
Daily Lifestyle and Inflammatory Skin Diseases

Abstract
1. Introduction
2. Pathogenesis of Inflammatory Skin Diseases
2.1. Psoriasis
2.2. Atopic Dermatitis
2.3. Contact Dermatitis
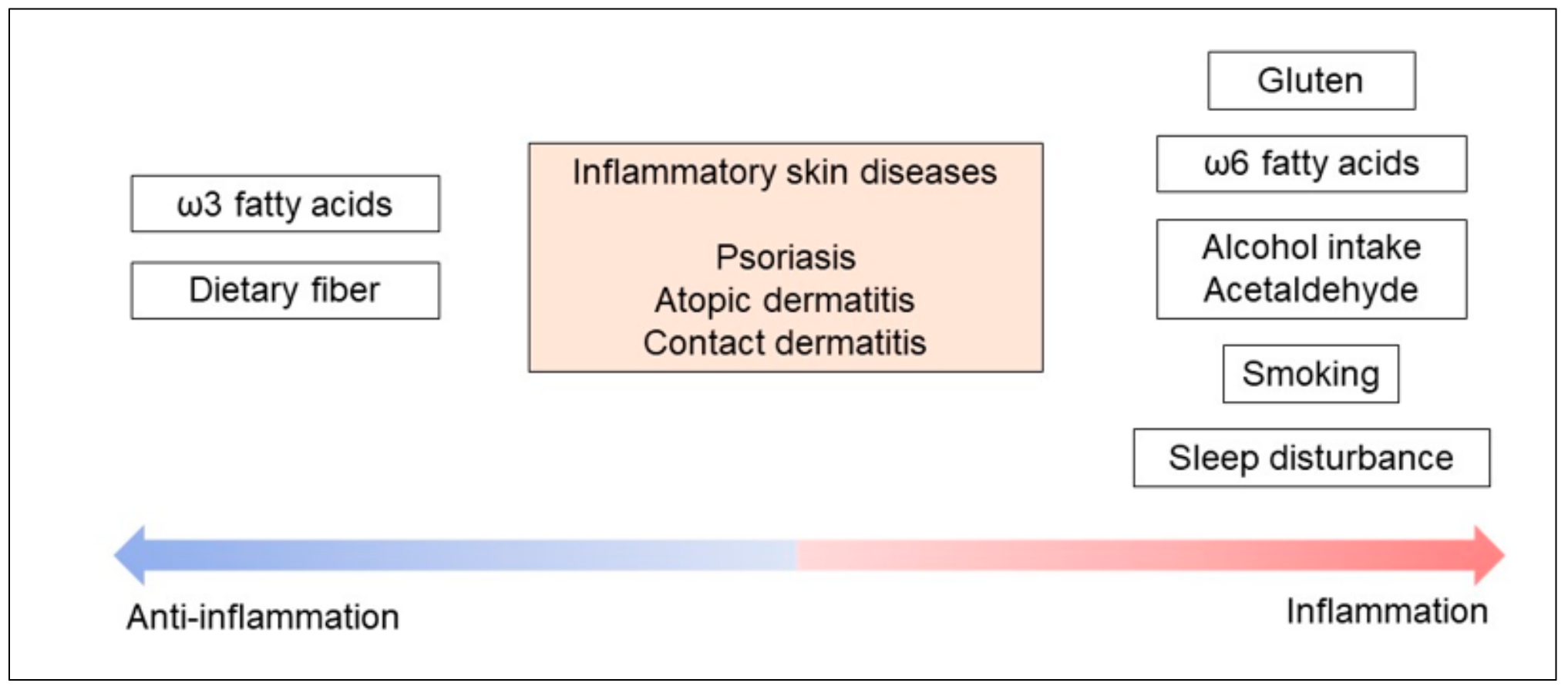
3. Daily Lifestyle and Inflammatory Skin Diseases
3.1. Gluten
3.2. Dietary Fiber
3.3. Omega (ω)-3 and Omega (ω)-6 Fatty Acids
3.4. Smoking
3.5. Alcohol
3.6. Sleep
3.7. Obesity
3.8. Others
4. Summary of Inflammatory Reactions to Daily Lifestyle Factors
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| CCL | C-C motif chemokine ligand |
| CXCL | C-X-C motif ligand |
| DC | dendritic cell |
| EPA | eicosapentaenoic acid |
| DHA | docosahexaenoic acid |
| IFN | interferon |
| IL | interleukine |
| LTB4 | leukotriene B4 |
| MaR1 | maresin 1 |
| PGE2 | prostaglaind E2 |
| RvD1 | resolvin D1 |
| RvE1 | resolvin E1 |
| TNF | tumor necrosis factor |
| TSLP | Thymic stromal lymphopoietin |
| TxA2 | thromoboxane A2 |
References
- McAlpine, C.S.; Swirski, F.K. Circadian Influence on Metabolism and Inflammation in Atherosclerosis. Circ. Res. 2016, 119, 131–141. [Google Scholar] [CrossRef]
- Chan, C.W.H.; Wong, R.S.; Law, P.T.W.; Wong, C.L.; Tsui, S.K.W.; Tang, W.P.Y.; Sit, J.W.H. Environmental Factors Associated with Altered Gut Microbiota in Children with Eczema: A Systematic Review. Int. J. Mol. Sci. 2016, 17, 1147. [Google Scholar] [CrossRef]
- Katsiki, N.; Purrello, F.; Tsioufis, C.; Mikhailidis, D.P. Cardiovascular disease prevention strategies for type 2 diabetes mellitus. Expert Opin. Pharmacother. 2017, 18, 1243–1260. [Google Scholar] [CrossRef] [PubMed]
- Waggoner, S.N. Circadian Rhythms in Immunity. Curr. Allergy Asthma Rep. 2020, 20, 2. [Google Scholar] [CrossRef] [PubMed]
- Kabashima, K.; Honda, T.; Ginhoux, F.; Egawa, G. The immunological anatomy of the skin. Nat. Rev. Immunol. 2019, 19, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Dainichi, T.; Kitoh, A.; Otsuka, A.; Nakajima, S.; Nomura, T.; Kaplan, D.H.; Kabashima, K. The epithelial immune microenvironment (EIME) in atopic dermatitis and psoriasis. Nat. Immunol. 2018, 19, 1286–1298. [Google Scholar] [CrossRef]
- Dainichi, T.; Matsumoto, R.; Mostafa, A.; Kabashima, K. Immune Control by TRAF6-Mediated Pathways of Epithelial Cells in the EIME (Epithelial Immune Microenvironment). Front. Immunol. 2019, 10, 1107. [Google Scholar] [CrossRef]
- Sawada, Y.; Gallo, R.L. Role of Epigenetics in the Regulation of Immune Functions of the Skin. J. Investig. Dermatol. 2021, 141, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Addolorato, G.; Parente, A.; de Lorenzi, G.; D’Angelo Di Paola, M.E.; Abenavoli, L.; Leggio, L.; Capristo, E.; De Simone, C.; Rotoli, M.; Rapaccini, G.L.; et al. Rapid regression of psoriasis in a coeliac patient after gluten-free diet. A case report and review of the literature. Digestion 2003, 68, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Barrea, L.; Macchia, P.E.; Tarantino, G.; Di Somma, C.; Pane, E.; Balato, N.; Napolitano, M.; Colao, A.; Savastano, S. Nutrition: A key environmental dietary factor in clinical severity and cardio-metabolic risk in psoriatic male patients evaluated by 7-day food-frequency questionnaire. J. Transl. Med. 2015, 13, 303. [Google Scholar] [CrossRef]
- Tokura, Y. Extrinsic and intrinsic types of atopic dermatitis. J. Dermatol. Sci. 2010, 58, 1–7. [Google Scholar] [CrossRef]
- Afifi, L.; Danesh, M.J.; Lee, K.M.; Beroukhim, K.; Farahnik, B.; Ahn, R.S.; Yan, D.; Singh, R.K.; Nakamura, M.; Koo, J.; et al. Dietary Behaviors in Psoriasis: Patient-Reported Outcomes from a U.S. National Survey. Dermatol. Ther. 2017, 7, 227–242. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Behrens, F.; Mease, P.J.; Kavanaugh, A.; Ritchlin, C.; Nash, P.; Masmitja, J.G.; Goupille, P.; Korotaeva, T.; Gottlieb, A.B.; et al. Secukinumab versus adalimumab for treatment of active psoriatic arthritis (EXCEED): A double-blind, parallel-group, randomised, active-controlled, phase 3b trial. Lancet 2020, 395, 1496–1505. [Google Scholar] [CrossRef]
- Reich, K.; Armstrong, A.W.; Langley, R.G.; Flavin, S.; Randazzo, B.; Li, S.; Hsu, M.-C.; Branigan, P.; Blauvelt, A. Guselkumab versus secukinumab for the treatment of moderate-to-severe psoriasis (ECLIPSE): Results from a phase 3, randomised controlled trial. Lancet 2019, 394, 831–839. [Google Scholar] [CrossRef]
- Yoshiki, R.; Kabashima, K.; Honda, T.; Nakamizo, S.; Sawada, Y.; Sugita, K.; Yoshioka, H.; Ohmori, S.; Malissen, B.; Tokura, Y.; et al. IL-23 from Langerhans Cells Is Required for the Development of Imiquimod-Induced Psoriasis-Like Dermatitis by Induction of IL-17A-Producing γδ T Cells. J. Investig. Dermatol. 2014, 134, 1912–1921. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Li, S.; Ying, S.; Tang, S.; Ding, Y.; Li, Y.; Qiao, J.; Fang, H. The IL-23/IL-17 Pathway in Inflammatory Skin Diseases: From Bench to Bedside. Front. Immunol. 2020, 11, 594735. [Google Scholar] [CrossRef]
- Bissonnette, R.; Luger, T.; Thaçi, D.; Toth, D.; Messina, I.; You, R.; Guana, A.; Fox, T.; Papavassilis, C.; Gilloteau, I.; et al. Secukinumab sustains good efficacy and favourable safety in moderate-to-severe psoriasis after up to 3 years of treatment: Results from a double-blind extension study. Br. J. Dermatol. 2017, 177, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Saunte, D.; Mrowietz, U.; Puig, L.; Zachariae, C. Candida infections in patients with psoriasis and psoriatic arthritis treated with interleukin-17 inhibitors and their practical management. Br. J. Dermatol. 2017, 177, 47–62. [Google Scholar] [CrossRef]
- Egawa, G.; Kabashima, K. Barrier dysfunction in the skin allergy. Allergol. Int. 2018, 67, 3–11. [Google Scholar] [CrossRef]
- Palmer, C.N.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.; et al. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef]
- Osawa, R.; Konno, S.; Akiyama, M.; Nemoto-Hasebe, I.; Nomura, T.; Nomura, Y.; Abe, R.; Sandilands, A.; McLean, W.H.I.; Hizawa, N.; et al. Japanese-Specific Filaggrin Gene Mutations in Japanese Patients Suffering from Atopic Eczema and Asthma. J. Investig. Dermatol. 2010, 130, 2834–2836. [Google Scholar] [CrossRef] [PubMed]
- Thyssen, J.P.; Kezic, S. Causes of epidermal filaggrin reduction and their role in the pathogenesis of atopic dermatitis. J. Allergy Clin. Immunol. 2014, 134, 792–799. [Google Scholar] [CrossRef]
- Nakajima, S.; Nomura, T.; Common, J.; Kabashima, K. Insights into atopic dermatitis gained from genetically defined mouse models. J. Allergy Clin. Immunol. 2019, 143, 13–25. [Google Scholar] [CrossRef]
- Otsuka, A.; Tanioka, M.; Nakagawa, Y.; Honda, T.; Ikoma, A.; Miyachi, Y.; Kabashima, K. Effects of cyclosporine on pruritus and serum IL-31 levels in patients with atopic dermatitis. Eur. J. Dermatol. EJD 2011, 21, 816–817. [Google Scholar] [CrossRef]
- Sonkoly, E.; Muller, A.; Lauerma, A.I.; Pivarcsi, A.; Soto, H.; Kemeny, L.; Alenius, H.; Dieu-Nosjean, M.-C.; Meller, S.; Rieker, J.; et al. IL-31: A new link between T cells and pruritus in atopic skin inflammation. J. Allergy Clin. Immunol. 2006, 117, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Beck, L.A.; Thaçi, D.; Hamilton, J.D.; Graham, N.M.; Bieber, T.; Rocklin, R.; Ming, J.E.; Ren, H.; Kao, R.; Simpson, E.; et al. Dupilumab Treatment in Adults with Moderate-to-Severe Atopic Dermatitis. N. Engl. J. Med. 2014, 371, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Ruzicka, T.; Hanifin, J.M.; Furue, M.; Pulka, G.; Mlynarczyk, I.; Wollenberg, A.; Galus, R.; Etoh, T.; Mihara, R.; Yoshida, H.; et al. Anti–Interleukin-31 Receptor A Antibody for Atopic Dermatitis. N. Engl. J. Med. 2017, 376, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, H.; Hirasawa, N.; Asakawa, S.; Okita, K.; Tokura, Y. Intrinsic atopic dermatitis shows high serum nickel concentration. Allergol. Int. 2015, 64, 282–284. [Google Scholar] [CrossRef]
- Frings, V.G.; Müller, D.; Storz, G.; Rossi, A.; Sennefelder, H.; Adam, C.; Goebeler, M.; Groeber-Becker, F.K.; Schmidt, M. Improved metal allergen reactivity of artificial skin models by integration of Toll-like receptor 4-positive cells. Contact Dermat. 2019, 81, 254–261. [Google Scholar] [CrossRef]
- Natsuaki, Y.; Egawa, G.; Nakamizo, S.; Ono, S.; Hanakawa, S.; Okada, T.; Kusuba, N.; Otsuka, A.; Kitoh, A.; Honda, T.; et al. Perivascular leukocyte clusters are essential for efficient activation of effector T cells in the skin. Nat. Immunol. 2014, 15, 1064–1069. [Google Scholar] [CrossRef]
- Sawada, Y.; Honda, T.; Hanakawa, S.; Nakamizo, S.; Murata, T.; Ueharaguchi-Tanada, Y.; Ono, S.; Amano, W.; Nakajima, S.; Egawa, G.; et al. Resolvin E1 inhibits dendritic cell migration in the skin and attenuates contact hypersensitivity responses. J. Exp. Med. 2015, 212, 1921–1930. [Google Scholar] [CrossRef] [PubMed]
- Honda, T.; Egawa, G.; Grabbe, S.; Kabashima, K. Update of Immune Events in the Murine Contact Hypersensitivity Model: Toward the Understanding of Allergic Contact Dermatitis. J. Investig. Dermatol. 2013, 133, 303–315. [Google Scholar] [CrossRef]
- Sharma, N.; Bhatia, S.; Chunduri, V.; Kaur, S.; Sharma, S.; Kapoor, P.; Kumari, A.; Garg, M. Pathogenesis of Celiac Disease and Other Gluten Related Disorders in Wheat and Strategies for Mitigating Them. Front. Nutr. 2020, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Hagforsen, E.; Michaëlsson, G.; Åhs, S.; Hammarström, I.; Lundin, I.P. Gluten-free Diet in Psoriasis Patients with Antibodies to Gliadin Results in Decreased Expression of Tissue Transglutaminase and Fewer Ki67+ Cells in the Dermis. Acta Derm. Venereol. 2003, 83, 425–429. [Google Scholar] [CrossRef]
- Castellanos-Rubio, A.; Santin, I.; Irastorza, I.; Castaño, L.; Vitoria, J.C.; Bilbao, J.R. TH17 (and TH1) signatures of intestinal biopsies of CD patients in response to gliadin. Autoimmunity 2009, 42, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Ciacci, C.; Cavallaro, R.; Iovino, P.; Sabbatini, F.; Palumbo, A.; Amoruso, D.; Tortora, R.; Mazzacca, G. Allergy prevalence in adult celiac disease. J. Allergy Clin. Immunol. 2004, 113, 1199–1203. [Google Scholar] [CrossRef]
- Ress, K.; Annus, T.; Putnik, U.; Luts, K.; Uibo, R.; Uibo, O. Celiac Disease in Children with Atopic Dermatitis. Pediatr. Dermatol. 2014, 31, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Drucker, A.M.; Qureshi, A.A.; Thompson, J.M.; Li, T.; Cho, E. Gluten intake and risk of psoriasis, psoriatic arthritis, and atopic dermatitis among United States women. J. Am. Acad. Dermatol. 2020, 82, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Ebaugh, S.; Martens, A.; Gao, H.; Olson, E.; Ng, P.K.; Gangur, V. A Mouse Model of Anaphylaxis and Atopic Dermatitis to Salt-Soluble Wheat Protein Extract. Int. Arch. Allergy Immunol. 2017, 174, 7–16. [Google Scholar] [CrossRef]
- Kuroda, Y.; Yuki, T.; Takahashi, Y.; Sakaguchi, H.; Matsunaga, K.; Itagaki, H. Long form of thymic stromal lymphopoietin of keratinocytes is induced by protein allergens. J. Immunotoxicol. 2017, 14, 178–187. [Google Scholar] [CrossRef] [PubMed]
- D’Alcamo, A.; Mansueto, P.; Soresi, M.; Iacobucci, R.; La Blasca, F.; Geraci, G.; Cavataio, F.; Fayer, F.; Arini, A.; Di Stefano, L.; et al. Contact Dermatitis Due to Nickel Allergy in Patients Suffering from Non-Celiac Wheat Sensitivity. Nutrients 2017, 9, 103. [Google Scholar] [CrossRef] [PubMed]
- Halnes, I.; Baines, K.J.; Berthon, B.S.; MacDonald-Wicks, L.K.; Gibson, P.G.; Wood, L.G. Soluble Fibre Meal Challenge Reduces Airway Inflammation and Expression of GPR43 and GPR41 in Asthma. Nutrients 2017, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, H.; Mander, I.; Zhang, Z.; Armstrong, D.; Wine, E. Not All Fibers Are Born Equal; Variable Response to Dietary Fiber Subtypes in IBD. Front. Pediatr. 2021, 8, 620189. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Takahashi, K.; Abe, S.; Yamada, K.; Suzuki, M.; Masahisa, M.; Endo, M.; Abe, K.; Inoue, R.; Hoshi, H. Improvement of Psoriasis by Alteration of the Gut Environment by Oral Administration of Fucoidan from Cladosiphon Okamuranus. Mar. Drugs 2020, 18, 154. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Schwarz, A.; Philippsen, R.; Schwarz, T. Induction of Regulatory T Cells and Correction of Cytokine Disbalance by Short-Chain Fatty Acids: Implications for Psoriasis Therapy. J. Investig. Dermatol. 2021, 141, 95–104.e2. [Google Scholar] [CrossRef] [PubMed]
- Reddel, S.; Del Chierico, F.; Quagliariello, A.; Giancristoforo, S.; Vernocchi, P.; Russo, A.; Fiocchi, A.; Rossi, P.; Putignani, L.; El Hachem, M. Gut microbiota profile in children affected by atopic dermatitis and evaluation of intestinal persistence of a probiotic mixture. Sci. Rep. 2019, 9, 4996. [Google Scholar] [CrossRef]
- Kim, J.A.; Kim, S.C.; Kim, I.S.; Yu, D.Y.; Lee, S.H.; Lee, S.S.; Yun, C.-H.; Choi, I.S.; Cho, K.K. Anti-Inflammatory Effects of a Mixture of Lactic Acid Bacteria and Sodium Butyrate in Atopic Dermatitis Murine Model. J. Med. Food 2018, 21, 716–725. [Google Scholar] [CrossRef]
- Schwarz, A.; Bruhs, A.; Schwarz, T. The Short-Chain Fatty Acid Sodium Butyrate Functions as a Regulator of the Skin Immune System. J. Investig. Dermatol. 2017, 137, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Sasajima, N.; Ogasawara, T.; Takemura, N.; Fujiwara, R.; Watanabe, J.; Sonoyama, K. Role of intestinal Bifidobacterium pseudolongum in dietary fructo-oligosaccharide inhibition of 2,4-dinitrofluorobenzene-induced contact hypersensitivity in mice. Br. J. Nutr. 2010, 103, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Sawada, Y.; Saito-Sasaki, N.; Nakamura, M. Omega 3 Fatty Acid and Skin Diseases. Front. Immunol. 2021, 11. [Google Scholar] [CrossRef]
- Horrobin, D.F. Low prevalences of coronary heart disease (CHD), psoriasis, asthma and rheumatoid arthritis in Eskimos: Are they caused by high dietary intake of eicosapentaenoic acid (EPA), a genetic variation of essential fatty acid (EFA) metabolism or a combination of both? Med. Hypotheses 1987, 22, 421–428. [Google Scholar] [CrossRef]
- Sumida, H.; Yanagida, K.; Kita, Y.; Abe, J.; Matsushima, K.; Nakamura, M.; Ishii, S.; Sato, S.; Shimizu, T. Interplay between CXCR2 and BLT1 Facilitates Neutrophil Infiltration and Resultant Keratinocyte Activation in a Murine Model of Imiquimod-Induced Psoriasis. J. Immunol. 2014, 192, 4361–4369. [Google Scholar] [CrossRef]
- Rola-Pleszczynski, M.; Bouvrette, L.; Gingras, D.; Girard, M. Identification of interferon-gamma as the lymphokine that mediates leukotriene B4-induced immunoregulation. J. Immunol. 1987, 139, 513–517. [Google Scholar] [PubMed]
- Maurice, P.; Allen, B.; Barkley, A.; Cockbill, S.; Stammers, J.; Bather, P. The effects of dietary supplementation with fish oil in patients with psoriasis. Br. J. Dermatol. 1987, 117, 599–606. [Google Scholar] [CrossRef]
- Ueharaguchi, Y.; Honda, T.; Kusuba, N.; Hanakawa, S.; Adachi, A.; Sawada, Y.; Otsuka, A.; Kitoh, A.; Dainichi, T.; Egawa, G.; et al. Thromboxane A(2) facilitates IL-17A production from Vγ4(+) γδ T cells and promotes psoriatic dermatitis in mice. J. Allergy Clin. Immunol. 2018, 142, 680–683.e2. [Google Scholar] [CrossRef] [PubMed]
- James, M.J.; Gibson, R.A.; Cleland, L.G. Dietary polyunsaturated fatty acids and inflammatory mediator production. Am. J. Clin. Nutr. 2000, 71, 343s–348s. [Google Scholar] [CrossRef] [PubMed]
- Sawada, Y.; Honda, T.; Nakamizo, S.; Otsuka, A.; Ogawa, N.; Kobayashi, Y.; Nakamura, M.; Kabashima, K. Resolvin E1 attenuates murine psoriatic dermatitis. Sci. Rep. 2018, 8, 11837. [Google Scholar] [CrossRef] [PubMed]
- Saito-Sasaki, N.; Sawada, Y.; Mashima, E.; Yamaguchi, T.; Ohmori, S.; Yoshioka, H.; Haruyama, S.; Okada, E.; Nakamura, M. Maresin-1 suppresses imiquimod-induced skin inflammation by regulating IL-23 receptor expression. Sci. Rep. 2018, 8, 5522. [Google Scholar] [CrossRef]
- Xu, J.; Duan, X.; Hu, F.; Poorun, D.; Liu, X.; Wang, X.; Zhang, S.; Gan, L.; He, M.; Zhu, K.; et al. Resolvin D1 attenuates imiquimod-induced mice psoriasiform dermatitis through MAPKs and NF-κB pathways. J. Derm. Sci. 2018, 89, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Kragballe, K. Dietary supplementation with a combination of n-3 and n-6 fatty acids (super gamma-oil marine) improves psoriasis. Acta Derm. Venereol. 1989, 69, 265–268. [Google Scholar] [PubMed]
- Venter, C.; Agostoni, C.; Arshad, S.H.; Ben-Abdallah, M.; Du Toit, G.; Fleischer, D.M.; Greenhawt, M.; Glueck, D.H.; Groetch, M.; Lunjani, N.; et al. Dietary factors during pregnancy and atopic outcomes in childhood: A systematic review from the European Academy of Allergy and Clinical Immunology. Pediatr. Allergy Immunol. 2020, 31, 889–912. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-C.; Tominaga, M.; Yasukawa, K.; Ohba, M.; Takahashi, N.; Honda, K.; Okuno, T.; Takamori, K.; Yokomizo, T. Dietary supplementation of omega-3 fatty acid eicosapentaenoic acid does not ameliorate pruritus in murine models of atopic dermatitis and psoriasis. J. Dermatol. Sci. 2019, 95, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Best, K.P.; Gold, M.; Kennedy, D.; Martin, J.; Makrides, M. Omega-3 long-chain PUFA intake during pregnancy and allergic disease outcomes in the offspring: A systematic review and meta-analysis of observational studies and randomized controlled trials. Am. J. Clin. Nutr. 2016, 103, 128–143. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Yasutomo, K.; Watanabe, T. Treatment with DHA/EPA ameliorates atopic dermatitis-like skin disease by blocking LTB4 production. J. Med Investig. 2016, 63, 187–191. [Google Scholar] [CrossRef]
- Kim, T.-H.; Kim, G.-D.; Jin, Y.-H.; Park, Y.S.; Park, C.-S. Omega-3 fatty acid-derived mediator, Resolvin E1, ameliorates 2,4-dinitrofluorobenzene-induced atopic dermatitis in NC/Nga mice. Int. Immunopharmacol. 2012, 14, 384–391. [Google Scholar] [CrossRef]
- Gardner, K.G.; Gebretsadik, T.; Hartman, T.J.; Rosa, M.J.; Tylavsky, F.A.; Adgent, M.A.; Moore, P.E.; Kocak, M.; Bush, N.R.; Davis, R.L.; et al. Prenatal Omega-3 and Omega-6 Polyunsaturated Fatty Acids and Childhood Atopic Dermatitis. J. Allergy Clin. Immunol. Pr. 2020, 8, 937–944. [Google Scholar] [CrossRef]
- Fogh, K.; Herlin, T.; Kragballe, K. Eicosanoids in skin of patients with atopic dermatitis: Prostaglandin E2 and leukotriene B4 are present in biologically active concentrations. J. Allergy Clin. Immunol. 1989, 83, 450–455. [Google Scholar] [CrossRef]
- Oyoshi, M.K.; He, R.; Li, Y.; Mondal, S.; Yoon, J.; Afshar, R.; Chen, M.; Lee, D.M.; Luo, H.R.; Luster, A.D.; et al. Leukotriene B4-driven neutrophil recruitment to the skin is essential for allergic skin inflammation. Immunity 2012, 37, 747–758. [Google Scholar] [CrossRef]
- Sawada, Y.; Honda, T.; Nakamizo, S.; Nakajima, S.; Nonomura, Y.; Otsuka, A.; Egawa, G.; Yoshimoto, T.; Nakamura, M.; Narumiya, S.; et al. Prostaglandin E2 (PGE2)–EP2 signaling negatively regulates murine atopic dermatitis–like skin inflammation by suppressing thymic stromal lymphopoietin expression. J. Allergy Clin. Immunol. 2019, 144, 1265–1273.e9. [Google Scholar] [CrossRef] [PubMed]
- Laouini, D.; Elkhal, A.; Yalcindag, A.; Kawamoto, S.; Oettgen, H.; Geha, R.S. COX-2 inhibition enhances the TH2 immune response to epicutaneous sensitization. J. Allergy Clin. Immunol. 2005, 116, 390–396. [Google Scholar] [CrossRef] [PubMed]
- Tomobe, Y.I.; Morizawa, K.; Tsuchida, M.; Hibino, H.; Nakano, Y.; Tanaka, Y. Dietary docosahexaenoic acid suppresses inflammation and immunoresponses in contact hypersensitivity reaction in mice. Lipids 2000, 35, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Sierra, S.; Lara-Villoslada, F.; Comalada, M.; Olivares, M.; Xaus, J. Dietary eicosapentaenoic acid and docosahexaenoic acid equally incorporate as decosahexaenoic acid but differ in inflammatory effects. Nutrition 2008, 24, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Arita, M.; Bianchini, F.; Aliberti, J.; Sher, A.; Chiang, N.; Hong, S.; Yang, R.; Petasis, N.A.; Serhan, C.N. Stereochemical assignment, antiinflammatory properties, and receptor for the omega-3 lipid mediator resolvin E1. J. Exp. Med. 2005, 201, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Honda, T.; Matsuoka, T.; Ueta, M.; Kabashima, K.; Miyachi, Y.; Narumiya, S. Prostaglandin E2–EP3 signaling suppresses skin inflammation in murine contact hypersensitivity. J. Allergy Clin. Immunol. 2009, 124, 809–818.e2. [Google Scholar] [CrossRef]
- Kabashima, K.; Sakata, D.; Nagamachi, M.; Miyachi, Y.; Inaba, K.; Narumiya, S. Prostaglandin E2–EP4 signaling initiates skin immune responses by promoting migration and maturation of Langerhans cells. Nat. Med. 2003, 9, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Robb, C.T.; McSorley, H.J.; Lee, J.; Aoki, T.; Yu, C.; Crittenden, S.; Astier, A.; Felton, J.M.; Parkinson, N.; Ayele, A.; et al. Prostaglandin E2 stimulates adaptive IL-22 production and promotes allergic contact dermatitis. J. Allergy Clin. Immunol. 2018, 141, 152–162. [Google Scholar] [CrossRef]
- Price, L.R.; Martinez, J. Cardiovascular, carcinogenic and reproductive effects of nicotine exposure: A narrative review of the scientific literature. F1000Research 2020, 8, 1586. [Google Scholar] [CrossRef] [PubMed]
- Kavli, G.; Forde, O.H.; Arnesen, E.; Stenvold, S.E. Psoriasis: Familial predisposition and environmental factors. BMJ 1985, 291, 999–1000. [Google Scholar] [CrossRef] [PubMed]
- Setty, A.R.; Curhan, G.; Choi, H.K. Smoking and the Risk of Psoriasis in Women: Nurses’ Health Study II. Am. J. Med. 2007, 120, 953–959. [Google Scholar] [CrossRef]
- Lee, E.J.; Han, K.D.; Han, J.H.; Lee, J.H. Smoking and risk of psoriasis: A nationwide cohort study. J. Am. Acad. Dermatol. 2017, 77, 573–575. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.W.; Armstrong, E.J.; Fuller, E.N.; Sockolov, M.E.; Voyles, S.V. Smoking and pathogenesis of psoriasis: A review of oxidative, inflammatory and genetic mechanisms. Br. J. Dermatol. 2011, 165, 1162–1168. [Google Scholar] [CrossRef]
- Shan, M.; Yuan, X.; Song, L.Z.; Roberts, L.; Zarinkamar, N.; Seryshev, A.; Zhang, Y.; Hilsenbeck, S.; Chang, S.H.; Dong, C.; et al. Cigarette smoke induction of osteopontin (SPP1) mediates T(H)17 inflammation in human and experimental emphysema. Sci. Transl. Med. 2012, 4, 117ra9. [Google Scholar] [CrossRef] [PubMed]
- Stelmach, I.; Bobrowska-Korzeniowska, M.; Smejda, K.; Majak, P.; Jerzynska, J.; Stelmach, W.; Polańska, K.; Sobala, W.; Krysicka, J.; Hanke, W. Risk factors for the development of atopic dermatitis and early wheeze. Allergy Asthma Proc. 2014, 35, 382–389. [Google Scholar] [CrossRef]
- Ahn, K. The role of air pollutants in atopic dermatitis. J. Allergy Clin. Immunol. 2014, 134, 993–999. [Google Scholar] [CrossRef]
- Egeberg, A.; Andersen, Y.M.; Gislason, G.H.; Skov, L.; Thyssen, J.P. Prevalence of comorbidity and associated risk factors in adults with atopic dermatitis. Allergy 2016, 72, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Lee, S.Y.; Lee, K.-S. The Use of Heated Tobacco Products is Associated with Asthma, Allergic Rhinitis, and Atopic Dermatitis in Korean Adolescents. Sci. Rep. 2019, 9, 17699. [Google Scholar] [CrossRef]
- Weidinger, S.; Beck, L.A.; Bieber, T.; Kabashima, K.; Irvine, A.D. Atopic dermatitis. Nat. Rev. Dis. Primers 2018, 4, 1. [Google Scholar] [CrossRef]
- Jachiet, M.; Nosbaum, A.; Staumont-Sallé, D.; Seneschal, J.; Viguier, M.; Soria, A.; Barbaud, A.; Carriou, A.; Chuffart-Delplanque, M.; Darrigade, A.; et al. Low cardiovascular risk and poor quality of life associated with tobacco use and skin infections in adult atopic dermatitis: Result of a French multicenter study. J. Eur. Acad. Dermatol. Venereol. 2019, 33, e451–e453. [Google Scholar] [CrossRef]
- Huang, C.-C.; Chiang, T.-L.; Chen, P.-C.; Lin, S.-J.; Wen, H.-J.; Guo, Y.L. Risk factors for asthma occurrence in children with early-onset atopic dermatitis: An 8-year follow-up study. Pediatr. Allergy Immunol. 2017, 29, 159–165. [Google Scholar] [CrossRef]
- Tanaka, K.; Miyake, Y.; Furukawa, S.; Arakawa, M. Pre- and Postnatal Smoking Exposure and Risk of Atopic Eczema in Young Japanese Children: A Prospective Prebirth Cohort Study. Nicotine Tob. Res. 2016, 19, 804–809. [Google Scholar] [CrossRef] [PubMed]
- Feleszko, W.; Zawadzka-Krajewska, A.; Matysiak, K.; Lewandowska, D.; Peradzyńska, J.; Dinh, Q.T.; Hamelmann, E.; Groneberg, D.A.; Kulus, M. Parental tobacco smoking is associated with augmented IL-13 secretion in children with allergic asthma. J. Allergy Clin. Immunol. 2006, 117, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Jiang, Y.; Xia, P.; Luo, G.; Huang, W.; Hu, Z.; Cheng, G.; Xiong, Y.; Wang, Y.; Cui, T. Cigarette Smoke Extract Promotes TIM4 Expression in Murine Dendritic Cells Leading to Th2 Polarization through ERK-Dependent Pathways. Int. Arch. Allergy Immunol. 2018, 178, 219–228. [Google Scholar] [CrossRef]
- Chen, Y.X.; Cheng, H.Y.; Li, L.F. Prevalence and risk factors of contact dermatitis among clothing manufacturing employees in Beijing: A cross-sectional study. Medicine 2017, 96, e6356. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, J.A.; Fisker, M.H.; Agner, T.; Clemmensen, K.K.B.; Ebbehøj, N.E. Associations between lifestyle factors and hand eczema severity: Are tobacco smoking, obesity and stress significantly linked to eczema severity? Contact Dermat. 2016, 76, 138–145. [Google Scholar] [CrossRef]
- Van Der Heiden, J.; Agner, T.; Rustemeyer, T.; Clemmensen, K.K.B. Hyperkeratotic hand eczema compared to other subgroups of hand eczema-a retrospective study with a follow-up questionnaire. Contact Dermat. 2018, 78, 216–222. [Google Scholar] [CrossRef]
- Zimmer, K.A.; Armbrecht, E.S.; Burkemper, N.M. The association of smoking with contact dermatitis and hand eczema-a review. Int. J. Dermatol. 2017, 57, 375–387. [Google Scholar] [CrossRef]
- Sahu, R.P.; Petrache, I.; Van Demark, M.J.; Rashid, B.M.; Ocana, J.A.; Tang, Y.; Yi, Q.; Turner, M.J.; Konger, R.L.; Travers, J.B. Cigarette Smoke Exposure Inhibits Contact Hypersensitivity via the Generation of Platelet-Activating Factor Agonists. J. Immunol. 2013, 190, 2447–2454. [Google Scholar] [CrossRef] [PubMed]
- Shaler, C.R.; Horvath, C.N.; McCormick, S.; Jeyanathan, M.; Khera, A.; Zganiacz, A.; Kasińska, J.; Stampfli, M.R.; Xing, Z. Continuous and Discontinuous Cigarette Smoke Exposure Differentially Affects Protective Th1 Immunity against Pulmonary Tuberculosis. PLoS ONE 2013, 8, e59185. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, X.; Chen, X.; Lin, Y.; Qiu, S.; Zhao, Y.; Tang, Q.; Liang, Y.; Zhong, X. Rapamycin attenuates Tc1 and Tc17 cell responses in cigarette smoke-induced emphysema in mice. Inflamm. Res. 2019, 68, 957–968. [Google Scholar] [CrossRef]
- Heidari-Japelaghi, R.; Valizadeh, M.; Haddad, R.; Dorani-Uliaie, E.; Jalali-Javaran, M. Fusion to elastin-like polypeptide increases production of bioactive human IFN-γ in tobacco. Transgenic Res. 2020, 29, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Alkhattabi, N.; Todd, I.; Negm, O.; Tighe, P.J.; Fairclough, L.C. Tobacco smoke and nicotine suppress expression of activating signaling molecules in human dendritic cells. Toxicol. Lett. 2018, 299, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Naldi, L.; Peli, L.; Parazzini, F. Association of early-stage psoriasis with smoking and male alcohol consumption: Evidence from an Italian case-control study. Arch. Derm. 1999, 135, 1479–1484. [Google Scholar] [CrossRef]
- Farkas, A.; Kemény, L.; Széll, M.; Dobozy, A.; Bata-Csörgo, Z. Ethanol and acetone stimulate the proliferation of HaCaT keratinocytes: The possible role of alcohol in exacerbating psoriasis. Arch. Derm. Res. 2003, 295, 56–62. [Google Scholar] [CrossRef]
- Ockenfels, H.M.; Keim-Maas, C.; Funk, R.; Nussbaum, G.; Goos, M. Ethanol enhances the IFN-gamma, TGF-alpha and IL-6 secretion in psoriatic co-cultures. Br. J. Dermatol. 1996, 135, 746–751. [Google Scholar] [CrossRef]
- Schopf, R.E.; Ockenfels, H.M.; Morsches, B. Ethanol enhances the mitogen-driven lymphocyte proliferation in patients with psoriasis. Acta Derm. Venereol. 1996, 76, 260–263. [Google Scholar]
- Halling-Overgaard, A.-S.; Hamann, C.R.; Holm, R.P.; Linneberg, A.; Silverberg, J.I.; Egeberg, A.; Thyssen, J.P. Atopic dermatitis and alcohol use-a meta-analysis and systematic review. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 1238–1245. [Google Scholar] [CrossRef]
- Nakashima, C.; Ishida, Y.; Kitoh, A.; Otsuka, A.; Kabashima, K. Interaction of peripheral nerves and mast cells, eosinophils, and basophils in the development of pruritus. Exp. Dermatol. 2019, 28, 1405–1411. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.-B.; Xu, L.; Wang, Y.; Zhang, R.; Wang, Y.-C.; Li, J.-B.; Mu, D. Posterior Thalamic Nucleus Mediates Facial Histaminergic Itch. Neuroscience 2020, 444, 54–63. [Google Scholar] [CrossRef]
- Kabashima, K.; Matsumura, T.; Komazaki, H.; Kawashima, M. Trial of Nemolizumab and Topical Agents for Atopic Dermatitis with Pruritus. N. Engl. J. Med. 2020, 383, 141–150. [Google Scholar] [CrossRef]
- Gonzales, R.A.; Crews, F.T. Effect of ethanol and aging on histamine release and membranes of mast cells. Alcohol 1985, 2, 313–316. [Google Scholar] [CrossRef]
- Wyczółkowska, J.; Korsak, Z.; Maśliński, C. The effect of ethanol administration into the mouse on mast cell histamine releasability in vitro. Pol. J. Pharmacol. Pharm. 1984, 36, 641–646. [Google Scholar] [PubMed]
- Kawano, T.; Matsuse, H.; Kondo, Y.; Machida, I.; Saeki, S.; Tomari, S.; Mitsuta, K.; Obase, Y.; Fukushima, C.; Shimoda, T.; et al. Acetaldehyde Induces Histamine Release from Human Airway Mast Cells to Cause Bronchoconstriction. Int. Arch. Allergy Immunol. 2004, 134, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Doggett, T.M.; Breslin, J.W. Acute Alcohol Intoxication-Induced Microvascular Leakage. Alcohol. Clin. Exp. Res. 2014, 38, 2414–2426. [Google Scholar] [CrossRef]
- Elamin, E.; Jonkers, D.; Juuti-Uusitalo, K.; Van Ijzendoorn, S.; Troost, F.; Duimel, H.; Broers, J.; Verheyen, F.; Dekker, J.; Masclee, A. Effects of Ethanol and Acetaldehyde on Tight Junction Integrity: In Vitro Study in a Three Dimensional Intestinal Epithelial Cell Culture Model. PLoS ONE 2012, 7, e35008. [Google Scholar] [CrossRef]
- Li, W.-Q.; Qureshi, A.A.; Schernhammer, E.S.; Han, J. Rotating night-shift work and risk of psoriasis in US women. J. Investig. Dermatol. 2013, 133, 565–567. [Google Scholar] [CrossRef]
- Ando, N.; Nakamura, Y.; Aoki, R.; Ishimaru, K.; Ogawa, H.; Okumura, K.; Shibata, S.; Shimada, S.; Nakao, A. Circadian Gene Clock Regulates Psoriasis-Like Skin Inflammation in Mice. J. Investig. Dermatol. 2015, 135, 3001–3008. [Google Scholar] [CrossRef]
- Lu, F.; Suggs, A.; Ezaldein, H.H.; Ya, J.; Fu, P.; Jamora, J.; Verallo-Rowel, V.; Baron, E.D. The Effect of Shift Work and Poor Sleep on Self-Reported Skin Conditions: A Survey of Call Center Agents in the Philippines. Clocks Sleep 2019, 1, 23. [Google Scholar] [CrossRef]
- Chang, Y.-S.; Chou, Y.-T.; Lee, J.-H.; Lee, P.-L.; Dai, Y.-S.; Sun, C.; Lin, Y.-T.; Wang, L.-C.; Yu, H.-H.; Yang, Y.-H.; et al. Atopic Dermatitis, Melatonin, and Sleep Disturbance. Pediatrics 2014, 134, e397–e405. [Google Scholar] [CrossRef]
- Matsunaga, N.; Itcho, K.; Hamamura, K.; Ikeda, E.; Ikeyama, H.; Furuichi, Y.; Watanabe, M.; Koyanagi, S.; Ohdo, S. 24-Hour Rhythm of Aquaporin-3 Function in the Epidermis Is Regulated by Molecular Clocks. J. Investig. Dermatol. 2014, 134, 1636–1644. [Google Scholar] [CrossRef]
- Takita, E.; Yokota, S.; Tahara, Y.; Hirao, A.; Aoki, N.; Nakamura, Y.; Nakao, A.; Shibata, S. Biological clock dysfunction exacerbates contact hypersensitivity in mice. Br. J. Dermatol. 2012, 168, 39–46. [Google Scholar] [CrossRef]
- Savetsky, I.L.; Albano, N.J.; Cuzzone, D.A.; Gardenier, J.C.; Torrisi, J.S.; Nores, G.D.G.; Nitti, M.D.; Hespe, G.E.; Nelson, T.S.; Kataru, R.P.; et al. Lymphatic Function Regulates Contact Hypersensitivity Dermatitis in Obesity. J. Investig. Dermatol. 2015, 135, 2742–2752. [Google Scholar] [CrossRef]
- Dupuy, A.; Benchikhi, H.; Roujeau, J.-C.; Bernard, P.; Vaillant, L.; Chosidow, O.; Sassolas, B.; Guillaume, J.-C.; Grob, J.-J.; Bastuji-Garin, S. Risk factors for erysipelas of the leg (cellulitis): Case-control study. BMJ 1999, 318, 1591–1594. [Google Scholar] [CrossRef]
- Budu-Aggrey, A.; Brumpton, B.; Tyrrell, J.; Watkins, S.; Modalsli, E.H.; Celis-Morales, C.; Ferguson, L.D.; Vie, G.Å.; Palmer, T.; Fritsche, L.G.; et al. Evidence of a causal relationship between body mass index and psoriasis: A mendelian randomization study. PLoS Med. 2019, 16, e1002739. [Google Scholar] [CrossRef] [PubMed]
- Vasseur, P.; Serres, L.; Jégou, J.-F.; Pohin, M.; Delwail, A.; Petit-Paris, I.; Levillain, P.; Favot, L.; Samson, M.; Yssel, H.; et al. High-Fat Diet–Induced IL-17A Exacerbates Psoriasiform Dermatitis in a Mouse Model of Steatohepatitis. Am. J. Pathol. 2016, 186, 2292–2301. [Google Scholar] [CrossRef] [PubMed]
- Nakamizo, S.; Honda, T.; Adachi, A.; Nagatake, T.; Kunisawa, J.; Kitoh, A.; Otsuka, A.; Dainichi, T.; Nomura, T.; Ginhoux, F.; et al. High fat diet exacerbates murine psoriatic dermatitis by increasing the number of IL-17-producing γδ T cells. Sci. Rep. 2017, 7, 14076. [Google Scholar] [CrossRef]
- Zhang, A.; Silverberg, J.I. Association of atopic dermatitis with being overweight and obese: A systematic review and metaanalysis. J. Am. Acad. Dermatol. 2015, 72, 606–616.e4. [Google Scholar] [CrossRef]
- Budu-Aggrey, A.; Watkins, S.H.; Brumpton, B.; Løset, M.; Tyrrell, J.; Modalsli, E.H.; Vie, G.Å.; Palmer, T.; Fritsche, L.G.; Nielsen, J.B.; et al. Assessment of a causal relationship between body mass index and atopic dermatitis. J. Allergy Clin. Immunol. 2021, 147, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Yosipovitch, G.; DeVore, A.; Dawn, A. Obesity and the skin: Skin physiology and skin manifestations of obesity. J. Am. Acad. Dermatol. 2007, 56, 901–916. [Google Scholar] [CrossRef] [PubMed]
- Sybilski, A.J.; Raciborski, F.; Lipiec, A.; Tomaszewska, A.; Lusawa, A.; Furmańczyk, K.; Krzych-Fałta, E.; Komorowski, J.; Samoliński, B. Obesity—A risk factor for asthma, but not for atopic dermatitis, allergic rhinitis and sensitization. Public Heal. Nutr. 2014, 18, 530–536. [Google Scholar] [CrossRef]
- Savetsky, I.L.; Torrisi, J.S.; Cuzzone, D.A.; Ghanta, S.; Albano, N.J.; Gardenier, J.C.; Joseph, W.J.; Mehrara, B.J. Obesity increases inflammation and impairs lymphatic function in a mouse model of lymphedema. Am. J. Physiol. Circ. Physiol. 2014, 307, H165–H172. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.Y.; Rutkowski, J.M.; Helft, J.; Reddy, S.T.; Swartz, M.A.; Randolph, G.J.; Angeli, V. Hypercholesterolemic Mice Exhibit Lymphatic Vessel Dysfunction and Degeneration. Am. J. Pathol. 2009, 175, 1328–1337. [Google Scholar] [CrossRef]
- Decock, P.A.; Shanon, J. Pseudo-atopic dermatitis. An example of pseudo-nomenclature. Dermatologica 1966, 133, 236–237. [Google Scholar]
- Sharma, A. Relationship between nickel allergy and diet. Indian J. Dermatol. Venereol. Leprol. 2007, 73, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Plonka, P.M.; Szaflarski, J.P.; Paus, R. How UV Light Touches the Brain and Endocrine System Through Skin, and Why. Endocrinology 2018, 159, 1992–2007. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Skobowiat, C.; Zbytek, B.; Slominski, R.M.; Steketee, J.D. Sensing the environment: Regulation of local and global homeostasis by the skin’s neuroendocrine system. Adv. Anat. Embryol. Cell Biol. 2012, 212, 1–115. [Google Scholar]


{kind=link}
| Psoriasis | AD | Contact Dermatitis | |
|---|---|---|---|
| Gluten or wheat | Gluten free diet CD4+ cell infiltration ↓ [34] IFN-γ, IL-12, IL-17 ↓ [35] | Gluten: TSLP ↑ [40] Salt soluble wheat protein: TNF-α, IFN-γ, IL-1β, IL-4, IL-10, IL-13, IL-17, CCL1, CCL5, CCL11, CCL12, CCL22, CCL24, CXCL9, CXCL16, E-selectin ↑ [39] | |
| Fiber | Dietary fiber: Treg ↓ [45] Butyrate:IL-17, IL-6↓IL-10 ↑ [46] | Butyrate: Treg ↑ IL-6 ↓ [49], IL-10 ↑ [49,50]. | |
| Lipid | LTB4: Neutrophil migration, IL-19 ↑ [53], DC migration↑ [31], IFN-γ ↑ [54] TxA2: IL-17 ↑ [56]. EPA: TNF-α, IL-1β ↓ [57]. RvE1: IL-17, IL-23, IL-12b ↓ [58] MaR1: IL-17 ↓ [59] RvD1: IL-23, IL-22, IL-17, TNF-α ↓ [60]. | EPA/DHA: IL-13, IL-17, LTB4, T cell migration ↓ [65] RvE1: IFN-γ, IL-4, IgE ↓ [66] LTB4: IL-4, IL-13, CD4 migration↑ [69] PGE2-EP2: TSLP ↓ [70] | DHA: IFN-γ, IL-6, IL-1β, IL-2↓ [72] EPA/DHA: TNF-α, IL-2, IL-4, IL-12 ↓ IL-10 ↑ [73] RvE1: DC migration ↓ [31,74], IFN-γ ↓ [31] PGE2-EP3: CXCL1 ↓ [75] PGE2-EP4: LC maturation, migration ↑ [76]. PGE2-EP2/EP4: IL-22 ↑ [77] |
| Smoking | IFN-γ, IL-17A ↑ [83] | IL-13 ↑ [92],, IL-4 ↑ [93] | IFN-γ ↑ [99,100,101] Nicotine: DC maturation, T cell activation/migration ↓ [102] |
| Alcohol | Ethanol: IFN-γ, TNF-α, IL-6 ↑ [105], Lymphocyte proliferation ↑ [106] | Ethanol: Histamine ↓ [111] Acetaldehyde: Histamine ↑ [113] | |
| Sleep disturbance | IL-17 ↑ [117] | Skin hydration ↓ [120] | Mast cell, IgE↑ [121] |
| Obesity | Th17 ↑ [125,126] | Skin burrier ↓ [129], TSLP ↑ [70] | Lymphatic vessel disfunction [131,132], IFN-γ ↑ [122] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sawada, Y.; Saito-Sasaki, N.; Mashima, E.; Nakamura, M. Daily Lifestyle and Inflammatory Skin Diseases. Int. J. Mol. Sci. 2021, 22, 5204. https://doi.org/10.3390/ijms22105204
Sawada Y, Saito-Sasaki N, Mashima E, Nakamura M. Daily Lifestyle and Inflammatory Skin Diseases. International Journal of Molecular Sciences. 2021; 22(10):5204. https://doi.org/10.3390/ijms22105204
Chicago/Turabian StyleSawada, Yu, Natsuko Saito-Sasaki, Emi Mashima, and Motonobu Nakamura. 2021. "Daily Lifestyle and Inflammatory Skin Diseases" International Journal of Molecular Sciences 22, no. 10: 5204. https://doi.org/10.3390/ijms22105204
APA StyleSawada, Y., Saito-Sasaki, N., Mashima, E., & Nakamura, M. (2021). Daily Lifestyle and Inflammatory Skin Diseases. International Journal of Molecular Sciences, 22(10), 5204. https://doi.org/10.3390/ijms22105204