
Hydrogen Sulfide in Plants: Crosstalk with Other Signal Molecules in Response to Abiotic Stresses



College of Horticulture, Gansu Agricultural University, Lanzhou 730070, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(21), 12068; https://doi.org/10.3390/ijms222112068
Submission received: 4 September 2021
/
Revised: 3 November 2021
/
Accepted: 5 November 2021
/
Published: 8 November 2021
(This article belongs to the Special Issue Hydrogen Sulfide and Reactive Oxygen Species, Antioxidant Defense, Abiotic Stress Tolerance Mechanisms in Plants)
Abstract
:Hydrogen sulfide (H2S) has recently been considered as a crucial gaseous transmitter occupying extensive roles in physiological and biochemical processes throughout the life of plant species. Furthermore, plenty of achievements have been announced regarding H2S working in combination with other signal molecules to mitigate environmental damage, such as nitric oxide (NO), abscisic acid (ABA), calcium ion (Ca2+), hydrogen peroxide (H2O2), salicylic acid (SA), ethylene (ETH), jasmonic acid (JA), proline (Pro), and melatonin (MT). This review summarizes the current knowledge within the mechanism of H2S and the above signal compounds in response to abiotic stresses in plants, including maintaining cellular redox homeostasis, exchanging metal ion transport, regulating stomatal aperture, and altering gene expression and enzyme activities. The potential relationship between H2S and other signal transmitters is also proposed and discussed.
1. Introduction
Several abiotic stresses such as salt, drought, flooding, heat, cold, and freezing easily result in the loss of crop production and a drop in economy in the world. Furthermore, with ongoing industrialization and pesticides application, plants are more likely subjected to some abiotic stresses including salinity and heavy metal (aluminum (Al); cadmium (Cd); chromium (Cr); lead (Pb); cobalt (Co); arsenic (As); nickel (Ni)) stresses [1,2]. In order to survive, plants must make a series of adjustments in morphology and physiological and biochemical metabolism when they are subjected to abiotic stresses. There are many kinds of mechanisms for plants to respond to abiotic stresses, including plant hormones, osmotic regulators, active oxygen scavenging systems, genes, and proteins. When plants are subjected to adversity stress, a series of changes will occur in the hormone levels, thereby initiating or regulating certain physiological and biochemical processes related to stress resistance to complete the response to adversity. Moreover, some inorganic and organic osmotic substances such as Na+, K+, Cl−, proline (Pro), and soluble sugars may accumulate when plants encounter stresses. Further, under normal circumstances, the reactive oxygen species (ROS) are tightly controlled in plants, because plants have a reactive oxygen scavenging system, which keeps the production and removal of reactive oxygen species in a dynamic balance. This ROS includes hydrogen peroxide (H2O2), superoxide anion (O2·-), singlet oxygen (·O2), and hydroxyl radical (·OH) [3]. Under the condition of adversity, this balance is broken, and a large amount of active oxygen is produced. Active oxygen attacks the membrane system, causing changes in membrane lipid components and conformation of various enzymes on the membrane, loss of membrane selective permeability, leakage of electrolytes and certain small molecular organic substances, and disorder of mitochondria and chloroplast functions [1,4]. The active oxygen scavenging system mainly includes two types of substances: one is an enzymatic protection system composed of superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT), etc.; the other is non-enzymatic antioxidants including reduced glutathione (GSH), carotenoids (Car), vitamin E, and other antioxidants [5]. Last but not least, some proteins such as NAC, WRKY, basic region/leucine zipper motif (bZTP), and salt overly sensitive1 (SOS1) participate in plant response to abiotic stresses [2,6].
Hydrogen sulfide (H2S) is a colorless, combustible, and hydrosoluble gas with an obvious smell of rotten eggs, which has been widely considered as the third gasotransmitter molecule besides nitric oxide (NO) and carbon monoxide (CO) [7]. The emission of H2S was studied a long time ago. In 1978, Wilson et al. (1978) firstly observed the emission of H2S in the leaves of cucumber (Cucumis sativus L.), squash and pumpkin (Cucurbita pepo L.), cantaloupe (Cucumis melo L.), maize, soybean (Glycine max L. Merr), and cotton (Gossypium hirsutum L.) [8]. Current studies show that H2S can be biosynthesized through a variety of enzymes such as cysteine synthase (CS), β-cyanoalanine synthase (CAS), L-cysteine desulfhydrase (LCD), D-cysteine desulfhydrase (DCD), and sulfite reductase (SiR) in mitochondria, cytosol, and chloroplast [9,10]. In mitochondria, H2S can be produced by CAS in the course of cyanide detoxification. The generation of H2S mainly occurs by inducing the activities of LCD and DCD from cysteine (Cys) in the cytosol, which is also accompanied by the formation of pyruvate and ammonia. SiR is the reaction catalyst in the photosynthetic sulfate-assimilation pathway which induces the release of H2S in the chloroplast [11,12]. Thus, endogenous H2S can be produced under the catalysis of the corresponding enzymes [8,9,10,11]. The changes in endogenous H2S level can influence cellular metabolisms, enzyme activities, and gene expressions, and thus modulate plant growth and development [5,13]. Therefore, H2S is widely considered as a signaling molecule within organic cells.
In the last few decades, increasing evidence has shown that H2S plays a vital role in the treatment of diseases for animals and humans, including cancer [13], burns [14], neurodegenerative diseases [15], and inflammation [16]. In addition, it is involved in many processes of growth and development in plants. It can influence the seed germination, root organogenesis, photosynthesis, stomatal movement, leaf senescence, fruit ripening and nodulation, and nitrogen fixation [17]. H2S can also enhance the plant’s tolerance to diverse biotic and abiotic stresses, such as bacterial and fungal pathogens, salinity, drought, heat, hyperosmotic, oxidative and heavy metal stresses, etc. [5,17,18,19].
As a gaseous signaling molecule, H2S can interact with other signal molecules to influence the growth and development of, and respond to abiotic stresses in, plants. Plenty of research demonstrates that H2S is involved in NO-alleviated salt stress and heavy metal stresses in the seedling roots of pea (Pisum sativum L. cv. Azad P-1) and barley (Hordeum vulgare L.), as well as the seeds of alfalfa (Medicago sativa L. cv. Victoria) [20,21,22]. Besides, some plant hormones such as abscisic acid (ABA), salicylic acid (SA), ethylene (ETH), jasmonic acid (JA), and melatonin (MT) could alleviate abiotic stresses together with H2S in the process of plant growth and development. Some ionic signals such as calcium ion (Ca2+) and H2S are interrelated under stresses [23]. Meanwhile, H2O2 and proline (Pro) have been reported to have a relationship with H2S under abiotic stresses during the process of plant growth [17,24,25]. Here, we comprehensively review the crosstalk between H2S and other signal molecules in response to abiotic stresses. Also, new research directions and future prospects in this area will be discussed in this review (Figure 1).
2. Crosstalk between H2S and NO in Response to Abiotic Stresses
NO is widely recognized as a gas transmitter in the regulation of seed germination, dormancy, stomatal aperture, adventitious root development, and photosynthesis in plants [26,27]. NO also takes part in many stress alleviation processes, such as heavy metal, extreme temperature, drought, salt, and UV-B radiation [4,28]. Moreover, the relationship between H2S and NO under different stress conditions has been explored at both the physiological and molecular levels, which remains a hot topic in plant science research in recently years. The obtained achievements in this field were collected and shown below.
2.1. Crosstalk between H2S and NO in Response to Heavy Metal Stress
There is considerable research on how H2S and NO interplay with each other in plants under heavy metal stress. In pea seedlings, As (V) reduced growth, photosynthesis capacity, and nitrogen content [29]. An application of exogenous NaHS alleviated As (V) toxicity by inducing H2S and NO generation. These results suggest a vital role of H2S in As (V) stress tolerance. Also, exogenous H2S and NO could reduce the influence of Cr (VI) toxicity in maize (Zea mays L.) in a similar manner [30]. Furthermore, H2S donor NaHS and NO donor sodium nitroprusside (SNP), rather than other derivatives, were found to specifically ameliorate Cd-induced oxidative damage in the root tissues of alfalfa seedlings [31]. This work further confirms that both H2S and NO may participate in alleviating heavy metal stress. In addition, the alleviation effects of NaHS and SNP were reversed by NO scavenger 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide potassium salt (cPTIO) [31], illustrating crosstalk between H2S and NO during the response to Cd stress. Another study in wheat (Triticum aestivum L.) obtained similar results that exogenous H2S might correlate with NO to enhance Co tolerance [32]. The above studies show that H2S may cooperate with the NO signal in managing different heavy metal stresses in plants.
The pharmacological method of introducing specific scavengers into different experimental conditions was further employed to research the relationship between H2S and NO under heavy metal ion stress in plants. Cd stress was shown to induce a burst of endogenous NO and H2S in bermudagrass [Cynodon dactylon (L). Pers.] [33]. Moreover, exogenous NO donor SNP and H2S donor NaHS could improve Cd stress tolerance, while the positive roles of SNP and NaHS were specifically blocked by H2S scavenger hypotaurine (HT, C2H7NO2S), but not by NO scavenger cPTIO and H2S inhibitors potassium pyruvate (PP, C3H3KO3) and hydroxylamine (HA, H3NO). PP is regarded as the substrate of dehydrogenase. H2S could interact with the dehydrogenase. HA is an alkaline inorganic amine, which can react with the acid gas H2S; thereby, PP and HA are able to inhibit the production of endogenous H2S [33]. Thus, NO could activate the H2S signal in response to Cd stress, and maybe H2S is downstream of the NO signal. This phenomenon was further proved by the study of Al stress in soybean roots, in which NO modulated GmMATE13 and GmMATE47 gene expressions to enhance citrate secretion, and regulated PM H+-ATPase activity through regulating H2S biosynthesis and degradation [34]. H2S and NO improved Pb tolerance in Sesamum indicum, while the H2S-induced response was completely eliminated by NO scavenger cPTIO [35]. Meanwhile, only part of the effect conducted by NO was weakened by H2S scavenger HT. It seems that NO acts downstream of H2S or independent of H2S in conferring plant tolerance to Pd stress. More recently, the downstream role of NO in cooperation with H2S was also discovered in pepper (Capsicum annuum L.) and wheat under Cd stress [36,37]. From the numerous studies of H2S and NO, a hypothesis may be drawn that there exists a two-side signal cascades mechanism between H2S and NO in mediating heavy metal damage (Figure 2).
2.2. Crosstalk between H2S and NO in Response to Salt Stress
It has long been recognized that H2S and NO participate in alleviating salt stress in different plant species. Salt treatment (conducted by NaCl) could increase endogenous H2S and NO generation in the leaves of Nicotiana tabacum L. cv. Havana by increasing L-Cys and L-Arg contents and enhancing H2S and NO biosynthesis enzyme activities [38]. Then, H2S and NO help plants to cope with oxidative stress induced by salinity. These results suggest that both H2S and NO contribute to enhancing salt tolerance. Moreover, H2S donor NaHS and NO donor SNP relieved the inhibition of seed germination under salt stress in alfalfa through reestablishing ion homeostasis and maintaining activities of antioxidant enzymes [39]. The attenuation effect of salinity damage by H2S was reversed by NO scavenger cPTIO, suggesting that H2S enhanced salt tolerance through the NO pathway [39]. Another report discovered a similar relationship between H2S and NO in rescuing salt-induced inhibition of plant growth by regulating ion homeostasis [22].
The relationship between H2S and NO in salt resistance is still puzzled. It has been found that NO accumulation occurred ahead of H2S, however, H2S could not stimulate NO accumulation during the initial stage in salt-treated tomato (Solanum lycopersicum) roots [40]. The results above illustrate that H2S acts downstream of NO under salt stress, and may further induce NO production to strengthen the signal cascade in a feedback manner (Figure 2). In addition, H2S and NO may act downstream of MT to alleviate salt stress in pepper seedlings [41].
2.3. Crosstalk between H2S and NO in Response to Other Stresses
There also exists multiple pieces of evidence that H2S and NO cooperate with each other in heat, drought, osmotic, and flooding stresses. The pretreatment of exogenous NO enhanced the survival rate of maize seedlings under heat stress, and NO increased H2S content [42]. Furthermore, NO-induced heat tolerance was eliminated by H2S synthesis inhibitors and a H2S scavenger [42], indicating that H2S may act downstream of the NO signal in NO-induced heat tolerance. Later, another study discovered that SNP treatment facilitated the survival of submerged maize by enhancing the antioxidant system and regulating ROS content, elevating intracellular Ca2+ content and ADH activity, and increasing expressions of hypoxia-induced genes in maize seedling roots [43]. Moreover, SNP induced endogenous H2S generation, and H2S increased the NO-enhanced acquisition of tolerance to flooding-induced hypoxia in maize seedling roots [43], suggesting an analogical pattern of H2S and NO signal cascades in relieving heat and hypoxia stresses.
H2S may act as a downstream component of NO in ethylene-induced stomatal closure in Vicia faba L. [44]. Also, NO represented downstream of H2S in ABA-triggered stomatal closure, which may suggest a paradoxical relationship between H2S and NO under drought condition [45]. As for osmotic stress in wheat seedlings, the application of exogenous NO markedly improved H2S synthesis enzymes LCD and DCD, as well as enhancing the activity of O-acetylserine (thiol)lyase (OAS-TL) to modulate Cys homeostasis [46]. On the other hand, NO scavenger cPTIO and H2S scavenger HT invalidated the effect of NO on endogenous H2S levels and Cys homeostasis in wheat [46]. Thus, both H2S and NO could contribute to reinforcing osmotic tolerance and direct stomatal closure, though the concrete mechanism is largely unknown.
The H2S donor GYY4137 released a less severe H2S shock and a more prolonged H2S flux; however, it decreased NO accumulation in guard cells of A. thaliana leaves, in accordance with another type of H2S donor, NaHS [47]. In Medicago sativa, pretreatment with NOSH or NOSH-aspirin, the novel donors, which can donate NO and H2S simultaneously to plants, could enhance plant tolerance to drought stress and improve the recovery phenotype followed by rewatering [48]. Considering the cooperative relationship between H2S and NO, acting as signal molecules in retarding environmental damages, NOSH or NOSH-aspirin seems to be more favorable compared with NaHS and GYY4137 when used in plant guard cells, however, the effect and dosage have yet to be demonstrated (Figure 2).
3. Crosstalk between H2S and ABA in Response to Abiotic Stresses
ABA has long been recognized as a significant phytohormone with the function of regulating plant growth, development processes, and responses to diverse environmental stresses [49]. Within drought stress, ABA may take a central role in endogenous physiological processes, including stomatal movement [50,51]. Stomata are pores of plant aerial tissues and consist of a pair of guard cells. The stomatal aperture can be modulated by these specialized cells to respond to external and internal stimuli [52]. Within the past 10 years, the research of H2S and ABA crosstalk in augmenting plant tolerance to abiotic stresses has always come along with the regulation mechanism of stomatal movement.
3.1. Crosstalk between H2S and ABA in Response to Abiotic Stresses through Regulating Stomatal Closure
H2S cooperates with ABA in modulating the stomatal aperture, which has long been reported since [53] found that exogenous H2S regulated stomatal movement and enhanced leaf relative water content (RWC) to strengthen plant drought tolerance in Arabidopsis thaliana. Furthermore, scavenging H2S by HT or inhibiting H2S biosynthesis partially blocked ABA-dependent stomatal closure through regulating ATP-binding cassette transporters [53]. Similarly, pretreatment with H2S could considerably enhance rice’s tolerance to drought stress by decreasing lipid peroxidation, maintaining antioxidant system activation, and improving ABA biosynthesis [54]. The results above affirm a role of H2S in ABA signaling under environmental stresses. Furthermore, the stomatal aperture was enlarged in lcd mutant plants, causing a sensitive drought phenotype [55]. In addition, LCD expression and H2S generation were down-regulated in ABA-related mutants aba3 and abi1, and NaHS application increased stomatal closure in these mutants [55]. Thus, H2S may regulate stomatal aperture in an ABA-dependent manner, and ABA may induce H2S biosynthesis under drought stress. Simultaneously, another report revealed that pretreatment of exogenous H2S enhanced wheat seedling tolerance to drought conditions through reinforcing antioxidant capacity [56]. Besides, the application of H2S modulated ABA metabolic pathway genes and up-regulated ABA receptors, indicating again that H2S alleviates drought stress, at least in part, through the ABA signaling pathway. Furthermore, exogenous ABA induced the endogenous H2S content under drought stress [56], illustrating a complex relationship between H2S and ABA signals in modulating drought stress.
Mitogen-activated protein kinases (MAPKs) belong to a crucial signaling molecule family which adjusts plants to multiple environmental stimuli [49]. In A. thaliana, drought stress fortified H2S generation and gene expression of MAPK, however, the induced MAPK expression was abolished in H2S synthesis double mutants lcd des1 [57]. Further, the contributions of ABA to stomatal movements were also inhibited in lcd des1 and mpk4 mutants. In addition, H2S-enhanced stomatal closure was impaired in slac1-3 mutants [57], in which SLAC1 is an S-type anion channel that responds to ABA signaling in stomatal closure [58]. A previous report announced that H2S could activate S-type anion currents via SLAC1 to induce stomatal closure [59]. In all, it could be proposed that H2S is involved in ABA-stimulated stomatal closure. Thus, MPK4 may act downstream of H2S, and H2S-MPK4 signal cascade is involved in ABA-stimulated stomatal closure in alleviating drought stress [57].
Osmotic stress adversely causes internal environmental disorder on account of the overproduction of ROS, which leads to a decrease in plant growth and productivity. Usually, plants resist osmotic stress by enhancing the antioxidant system and stimulating signal transductions [60]. Wheat could adjust itself to resisting osmotic stress by enhancing antioxidant systems and inducing H2S biosynthesis [61]. Furthermore, exogenous ABA induced AsA-GSH cycle activity, but H2S scavenger HT and synthesis inhibitor aminooxy acetic acid (AOA) reversed the activities mentioned above [61]. These results suggest that H2S induced by exogenous ABA is a signal that triggers the up-regulation of the AsA-GSH cycle under osmotic stress. Obviously, H2S takes part in ABA-related stomatal closure in response to different environmental stresses; however, the relationship between them is complicated (Figure 3).
3.2. Crosstalk between DES1/H2S and ABA in Response to Drought Stress through Regulating Protein Persulfidation
ABA could stimulate H2S generation under stresses, but how H2S synthesis enzyme DES1 contributes to the crosstalk between ABA and H2S is puzzled. Recently, by creating transgenic lines that expressed DES1 in a tissue-specific pattern, it was found that the guard cell-specific DES1 was involved in ABA-induced physiological molecular responses [62]. ABA-induced DES1 expression and H2S production in guard cells were inhibited by H2S scavenger and restored by H2S donor [62]. The above genetic and pharmacological evidence further confirmed the hypothesis that DES1 is a unique component in ABA signaling in guard cells, and guard cell in situ DES1, together with H2S, participates in ABA-guided stomatal closure [63].
Excitingly, another report discovered that the ABA signal was, in turn, commanded by H2S-induced persulfidation of Open stomata 1 (OST1)/Snf1-related protein kinase 2.6 (SnRK2.6) on Cys131 and Cys137 residues in A. thaliana [64]. The persulfidated SnRK2.6 then interacted with ABA response element-binding factor 2 (ABF2), an ABA downstream protein, to modulate stomatal movement. Also, ABA was detected to induce DES1 and DCD expressions within 5–30 min previously [63,65], which suggests that the accumulation of H2S by ABA is ahead of the occurrence of protein persulfidation. Together with the works above, a hypothesis that ABA induces H2S accumulation, which further persulfidates SnRK2.6 continuously to promote ABA signaling in guard cells, would be proposed. The persulfidated SnRK2.6 then enhanced ABA- and H2S-induced Ca2+ influx, which subsequently caused stomatal closure through the inhibition of inward K+ channels and activation of outward anion channels [66]. To be encouraged continually, the DES1/H2S-triggered persulfidation mechanism in ABA-regulated stomatal movement has been confirmed in another two reports [67,68]. One of their works found that ABA triggered DES1 accumulation, and DES1 auto-presulfidated at Cys44 and Cys205 in a redox-dependent fashion, causing a trigger of transient H2S overproduction in guard cells [67]. They also found that the sustained DES1/H2S drove persulfidation of the NADPH oxidase respiratory burst oxidase homolog protein d (RBOHD) at Cys825 and Cys890 to strengthen its ability to introduce a ROS burst, which in turn induced stomatal closure [67]. Together, this work suggests that H2S-guided persulfidantion of DES1 and RBOHD may form a negative feedback loop that fine-tunes guard cell redox homeostasis and ABA signaling. Abscisic acid insensitive 4 (ABI4) could also be persulfidated by DES1 at Cys250 in vitro and in vivo, and served as a downstream target of H2S in plant’s response to ABA under stress conditions [68]. In addition, DES1-linked persulfication of ABI4 induced MPAKKK18 transactivation through binding to the CE1 motif in the MAPKKK18 promoter, which further enlarged the MAPK signaling cascade induced by ABA. Meanwhile, ABI4 could bind to the DES1 promoter and, in turn, activate its transcription, forming a DES1-ABI4 loop to fine-tune ABA-MAPK signals [68]. The results above illustrate a redox-based protein persulfidation mechanism within the crosstalk between H2S- and ABA-involved stomatal movement [69]. Further work may focus on the molecular mechanisms of persulfidation and other post-translational modification events in H2S-regulated ABA signaling in guard cells (Figure 3).
4. Crosstalk between H2S and Ca2+ in Response to Abiotic Stresses
Ca2+ is another well-known second messenger in plant cells with the function of regulating intracellular physiological and biochemical processes, including alleviating abiotic stresses. Calmodulin (CaM) is a receptor protein in calcium signal transduction, and its main function is to perceive the volatility of intracellular calcium ions [10,70]. Recent studies uncovered a new signal transduction pattern in which Ca2+ and H2S cooperate to help plants resist environmental stresses.
4.1. Crosstalk between H2S and Ca2+ in Response to Heavy Metal Stress
Ca2+ influx was found to participate in restraining heavy metal contamination together with H2S signal cascade. H2S synthesis inhibitor and Ca2+ chelators aggravated the toxic phenotypes of foxtail millet (Setaria italica) exposed to Cr (VI) damage, demonstrating the involvement of H2S and Ca2+ signals during this process [71]. Furthermore, Ca2+ enhanced the expressions of heavy metal chelator biosynthesis genes Metallothionein-like type 3 (MT3A) and Phytochelatin Synthase (PCS) and activated the antioxidant system, which was partially dependent on the H2S signal [71], indicating a downstream role of H2S in Ca2+ signaling. A later report in A. thaliana further discovered that the expression of H2S synthesis enzyme LCD was increased through a Ca2+/calmodulin 2 (CaM2)-directed pathway, which may explain the generation of H2S in the defense of plants against the Cr (VI) toxic condition [72,73]. The detailed mechanism was that the extracellular Cr (VI) stimulated Ca2+ influx, and the CaM2 protein then bound Ca2+ and interacted with the bZIP transcription factor TGA3, which further reinforced LCD gene expression and enhanced H2S production [72]. Ca2+ and H2S donor NaHS induced AsA-GSH cycle, redox homeostasis, and Ca2+-dependent protein kinase (CDPK) and Phytochelatins (PCs) genes expressions under Ni toxicity in zucchini seedlings [74]. In addition, H2S scavenger HT inhibited H2S accumulation induced by Ca2+, and Ca2+ chelator ethylene glycol-bis(b-aminoethylether)-N,N,N’,N’-tetra-acetic acid (EGTA) eliminated the impacts of seed priming induced by NaHS [74]. Thus, Ca2+ and H2S may manifest a two-side crosstalk in inoculating plants against heavy metal conditions. The relationship between NO and H2S has been discussed in another part of the present article, and it was put forward that Ca2+, in association with NO and H2S, improved chlorophyll metabolism, photosynthesis, carbohydrate accumulation, and maintained redox homeostasis in Vigna radiata under Cd stress [32]. The study also discovered that NO scavenger cPTIO could reduce Ca2+ content, and that EGTA reduced H2S content and altered Ca2+-dependent LCD and DCD enzyme activities, but that HT could not considerably reduce Ca2+ content [32]. Therefore, Ca2+, as a downstream signal of NO, may act in a two-side crosstalk pattern with H2S during plants’ adjustment to heavy mental contamination (Figure 2).
4.2. Crosstalk between H2S and Ca2+ in the Regulation of Stomatal Closure
Stomatal closure is an important physiological process under stress conditions; thus, the role of Ca2+ in stomatal closure was also summarized here. As mentioned above, H2S contributed to regulate S-type anion channel activation in guard cells, and this process was correlated with the SnRK2.6 function and the level of cytosolic free Ca2+ [59]. Further, H2S induced the Ca2+ influx in guard cells by stimulating the accumulation of ROS [75]. H2S triggered the persulfidation of SnRK2.6, and the persulfidated SnRK2.6 enhanced ABA- and H2S-induced Ca2+ influx, which subsequently caused stomatal closure [64]. Therefore, Ca2+ may function downstream of H2S-driven stomatal closure in a redox- and post-translational persulfidation-dependent manner (Figure 2).
4.3. Crosstalk between H2S and Ca2+ in Response to Other Stresses
As signal messengers, the crosstalk between H2S and Ca2+ has also been validated in many kinds of other stress conditions. Pretreating with H2S enhanced the heat tolerance of tobacco (Nicotiana tabacum L.) suspension-cultured cells by inhibiting electrolyte leakage and MDA accumulation, and exogenous Ca2+ and its ionophore A23187 intensified these effects [76]. However, H2S-induced heat tolerance was restrained by the application of Ca2+ chelator EGTA, as well as CaM antagonists chlorpromazine (CPZ) and trifluoperazine (TFP), illustrating a role of Ca2+ and CaM in H2S-triggered heat tolerance [76]. Afterward, another study announced that exogenous H2S enhanced the heat resistance of wheat coleoptiles through strengthening antioxidant enzyme activities in a Ca2+-dependent manner [77]. Thus, Ca2+ and CaM participate in H2S-induced heat tolerance in plants.
As for K+ deficiency under NaCl stress in Vigna radiata seedlings, Ca2+ increased endogenous H2S generation, and Ca2+ and H2S then cooperated with each other to induce an Na+/H+ antiport system and antioxidant defense [78]. Considering another result that adding of Ca2+-chelator EGTA and H2S scavenger HT reversed the effects of Ca2+ [78], a hypothesis may be drawn that H2S acts downstream during Ca2+-mediated plant adaptive responses to NaCl stress (Figure 2).
5. Crosstalk between H2S and H2O2 in Response to Abiotic Stresses
H2O2 is a colorless transparent liquid and crucial signaling molecule. Various studies have shown that H2O2 plays important roles in seed germination, stomatal movement, shoot and root development, pollination, and fruit ripening [79]. Also, it can modulate the plant growth and development under abiotic stresses [80]. The crosstalk between H2S and H2O2 under stress has been studied in recent years.
5.1. Crosstalk between H2S and H2O2 in Response to Heavy Metal Stress
Cd stress could regulate the homeostasis of ROS and promote oxidative injury, which may cause cell death [81]. Cd could decrease vacuolar H+-ATPase activity, which was able to generate a proton gradient across the vacuolar membrane [82]. Under high Cd concentration stress, H2O2 and O2·- significantly enhanced and triggered the oxidative injury, thus resulting in cell death in Brassica rapa root tips [81]. However, when B. rapa was exposed to low concentration Cd stress, the transcript levels of H2S biosynthesis-related genes LCD and DCD were significantly increased. Simultaneously, H2O2 had a remarkable increase and O2·- went down, whereas H2S biosynthesis inhibitor or H2S scavenger reversed the positive effects, indicating a role of H2S in alleviating low Cd stress by adjusting the balance between H2O2 and O2·− [81]. H2S donor NaHS treatment increased the photosynthetic fluorescence parameters in cotyledons of cucumber (C. sativus L. var. Wisconsin) seedling roots exposed to 100 μM CdCl2 for 24 h [82]. In addition, both the enhancement of H2O2 content and the decline in H2S content in roots decreased vacuolar H+-ATPase activity under Cd stress. Further, the increase in H2S content in root tissue by exogenous H2O2 had nothing to do with the desulfurization enzyme activity. Exogenous H2S remarkably enhanced the NADPH oxidase activity and the relative gene expression; however, it did not have an effect on the accumulation of H2O2 in cucumber roots under Cd stress [82]. Hence, H2S content might be partially enhanced through the H2O2/NADPH oxidase-induced pathway, independent of desulfhydrase activity (Figure 4).
5.2. Crosstalk between H2S and H2O2 in Response to Salt Stress
H2S donor NaHS could enhance the activity of PM H+-ATPase under salt or low-temperature stress in cucumber, and the transcript levels of the plasma membrane proton pump-related genes including CsHA2, CsH4, CsH8, CsH9, and CsHA10 were also increased [83]. However, NO and H2O2 only enhanced the expression of CsHA1. Therefore, H2S, NO, and H2O2 could resist the salt stress by regulating the plasma membrane proton pump at different standards. Usually, salt stress could induce stomata closure. However, the H2S scavengers HT, AOA, hydroxylamine (NH2OH), potassium pyruvate (C3H3KO3), ammonia (NH3), H2O2, ascorbic acid (AsA), CAT, and diphenyl iodide (DPI) suppressed the closure of stomata in V. faba L. [44], suggesting that both H2S and H2O2 could regulate stomatal movement under salt stress. Furthermore, endogenous H2S and H2O2 accumulation and the activities of LCD and DCD were enhanced by salt treatment in guard cells. Nevertheless, these effects were inhibited by H2O2 and H2S scavengers. Exogenous H2O2 scavengers prevented the increase in endogenous H2S level as well as the stomatal closure; however, H2O2 generation was barely influenced with the application of H2S scavengers in guard cells responding to salt stress [44]. Hence, H2S may act as the downstream of H2O2-alleviated salt stress (Figure 4).
5.3. Crosstalk between H2S and H2O2 in Response to Drought Stress
Drought stress is one of the most serious abiotic stresses in the world. Treatment by spermidine (Spd) remarkably enhanced H2S production and activities of antioxidant enzymes [SOD, CAT, guaiacol peroxidase (GPOX), APX, GR, dehydroascorbate reductase (DHAR), and monodehydroascorbate reductase (MDHAR)] in white clover (Trifolium repens) under dehydration conditions [84]. Furthermore, NO and H2S scavengers could not reduce the generation of H2O2 induced by Spd, but H2O2 scavengers could effectively inhibit the increase of NO and H2S induced by Spd. The H2S signal induced by Spd was also significantly inhibited by NO scavenger [84]. Hence, in response to dehydration, H2S may be the downstream signaling molecule to interact with NO and H2O2 (Figure 4).
5.4. Crosstalk between H2S and H2O2 in Response to Other Stresses
UV-B is a common stress in practical agricultural production. When plants encounter the UV-B stress, the levels of electrolyte leakage, MDA, and ultraviolet absorbing compounds decreased, and the activities of antioxidant enzymes, GSH, and AsA also declined [85]. However, exogenous H2S, H2O2, and putrescence (Put) could alleviate the negative effects of UV-B stress. The protective role of Put in UV-B radiation damage was reduced by the inhibitors of H2S, H2O2, and Put [86]. Moreover, the level of H2O2 was increased by exogenous H2S, and the enhanced H2O2 promoted the accumulation of UV absorbing compounds in hulless barley (H. vulgare L. var. nude, Kunlun-12) seedlings, thus preserving the steady state of oxidation-reduction under UV-B stress and improving its UV-B tolerance [86].
In addition, extreme temperature is a key factor which influences plant growth and development. H2S, NO, and H2O2 had a significant impact in response to low temperature (10 °C) by modulating the plasma membrane proton pump in cucumber roots [83]. Moreover, H2O2 treatment could improve the heat resistance in maize (Z. mays L., Huidan No. 4) seedlings, and this effect could be strengthened by NO and H2S donors but abolished by NO and H2S scavengers or synthesis inhibitors [87]. It seems that NO and H2S act downstream of H2O2 in the acquisition of heat resistance in plants (Figure 4).
6. Crosstalk between H2S and Other Signal Molecules in Response to Abiotic Stresses
In recent years, many kinds of signal transmitters have emerged to regulate plant growth and development, and to acclimate to environment changes. The protective role of H2S related to these signal molecules such as SA, ETH, JA, Pro, and MT (mentioned in another part of the article) under toxic environment in plants has also been explored to some extent.
6.1. Crosstalk between H2S and SA in Response to Abiotic Stresses
SA has long been recognized as a pivotal signal messenger, manifesting multiple functions in defending plant disease and adverse environmental conditions. Endogenous SA biosynthesis is mainly proceeded in the cytoplasm through the phenylalanine route by phenylalanine ammonia lyase (PAL) and benzoic-acid-2-hydroxylase (BA2H) [10,88,89]. SA and H2S enhanced heat tolerance by strengthening the activities of antioxidant enzymes and increasing osmolyte content in maize seedlings [90]. Further, SA induced endogenous H2S generation by enhancing the activity of H2S synthesis enzyme DES [91]. While the increase in SA production and the relative enzyme activities of PAL and BA2H were rarely influenced by H2S, this downstream role of H2S in SA-induced stress responses was also similarly reported in Cd tolerance in A. thaliana [92]. Thus, the positive role of SA under the stress condition is partially dependent on H2S. Pb stress accelerated endogenous H2S production [35]. Moreover, SA improved enzyme activities of the AsA-GSH cycle system in pepper under Pb stress [93]. In addition, exogenous SA enhanced the H2S content, which was further reinforced by H2S donor NaHS. It seems that SA triggers endogenous H2S accumulation, which further regulates the AsA-GSH cycle to resist Pb toxicity (Figure 5).
6.2. Crosstalk between H2S and ETH in Response to Abiotic Stresses
Ethylene induced H2S biosynthesis in guard cells in tomatoes under osmotic stress [94]. Moreover, the effect of ethylene on resisting osmotic stress was reversed by H2S scavenger HT or H2S synthetic inhibitor PAG, suggesting a downstream component of H2S in ethylene-triggered stomatal closure under osmotic stress. Further, H2S induced the persulfidation of 1-aminocyclopropane-1-carboxylic acid oxidase1 (ACO1) and ACO2, and restrained their expressions. As a result, H2S negatively regulated ethylene generation in response to osmotic stress [94]. These results are parallel with a recently published mechanism of waterlogging damage resistance in peach (Prunus persica L. Batsch) seedlings [95], in which H2S restrained over-synthesis of ethylene as well as inhibited oxidative damage under waterlogging stress (Figure 5).
6.3. Crosstalk between H2S and JA in Response to Abiotic Stresses
JA is another phytohormone kind signal transmitter with extensive modulation functions in plant root elongation [96], anthocyanin accumulation and trichome initiation [97], stamen development and flowing [98], leaf senescence [99], and stress resistance [100]. A recent study announced a critical role of JA in inhibiting stomatal development in A. thaliana [101]. Furthermore, JA positively modified LCD activity and H2S production. The JA-deficient mutants represented a high stomatal density phenotype, which could be reversed by exogenous H2S, whereas the H2S synthesis-deficient mutants lcd displayed similar stomatal development phenotype as the JA-deficient mutants, which could be rescued by H2S donor NaHS but not by JA [102]. Thus, H2S may act as a downstream member of JA in stomatal development (Figure 5).
6.4. Crosstalk between H2S and Pro in Response to Abiotic Stresses
Pro is a kind of organic osmolyte with a wide distribution in plant cells. Previous studies have demonstrated the increase of Pro after the application of signal transmitter agents in defense of abiotic stresses [89,103,104]. Pretreatment with exogenous H2S increased endogenous Pro content, and the activities and transcription levels of proline-5-carboxylate reductase (P5CR) and proline dehydrogenase (PDH) in foxtail millet, whereas H2S scavenger or inhibitor reduced the above effects [105]. Moreover, the combined application of H2S and Pro resulted in preferable growth status, stomatal movement, and oxidative remission under stress conditions. These results indicate a cooperation of Pro and H2S under adverse environments (Figure 5).
7. Conclusions and Outlook
The disadvantageous environment conditions cause oxidative damage, ionic imbalance, and osmotic stress to plants, resulting in a weakened growth and development status. H2S can reinforce plant tolerance to these stresses through constructing a luxuriant crosstalk with other signal molecules, such as NO, ABA, Ca2+, H2O2, SA, ETH, JA, Pro, and MT. The genes regulated by H2S and other molecules under abiotic stress conditions are displayed in Table 1. There exists a legible clue that environmental stresses and various signal transmitters stimulate endogenous H2S generation and improve the activities of H2S synthesis enzymes under the stress condition. Meanwhile, H2S represents a feedback manner to enhance the signal cascades in inducing the accumulation of some signal messengers, especially NO, ABA, and Ca2+. In addition, the existence of DES1-related auto-persulfidation and persulfidation may be the reason for the extensive inspiration of its enzyme activity in different stress conditions. In summary, H2S acts as a downstream signal member in cooperation with ABA, H2O2, SA, ETH, JA, and MT, but an upstream signal member of Pro under stress condition. Nevertheless, the crosstalk between H2S, NO, and Ca2+ represents a two-side signal cascades manner, whereas relationships between H2S and other signal molecules vary on account of the specific stress pattern.
Multiple types of research need to be done to explore the point-to-point mechanism within the crosstalk between H2S and one single signal transducer under abiotic stress conditions. Firstly, the feedback molecular mechanism of H2S and NO, and the interactions within protein persulfidation, S-sulfhydration, and S-nitrosylation, remain unclear. Next, more post-translational modification proteins need to be discovered and identified that are triggered by H2S in ABA- or NO-dependent signal pathways under stress condition. Finally, new signal messengers related to H2S activity are waiting to be discovered.
Author Contributions
Conceptualization W.L. and C.W.; Collection and analysis of bibliography, Z.L. and Y.D.; Writing original draft, W.L., C.W., Y.D., and Z.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Scientific research start-up funds for openly-recruited doctors (GAU-KYQD-2017RCZX-29); the National Natural Science Foundation of China (Nos. 32102370, 32072559, 31860568, 31560563, and 31160398); the National Key Research and Development Program of China (Grant No. 2018YFD1000800); the Natural Science Foundation of Gansu Province, China (No. 20JR5RA027); the Fuxi Young Talents Fund of Gansu Agricultural University (No. Gaufx-03Y07).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kazan, K. Diverse roles of jasmonates and ethylene in abiotic stress tolerance. Trends Plant Sci. 2015, 20, 219–229. [Google Scholar] [CrossRef]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy metal stress and some mechanisms of plant defense response. Sci. World J. 2015, 2015, 756120. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Devireddy, A.R.; Zandalinas, S.I.; Fichman, Y.; Mittler, R. Integration of reactive oxygen species and hormone signaling during abiotic stress. Plant J. 2021, 105, 459–476. [Google Scholar] [CrossRef] [PubMed]
- Fotopoulos, V.; Christou, A.; Manganaris, G.A. Hydrogen sulfide as a potent regulator of plant responses to abiotic stress factors. Mol. Approaches Plant Abiotic Stress 2013, 353–373. [Google Scholar] [CrossRef]
- Li, W.; Pang, S.; Lu, Z.; Jin, B. Function and mechanism of WRKY transcription factors in abiotic stress responses of plants. Plants 2020, 9, 1515. [Google Scholar] [CrossRef] [PubMed]
- Calderwood, A.; Kopriva, S. Hydrogen sulfide in plants: From dissipation of excess sulfur to signaling molecule. Nitric oxide. 2014, 41, 72–78. [Google Scholar] [CrossRef]
- Wilson, L.G.; Bressan, R.A.; Filner, P. Light-dependent Emission of Hydrogen Sulfide from Plants. Plant Physiol. 1978, 61, 184–189. [Google Scholar] [CrossRef] [Green Version]
- Papenbrock, J.; Riemenschneider, A.; Kamp, A.; Schulz-Vogt, H.N.; Schmidt, A. Characterization of Cysteine-Degrading and H2S-Releasing Enzymes of Higher Plants-From the Field to the Test Tube and Back. Plant Biol. 2007, 9, 582–588. [Google Scholar] [CrossRef]
- Huang, D.; Huo, J.; Liao, W. Hydrogen sulfide: Roles in plant abiotic stress response and crosstalk with other signals. Plant Sci. 2021, 302, 110733. [Google Scholar] [CrossRef]
- Takahashi, H.; Kopriva, S.; Giordano, M.; Saito, K.; Hell, R. Sulfur assimilation in photosynthetic organisms: Molecular functions and regulations of transporters and assimilatory enzymes. Annu. Rev. Plant Biol. 2011, 62, 157–184. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Kumar, V.; Kapoor, D.; Kumar, S.; Singh, S.; Dhanjal, D.S.; Datta, S.; Samuel, J.; Dey, P.; Wang, S.; et al. Revealing on hydrogen sulfide and nitric oxide signals coordination for plant growth under stress conditions. Physiol. Plant. 2019, 168, 301–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Z.W.; Deng, L.W. Role of H2S Donors in Cancer Biology. Handb. Exp. Pharmacol. 2015, 230, 243–265. [Google Scholar] [CrossRef] [PubMed]
- Akter, F. The role of hydrogen sulfide in burns. Burns 2016, 42, 519–525. [Google Scholar] [CrossRef]
- Tabassum, R.; Jeong, N.; Jung, J. Therapeutic importance of hydrogen sulfide in age-associated neurodegenerative diseases. Neural. Regen. Res. 2020, 15, 653–662. [Google Scholar] [CrossRef]
- Bhatia, M. H2S and inflammation: An overview. Handb. Exp. Pharmacol. 2015, 230, 165–180. [Google Scholar] [CrossRef]
- Mukherjee, S.; Corpas, F.J. Crosstalk among hydrogen sulfide (H2S), nitric oxide (NO) and carbon monoxide (CO) in root-system development and its rhizosphere interactions: A gaseous interactome. Plant Physiol. Bioch. 2020, 155, 800–814. [Google Scholar] [CrossRef]
- Aroca, A.; Gotor, C.; Romero, L.C. Hydrogen Sulfide Signaling in Plants: Emerging Roles of Protein Persulfidation. Front. Plant Sci. 2018, 9, 1369. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhou, H.; Zhou, M.; Ge, Z.; Zhang, F.; Foyer, C.H.; Yuan, X.; Xie, Y. The coordination of guard-cell autonomous ABA synthesis and DES1 function in situ regulates plant water deficit responses. J. Adv. Res. 2020, 27, 191–197. [Google Scholar] [CrossRef]
- Singh, V.P.; Tripathi, D.K.; Fotopoulos, V. Hydrogen sulfide and nitric oxide signal integration and plant development under stressed/non-stressed conditions. Physiol. Plant. 2020, 168, 239–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corpas, F.J.; Barroso, J.B.; González-Gordo, S.; Muñoz-Vargas, M.A.; Palma, J.M. Hydrogen sulfide: A novel component in Arabidopsis peroxisomes which triggers catalase inhibition. J. Integr. Plant Biol. 2019, 61, 871–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Wang, W.H.; Wu, F.H.; He, E.M.; Liu, X.; Shangguan, Z.P.; Zheng, H.L. Hydrogen sulfide enhances salt tolerance through nitric oxide-mediated maintenance of ion homeostasis in barley seedling roots. Sci. Rep. 2015, 5, 12516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xuan, L.; Li, J.; Wang, X.; Wang, C. Crosstalk between hydrogen sulfide and other signal molecules regulates plant growth and development. Int. J. Mol. Sci. 2020, 21, 4593. [Google Scholar] [CrossRef]
- Ahmad, R.; Ali, S.; Rizwan, M.; Dawood, M.; Farid, M.; Hussain, A.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Hydrogen sulfide alleviates chromium stress on cauliflower by restricting its uptake and enhancing antioxidative system. Physiol. Plant. 2020, 168, 289–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Cheng, P.; Wang, Y.; Li, Y.; Su, J.; Chen, Z.; Yu, X.; Shen, W. Genetic elucidation of hydrogen signaling in plant osmotic tolerance and stomatal closure via hydrogen sulfide. Free Radic. Bio. Med. 2020, 161, 1–14. [Google Scholar] [CrossRef]
- Deng, Y.; Wang, C.; Wang, N.; Wei, L.; Li, W.; Yao, Y.; Liao, W. Roles of Small-Molecule Compounds in Plant Adventitious Root Development. Biomolecules 2019, 9, 420. [Google Scholar] [CrossRef] [Green Version]
- Corpas, F.J. Nitric oxide and hydrogen sulfide in higher plants under physiological and stress conditions. Antioxidants 2019, 8, 457. [Google Scholar] [CrossRef] [Green Version]
- Palma, J.M.; Mateos, R.M.; López-Jaramillo, J.; Rodríguez-Ruiz, M.; González-Gordo, S.; Lechuga-Sancho, A.M.; Corpas, F.J. Plant catalases as NO and H2S targets. Redox Biol. 2020, 34, 101525. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.P.; Singh, S.; Kumar, J.; Prasad, S.M. Hydrogen sulfide alleviates toxic effects of arsenate in pea seedlings through up-regulation of the ascorbate-glutathione cycle: Possible involvement of nitric oxide. J. Plant Physiol. 2015, 181, 20–29. [Google Scholar] [CrossRef]
- Kharbech, O.; Houmani, H.; Chaoui, A.; Corpas, F.J. Alleviation of Cr(VI)-induced oxidative stress in maize (Zea mays L.) seedlings by NO and H2S donors through differential organ-dependent regulation of ROS and NADPH-recycling metabolisms. J. Plant Physiol. 2017, 219, 71–80. [Google Scholar] [CrossRef]
- Li, L.; Wang, Y.; Shen, W. Roles of hydrogen sulfide and nitric oxide in the alleviation of cadmium-induced oxidative damage in alfalfa seedling roots. Biometals 2012, 25, 617–631. [Google Scholar] [CrossRef]
- Ozfidan-Konakci, C.; Yildiztugay, E.; Elbasan, F.; Kucukoduk, M.; Turkan, I. Hydrogen sulfide (H2S) and nitric oxide (NO) alleviate cobalt toxicity in wheat (Triticum aestivum L.) by modulating photosynthesis, chloroplastic redox and antioxidant capacity. J. Hazard. Mater. 2020, 388, 122061. [Google Scholar] [CrossRef]
- Shi, H.; Ye, T.; Chan, Z. Nitric oxide-activated hydrogen sulfide is essential for cadmium stress response in bermudagrass (Cynodon dactylon (L). Pers.). Plant Physiol. Bioch. 2014, 74, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ji, F.; Zhang, Y.; Hou, J.; Liu, W.; Huang, J.; Liang, W. Interactions between hydrogen sulphide and nitric oxide regulate two soybean citrate transporters during the alleviation of aluminium toxicity. Plant Cell Environ. 2019, 42, 2340–2356. [Google Scholar] [CrossRef]
- Amooaghaie, R.; Zangene-Madar, F.; Enteshari, S. Role of two-sided crosstalk between NO and H2S on improvement of mineral homeostasis and antioxidative defense in Sesamum indicum under lead stress. Ecotox. Environ. Saf. 2017, 139, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Akram, N.A.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Exogenously supplied silicon (Si) improves cadmium tolerance in pepper (Capsicum annuum L.) by up-regulating the synthesis of nitric oxide and hydrogen sulfide. J. Biotechnol. 2020, 316, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Responses of nitric oxide and hydrogen sulfide in regulating oxidative defence system in wheat plants grown under cadmium stress. Physiol. Plant. 2019, 168, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, C.J.; Batista Fontes, E.P.; Modolo, L.V. Salinity-induced accumulation of endogenous H2S and NO is associated with modulation of the antioxidant and redox defense systems in Nicotiana tabacum L. cv. Havana. Plant Sci. 2017, 256, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, L.; Cui, W.; Xu, S.; Shen, W.; Wang, R. Hydrogen sulfide enhances alfalfa (Medicago sativa) tolerance against salinity during seed germination by nitric oxide pathway. Plant Soil 2012, 351, 107–119. [Google Scholar] [CrossRef]
- Da-Silva, C.J.; Mollica, D.C.F.; Vicente, M.H.; Peres, L.E.P.; Modolo, L.V. NO, hydrogen sulfide does not come first during tomato response to high salinity. Nitric Oxide 2018, 76, 164–173. [Google Scholar] [CrossRef]
- Kaya, C.; Higgs, D.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Integrative roles of nitric oxide and hydrogen sulfide in melatonin-induced tolerance of pepper (Capsicum annuum L.) plants to iron deficiency and salt stress alone or in combination. Physiol. Plant. 2020, 168, 256–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Yang, S.; Long, W.; Yang, G.; Shen, Z. Hydrogen sulphide may be a novel downstream signal molecule in nitric oxide-induced heat tolerance of maize (Zea mays L.) seedlings. Plant Cell Environ. 2013, 36, 1564–1572. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.; Bian, Z.; Zhou, L.; Cheng, W.; Hai, N.; Yang, C.; Yang, T.; Wang, X.; Wang, C. Hydrogen sulfide enhances nitric oxide-induced tolerance of hypoxia in maize (Zea mays L.). Plant Cell Rep. 2016, 35, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hou, Z.; Liu, G.; Hou, L.; Liu, X. Hydrogen sulfide may function downstream of nitric oxide in ethylene-induced stomatal closure in Vicia faba L. J. Integr. Agric. 2012, 11, 1644–1653. [Google Scholar] [CrossRef]
- Mishra, V.; Singh, P.; Tripathi, D.K.; Corpas, F.J.; Singh, V.P. Nitric oxide and hydrogen sulfide: An indispensable combination for plant functioning. Trends Plant Sci. 2021, 1360–1385. [Google Scholar] [CrossRef]
- Khan, M.N.; Mobin, M.; Abbas, Z.K.; Siddiqui, M.H. Nitric oxide-induced synthesis of hydrogen sulfide alleviates osmotic stress in wheat seedlings through sustaining antioxidant enzymes, osmolyte accumulation and cysteine homeostasis. Nitric Oxide 2017, 68, 91–102. [Google Scholar] [CrossRef]
- Lisjak, M.; Srivastava, N.; Teklic, T.; Civale, L.; Lewandowski, K.; Wilson, I.; Wood, M.E.; Whiteman, M.; Hancock, J.T. A novel hydrogen sulfide donor causes stomatal opening and reduces nitric oxide accumulation. Plant Physiol. Bioch. 2010, 48, 931–935. [Google Scholar] [CrossRef]
- Antoniou, C.; Xenofontos, R.; Chatzimichail, G.; Christou, A.; Kashfi, K.; Fotopoulos, V. Exploring the potential of nitric oxide and hydrogen sulfide (NOSH)-releasing synthetic compounds as novel priming agents against drought stress in Medicago sativa plants. Biomolecules 2020, 10, 120. [Google Scholar] [CrossRef] [Green Version]
- Urhonen, T.; Lie, A.; Aamodt, G. Associations between long commutes and subjective health complaints among railway workers in Norway. Prev. Med. Rep. 2016, 4, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Duan, Y.; Hua, D.; Fan, G.; Wang, L.; Liu, Y.; Chen, Z.; Han, L.; Qu, L.; Gong, Z. DEXH box RNA helicase-mediated mitochondrial reactive oxygen species production in Arabidopsis mediates crosstalk between abscisic acid and auxin signaling. Plant Cell 2012, 24, 1815–1833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Xue, S. Interplay between hydrogen sulfide and other signaling molecules in the regulation of guard cell signaling and abiotic/biotic stress response. Plant Commun. 2021, 2, 100179. [Google Scholar] [CrossRef] [PubMed]
- García-Mata, C.; Lamattina, L. Hydrogen sulphide, a novel gasotransmitter involved in guard cell signalling. New Phytol. 2010, 188, 977–984. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, Y.; Zhai, F.; Zhang, J.; Zhang, F.; Yuan, X.; Xie, Y. Hydrogen sulfide promotes rice drought tolerance via reestablishing redox homeostasis and activation of ABA biosynthesis and signaling. Plant Physiol. Bioch. 2020, 155, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Xue, S.; Luo, Y.; Tian, B.; Fang, H.; Li, H.; Pei, Y. Hydrogen sulfide interacting with abscisic acid in stomatal regulation responses to drought stress in Arabidopsis. Plant Physiol. Bioch. 2013, 62, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Ding, H.; Wang, C.; Qin, H.; Han, Q.; Hou, J.; Lu, H.; Xie, Y.; Guo, T. Alleviation of drought stress by hydrogen sulfide is partially related to the abscisic acid signaling pathway in wheat. PLoS ONE 2016, 11, e163082. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Jin, Z.; Zhang, L.; Liu, X.; Yang, G.; Pei, Y. H2S is involved in ABA-mediated stomatal movement through MPK4 to alleviate drought stress in Arabidopsis thaliana. Plant Soil 2019, 435, 295–307. [Google Scholar] [CrossRef]
- Hua, D.; Wang, C.; He, J.; Liao, H.; Duan, Y.; Zhu, Z.; Guo, Y.; Chen, Z.; Gong, Z. A plasma membrane receptor kinase, GHR1, mediates abscisic acid- and hydrogen peroxide-regulated stomatal movement in Arabidopsis. Plant Cell 2012, 24, 2546–2561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wan, R.; Shi, Y.; Xue, S. Hydrogen sulfide activates S-type anion channel via OST1 and Ca2+ Modules. Mol. Plant 2016, 9, 489–491. [Google Scholar] [CrossRef] [Green Version]
- Golldack, D.; Lüking, I.; Yang, O. Plant tolerance to drought and salinity: Stress regulating transcription factors and their functional significance in the cellular transcriptional network. Plant Cell Rep. 2011, 30, 1383–1391. [Google Scholar] [CrossRef]
- Shan, C.; Zhang, S.; Zhou, Y. Hydrogen sulfide is involved in the regulation of ascorbate-glutathione cycle by exogenous ABA in wheat seedling leaves under osmotic stress. Cereal Res. Commun. 2017, 45, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhou, M.; Ge, Z.; Shen, J.; Zhou, C.; Gotor, C.; Romero, L.C.; Duan, X.; Liu, X.; Wu, D.; et al. Abscisic acid-triggered guard cell L-cysteine desulfhydrase function and in situ hydrogen sulfide production contributes to heme oxygenase-modulated stomatal closure. Plant Cell Environ. 2020, 43, 624–636. [Google Scholar] [CrossRef] [PubMed]
- Scuffi, D.; álvarez, C.; Laspina, N.; Gotor, C.; Lamattina, L.; García-Mata, C. Hydrogen sulfide generated by L-cysteine desulfhydrase acts upstream of nitric oxide to modulate abscisic acid-dependent stomatal closure. Plant Physiol. 2014, 166, 2065–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Jia, H.; Wang, X.; Shi, C.; Wang, X.; Ma, P.; Wang, J.; Ren, M.; Li, J. Hydrogen sulfide positively regulates abscisic acid signaling through persulfidation of SnRK2.6. Guard Cells 2020, 13, 732–744. [Google Scholar] [CrossRef]
- Hou, Z.; Wang, L.; Liu, J.; Hou, L.; Liu, X. Hydrogen sulfide regulates ethylene-induced stomatal closure in Arabidopsis thaliana. J. Integr. Plant Biol. 2013, 55, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Böhmer, M.; Hu, H.; Nishimura, N.; Schroeder, J.I. Guard cell signal transduction network: Advances in understanding abscisic acid, CO2, and Ca2+ signaling. Annu. Rev. Plant Biol. 2010, 61, 561–591. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Zhang, J.; Zhou, M.; Zhou, H.; Cui, B.; Gotor, C.; Romero, L.C.; Fu, L.; Yang, J.; Foyer, C.H.; et al. Persulfidation-based modification of cysteine desulfhydrase and the NADPH oxidase RBOHD controls guard cell abscisic acid signaling. Plant Cell 2020, 32, 1000–1017. [Google Scholar] [CrossRef]
- Zhou, M.; Zhang, J.; Shen, J.; Zhou, H.; Zhao, D.; Gotor, C.; Romero, L.C.; Fu, L.; Li, Z.; Yang, J.; et al. Hydrogen sulfide-linked persulfidation of ABI4 controls ABA responses through the transactivation of MAPKKK18. Arabidopsis 2021, 14, 921–936. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, J.; Shen, J.; Zhou, M.; Yuan, X.; Xie, Y. Redox-based protein persulfidation in guard cell ABA signaling. Plant Signal. Behav. 2020, 15, 1741987. [Google Scholar] [CrossRef]
- Peiter, E. The plant vacuole: Emitter and receiver of calcium signals. Cell Calcium 2011, 50, 120–128. [Google Scholar] [CrossRef]
- Fang, H.; Jing, T.; Liu, Z.; Zhang, L.; Jin, Z.; Pei, Y. Hydrogen sulfide interacts with calcium signaling to enhance the chromium tolerance in Setaria italica. Cell Calcium 2014, 56, 472–481. [Google Scholar] [CrossRef]
- Fang, H.; Liu, Z.; Long, Y.; Liang, Y.; Jin, Z.; Zhang, L.; Liu, D.; Li, H.; Zhai, J.; Pei, Y. The Ca2/calmodulin2-binding transcription factor TGA3 elevates LCD expression and H2S production to bolster Cr6+ tolerance in Arabidopsis. Plant J. 2017, 91, 1038–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, H.; Li, Y.; He, L. The central role of hydrogen sulfide in plant responses to toxic metal stress. Ecotox. Environ. Saf. 2018, 157, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Valivand, M.; Amooaghaie, R.; Ahadi, A. Seed priming with H2S and Ca2+ trigger signal memory that induces cross-adaptation against nickel stress in zucchini seedlings. Plant Physiol. Biochem. 2019, 143, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Shi, C.; Ding, X.; Liu, C.; Li, J. Hydrogen sulfide induces Ca2+ signal in guard cells by regulating reactive oxygen species accumulation. Plant Signal. Behav. 2020, 15, 1805228. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Gong, M.; Xie, H.; Yang, L.; Li, J. Hydrogen sulfide donor sodium hydrosulfide-induced heat tolerance in tobacco (Nicotiana tabacum L) suspension cultured cells and involvement of Ca2+ and calmodulin. Plant Sci. 2012, 185, 185–189. [Google Scholar] [CrossRef]
- Kolupaev, Y.E.; Firsova, E.N.; Yastreb, T.O.; Lugovaya, A.A. The participation of calcium ions and reactive oxygen species in the induction of antioxidant enzymes and heat resistance in plant cells by hydrogen sulfide donor. Appl. Biochem. Microbiol. 2017, 53, 573–579. [Google Scholar] [CrossRef]
- Khan, M.N.; Siddiqui, M.H.; Mukherjee, S.; Alamri, S.; Al-Amri, A.A.; Alsubaie, Q.D.; Al-Munqedhi, B.; Ali, H.M. Calcium-hydrogen sulfide crosstalk during K+-deficient NaCl stress operates through regulation of Na+/H+ antiport and antioxidative defense system in mung bean roots. Plant Physiol. Biochem. 2021, 159, 211–225. [Google Scholar] [CrossRef] [PubMed]
- Waszczak, C.; Carmody, M.; Kangasjarvi, J. Reactive oxygen species in plant signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [Green Version]
- Černý, M.; Habánová, H.; Berka, M.; Luklová, M.; Brzobohatý, B. Hydrogen peroxide: Its role in plant biology and crosstalk with signalling networks. Int. J. Mol. Sci. 2018, 19, 2812. [Google Scholar] [CrossRef] [Green Version]
- Lv, W.; Yang, L.; Xu, C.; Shi, Z.; Shao, J.; Xian, M.; Chen, J. Cadmium disrupts the balance between hydrogen peroxide and superoxide radical by regulating endogenous hydrogen sulfide in the root tip of Brassica rapa. Front. Plant Sci. 2017, 8, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabała, K.; Zboińska, M.; Głowiak, D.; Reda, M.; Jakubowska, D.; Janicka, M. Interaction between the signaling molecules hydrogen sulfide and hydrogen peroxide and their role in vacuolar H+-ATPase regulation in cadmium-stressed cucumber roots. Physiol. Plant. 2019, 166, 688–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janicka, M.; Reda, M.; Czyżewska, K.; Kabała, K. Involvement of signalling molecules NO, H2O2 and H2S in modification of plasma membrane proton pump in cucumber roots subjected to salt or low temperature stress. Funct. Plant Biol. 2018, 45, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhu, Y.; He, X.; Yong, B.; Peng, Y.; Zhang, X.; Ma, X.; Yan, Y.; Huang, L.; Nie, G. The hydrogen sulfide, a downstream signaling molecule of hydrogen peroxide and nitric oxide, involves spermidine-regulated transcription factors and antioxidant defense in white clover in response to dehydration. Environ. Exp. Bot. 2019, 161, 255–264. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, X.; Wang, X.; Wu, K.; Li, P.; Chang, N.; Wang, J.; Wang, F.; Li, J.; Bi, Y. Glucose-6-phosphate dehydrogenase and alternative oxidase are involved in the cross tolerance of highland barley to salt stress and UV-B radiation. J. Plant Physiol. 2015, 181, 83–95. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Z.; Zhao, Y.; Zhang, X.; Zhang, S.; Bo, L.; Wang, Y.; Ding, Y.; An, L. Putrescine protects hulless barley from damage due to UV-B stress via H2S- and H2O2-mediated signaling pathways. Plant Cell Rep. 2016, 35, 1155–1168. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Luo, L.; Sun, Y. Signal crosstalk between nitric oxide and hydrogen sulfide may be involved in hydrogen peroxide-induced thermotolerance in maize seedlings. Russ. J. Plant Physiol. 2015, 62, 507–514. [Google Scholar] [CrossRef]
- Janda, M.; Ruelland, E. Magical mystery tour: Salicylic acid signalling. Environ. Exp. Bot. 2015, 114, 117–128. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, M.; Zhou, H.; Zhao, D.; Gotor, C.; Romero, L.C.; Shen, J.; Ge, Z.; Zhang, Z.; Shen, W.; et al. Hydrogen sulfide, a signaling molecule in plant stress responses. J. Integr. Plant Biol. 2021, 63, 146–160. [Google Scholar] [CrossRef]
- Li, Z. Synergistic effect of antioxidant system and osmolyte in hydrogen sulfide and salicylic acid crosstalk-induced heat tolerance in maize (Zea mays L.) seedlings. Plant Signal. Behav. 2015, 10, e1051278. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Xie, L.; Li, X. Hydrogen sulfide acts as a downstream signal molecule in salicylic acid-induced heat tolerance in maize (Zea mays L.) seedlings. J. Plant Physiol. 2015, 177, 121–127. [Google Scholar] [CrossRef]
- Qiao, Z.; Jing, T.; Liu, Z.; Zhang, L.; Jin, Z.; Liu, D.; Pei, Y. H2S acting as a downstream signaling molecule of SA regulates Cd tolerance in Arabidopsis. Plant Soil. 2015, 393, 137–146. [Google Scholar] [CrossRef]
- Kaya, C. Salicylic acid-induced hydrogen sulphide improves lead stress tolerance in pepper plants by upraising the ascorbate-glutathione cycle. Physiol. Plant. 2020, 173, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Chen, S.; Liu, D.; Liesche, J.; Shi, C.; Wang, J.; Ren, M.; Wang, X.; Yang, J.; Shi, W.; et al. Ethylene-induced hydrogen sulfide negatively regulates ethylene biosynthesis by persulfidation of ACO in tomato under osmotic stress. Front. Plant Sci. 2018, 9, 1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Wu, X.; Sun, M.; Peng, F. Hydrogen sulfide alleviates waterlogging-induced damage in peach seedlings via enhancing antioxidative system and inhibiting ethylene synthesis. Front Plant Sci. 2020, 11, 696. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, L.; Barbero, G.F.; Geerinck, J.; Tilleman, S.; Grunewald, W.; Pérez, A.C.; Chico, J.M.; Bossche, R.V.; Sewell, J.; Gil, E.; et al. NINJA connects the co-repressor TOPLESS to jasmonate signalling. Nature 2010, 464, 788–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, T.; Song, S.; Ren, Q.; Wu, D.; Huang, H.; Chen, Y.; Fan, M.; Peng, W.; Ren, C.; Xie, D. The Jasmonate-ZIM-domain proteins interact with the WD-Repeat/bHLH/MYB complexes to regulate Jasmonate-mediated anthocyanin accumulation and trichome initiation in Arabidopsis thaliana. Plant Cell 2011, 23, 1795–1814. [Google Scholar] [CrossRef] [Green Version]
- Qi, T.; Huang, H.; Song, S.; Xie, D. Regulation of Jasmonate-mediated stamen development and seed production by a bHLH-MYB complex in Arabidopsis. Plant Cell 2015, 27, 1620–1633. [Google Scholar] [CrossRef] [Green Version]
- Qi, T.; Wang, J.; Huang, H.; Liu, B.; Gao, H.; Liu, Y.; Song, S.; Xie, D. Regulation of Jasmonate-Induced Leaf Senescence by Antagonism between bHLH Subgroup IIIe and IIId Factors in Arabidopsis. Plant Cell 2015, 27, 1634–1649. [Google Scholar] [CrossRef] [Green Version]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Hu, Y.; Zhang, G.; Jiang, Y.; Chen, X.; Yu, D. Jasmonate negatively regulates stomatal development in Arabidopsis cotyledons. Plant Physiol. 2018, 176, 2871–2885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, G.; Zhou, L.; Wang, Y.; Zhang, G.; Chen, X. Hydrogen sulfide acts downstream of jasmonic acid to inhibit stomatal development in Arabidopsis. Planta 2020, 251, 42. [Google Scholar] [CrossRef]
- He, H.; He, L. Regulation of gaseous signaling molecules on proline metabolism in plants. Plant Cell Rep. 2018, 37, 387–392. [Google Scholar] [CrossRef]
- Yao, Y.; Yang, Y.; Li, C.; Huang, D.; Zhang, J.; Wang, C.; Li, W.; Wang, N.; Deng, Y.; Liao, W. Research progress on the functions of gasotransmitters in plant responses to abiotic stresses. Plants 2019, 8, 605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, B.; Qiao, Z.; Zhang, L.; Li, H.; Pei, Y. Hydrogen sulfide and proline cooperate to alleviate cadmium stress in foxtail millet seedlings. Plant Physiol. Bioch. 2016, 109, 293–299. [Google Scholar] [CrossRef] [PubMed]
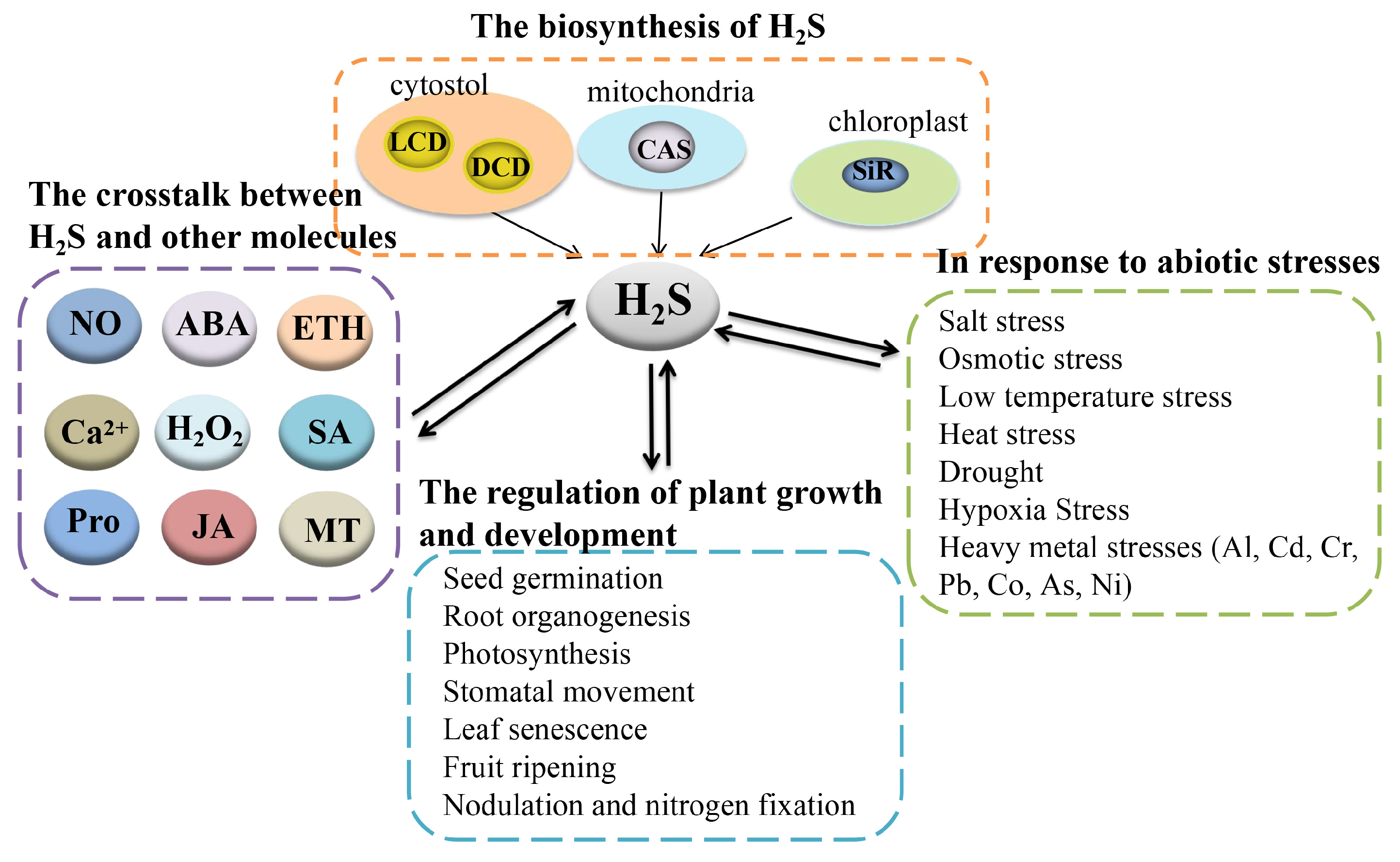
Figure 1.
The summary of the biosynthesis of H2S, the crosstalk between H2S and other molecules, the regulation of plant growth and development, and the response to abiotic stresses by H2S. H2S, hydrogen sulfide; LCD, L-cysteine desulfhydrase; DCD, D-cysteine desulfhydrase; CAS, β-cyanoalanine synthase; SiR, sulfite reductase; NO, nitric oxide; ABA, abscisic acid; Ca2+, calcium ion; H2O2, hydrogen peroxide; SA, salicylic acid; JA, jasmonic acid; Pro, proline; MT, melatonin; Al, aluminum; Cd, cadmium; Cr, chromium; Pb, lead; Co, cobalt; As, arsenic; Ni, nickel.
Figure 1.
The summary of the biosynthesis of H2S, the crosstalk between H2S and other molecules, the regulation of plant growth and development, and the response to abiotic stresses by H2S. H2S, hydrogen sulfide; LCD, L-cysteine desulfhydrase; DCD, D-cysteine desulfhydrase; CAS, β-cyanoalanine synthase; SiR, sulfite reductase; NO, nitric oxide; ABA, abscisic acid; Ca2+, calcium ion; H2O2, hydrogen peroxide; SA, salicylic acid; JA, jasmonic acid; Pro, proline; MT, melatonin; Al, aluminum; Cd, cadmium; Cr, chromium; Pb, lead; Co, cobalt; As, arsenic; Ni, nickel.
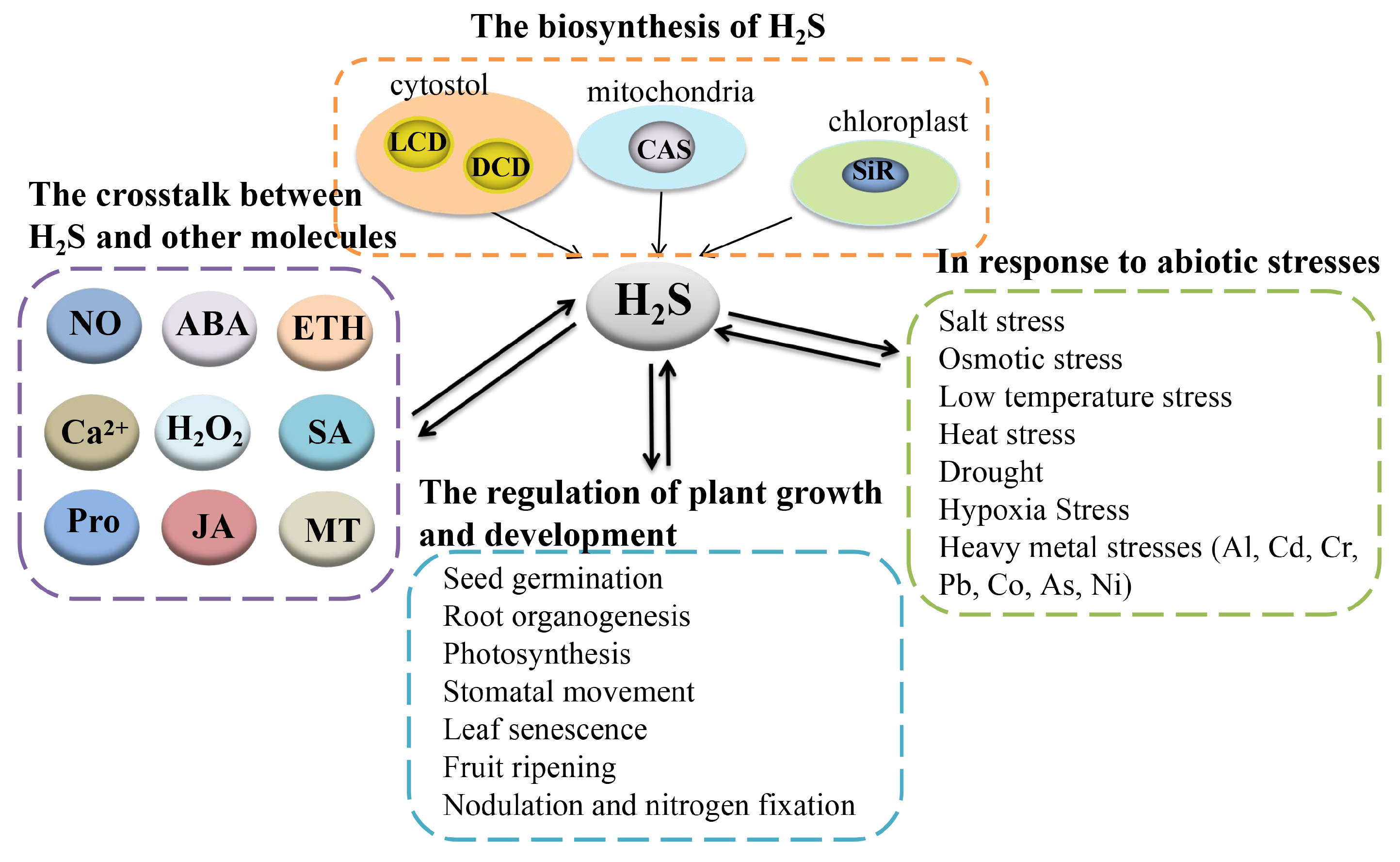
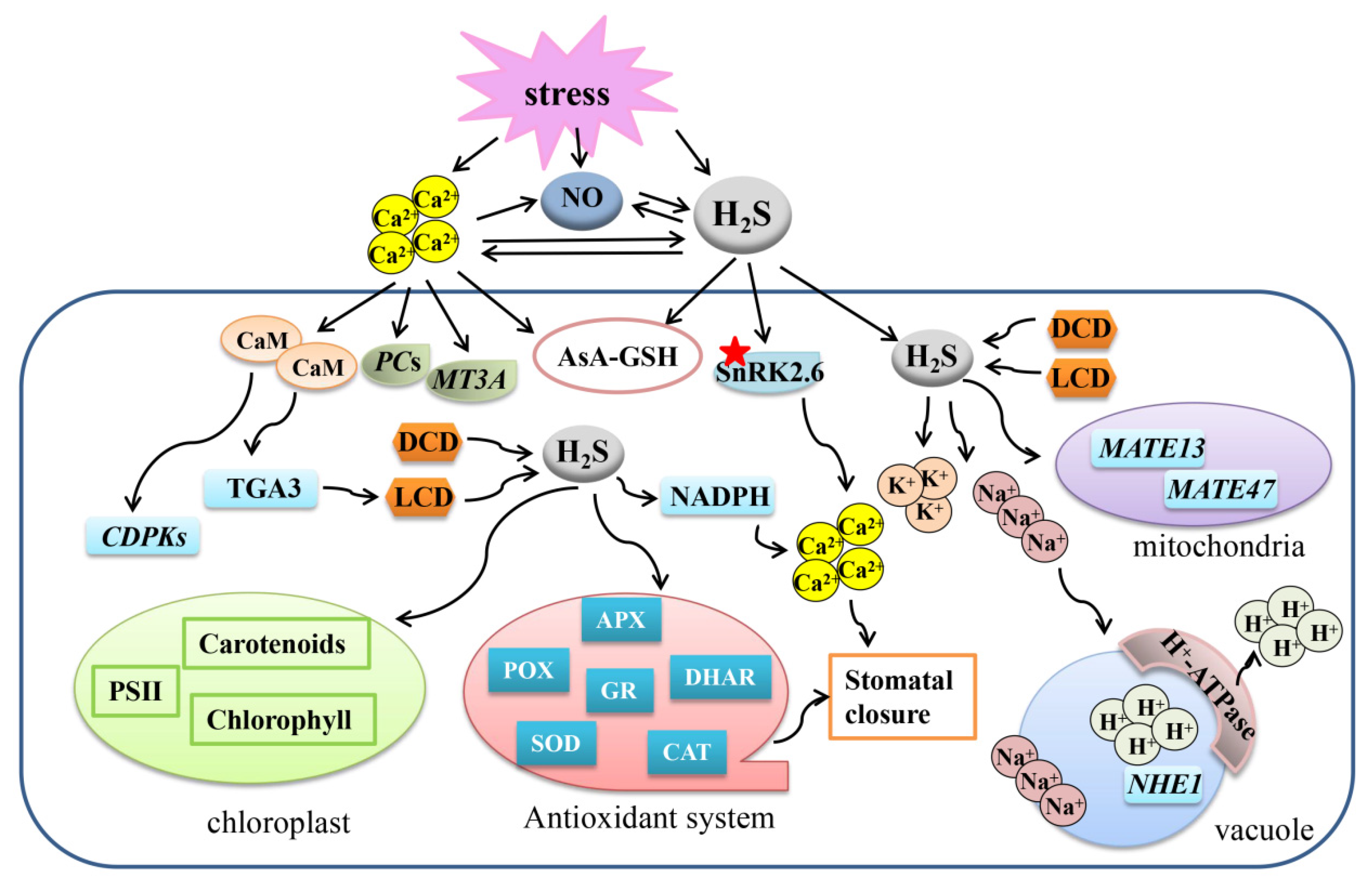
Figure 2.
Overview for the mechanisms of the crosstalk between Ca2+, NO, and H2S to regulate plant response to abiotic stresses. A protein marked with a red asterisk means that the protein can be persulfided. Ca2+, calcium ion; NO, nitric oxide; H2S, hydrogen sulfide; LCD, L-cysteine desulfhydrase; DCD, D-cysteine desulfhydrase; APX, ascorbate peroxidase; SOD, superoxide dismutase; GR, glutathione reductase; POD, peroxidase; CAT, catalase; CaM, calmodulin; PCs, phytochelatin synthase; MT3A, metallothionein-like type 3; CDPKs, Ca2+-dependent protein kinases; AsA-GSH, ascorbate-glutathione cycle; DHAR, dehydroascorbate reductase; POD, peroxidase; CAT, catalase.
Figure 2.
Overview for the mechanisms of the crosstalk between Ca2+, NO, and H2S to regulate plant response to abiotic stresses. A protein marked with a red asterisk means that the protein can be persulfided. Ca2+, calcium ion; NO, nitric oxide; H2S, hydrogen sulfide; LCD, L-cysteine desulfhydrase; DCD, D-cysteine desulfhydrase; APX, ascorbate peroxidase; SOD, superoxide dismutase; GR, glutathione reductase; POD, peroxidase; CAT, catalase; CaM, calmodulin; PCs, phytochelatin synthase; MT3A, metallothionein-like type 3; CDPKs, Ca2+-dependent protein kinases; AsA-GSH, ascorbate-glutathione cycle; DHAR, dehydroascorbate reductase; POD, peroxidase; CAT, catalase.
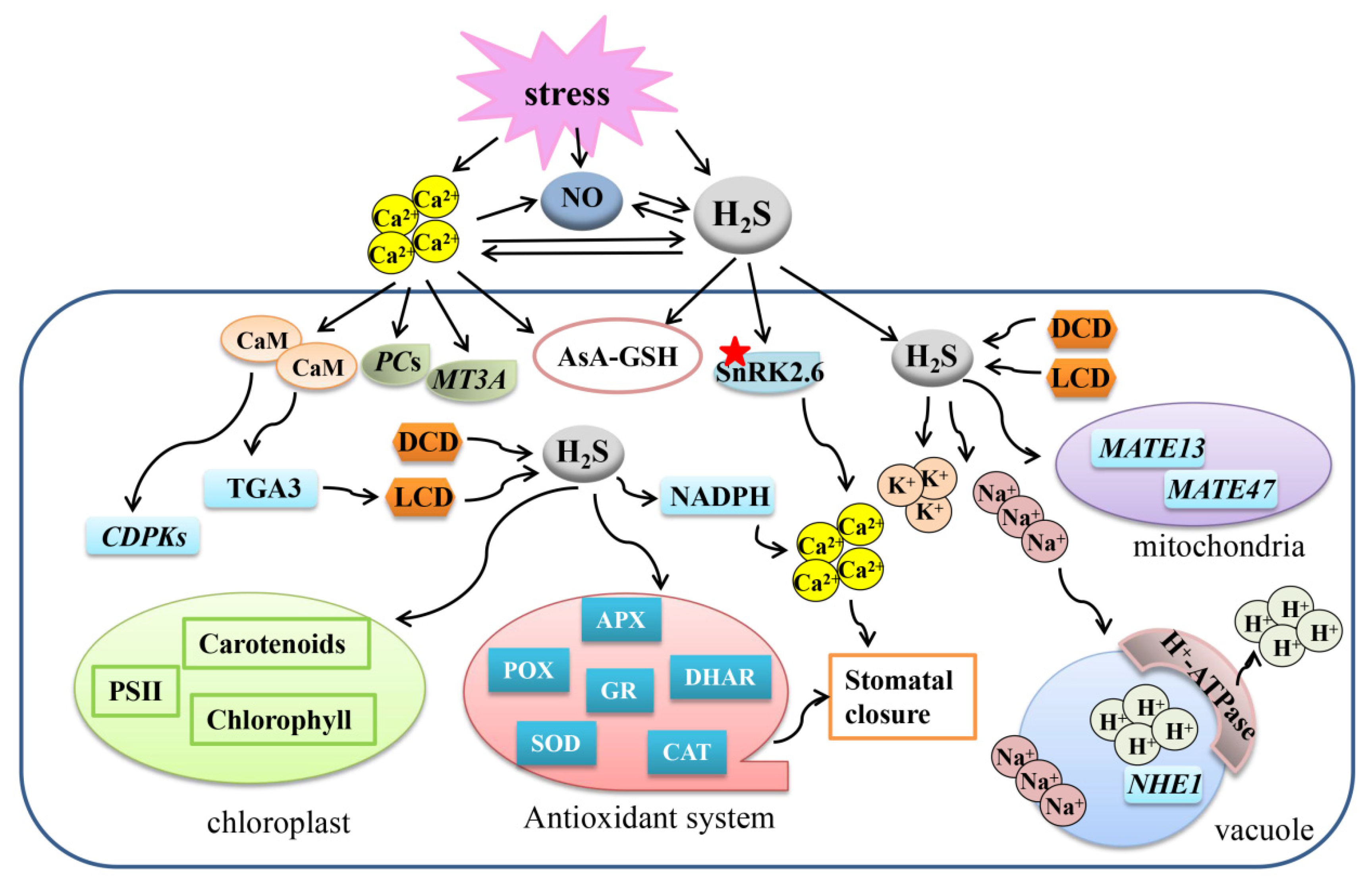
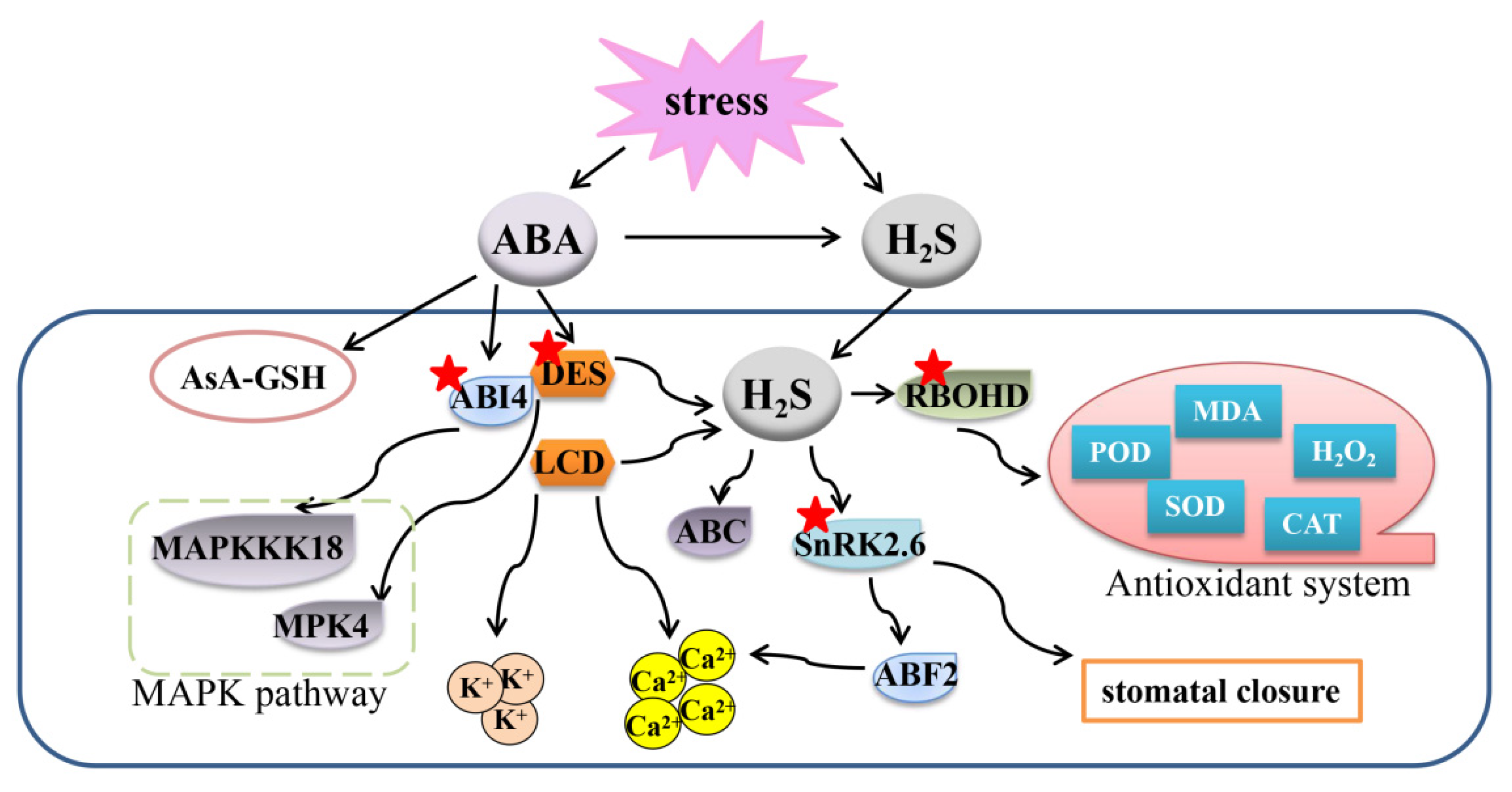
Figure 3.
Overview of the mechanisms of the crosstalk between ABA and H2S to regulate plant response to abiotic stresses. A gene or protein marked with a red asterisk means that the protein can be persulfided. H2S, hydrogen sulfide; ABA, abscisic acid; ABF2, ABA response element-binding factor 2; AsA-GSH, ascorbate-glutathione cycle; SnRK2.6, snf1-related protein kinase 2.6; RBOHD, respiratory burst oxidase homolog protein d; MDA, malondialdehyde, ABI4, abscisic acid insensitive 4; MAPK, mitogen-activated protein kinase.
Figure 3.
Overview of the mechanisms of the crosstalk between ABA and H2S to regulate plant response to abiotic stresses. A gene or protein marked with a red asterisk means that the protein can be persulfided. H2S, hydrogen sulfide; ABA, abscisic acid; ABF2, ABA response element-binding factor 2; AsA-GSH, ascorbate-glutathione cycle; SnRK2.6, snf1-related protein kinase 2.6; RBOHD, respiratory burst oxidase homolog protein d; MDA, malondialdehyde, ABI4, abscisic acid insensitive 4; MAPK, mitogen-activated protein kinase.
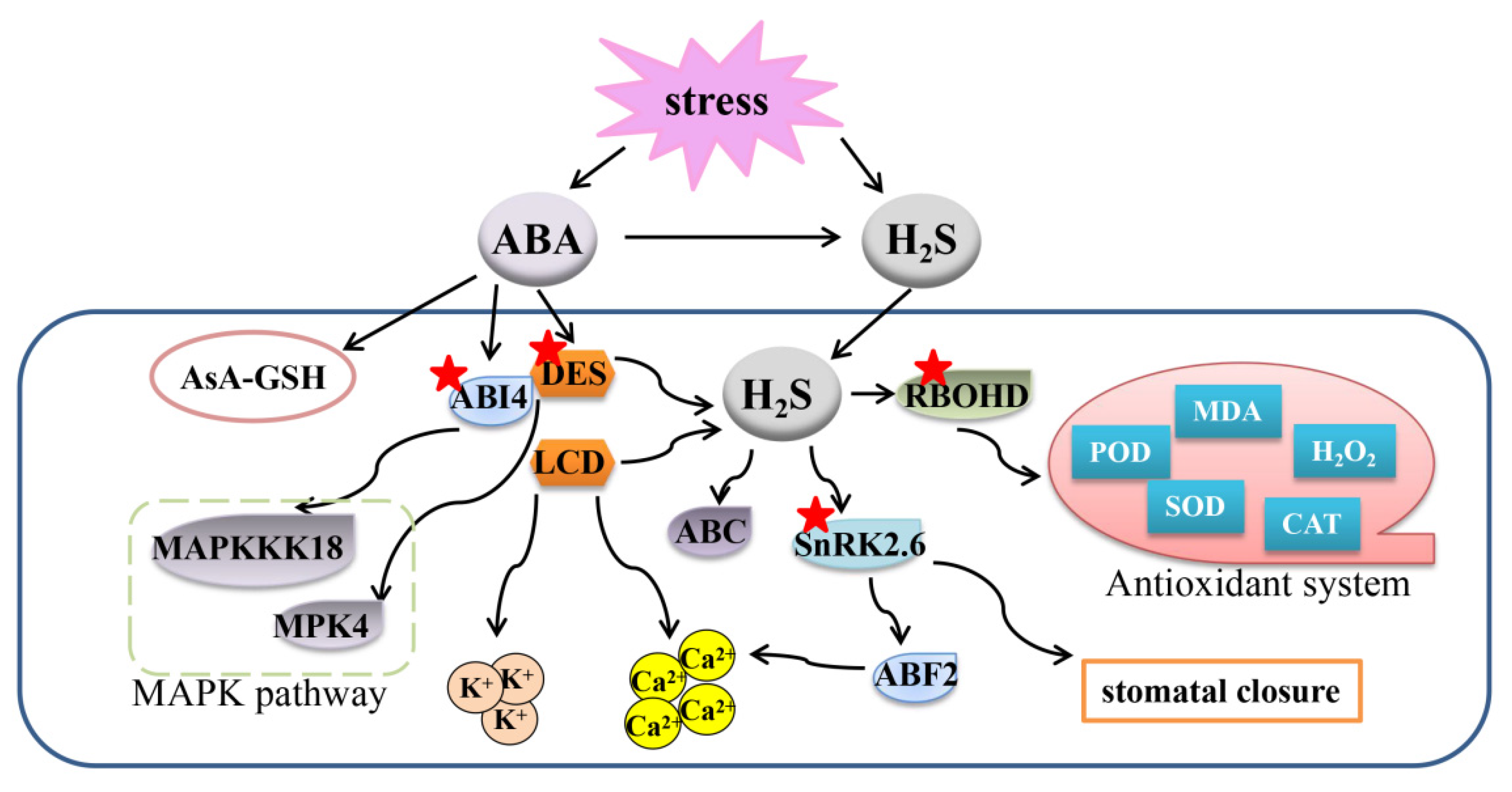
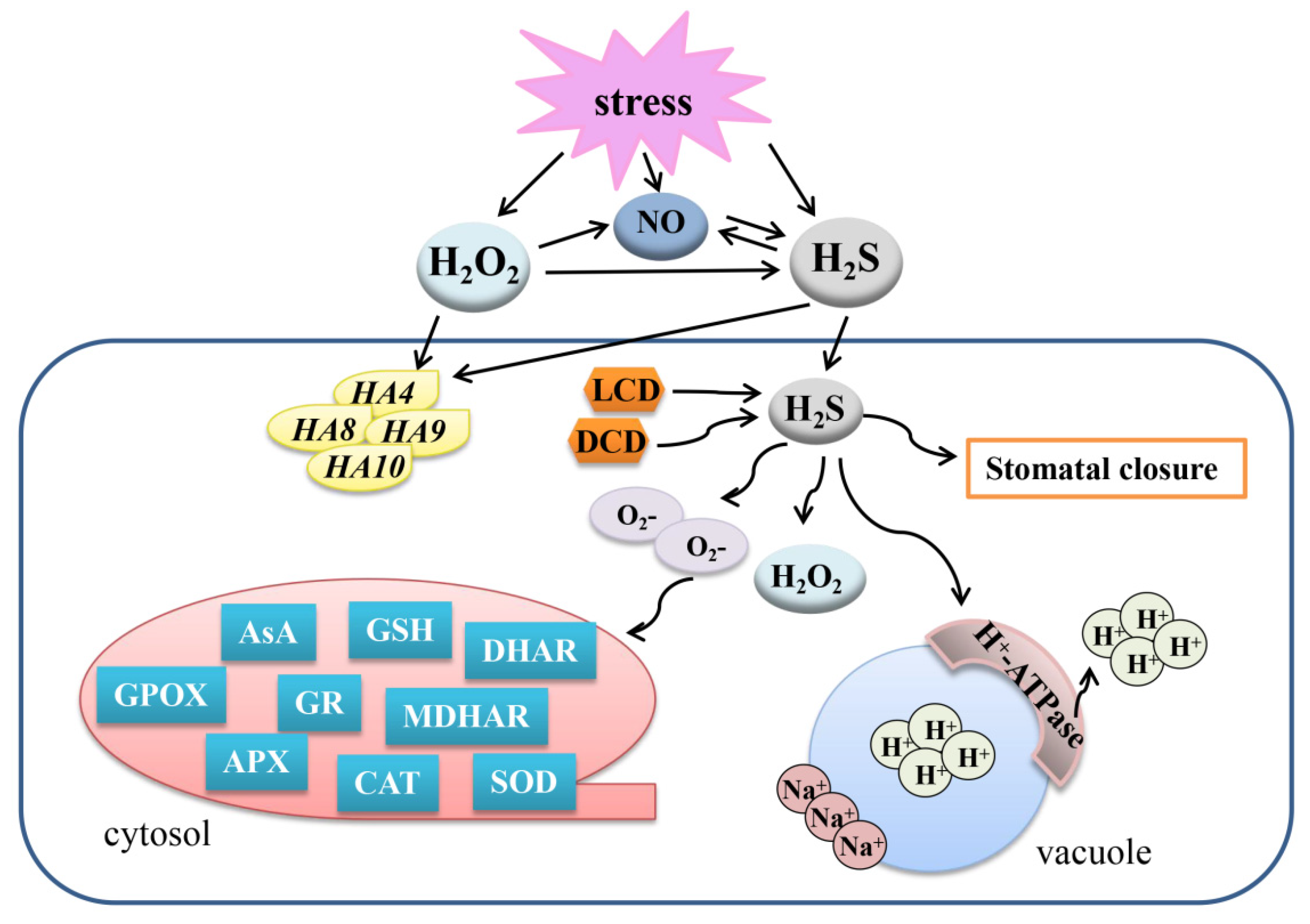
Figure 4.
Overview of the mechanisms of the crosstalk between H2S, NO, and H2O2 to regulate plant response to abiotic stresses. H2S, hydrogen sulfide; H2O2, hydrogen peroxide; NO, nitric oxide; LCD, L-cysteine desulfhydrase; DCD, D-cysteine desulfhydrase; AsA, ascorbic acid; GSH, glutathione; GR, glutathione reductase; APX, ascorbate peroxidase; GPOX, guaiacol peroxidase; CAT, catalase; SOD, superoxide dismutase; DHAR, dehydroascorbate reductase; MDHAR, monodehydroascorbate reductase.
Figure 4.
Overview of the mechanisms of the crosstalk between H2S, NO, and H2O2 to regulate plant response to abiotic stresses. H2S, hydrogen sulfide; H2O2, hydrogen peroxide; NO, nitric oxide; LCD, L-cysteine desulfhydrase; DCD, D-cysteine desulfhydrase; AsA, ascorbic acid; GSH, glutathione; GR, glutathione reductase; APX, ascorbate peroxidase; GPOX, guaiacol peroxidase; CAT, catalase; SOD, superoxide dismutase; DHAR, dehydroascorbate reductase; MDHAR, monodehydroascorbate reductase.
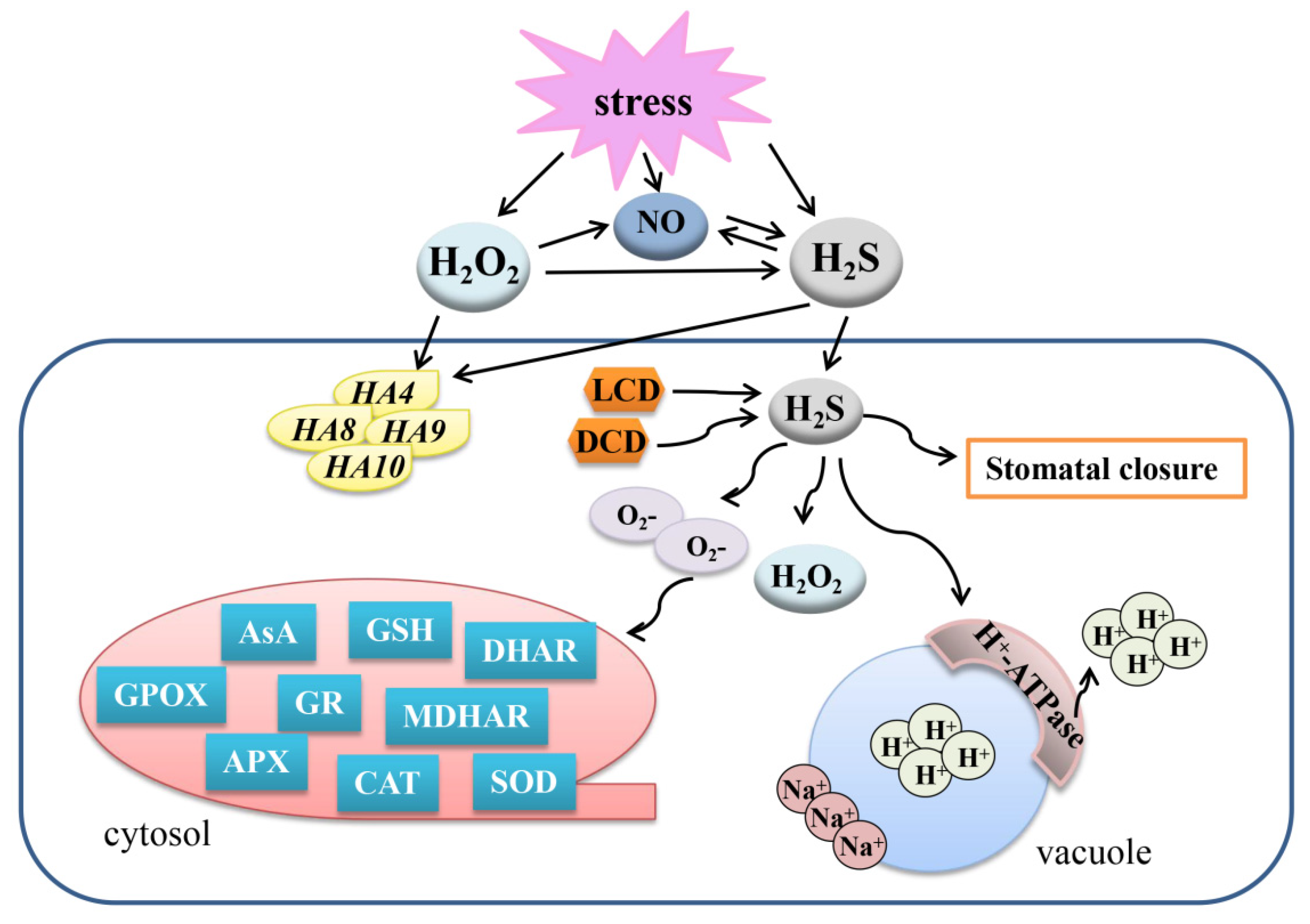
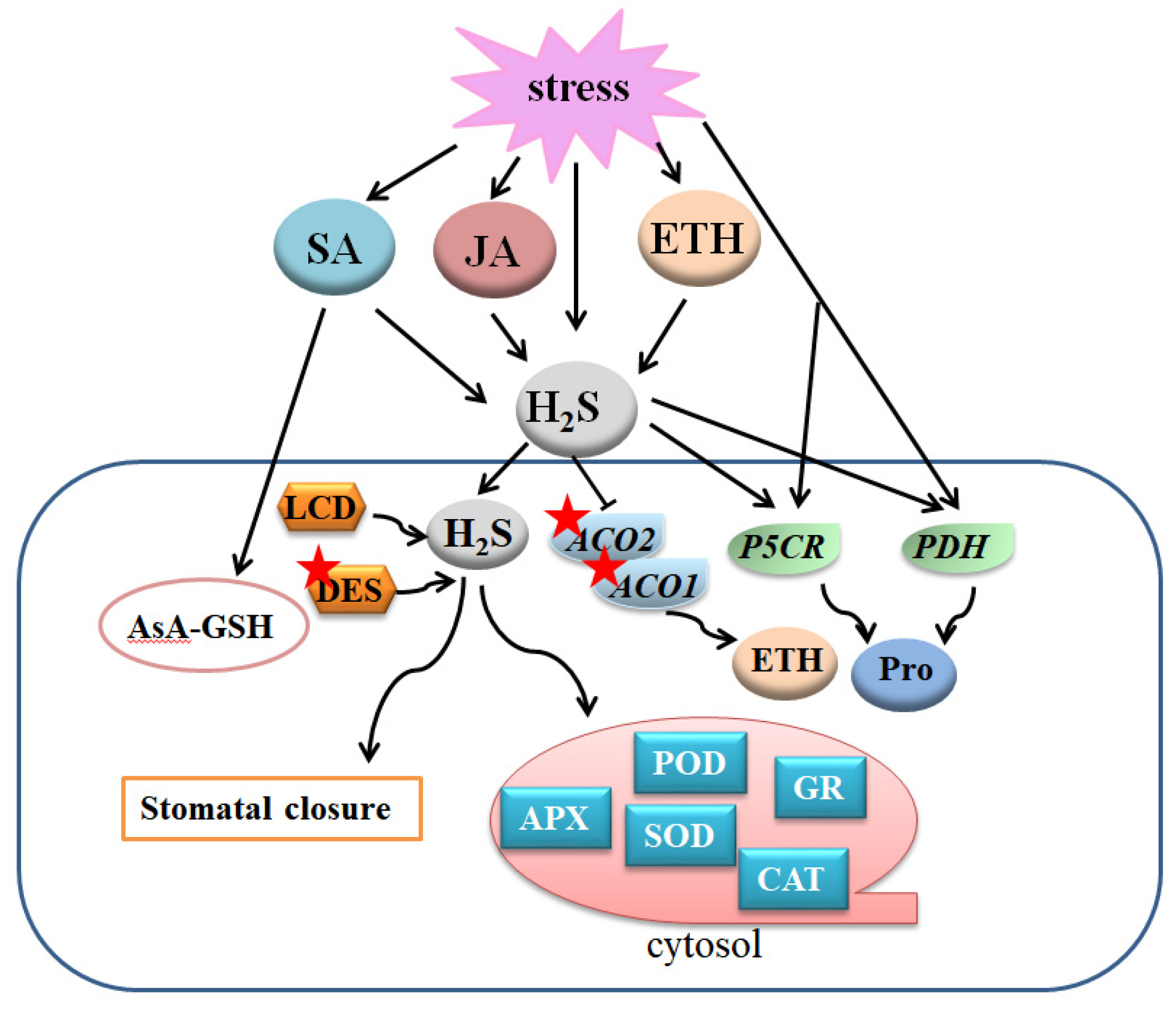
Figure 5.
Overview of the mechanisms of the crosstalk between H2S and JA, SA, ETH, and Pro to regulate plant response to abiotic stresses. A gene or protein marked with a red asterisk means that the protein can be persulfided. H2S, hydrogen sulfide; LCD, L-cysteine desulfhydrase; DES, desulfhydrase; SA, salicylic acid; JA, jasmonic acid; ETH, ethylene; Pro, proline; APX, ascorbate peroxidase; SOD, superoxide dismutase; GR, glutathione reductase; POD, peroxidase; CAT, catalase; ACO1, 1-aminocyclopropane-1-carboxylic acid oxidase 1; ACO2, 1-aminocyclopropane-1-carboxylic acid oxidase 2; P5CR, proline-5-carboxylate reductase; PDH, proline dehydrogenase.
Figure 5.
Overview of the mechanisms of the crosstalk between H2S and JA, SA, ETH, and Pro to regulate plant response to abiotic stresses. A gene or protein marked with a red asterisk means that the protein can be persulfided. H2S, hydrogen sulfide; LCD, L-cysteine desulfhydrase; DES, desulfhydrase; SA, salicylic acid; JA, jasmonic acid; ETH, ethylene; Pro, proline; APX, ascorbate peroxidase; SOD, superoxide dismutase; GR, glutathione reductase; POD, peroxidase; CAT, catalase; ACO1, 1-aminocyclopropane-1-carboxylic acid oxidase 1; ACO2, 1-aminocyclopropane-1-carboxylic acid oxidase 2; P5CR, proline-5-carboxylate reductase; PDH, proline dehydrogenase.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Genes regulated by H2S and other molecules under abiotic stress conditions.
| Crosstalk between H2S and other Molecules | Stresses | Plant Species | Tissue | Regulated Genes | References |
|---|---|---|---|---|---|
| H2S and NO | salt stress | Medicago sativa | seeds | APX-1, APX-2, and Cu/Zn-SOD | [39] |
| Hordeum vulgare L. | seedlings | HvHA, HvVHA-β, HvSOS1, HvVNHX2, HvAKT1 and HvHAK4 | [22] | ||
| Solanum lycopersicum | seedlings | SlL-DES, SlCAS and SlCS | [40] | ||
| drought | M. sativa L. | leaves | GST17, Cu/ZnSOD, FeSOD, NR, cAPX, PIP | [48] | |
| hypoxia stress | Zea mays L. | seedlings | P4H, ADH, CRT1, GS, CYP51 and ME | [43] | |
| cadmium stress | M. sativa L. | seedlings | Cu/Zn–SOD, APX and POD | [31] | |
| cobalt stress | Triticum aestivum L. | seedlings | RbcL | [32] | |
| aluminum stress | Glycine max L. | seedlings | MATE13, MATE47, MATE58, MATE74, MATE79, MATE84, and MATE87 | [34] | |
| H2S and ABA | drought | Oryza sativa L. | seedlings | NCED2, NCED3, NCED5, AREB1, AREB8, bZIP23 and LEA3 | [54] |
| Arabidopsis | seedlings | TPC1, GORK, SKOR, KCO1, MYP5, ACA9, ACA11, CAX1, SLAC1, AKT1A, KT2, KC1 and KAT1 | [55] | ||
| T. aestivum L. | leaves and roots | TaZEP, TaNCED, TaAAO and TaSDR | [56] | ||
| Arabidopsis thaliana | - | MAPKs | [57] | ||
| chromium stress | A. thaliana | seedlings | LCD | [72] | |
| nickel stress | Cucurbita pepo L. | seedlings | CDPK and PCS1 | [74] | |
| H2S and Ca2+ | chromium stress | Setaria italica | seedlings | MT3A, PCS, CaM, CBL and CDPK | [71] |
| H2S-H2O2 | cadmium stress | Brassica rapa. | seedlings | Br_UPB1A, Br_UPB1B↑; Bra035235, Bra033551, Bra006423, ra023639 | [89] |
| cadmium stress | Cucumis sativus L. | roots | CsVHA-A, CsVHA-B, CsVHA-a1, CsVHA-a2, CsVHA-a3, CsVHA-c1, CsVHA-c2 and CsVHA-c3 | [82] | |
| H2S, NO and H2O2 | salt or low temperature | C. sativus L. | roots | CsHA1, CsHA2, CsH4, CsH8, CsH9 and CsHA10 | [83] |
| dehydration | Trifolium repens | seedlings | bZIP37, bZIP107, DREB2, DREB4 and WRKY108715 | [84] | |
| H2S and ETH | osmotic stress | S. lycopersicum | seedlings | LeACO1 and LeACO2 | [94] |
| H2S and Pro | cadmium stress | Foxtail millet | seedlings | PDH and P5CR | [105] |
APX, ascorbate peroxidase; SOD, superoxide dismutase; HA, H+-ATPase; VNHX2, vacuolar Na+/H+ antiporter; VHA-β, H+-ATPase subunit β; HAK4, high-affinity K+ uptake system; L-DES, L-cysteine desulfhydrase; CAS, β-cyanoalanine synthase; CS, L-cysteine synthase; P4H, prolyl 4-hydroxylase; ADH, alcohol dehydrogenase; CRT1, calcium binding protein; CYP51, cytochrome P450 14a-sterol demethylase; GS, glutamate synthase 1; ME, NADP-dependent malic enzyme; POD, peroxidase; rbcL, rubisco large subunit; NCED, 9′-cis-epoxycarotenoid dioxygenase; TPC1, two pore segment channel 1; GORK, guard cell outward-rectifying Kþ channel; SKOR, SKI family transcriptional corepressor; KCO, outward-rectifying K+ channel; ACA, adenylyl cyclase-associated protein; CAX, calcium exchanger; SLAC1, slow anion channel associated 1; AKT, Arabidopsis potassium transporter; KC1, potassium channel 1; KAT1, potassium channel in Arabidopsis thaliana 1; ZEP, zeaxanthin epoxidase; AAO, abscisic aldehyde oxidase; SDR, short-chain dehydrogenase; MAPK, mitogen-activated protein kinase; LCD, L-cysteine desulfhydrase; CDPK, Ca2+-dependent protein kinase; PCS, phytochelatin; CaM, calmodulin; CBL, calcineurin B-like; ACO, 1-aminocyclopropane-1-carboxylic oxidase; PDH, proline dehydrogenase; P5CR, proline-5-carboxylate reductase.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, C.; Deng, Y.; Liu, Z.; Liao, W. Hydrogen Sulfide in Plants: Crosstalk with Other Signal Molecules in Response to Abiotic Stresses. Int. J. Mol. Sci. 2021, 22, 12068. https://doi.org/10.3390/ijms222112068
AMA Style
Wang C, Deng Y, Liu Z, Liao W. Hydrogen Sulfide in Plants: Crosstalk with Other Signal Molecules in Response to Abiotic Stresses. International Journal of Molecular Sciences. 2021; 22(21):12068. https://doi.org/10.3390/ijms222112068
Chicago/Turabian StyleWang, Chunlei, Yuzheng Deng, Zesheng Liu, and Weibiao Liao. 2021. "Hydrogen Sulfide in Plants: Crosstalk with Other Signal Molecules in Response to Abiotic Stresses" International Journal of Molecular Sciences 22, no. 21: 12068. https://doi.org/10.3390/ijms222112068
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.