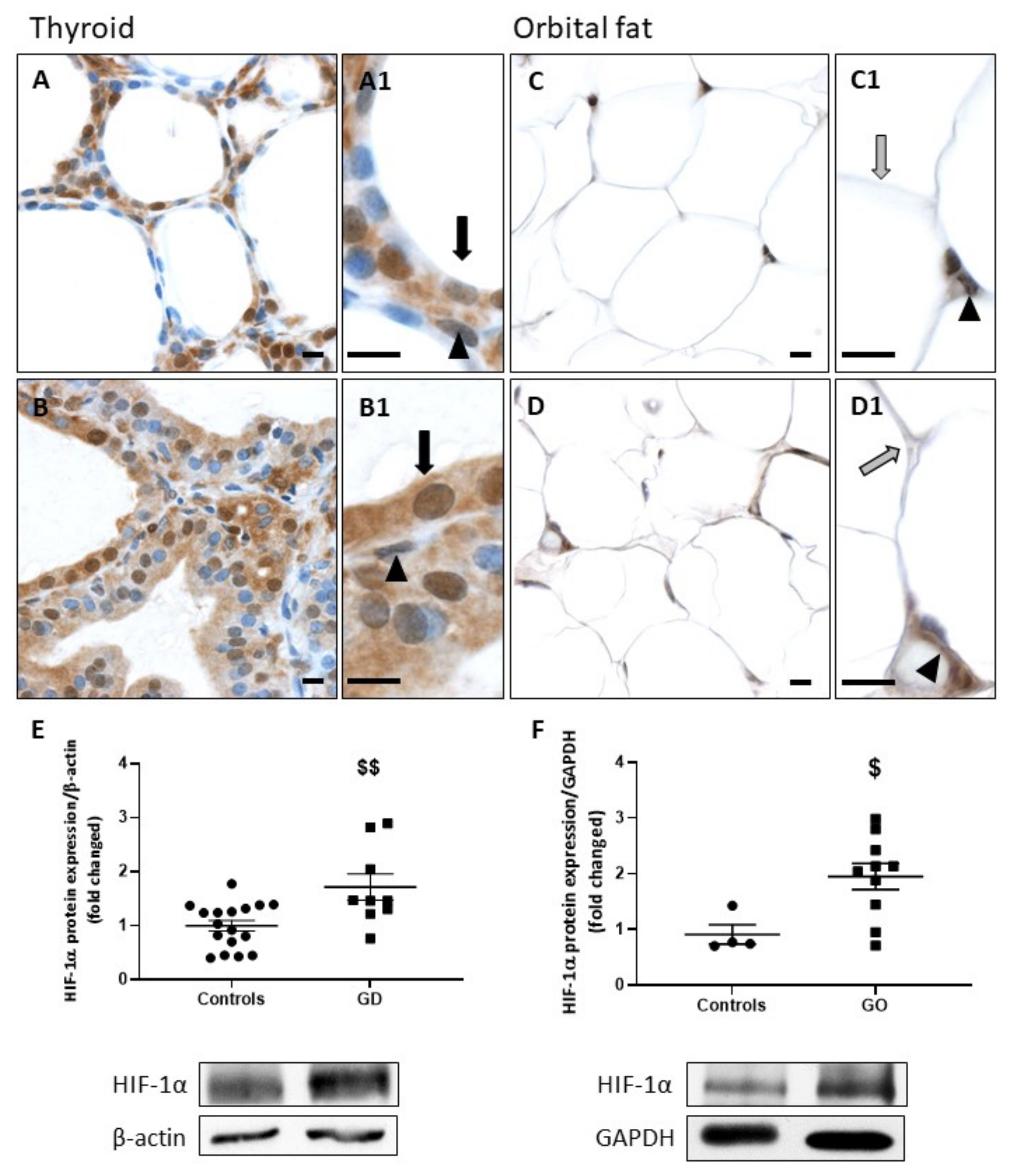
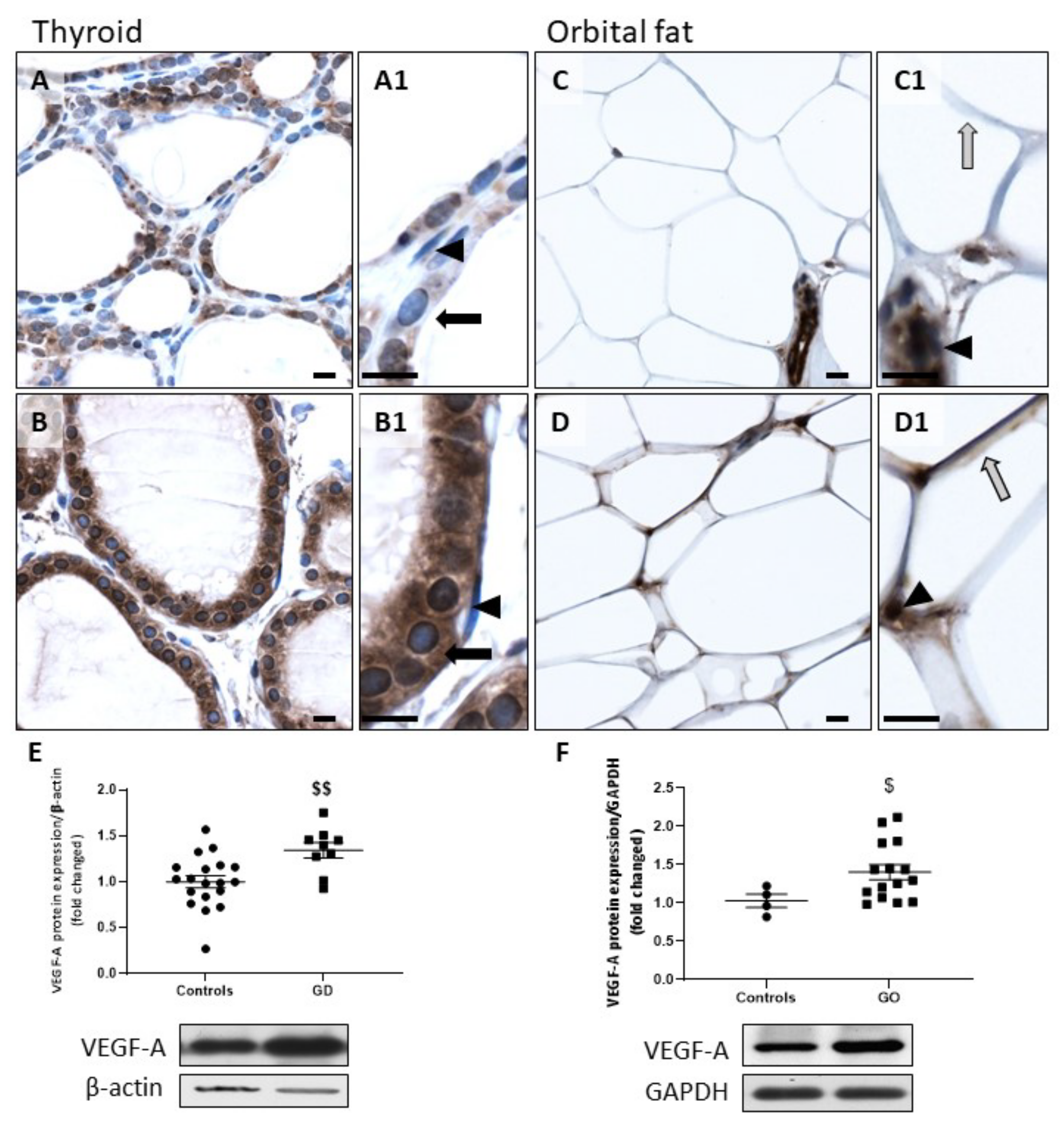
2.2. HIF-1α and VEGF-A Protein Expression Is Increased in GD and GO Tissues
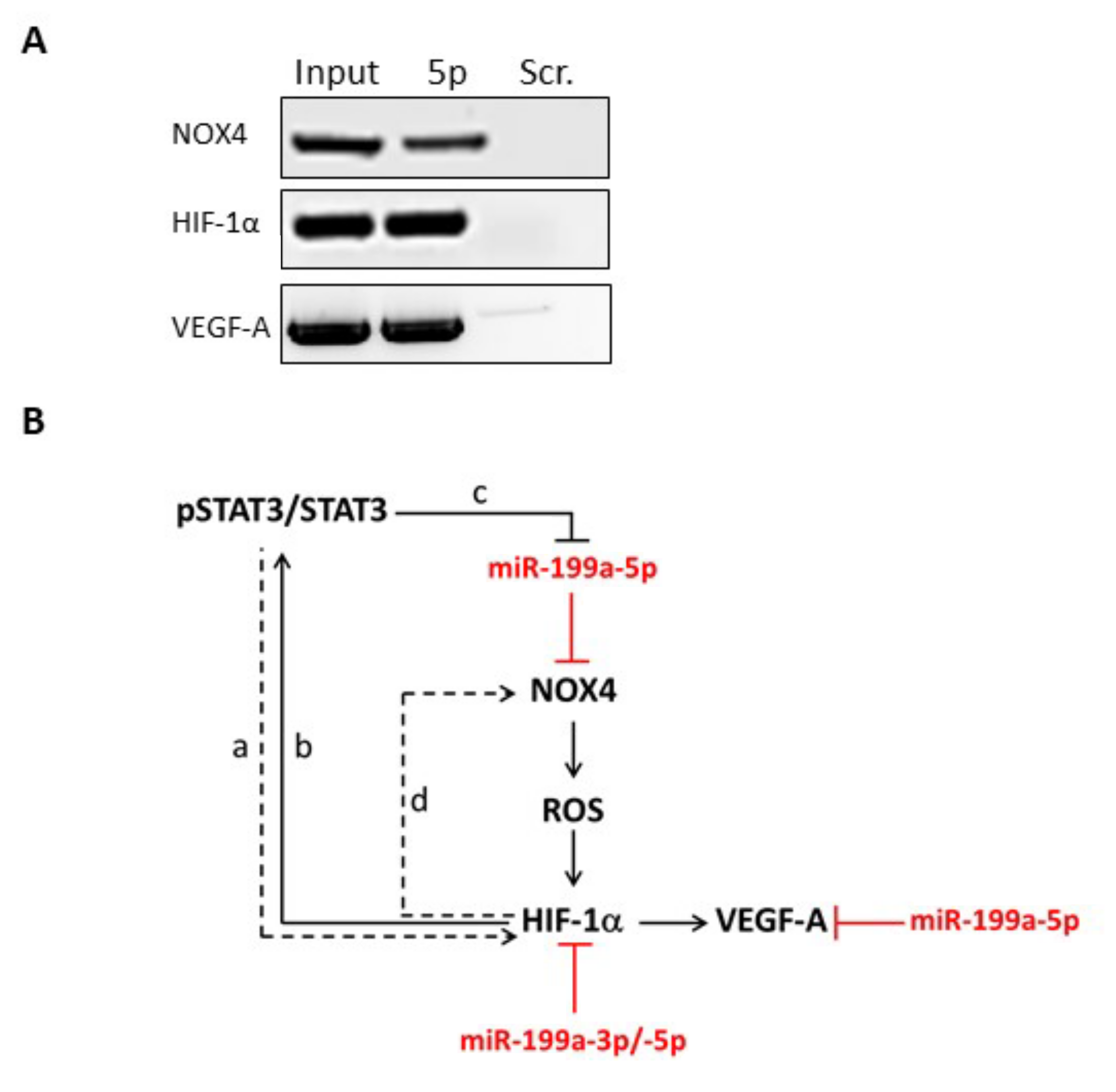
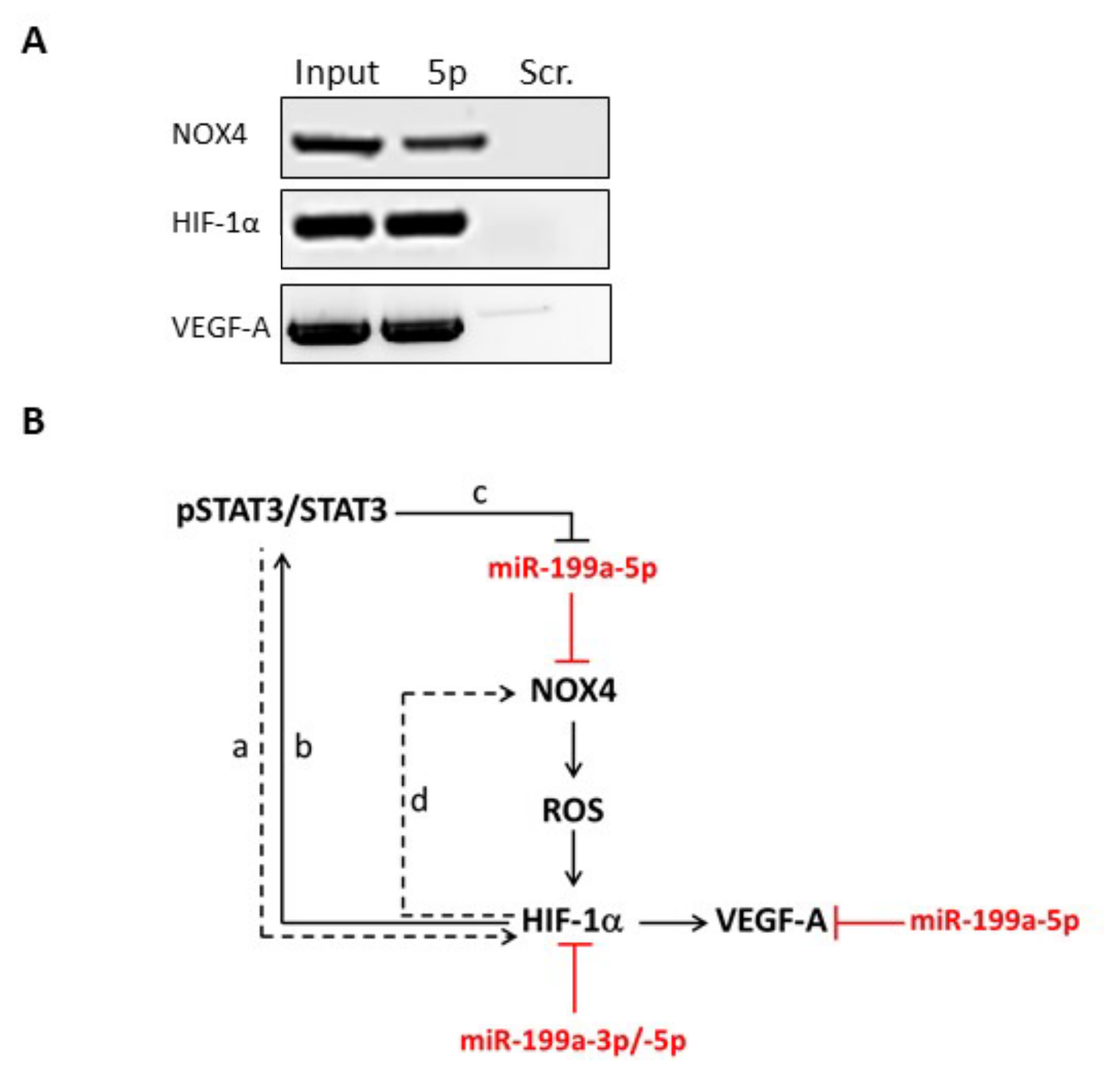
2.3. NOX4, HIF-1α and VEGF-A mRNAs Are Direct Targets of miR-199a-5p in Thyrocytes
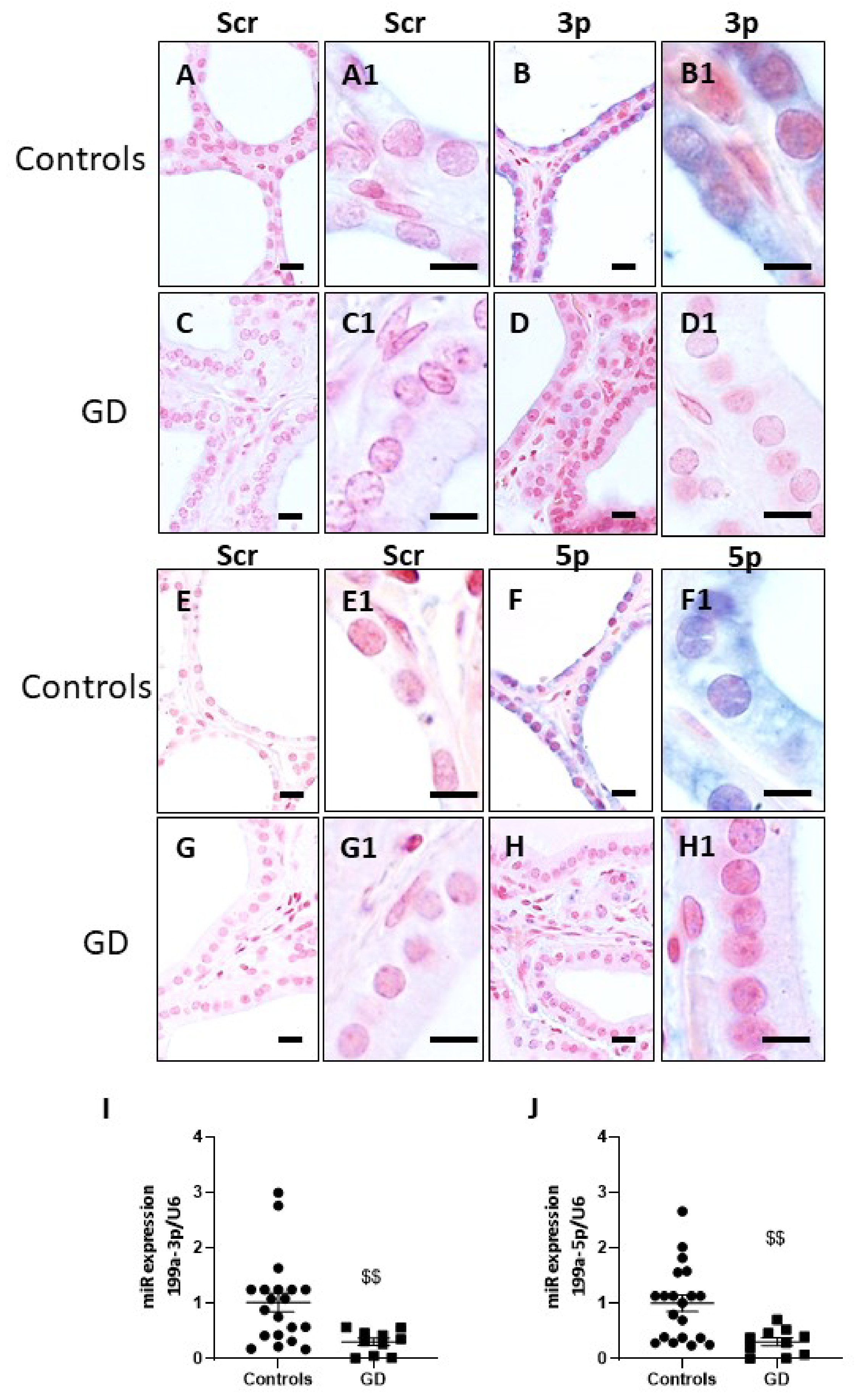
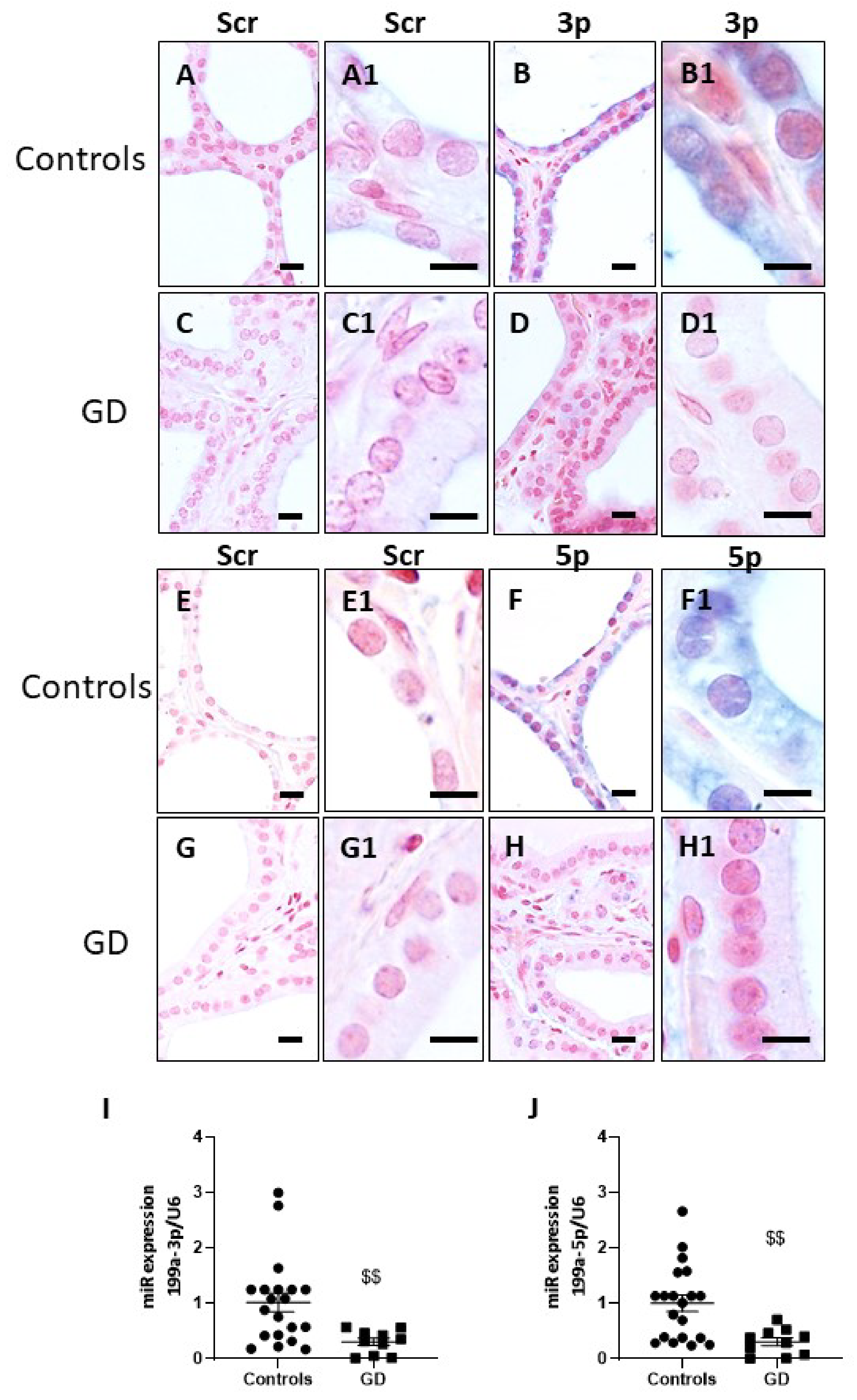
2.4. miR-199a-3p and miR-199a-5p Are Downregulated in GD Thyroids
2.5. A Higher Expression of VEGF-A and a Downregulation of miR-199a-3p and miR-199a-5p Are Observed in Human Primary Thyrocytes Treated with IL-4
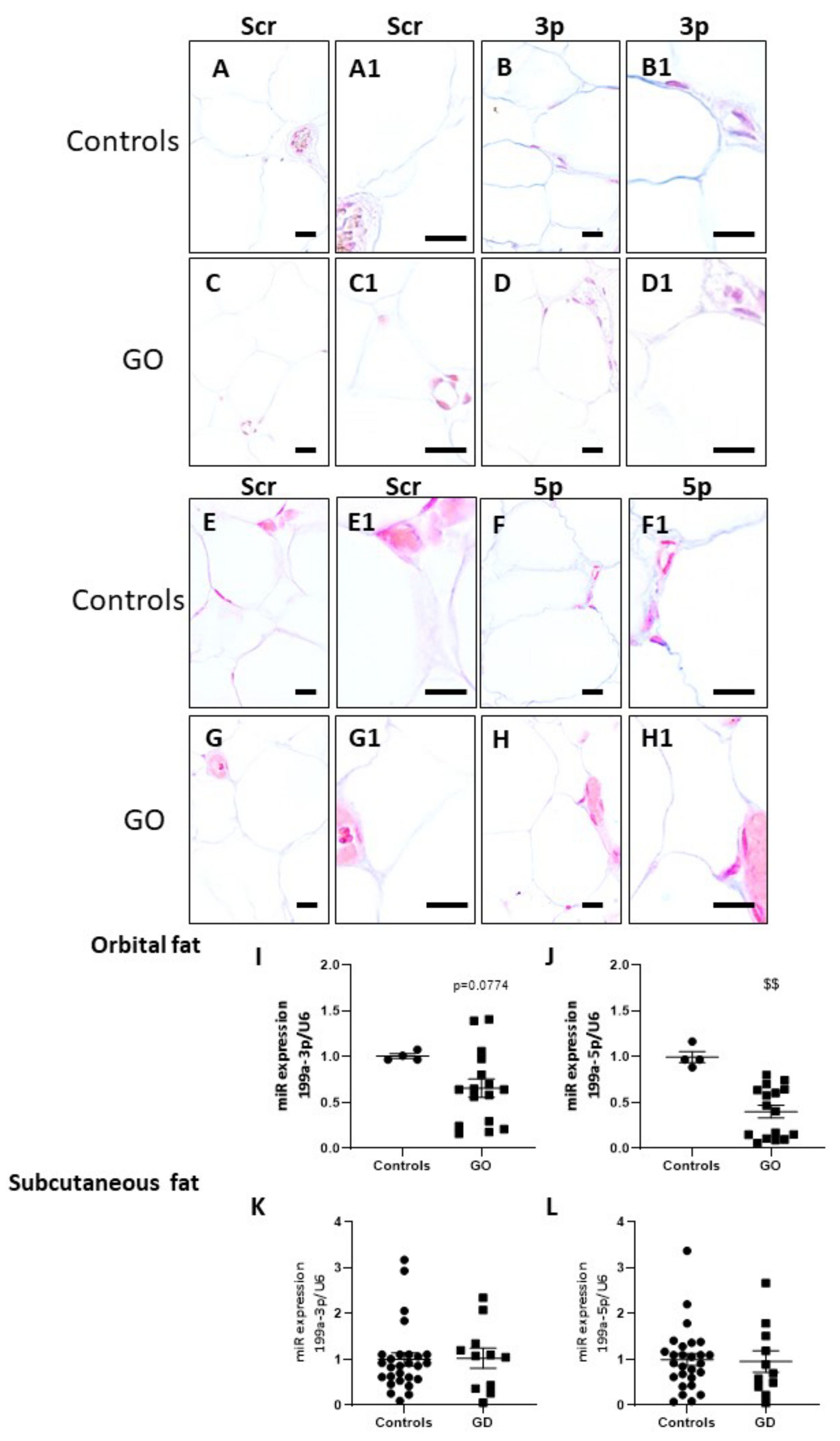
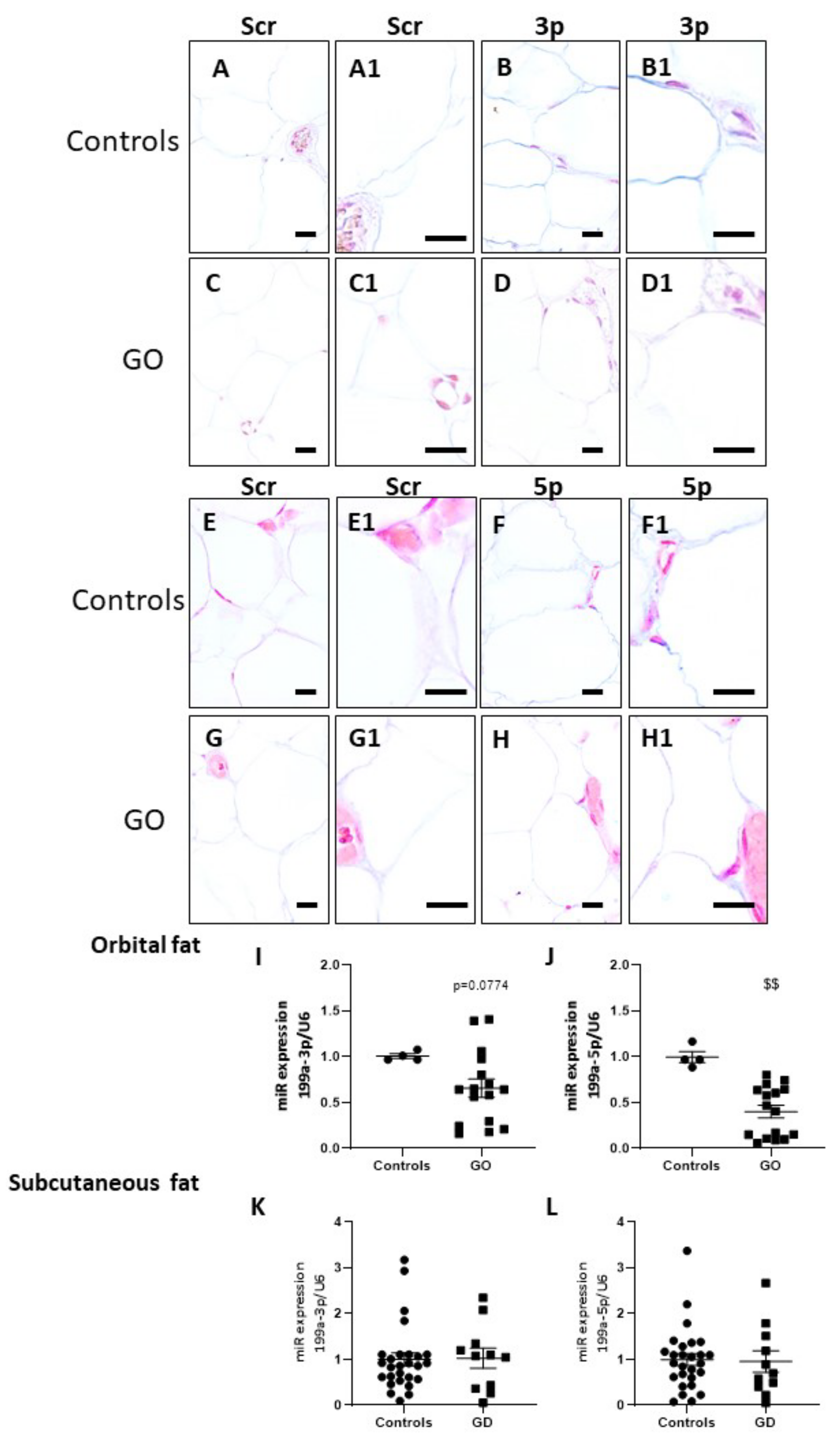
2.6. miR-199a-3p and miR-199a-5p Are Downregulated in GO Orbital Fat Tissues and in Plasma of GD Patients
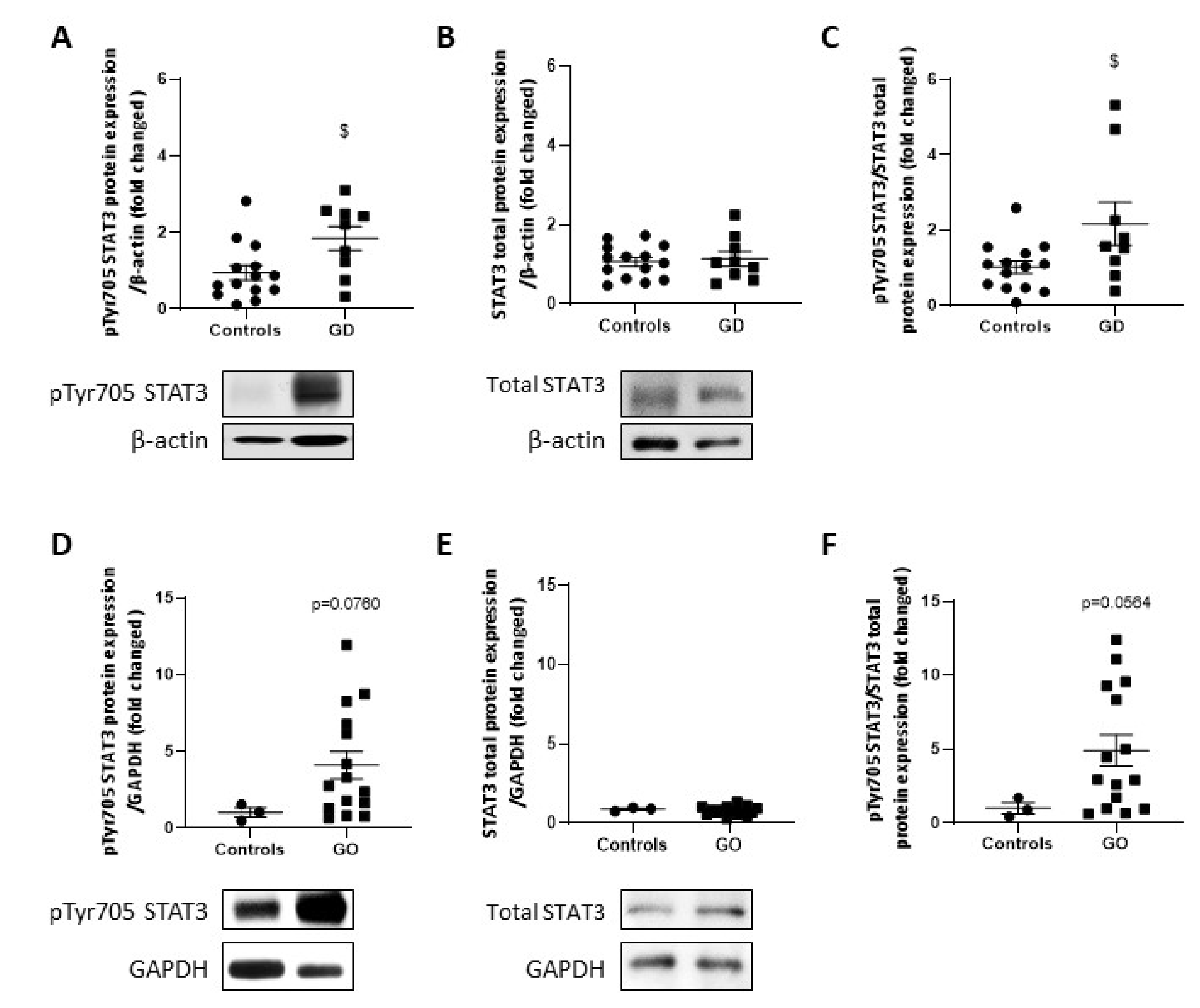
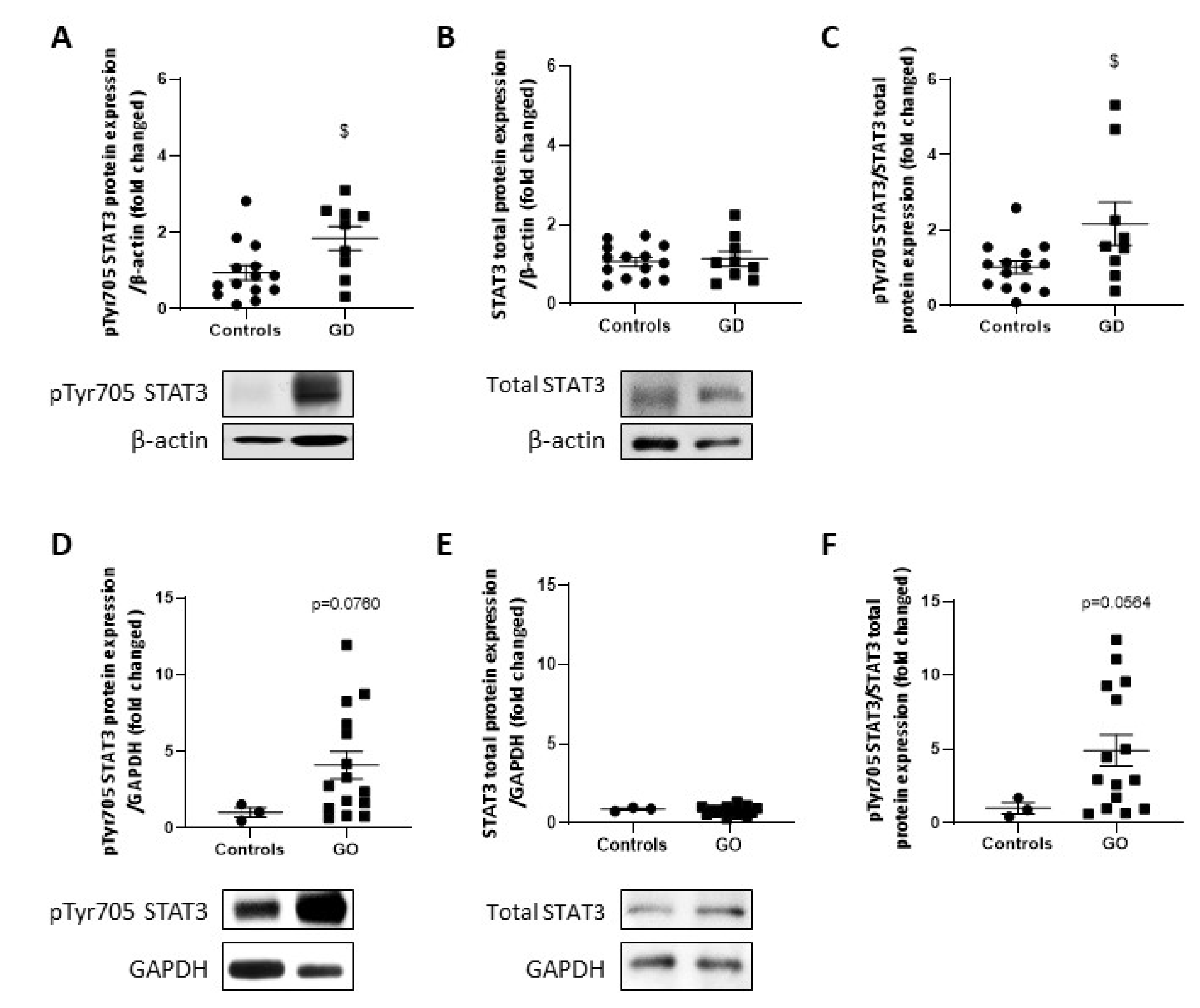
2.7. STAT3 Activity Is Upregulated in GD Thyroids and GO Orbital Fats
3. Discussion
4. Patients and Methods
4.1. Patients
4.2. Human Thyroid Cell Cultures
4.3. Transfection
4.4. Maxwell RNA Isolation
4.5. microRNA Reverse Transcription and Quantitative Real-Time PCR (qRT-PCR)
4.6. Western Blot Analysis
4.7. Immunohistochemistry
4.8. In Situ Hybridization
4.9. Biotin Pull-Down, Reverse Transcription, and Qualitative PCR
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AKT | protein kinase B |
| Cav-1 | Caveolin-1 |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| GD | Graves’ Disease |
| GO | Graves’ Orbitopathy |
| HIF | Hypoxia-inducible-factor |
| HT | Hashimoto’s thyroiditis |
| IHC | Immunohistochemistry |
| IL | Interleukin |
| miRs | microRNAs |
| NOX | NADPH-oxidase |
| OS | Oxidative stress |
| O2.- | Superoxide anion |
| PI3K | Phosphatidylinositol 3,4,5 triphosphate kinase |
| PLC | Phospholipase C |
| PRDX | Peroxiredoxin |
| PTC | Papillary thyroid cancers |
| ROS | Reactive oxygen species |
| SEM | Standard error of the mean |
| SOD | Superoxide dismutase |
| STAT3 | Signal transducer and activator of transcription 3 |
| Th2 | T helper type two |
| TRAb | Thyrotropin receptor-specific stimulatory autoantibodies |
| TSH | Thyrotropin |
| VEGF | Vascular endothelial growth factor |
| WB | Western Blot |
References
- Maheshwari, R.; Weis, E. Thyroid associated orbitopathy. Indian J. Ophthalmol. 2012, 60, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Shanmuganathan, T.; Girgis, C.; Lahooti, H.; Champion, B.; Wall, J.R. Does autoimmunity against thyroglobulin play a role in the pathogenesis of Graves’ ophthalmopathy: A review. Clin. Ophthalmol. 2015, 9, 2271–2276. [Google Scholar] [PubMed] [Green Version]
- Lanzolla, G.; Marcocci, C.; Marinò, M. Oxidative Stress in Graves’ Disease and Graves’ Orbitopathy. Eur. Thyroid. J. 2012, 9, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Marique, L.; Senou, M.; Craps, J.; Delaigle, A.; Van Regemorter, E.; Werion, A.; Van Regemorter, V.; Mourad, M.; Nyssen-Behets, C.; Lengele, B.; et al. Oxidative Stress and Upregulation of Antioxidant Proteins, Including Adiponectin, in Extraocular Muscular Cells, Orbital Adipocytes, and Thyrocytes in Graves’ Disease Associated with Orbitopathy. Thyroid 2015, 25, 1033–1042. [Google Scholar] [CrossRef]
- Iitaka, M.; Miura, S.; Yamanaka, K.; Kawasaki, S.; Kitahama, S.; Kawakami, Y.; Kakinuma, S.; Oosuga, I.; Wada, S.; Katayama, S. Increased Serum Vascular Endothelial Growth Factor Levels and Intrathyroidal Vascular Area in Patients with Graves’ Disease and Hashimoto’s Thyroiditis. J. Clin. Endocrinol. Metab. 1998, 83, 3908–3912. [Google Scholar] [CrossRef] [Green Version]
- Tseleni-Balafouta, S.; Kavantzas, N.; Balafoutas, D.; Patsouris, E. Comparative study of angiogenesis in thyroid glands with Graves’ disease and Hashimoto’s thyroiditis. Appl. Immunohistochem. Mol. Morphol. 2006, 14, 203–207. [Google Scholar] [CrossRef]
- Wong, L.L.; Lee, N.G.; Amarnani, D.; Choi, C.J.; Bielenberg, D.R.; Freitag, S.K.; D’Amore, P.A.; Kim, L.A. Orbital Angiogenesis and Lymphangiogenesis in Thyroid Eye Disease: An Analysis of Vascular Growth Factors with Clinical Correlation. Ophthalmology 2016, 123, 2028–2036. [Google Scholar] [CrossRef] [Green Version]
- Guerra, L.N.; Moiguer, S.; Karner, M.; de Molina, M.C.; Sreider, C.M.; Burdman, J.A. Antioxidants in the treatment of Graves’ disease. IUBMB Life 2001, 51, 105–109. [Google Scholar] [CrossRef]
- Karbownik, M.; Lewinski, A. The role of oxidative stress in physiological and pathological processes in the thyroid gland; possible involvement in pineal-thyroid interactions. Neuro Endocrinol. Lett. 2003, 24, 293–303. [Google Scholar]
- Poncin, S.; Gérard, A.-C.; Boucquey, M.; Senou, M.; Calderon, P.B.; Knoops, B.; Lengelé, B.; Many, M.-C.; Colin, I. Oxidative Stress in the Thyroid Gland: From Harmlessness to Hazard Depending on the Iodine Content. Endocrinology 2007, 149, 424–433. [Google Scholar] [CrossRef] [Green Version]
- Marcocci, C.; Leo, M.; Altea, M.A. Oxidative Stress in Graves’ Disease. Eur. Thyroid. J. 2012, 1, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Marique, L.; Van Regemorter, V.; Gérard, A.C.; Craps, J.; Senou, M.; Marbaix, E.; Rahier, J.; Daumerie, C.; Mourad, M.; Lengelé, B.; et al. The expression of dual oxidase, thyroid peroxidase, and caveolin-1 differs according to the type of immune response (TH1/TH2) involved in thyroid autoimmune disorders. J. Clin. Endocrinol. Metab. 2014, 99, 1722–1732. [Google Scholar] [CrossRef] [Green Version]
- Dupuy, C.; Ohayon, R.; Valent, A.; Noël-Hudson, M.-S.; Dème, D.; Virion, A. Purification of a novel flavoprotein involved in the thyroid NADPH oxidase: Cloning of the porcine and human cDNAs. J. Biol. Chem. 1999, 274, 37265–37269. [Google Scholar] [CrossRef] [Green Version]
- De Deken, X.; Wang, D.; Dumont, J.E.; Miot, F. Characterization of ThOX proteins as components of the thyroid H2O2-generating system. Exp. Cell Res. 2002, 273, 187–196. [Google Scholar] [CrossRef]
- Weyemi, U.; Caillou, B.; Talbot, M.; Ameziane-El-Hassani, R.; Lacroix, L.; Lagent-Chevallier, O.; Al Ghuzlan, A.; Roos, D.; Bidart, J.-M.; Virion, A.; et al. Intracellular expression of reactive oxygen species-generating NADPH oxidase NOX4 in normal and cancer thyroid tissues. Endocr. Relat. Cancer 2010, 17, 27–37. [Google Scholar] [CrossRef]
- Ameziane-El-Hassani, R.; Schlumberger, M.; Dupuy, C. NADPH oxidases: New actors in thyroid cancer? Nat. Rev. Endocrinol. 2016, 12, 485–494. [Google Scholar] [CrossRef]
- El Hassani, R.A.; Buffet, C.; Leboulleux, S.; Dupuy, C. Oxidative stress in thyroid carcinomas: Biological and clinical significance. Endocr. Relat. Cancer 2019, 26, R131–R143. [Google Scholar] [CrossRef] [Green Version]
- Hepp, M.; Werion, A.; De Greef, A.; de Ville de Goyet, C.; de Bournonville, M.; Behets, C.; Lengelé, B.; Daumerie, C.; Mourad, M.; Ludgate, M.; et al. Oxidative Stress-Induced Sirtuin1 Downregulation Correlates to HIF-1, GLUT-1, and VEGF-A Upregulation in Th1 Autoimmune Hashimoto’s Thyroiditis. Int. J. Mol. Sci. 2021, 22, 3806. [Google Scholar] [CrossRef]
- Wang, B.; Shao, X.; Song, R.; Xu, D.; Zhang, J.A. The Emerging Role of Epigenetics in Autoimmune Thyroid Diseases. Front. Immunol. 2017, 8, 396. [Google Scholar] [CrossRef] [Green Version]
- Razmara, E.; Salehi, M.; Aslani, S.; Bitaraf, A.; Yousefi, H.; Colón, J.R.; Mahmoudi, M. Graves’ disease: Introducing new genetic and epigenetic contributors. J. Mol. Endocrinol. 2021, 66, R33–R55. [Google Scholar] [CrossRef]
- Qin, Q.; Wang, X.; Yan, N.; Song, R.; Cai, T.-T.; Zhang, W.; Guan, L.-J.; Muhali, F.-S.; Zhang, J.-A. Aberrant Expression of miRNA and mRNAs in Lesioned Tissues of Graves’ Disease. Cell. Physiol. Biochem. 2015, 35, 1934–1942. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, S.-J.; Yao, X.; Tian, D.-M.; Zhang, K.-Q.; She, D.-M.; Guo, F.-F.; Zhai, Q.-W.; Ying, H.; Xue, Y. Circulating microRNA-1a is a biomarker of Graves’ disease patients with atrial fibrillation. Endocrine 2017, 57, 125–137. [Google Scholar] [CrossRef]
- Li, J.; Cai, Y.; Sun, X.; Yao, D.; Xia, J. MiR-346 and TRAb as Predicative Factors for Relapse in Graves’ Disease Within One Year. Horm. Metab. Res. 2017, 49, 180–184. [Google Scholar] [CrossRef]
- Joris, V.; Gomez, E.L.; Menchi, L.; Lobysheva, I.; Di Mauro, V.; Esfahani, H.; Condorelli, G.; Balligand, J.L.; Catalucci, D.; Dessy, C. MicroRNA-199a-3p and MicroRNA-199a-5p Take Part to a Redundant Network of Regulation of the NOS (NO Synthase)/NO Pathway in the Endothelium. Arterioscler. Thromb. Vasc. Biol. 2018, 3, 2345–2357. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Cao, Y.; Cui, X.; Wang, X.; Zhou, Y.; Huang, F.; Wang, X.; Wen, J.; Xie, K.; Xu, P.; et al. Mir-199a-3p regulates brown adipocyte differentiation through mTOR signaling pathway. Mol. Cell. Endocrinol. 2018, 476, 155–164. [Google Scholar] [CrossRef]
- Shi, X.-E.; Li, Y.-F.; Jia, L.; Ji, H.-L.; Song, Z.-Y.; Cheng, J.; Wu, G.-F.; Song, C.-C.; Zhang, Q.-L.; Zhu, J.-Y.; et al. MicroRNA-199a-5p Affects Porcine Preadipocyte Proliferation and Differentiation. Int. J. Mol. Sci. 2014, 15, 8526–8538. [Google Scholar] [CrossRef] [Green Version]
- Minna, E.; Romeo, P.; de Cecco, L.; Dugo, M.; Cassinelli, G.; Pilotti, S.; Degl’Innocenti, D.; Lanzi, C.; Casalini, P.; Pierotti, M.A.; et al. Mir-199a-3p displays tumor suppressor functions in papillary thyroid carcinoma. Oncotarget 2014, 5, 2513–2528. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Han, S.; Liu, C.; Zhou, R.; Sun, W.; Zhang, Z.; Qu, J. Microrna-199a-5p Functions as a Tumor Suppressor via Suppressing Connective Tissue Growth Factor (CTGF) in Follicular Thyroid Carcinoma. Med. Sci. Monit. 2016, 22, 1210–2117. [Google Scholar] [CrossRef]
- Görtz, G.-E.; Horstmann, M.; Aniol, B.; Reyes, B.D.; Fandrey, J.; Eckstein, A.; Berchner-Pfannschmidt, U. Hypoxia-Dependent HIF-1 Activation Impacts on Tissue Remodeling in Graves’ Ophthalmopathy—Implications for Smoking. J. Clin. Endocrinol. Metab. 2016, 101, 4834–4842. [Google Scholar] [CrossRef]
- Craps, J.; Wilvers, C.; Joris, V.; De Jongh, B.; Vanderstraeten, J.; Lobysheva, I.; Balligand, J.-L.; Sonveaux, P.; Gilon, P.; Many, M.-C.; et al. Involvement of Nitric Oxide in Iodine Deficiency-Induced Microvascular Remodeling in the Thyroid Gland: Role of Nitric Oxide Synthase 3 and Ryanodine Receptors. Endocrinology 2014, 156, 707–720. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; He, L.; Zuo, D.; He, W.; Wang, Y.; Zhang, Y.; Liu, W.; Yuan, Y. Mutual Regulation of MiR-199a-5p and HIF-1α Modulates the Warburg Effect in Hepatocellular Carcinoma. J. Cancer 2017, 8, 940–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rane, S.; He, M.; Sayed, D.; Vashistha, H.; Malhotra, A.; Sadoshima, J.; Vatner, D.E.; Vatner, S.F.; Abdellatif, M. Downregulation of miR-199a Derepresses Hypoxia-Inducible Factor-1α and Sirtuin 1 and Recapitulates Hypoxia Preconditioning in Cardiac Myocytes. Circ. Res. 2009, 104, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-Y.; Hsieh, T.-H.; Tsai, C.-F.; Tsai, H.-P.; Chen, H.-S.; Chang, Y.; Chuang, H.-Y.; Lee, J.-N.; Hsu, Y.-L.; Tsai, E.-M. miRNA-199a-5p regulates VEGFA in endometrial mesenchymal stem cells and contributes to the pathogenesis of endometriosis. J. Pathol. 2013, 232, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Diebold, I.; Petry, A.; Hess, J.; Görlach, A. The NADPH Oxidase Subunit NOX4 Is a New Target Gene of the Hypoxia-inducible Factor-1. Mol. Biol. Cell 2010, 21, 2087–2096. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.E.; Lee, H.-G.; Cho, I.-H.; Chung, D.H.; Yoon, S.-H.; Yang, Y.M.; Lee, J.W.; Choi, S.; Park, J.-W.; Ye, S.-K.; et al. STAT3 is a potential modulator of HIF-1-mediated VEGF expression in human renal carcinoma cells. FASEB J. 2005, 19, 1296–1298. [Google Scholar] [CrossRef]
- Yang, X.; Zheng, Y.; Tan, J.; Tian, R.; Shen, P.; Cai, W.; Liao, H. MiR-199a-5p–HIF-1α-STAT3 Positive Feedback Loop Contributes to the Progression of Non-Small Cell Lung Cancer. Front. Cell Dev. Biol. 2021, 8, 620615. [Google Scholar] [CrossRef]
- Haghikia, A.; Missol-Kolka, E.; Tsikas, D.; Venturini, L.; Brundiers, S.; Castoldi, M.; Muckenthaler, M.U.; Eder, M.; Stapel, B.; Thum, T.; et al. Signal transducer and activator of transcription 3-mediated regulation of miR-199a-5p links cardiomyocyte and endothelial cell function in the heart: A key role for ubiquitin-conjugating enzymes. Eur. Heart J. 2011, 32, 1287–1297. [Google Scholar] [CrossRef]
- De Deken, X.; Wang, D.; Many, M.C.; Costagliola, S.; Libert, F.; Vassart, G.; Dumont, J.E.; Miot, F. Cloning of two human thyroid cDNAs encoding new members of the NADPH oxidase family. J. Biol. Chem. 2000, 275, 23227–23233. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, D.; Dupuy, C. Role of the NADPH Oxidases DUOX and NOX4 in Thyroid Oxidative Stress. Eur. Thyroid J. 2013, 2, 160–167. [Google Scholar] [CrossRef] [Green Version]
- Gérard, A.-C.; Many, M.-C.; Daumerie, C.; Knoops, B.; Colin, I. Peroxiredoxin 5 Expression in the Human Thyroid Gland. Thyroid 2005, 15, 205–209. [Google Scholar] [CrossRef]
- Abalovich, M.; Llesuy, S.; Gutierrez, S.; Repetto, M. Peripheral parameters of oxidative stress in Graves’ disease: The effects of methimazole and 131 iodine treatments. Clin. Endocrinol. 2003, 59, 321–327. [Google Scholar] [CrossRef]
- Diana, T.; Daiber, A.; Oelze, M.; Neumann, S.; Olivo, P.D.; Kanitz, M.; Stamm, P.; Kahaly, G.J. Stimulatory TSH-Receptor Antibodies and Oxidative Stress in Graves’ Disease. J. Clin. Endocrinol. Metab. 2018, 103, 3668–3677. [Google Scholar] [CrossRef] [Green Version]
- Weyemi, U.; Lagente chevallier, O.; Boufraqech, M.; Prenois, F.; Courtin, F.; Caillou, B.; Talbot, M.R.; Dardalhon, M.; Al Ghuzlan, A.; Bidart, J.-M.; et al. ROS-generating NADPH oxidase NOX4 is a critical mediator in oncogenic H-Ras-induced DNA damage and subsequent senescence. Oncogene 2011, 31, 1117–1129. [Google Scholar] [CrossRef] [Green Version]
- Van Regemorter, E.; Joris, V.; Van Regemorter, V.; Marique, L.; Behets, C.; Lengelé, B.; Boschi, A.; Baldeschi, L.; Daumerie, C.; Many, M.C.; et al. Downregulation of Caveolin-1 and Upregulation of Deiodinase 3, Associated with Hypoxia-Inducible Factor-1 Increase, Are Involved in the Oxidative Stress of Graves’ Orbital Adipocytes. Thyroid 2020, 31, 627–637. [Google Scholar] [CrossRef]
- Mouche, S.; Ben Mkaddem, S.; Wang, W.; Katic, M.; Tseng, Y.-H.; Carnesecchi, S.; Steger, K.; Foti, M.; Meier, C.A.; Muzzin, P.; et al. Reduced expression of the NADPH oxidase NOX4 is a hallmark of adipocyte differentiation. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2007, 1773, 1015–1027. [Google Scholar] [CrossRef]
- Schröder, K.; Wandzioch, K.; Helmcke, I.; Brandes, R.P. Nox4 acts as a switch between differentiation and proliferation in preadipocytes. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 239–245. [Google Scholar] [CrossRef] [Green Version]
- DeVallance, E.; Li, Y.; Jurczak, M.J.; Cifuentes-Pagano, E.; Pagano, P.J. The Role of NADPH Oxidases in the Etiology of Obesity and Metabolic Syndrome: Contribution of Individual Isoforms and Cell Biology. Antioxid. Redox Signal. 2019, 31, 687–709. [Google Scholar] [CrossRef]
- Colin, I.M.; Denef, J.-F.; Lengelé, B.; Many, M.-C.; Gérard, A.-C. Recent Insights into the Cell Biology of Thyroid Angiofollicular Units. Endocr. Rev. 2013, 34, 209–238. [Google Scholar] [CrossRef] [Green Version]
- Patten, D.A.; Lafleur, V.N.; Robitaille, G.A.; Chan, D.A.; Giaccia, A.J.; Richard, D.E. Hypoxia-inducible Factor-1 Activation in Nonhypoxic Conditions: The Essential Role of Mitochondrial-derived Reactive Oxygen Species. Mol. Biol. Cell 2010, 21, 3247–3257. [Google Scholar] [CrossRef] [Green Version]
- Bonello, S.; Zähringer, C.; BelAiba, R.S.; Djordjevic, T.; Hess, J.; Michiels, C.; Kietzmann, T.; Gärlach, A. Reactive oxygen species activate the HIF-1alpha promoter via a functional NFkappaB site. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 755–761. [Google Scholar] [CrossRef] [Green Version]
- Diebold, I.; Petry, A.; Sabrane, K.; Djordjevic, T.; Hess, J.; Görlach, A. The HIF1 target gene NOX2 promotes angiogenesis through urotensin-II. J. Cell Sci. 2012, 125, 956–964. [Google Scholar] [CrossRef] [Green Version]
- Szanto, I.; Pusztaszeri, M.; Mavromati, M. H2O2 Metabolism in Normal Thyroid Cells and in Thyroid Tumorigenesis: Focus on NADPH Oxidases. Antioxidants 2019, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Gérard, A.-C.; Poncin, S.; Audinot, J.-N.; Denef, J.-F.; Colin, I.M. Iodide deficiency-induced angiogenic stimulus in the thyroid occurs via HIF- and ROS-dependent VEGF-A secretion from thyrocytes. Am. J. Physiol. Metab. 2009, 296, E1414–E1422. [Google Scholar] [CrossRef]
- Zhou, Y.; Pang, B.; Xiao, Y.; Zhou, S.; He, B.; Zhang, F.; Liu, W.; Peng, H.; Li, P. The protective microRNA-199a-5p-mediated unfolded protein response in hypoxic cardiomyocytes is regulated by STAT3 pathway. J. Physiol. Biochem. 2018, 75, 73–81. [Google Scholar] [CrossRef]
- Ko, J.; Kim, J.-Y.; Kim, B.R.; Lee, E.J.; Kikkawa, D.O.; Yoon, J.S. Signal transducer and activator of transcription 3 as a potential therapeutic target for Graves’ orbitopathy. Mol. Cell. Endocrinol. 2021, 534, 111363. [Google Scholar] [CrossRef]
- Frohlich, E.; Wahl, R. Thyroid Autoimmunity: Role of Anti-thyroid Antibodies in Thyroid and Extra-Thyroidal Diseases. Front. Immunol. 2017, 8, 521. [Google Scholar] [CrossRef]
- Matà, R.; Palladino, C.; Nicolosi, M.L.; Presti, A.R.L.; Malaguarnera, R.; Ragusa, M.; Sciortino, D.; Morrione, A.; Maggiolini, M.; Vella, V.; et al. IGF-I induces upregulation of DDR1 collagen receptor in breast cancer cells by suppressing MIR-199a-5p through the PI3K/AKT pathway. Oncotarget 2016, 7, 7683–7700. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.; Chen, L.; Chen, B.; Lu, K.; Belguise, K.; Wang, X.; Yi, B. MicroRNA-199a-5p Regulates the Proliferation of Pulmonary Microvascular Endothelial Cells in Hepatopulmonary Syndrome. Cell Physiol. Biochem. 2015, 37, 1289–1300. [Google Scholar] [CrossRef]
- Lino Cardenas, C.L.; Henaoui, I.S.; Courcot, E.; Roderburg, C.; Cauffiez, C.; Aubert, S.; Copin, M.C.; Wallaert, B.; Glowacki, F.; Dewaeles, E.; et al. Mir-199a-5p Is upregulated during fibrogenic response to tissue injury and mediates TGFbeta-induced lung fibroblast activation by targeting caveolin-1. PLoS Genet. 2013, 9, e1003291. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Lin, X.; Shan, S.K.; Li, F.; Xu, F.; Zhong, J.Y.; Guo, B.; Zheng, M.H.; Wang, Y.; Mo, Z.H.; et al. The Suppression of miR-199a-3p by Promoter Methylation Contributes to Papillary Thyroid Carcinoma Aggressiveness by Targeting RAP2a and DNMT3a. Front. Cell Dev. Biol. 2020, 8, 594528. [Google Scholar] [CrossRef]
- Liu, C.; Xing, M.; Wang, L.; Zhang, K. miR-199a-3p downregulation in thyroid tissues is associated with invasion and metastasis of papillary thyroid carcinoma. Br. J. Biomed. Sci. 2017, 74, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Aldred, M.A.; Ginn-Pease, M.E.; Morrison, C.D.; Popkie, A.P.; Gimm, O.; Hoang-Vu, C.; Krause, U.; Dralle, H.; Jhiang, S.M.; Plass, C.; et al. Caveolin-1 and caveolin-2, together with three bone morphogenetic protein-related genes, may encode novel tumor suppressors down-regulated in sporadic follicular thyroid carcinogenesis. Cancer Res. 2003, 63, 2864–2871. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of GD patients | n = 10 | |
| Mean (range) age | 34.4 (25–49) years | |
| Sex | Female/Male | n = 8 / n = 2 |
| Mean (range) GD duration | 52.2 (3–156) months | |
| Mean (range) TSH levels at biopsy time | 1.13 (<0.01–2.33) mU/mL | |
| Mean (range) TRAb levels at biopsy time | 49.7 (1.7–151) U/mL |
| Number of GO patients | n = 16 | |
| Mean (range) age | 52.0625 (24–75) years | |
| Sex | Female/Male | n = 12/n = 4 |
| Mean (range) GO duration | 47 (4–240) months | |
| Smoking habits | Smokers/Non-smokers | n = 10/n= 6 |
| Mean (range) TSH levels at biopsy time | 1.71 (0.005–3.15) mU/mL | |
| Mean (range) TRAb levels at biopsy time | 15.85 (0.3–33.5) U/L | |
| Corticotherapy | n = 8 | |
| Mean (range) steroid therapy discontinuation time before surgery | 5.5 (0–12) months |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Craps, J.; Joris, V.; Baldeschi, L.; Daumerie, C.; Camboni, A.; Buemi, A.; Lengelé, B.; Behets, C.; Boschi, A.; Mourad, M.; et al. miR-199a Downregulation as a Driver of the NOX4/HIF-1α/VEGF-A Pathway in Thyroid and Orbital Adipose Tissues from Graves′ Patients. Int. J. Mol. Sci. 2022, 23, 153. https://doi.org/10.3390/ijms23010153
Craps J, Joris V, Baldeschi L, Daumerie C, Camboni A, Buemi A, Lengelé B, Behets C, Boschi A, Mourad M, et al. miR-199a Downregulation as a Driver of the NOX4/HIF-1α/VEGF-A Pathway in Thyroid and Orbital Adipose Tissues from Graves′ Patients. International Journal of Molecular Sciences. 2022; 23(1):153. https://doi.org/10.3390/ijms23010153
Chicago/Turabian StyleCraps, Julie, Virginie Joris, Lelio Baldeschi, Chantal Daumerie, Alessandra Camboni, Antoine Buemi, Benoit Lengelé, Catherine Behets, Antonella Boschi, Michel Mourad, and et al. 2022. "miR-199a Downregulation as a Driver of the NOX4/HIF-1α/VEGF-A Pathway in Thyroid and Orbital Adipose Tissues from Graves′ Patients" International Journal of Molecular Sciences 23, no. 1: 153. https://doi.org/10.3390/ijms23010153