Role of Ca2+ in Mediating Plant Responses to Extracellular ATP and ADP



{kind=link}
Abstract
1. Introduction
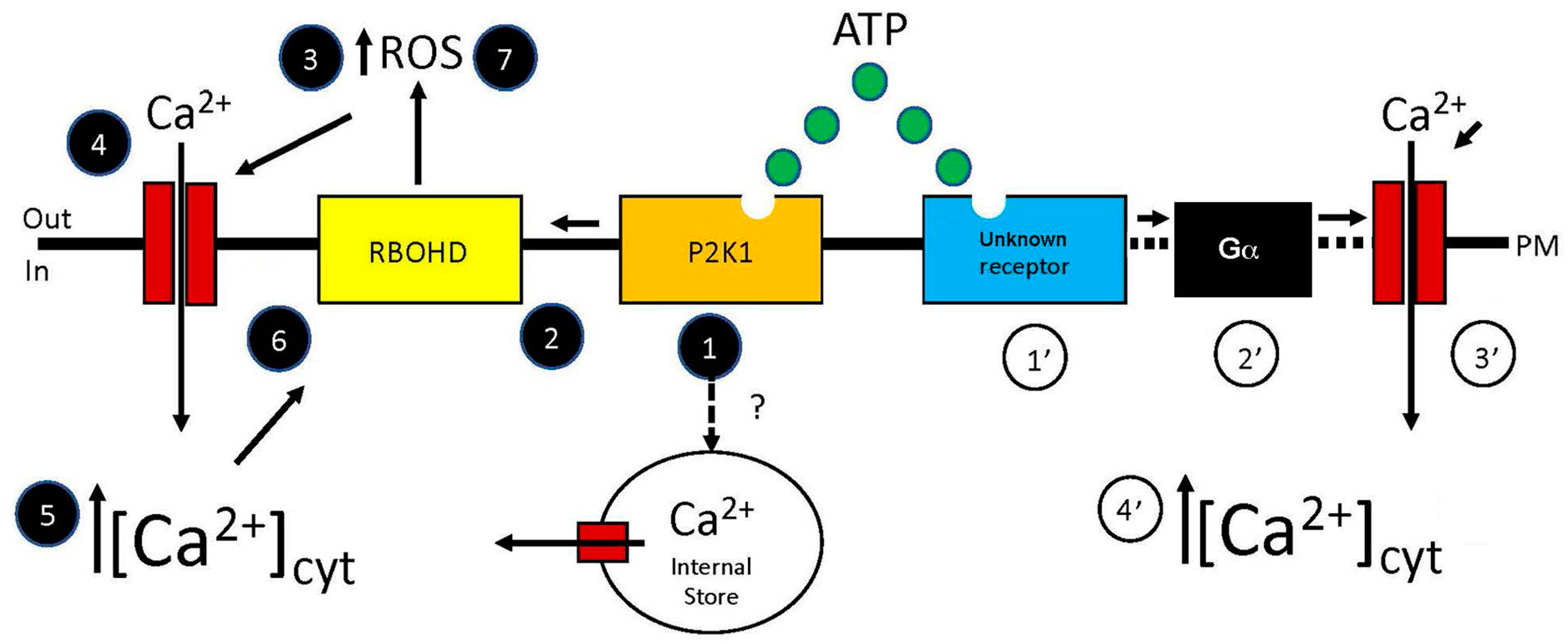
2. Rapid Changes in [Ca2+]cyt Induced by eATP
3. Role of eATP-Induced Increase in [Ca2+]cyt in Triggering Downstream Signaling Changes
4. Stress-Induced Growth Responses Regulated by eATP through Ca2+
5. Role of Ca2+/CaM-Activated Apyrase in Mediating Growth and Development Changes in Plants
6. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Clark, G.B.; Morgan, R.O.; Fernandez, M.P.; Salmi, M.L.; Roux, S.J. Breakthroughs spotlighting roles for extracellular nucleotides and apyrases in stress responses and growth and development. Plant Sci. 2014, 225, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Burnstock, G. Biology of purinergic signalling: Its ancient evolutionary roots, its omnipresence and its multiple functional significance. Bioessays 2014, 36, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Tanaka, K.; Cao, Y.R.; Qi, Y.; Qiu, J.; Liang, Y.; Lee, S.Y.; Stacey, G. Identification of a Plant Receptor for Extracellular ATP. Science 2014, 343, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G.; Verkhratsky, A. Evolutionary origins of the purinergic signalling system. Acta Physiol. 2009, 195, 415–447. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V.; Shang, Z.; Shin, R.; Thompson, E.; Rubio, L.; Laohavisit, A.; Mortimer, J.C.; Chivasa, S.; Slabas, A.R.; Glover, B.J.; et al. Plant extracellular ATP signalling by plasma membrane NADPH oxidase and Ca2+ channels. Plant J. 2009, 58, 903–913. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V.; Shang, Z.; Shin, R.; Colaco, R.; Laohavisit, A.; Shabala, S.; Davies, J.M. Receptor-Like Activity Evoked by Extracellular ADP in Arabidopsis Root Epidermal Plasma Membrane. Plant Physiol. 2011, 156, 1375–1385. [Google Scholar] [CrossRef] [PubMed]
- Jeter, C.R.; Tang, W.Q.; Henaff, E.; Butterfield, T.; Roux, S.J. Evidence of a novel cell signaling role for extracellular adenosine triphosphates and diphosphates in Arabidopsis. Plant Cell 2004, 16, 2652–2664. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.R.; Datta, N.; Roux, S.J. Purification and partial characterization of a calmodulin-stimulated nucleoside triphosphatase from pea nuclei. J. Biol. Chem. 1987, 262, 10689–10694. [Google Scholar] [PubMed]
- Steinebrunner, I.; Jeter, C.; Song, C.; Roux, S.J. Molecular and biochemical comparison of two different apyrases from Arabidopsis thaliana. Plant Physiol. Biochem. 2000, 38, 913–922. [Google Scholar] [CrossRef]
- Khakh, B.S.; Burnstock, G. The Double Life of ATP. Sci. Am. 2009, 301, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V.; Nichols, C.; Oliynyk, M.; Dark, A.; Glover, B.J.; Davies, J.M. Is ATP a signaling agent in plants? Plant Physiol. 2003, 133, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Tanaka, K.; Liang, Y.; Cao, Y.R.; Lee, S.Y.; Stacey, G. Extracellular ATP, a danger signal, is recognized by DORN1 in Arabidopsis. Biochem. J. 2014, 463, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.; Zhang, T.; Koo, A.J.; Stacey, G.; Tanaka, K. Extracellular ATP Acts on Jasmonate Signaling to Reinforce Plant Defense. Plant Physiol. 2018, 176, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.Q.; Cao, Y.R.; Li, H.; Kim, D.; Ahsan, N.; Thelen, J.; Stacey, G. Extracellular ATP elicits DORN1-mediated RBOHD phosphorylation to regulate stomatal aperture. Nat. Commun. 2017, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.M.; Wilkins, K.A.; Davies, J.M. Arabidopsis DORN1 extracellular ATP receptor; activation of plasma membrane K+-and Ca2+-permeable conductances. New Phytol. 2018, 218, 1301–1304. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.H.; Zhu, S.; Ye, R.; Xue, Y.; Chen, A.; An, L.Z.; Pei, Z.M. Relationship between NaCl- and H2O2-Induced Cytosolic Ca2+ Increases in Response to Stress in Arabidopsis. PLoS ONE 2013, 8, e76130. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.J.; Dong, X.X.; Hao, W.W.; Gao, W.; Zhang, W.Z.; Xia, S.Y.; Liu, T.; Shang, Z.L. Heterotrimeric G Protein-Regulated Ca2+ Influx and PIN2 Asymmetric Distribution Are Involved in Arabidopsis thaliana Roots’ Avoidance Response to Extracellular ATP. Front. Plant Sci. 2017, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Weerasinghe, R.R.; Swanson, S.J.; Okada, S.F.; Garrett, M.B.; Kim, S.-Y.; Stacey, G.; Boucher, R.C.; Gilroy, S.; Jones, A.M. Touch induces ATP release in Arabidopsis roots that is modulated by the heterotrimeric G-protein complex. FEBS Lett. 2009, 583, 2521–2526. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Swanson, S.J.; Gilroy, S.; Stacey, G. Extracellular Nucleotides Elicit Cytosolic Free Calcium Oscillations in Arabidopsis. Plant Physiol. 2010, 154, 705–719. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.-H.; Wang, W.-X.; Chen, C.; Wang, Y.-F.; Liu, T.; Li, X.; Shang, Z.-L. Extracellular ATP Promotes Stomatal Opening of Arabidopsis thaliana through Heterotrimeric G Protein a Subunit and Reactive Oxygen Species. Mol. Plant 2012, 5, 852–864. [Google Scholar] [CrossRef] [PubMed]
- Fountain, S.J.; Cao, L.S.; Young, M.T.; North, R.A. Permeation properties of a P2X receptor in the green algae Ostreococcus tauri. J. Biol. Chem. 2008, 283, 15122–15126. [Google Scholar] [CrossRef] [PubMed]
- Lew, R.R.; Dearnaley, J.D.W. Extracellular nucleotide effects on the electrical properties of growing Arabidopsis thaliana root hairs. Plant Sci. 2000, 153, 1–6. [Google Scholar] [CrossRef]
- Clark, G.; Roux, S.J. Apyrases, extracellular ATP and the regulation of growth. Curr. Opin. Plant Biol. 2011, 14, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Song, C.J.; Steinebrunner, I.; Wang, X.Z.; Stout, S.C.; Roux, S.J. Extracellular ATP induces the accumulation of superoxide via NADPH oxidases in Arabidopsis. Plant Physiol. 2006, 140, 1222–1232. [Google Scholar] [CrossRef] [PubMed]
- Seybold, H.; Trempel, F.; Ranf, S.; Scheel, D.; Romeis, T.; Lee, J. Ca2+ signalling in plant immune response: From pattern recognition receptors to Ca2+ decoding mechanisms. New Phytol. 2014, 204, 782–790. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Blumwald, E. The Roles of ROS and ABA in Systemic Acquired Acclimation. Plant Cell 2015, 27, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.J.; Liao, W.B. Hydrogen Peroxide Signaling in Plant Development and Abiotic Responses: Crosstalk with Nitric Oxide and Calcium. Front. Plant Sci. 2016, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Gilroy, S.; Bialasek, M.; Suzuki, N.; Gorecka, M.; Devireddy, A.R.; Karpinski, S.; Mittler, R. ROS, Calcium, and Electric Signals: Key Mediators of Rapid Systemic Signaling in Plants. Plant Physiol. 2016, 171, 1606–1615. [Google Scholar] [CrossRef] [PubMed]
- Peiter, E. The Ever-Closer Union of Signals: Propagating Waves of Calcium and ROS Are Inextricably Linked. Plant Physiol. 2016, 172, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V. ROS-Activated Ion Channels in Plants: Biophysical Characteristics, Physiological Functions and Molecular Nature. Int. J. Mol. Sci. 2018, 19, 1263. [Google Scholar] [CrossRef] [PubMed]
- Shang, Z.; Laohavisit, A.; Davies, J.M. Extracellular ATP activates an Arabidopsis plasma membrane Ca2+-permeable conductance. Plant Signal. Behav. 2009, 4, 989–991. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.L.; Laohavisit, A.; Mortimer, J.C.; Shabala, L.; Swarbreck, S.M.; Shabala, S.; Davies, J.M. Annexin 1 regulates the H2O2-induced calcium signature in Arabidopsis thaliana roots. Plant J. 2014, 77, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Ohura, I.; Kawakita, K.; Yokota, N.; Fujiwara, M.; Shimamoto, K.; Doke, N.; Yoshioka, H. Calcium-dependent protein kinases regulate the production of reactive oxygen species by potato NADPH oxidase. Plant Cell 2007, 19, 1065–1080. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.B.; Chen, J.P.; Wang, Q.; Yang, Y.O. Direct Phosphorylation and Activation of a Mitogen-Activated Protein Kinase by a Calcium-Dependent Protein Kinase in Rice. Plant Cell 2014, 26, 3077–3089. [Google Scholar] [CrossRef] [PubMed]
- Roux, S.J.; Song, C.; Jeter, C. Regulation of plant growth and development by extracellular nucleotides. In Communication in Plants; Springer: Berlin/Heidelberg, Germany, 2006; pp. 221–234. [Google Scholar]
- Reichler, S.A.; Torres, J.; Rivera, A.L.; Cintolesi, V.A.; Clark, G.; Roux, S.J. Intersection of two signalling pathways: Extracellular nucleotides regulate pollen germination and pollen tube growth via nitric oxide. J. Exp. Bot. 2009, 60, 2129–2138. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.; Wu, M.; Wat, N.; Onyirimba, J.; Pham, T.; Herz, N.; Ogoti, J.; Gomez, D.; Canales, A.A.; Aranda, G.; et al. Both the stimulation and inhibition of root hair growth induced by extracellular nucleotides in Arabidopsis are mediated by nitric oxide and reactive oxygen species. Plant Mol. Biol. 2010, 74, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.; Torres, J.; Finlayson, S.; Guan, X.Y.; Handley, C.; Lee, J.; Kays, J.E.; Chen, Z.J.; Roux, S.J. Apyrase (Nucleoside Triphosphate-Diphosphohydrolase) and Extracellular Nucleotides Regulate Cotton Fiber Elongation in Cultured Ovules. Plant Physiol. 2010, 152, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.; Fraley, D.; Steinebrunner, I.; Cervantes, A.; Onyirimba, J.; Liu, A.; Torres, J.; Tang, W.Q.; Kim, J.; Roux, S.J. Extracellular Nucleotides and Apyrases Regulate Stomatal Aperture in Arabidopsis. Plant Physiol. 2011, 156, 1740–1753. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.R.; Sun, J.; Zhao, R.; Ding, M.Q.; Zhang, Y.N.; Sun, Y.L.; Wang, W.; Tan, Y.Q.; Liu, D.D.; Ma, X.J.; et al. Populus euphratica APYRASE2 Enhances Cold Tolerance by Modulating Vesicular Trafficking and Extracellular ATP in Arabidopsis Plants. Plant Physiol. 2015, 169, 530–548. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.X.; Cao, J.; Ma, Q.A.; Gao, X.J.; Ren, J.A.; Xue, Y. GPS-YNO2: Computational prediction of tyrosine nitration sites in proteins. Mol. Biosyst. 2011, 7, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Kolbert, Z.; Feigl, G.; Borde, A.; Molnar, A.; Erdei, L. Protein tyrosine nitration in plants: Present knowledge, computational prediction and future perspectives. Plant Physiol. Biochem. 2017, 113, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Gzyl, J.; Izbianska, K.; Floryszak-Wieczorek, J.; Jelonek, T.; Arasimowicz-Jelonek, M. Cadmium affects peroxynitrite generation and tyrosine nitration in seedling roots of soybean (Glycine max L.). Environ. Exp. Bot. 2016, 131, 155–163. [Google Scholar] [CrossRef]
- Malerba, M.; Cerana, R. Role of peroxynitrite in the responses induced by heat stress in tobacco BY-2 cultured cells. Protoplasma 2018, 255, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Berkowitz, G.A. NO and Ca2+: Critical Components of Cytosolic Signaling Systems Involved in Stomatal Immune Responses. In Nitric Oxide and Signaling in Plants; Wendehenne, D., Ed.; Academic Press Ltd.-Elsevier Science Ltd.: London, UK, 2016; Volume 77, pp. 285–323. [Google Scholar]
- Salmi, M.L.; Clark, G.; Roux, S.J. Current status and proposed roles for nitric oxide as a key mediator of the effects of extracellular nucleotides on plant growth. Front. Plant Sci. 2013, 4, 427. [Google Scholar] [CrossRef] [PubMed]
- Sueldo, D.J.; Foresi, N.P.; Casalongue, C.A.; Lamattina, L.; Laxalt, A.M. Phosphatidic acid formation is required for extracellular ATP-mediated nitric oxide production in suspension-cultured tomato cells. New Phytol. 2010, 185, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Testerink, C.; Munnik, T. Molecular, cellular, and physiological responses to phosphatidic acid formation in plants. J. Exp. Bot. 2011, 62, 2349–2361. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.C.; Ufer, G.D.; Bartels, D. Lipid signalling in plant responses to abiotic stress. Plant Cell Environ. 2016, 39, 1029–1048. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-J.; Wu, J.-Y. Extracellular ATP-induced NO production and its dependence on membrane Ca(2+) flux in Salvia miltiorrhiza hairy roots. J. Exp. Bot. 2008, 59, 4007–4016. [Google Scholar] [CrossRef] [PubMed]
- Jeandroz, S.; Lamotte, O.; Astier, J.; Rasul, S.; Trapet, P.; Besson-Bard, A.; Bourque, S.; Nicolas-Frances, V.; Ma, W.; Berkowitz, G.A.; et al. There’s More to the Picture Than Meets the Eye: Nitric Oxide Cross Talk with Ca2+ Signaling. Plant Physiol. 2013, 163, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Sawa, T.; Zaki, M.H.; Okamoto, T.; Akuta, T.; Tokutomi, Y.; Kim-Mitsuyama, S.; Ihara, H.; Kobayashi, A.; Yamamoto, M.; Fujii, S.; et al. Protein S-guanylation by the biological signal 8-nitroguanosine 3′,5′-cyclic monophosphate. Nat. Chem. Biol. 2007, 3, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Joudoi, T.; Shichiri, Y.; Kamizono, N.; Akaike, T.; Sawa, T.; Yoshitake, J.; Yamada, N.; Iwai, S. Nitrated Cyclic GMP Modulates Guard Cell Signaling in Arabidopsis. Plant Cell 2013, 25, 558–571. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.; Bhatla, S.C. Molecular mechanisms accompanying nitric oxide signalling through tyrosine nitration and S-nitrosylation of proteins in plants. Funct. Plant Biol. 2018, 45, 70–82. [Google Scholar] [CrossRef]
- Chivasa, S.; Simon, W.J.; Murphy, A.M.; Lindsey, K.; Carr, J.P.; Slabas, A.R. The effects of extracellular adenosine 5′-triphosphate on the tobacco proteome. Proteomics 2010, 10, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.S.N.; Ali, G.S.; Celesnik, H.; Day, I.S. Coping with Stresses: Roles of Calcium- and Calcium/Calmodulin-Regulated Gene Expression. Plant Cell 2011, 23, 2010–2032. [Google Scholar] [CrossRef] [PubMed]
- Lang, T.; Deng, S.R.; Zhao, N.; Deng, C.; Zhang, Y.N.; Zhang, Y.L.; Zhang, H.L.; Sa, G.; Yao, J.; Wu, C.W.; et al. Salt-Sensitive Signaling Networks in the Mediation of K+/Na+ Homeostasis Gene Expression in Glycyrrhiza uralensis Roots. Front. Plant Sci. 2017, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.R.; Tanaka, K.; Nguyen, C.T.; Stacey, G. Extracellular ATP is a central signaling molecule in plant stress responses. Curr. Opin. Plant Biol. 2014, 20, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, Y.; Kaya, H.; Hiraoka, G.; Yumoto, F.; Kimura, S.; Kadota, Y.; Hishinuma, H.; Senzaki, E.; Yamagoe, S.; Nagata, K.; et al. Synergistic activation of the Arabidopsis NADPH oxidase AtrbohD by Ca2+ and phosphorylation. J. Biol. Chem. 2008, 283, 8885–8892. [Google Scholar] [CrossRef] [PubMed]
- Balague, C.; Gouget, A.; Bouchez, O.; Souriac, C.; Haget, N.; Boutet-Mercey, S.; Govers, F.; Roby, D.; Canut, H. The Arabidopsis thaliana lectin receptor kinase LecRK-I.9 is required for full resistance to Pseudomonas syringae and affects jasmonate signalling. Mol. Plant Pathol. 2017, 18, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.T.; Ma, S.L.; Bai, L.P.; Zhang, L.; Ma, H.; Jia, P.; Liu, J.; Zhong, M.; Guo, Z.F. Signal transduction during cold, salt, and drought stresses in plants. Mol. Biol. Rep. 2012, 39, 969–987. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Oku, H.; Nahar, K.; Bhuyan, M.; Al Mahmud, J.; Baluska, F.; Fujita, M. Nitric oxide-induced salt stress tolerance in plants: ROS metabolism, signaling, and molecular interactions. Plant Biotechnol. Rep. 2018, 12, 77–92. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, X.; Deng, S.; Zhang, C.; Wang, M.; Ding, M.; Zhao, R.; Shen, X.; Zhou, X.; Lu, C.; et al. Extracellular ATP Signaling Is Mediated by H2O2 and Cytosolic Ca2+ in the Salt Response of Populus euphratica Cells. PLoS ONE 2012, 7, e53136. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Wang, S.J.; Ma, X.J.; Zhu, H.P.; Sa, G.; Sun, J.; Li, N.F.; Zhao, C.J.; Zhao, R.; Chen, S.L. Extracellular ATP mediates cellular K+/Na+ homeostasis in two contrasting poplar species under NaCl stress. Trees-Struct. Funct. 2016, 30, 825–837. [Google Scholar] [CrossRef]
- Zhang, X.; Shen, Z.D.; Sun, J.; Yu, Y.C.; Deng, S.R.; Li, Z.Y.; Sun, C.H.; Zhang, J.; Zhao, R.; Shen, X.; et al. NaCl-elicited, vacuolar Ca2+ release facilitates prolonged cytosolic Ca2+ signaling in the salt response of Populus euphratica cells. Cell Calcium 2015, 57, 348–365. [Google Scholar] [CrossRef] [PubMed]
- Lang, T.; Sun, H.M.; Li, N.Y.; Lu, Y.J.; Shen, Z.D.; Jing, X.S.; Xiang, M.; Shen, X.; Chen, S.L. Multiple signaling networks of extracellular ATP, hydrogen peroxide, calcium, and nitric oxide in the mediation of root ion fluxes in secretor and non-secretor mangroves under salt stress. Aquat. Bot. 2014, 119, 33–43. [Google Scholar] [CrossRef]
- Knight, M.R.; Campbell, A.K.; Smith, S.M.; Trewavas, A.J. Transgenic plant aequorin reports the effects of touch and cold-shock and elicitors on cytoplasmic calcium. Nature 1991, 352, 524–526. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Serrano, M.; Romero-Puertas, M.C.; Pazmino, D.M.; Testillano, P.S.; Risueno, M.C.; del Rio, L.A.; Sandalio, L.M. Cellular Response of Pea Plants to Cadmium Toxicity: Cross Talk between Reactive Oxygen Species, Nitric Oxide, and Calcium. Plant Physiol. 2009, 150, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.Z.; Ye, G.J.; Wang, R.F.; Jia, L.Y.; Liang, J.Y.; Feng, H.Q.; Wen, J.; Shi, D.L.; Wang, Q.W. Changes by cadmium stress in lipid peroxidation and activities of lipoxygenase and antioxidant enzymes in Arabidopsis are associated with extracellular ATP. Biologia 2017, 72, 1467–1474. [Google Scholar] [CrossRef]
- Wu, J.; Steinebrunner, I.; Sun, Y.; Butterfield, T.; Torres, J.; Arnold, D.; Gonzalez, A.; Jacob, F.; Reichler, S.; Roux, S.J. Apyrases (nucleoside triphosphate-diphosphohydrolases) play a key role in growth control in arabidopsis. Plant Physiol. 2007, 144, 961–975. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.; Wu, J.; Yao, J.; Gallardo, I.; Dugger, J.; Webb, L.; Huang, J.; Salmi, M.; Song, J.; Clark, G.; et al. Apyrase suppression raises extracellular ATP levels and induces gene expression and cell wall changes characteristic of stress responses. Plant Physiol. 2014, 164, 2054–2067. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.Q.; Brady, S.R.; Sun, Y.; Muday, G.K.; Roux, S.J. Extracellular ATP inhibits root gravitropism at concentrations that inhibit polar auxin transport. Plant Physiol. 2003, 131, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, J.; Clark, G.; Lundy, S.; Lim, M.; Arnold, D.; Chan, J.; Tang, W.Q.; Muday, G.K.; Gardner, G.; et al. Role for Apyrases in Polar Auxin Transport in Arabidopsis. Plant Physiol. 2012, 160, 1985–1995. [Google Scholar] [CrossRef] [PubMed]
- Tonon, C.; Cecilia Terrile, M.; Jose Iglesias, M.; Lamattina, L.; Casalongue, C. Extracellular ATP, nitric oxide and superoxide act coordinately to regulate hypocotyl growth in etiolated Arabidopsis seedlings. J. Plant Physiol. 2010, 167, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Terrile, M.; Tonón, C.V.; Iglesias, M.J.; Lamattina, L.; Casalongué, C.A. Extracellular ATP and nitric oxide signaling pathways regulate redox-dependent responses associated to root hair growth in etiolated Arabidopsis seedlings. Plant Signal. Behav. 2010, 5, 698–701. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Y.; Wang, B.C.; Farris, B.; Clark, G.; Roux, S.J. Modulation of Root Skewing in Arabidopsis by Apyrases and Extracellular ATP. Plant Cell Physiol. 2015, 56, 2197–2206. [Google Scholar] [CrossRef] [PubMed]
- Haruta, M.; Sussman, M.R. The Effect of a Genetically Reduced Plasma Membrane Protonmotive Force on Vegetative Growth of Arabidopsis. Plant Physiol. 2012, 158, 1158–1171. [Google Scholar] [CrossRef] [PubMed]
- Chivasa, S.; Ndimba, B.K.; Simon, W.J.; Lindsey, K.; Slabas, A.R. Extracellular ATP functions as an endogenous external metabolite regulating plant cell viability. Plant Cell 2005, 17, 3019–3034. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, C.-L.; Deng, S.-R.; Lu, C.-F.; Shen, X.; Zhou, X.-Y.; Zheng, X.-J.; Hu, Z.-M.; Chen, S.-L. An ATP signalling pathway in plant cells: Extracellular ATP triggers programmed cell death in Populus euphratica. Plant Cell Environ. 2012, 35, 893–916. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.Q.; Guan, D.D.; Sun, K.; Fang, Y.; Zhao, Y.; Jia, L.Y. Extracellular ATP is Involved in the Salicylic Acid-Induced Cell Death in Suspension-Cultured Tobacco Cells. Plant Prod. Sci. 2015, 18, 154–160. [Google Scholar] [CrossRef]
- Feng, H.Q.; Guan, D.D.; Bai, J.Y.; Jia, L.Y.; Fang, Y.; Sun, K. Extracellular ATP alleviates the salicylic acid-induced inhibition of cell viability and respiration through a Ca2+-dependent mechanism. Biol. Plant. 2015, 59, 193–197. [Google Scholar] [CrossRef]
- Feng, H.Q.; Jiao, Q.S.; Sun, K.; Jia, L.Y.; Tian, W.Y. Extracellular ATP affects chlorophyll fluorescence of kidney bean (Phaseolus vulgaris) leaves through Ca2+ and H2O2-dependent mechanism. Photosynthetica 2015, 53, 201–206. [Google Scholar] [CrossRef]
- Hou, Q.Z.; Sun, K.; Zhang, H.; Su, X.; Fan, B.Q.; Feng, H.Q. The responses of photosystem II and intracellular ATP production of Arabidopsis leaves to salt stress are affected by extracellular ATP. J. Plant Res. 2018, 131, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Q.S.; Feng, H.Q.; Tian, W.Y.; Bai, J.Y.; Sun, K.; Jia, L.Y.; Zhang, J.P. Extracellular ATP functions in alleviating the decrease of PSII photochemistry caused by the infection of Xanthomonas campestris pv. phaseoli. Plant Pathol. 2016, 65, 819–825. [Google Scholar] [CrossRef]
- Shope, J.C.; DeWald, D.B.; Mott, K.A. Changes in surface area of intact guard cells are correlated with membrane internalization. Plant Physiol. 2003, 133, 1314–1321. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.; Darwin, C.; Mehta, V.; Jackobs, F.; Perry, T.; Hougaard, K.; Roux, S. Effects of chemical inhibitors and apyrase enzyme further document a role for apyrases and extracellular ATP in the opening and closing of stomates in Arabidopsis. Plant Signal. Behav. 2013, 8, e26093. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Jia, J.J.; Wang, Y.F.; Wang, W.X.; Chen, Y.L.; Liu, T.; Shang, Z.L. Hyperpolization-activated Ca2+ channels in guard cell plasma membrane are involved in extracellular ATP-promoted stomatal opening in Vicia faba. J. Plant Physiol. 2014, 171, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Trapet, P.; Kulik, A.; Lamotte, O.; Jeandroz, S.; Bourque, S.; Nicolas-Frances, V.; Rosnoblet, C.; Besson-Bard, A.; Wendehenne, D. NO signaling in plant immunity: A tale of messengers. Phytochemistry 2015, 112, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.X.; Ma, X.Y.; Che, Y.M.; Hou, L.X.; Liu, X.; Zhang, W. Extracellular ATP mediates H2S-regulated stomatal movements and guard cell K+ current in a H2O2-dependent manner in Arabidopsis. Sci. Bull. 2015, 60, 419–427. [Google Scholar] [CrossRef]
- Knowles, A.F. The GDA1_CD39 superfamily: NTPDases with diverse functions. Purinergic Signal. 2011, 7, 21–45. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Sun, Y.; Naus, K.; Lloyd, A.; Roux, S. Apyrase functions in plant phosphate nutrition and mobilizes phosphate from extracellular ATP. Plant Physiol. 1999, 119, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Veerappa, R.; Slocum, R.D.; Siegenthaler, A.; Wang, J.; Clark, G.; Roux, S.J. Ectopic Expression of a Pea Apyrase Enhances Root System Architecture and Drought Survival in Arabidopsis and Soybean. Plant Cell Environ. 2018, in press. [Google Scholar] [CrossRef] [PubMed]
- Steinebrunner, I.; Wu, J.; Sun, Y.; Corbett, A.; Roux, S.J. Disruption of apyrases inhibits pollen germination in Arabidopsis. Plant Physiol. 2003, 131, 1638–1647. [Google Scholar] [CrossRef] [PubMed]
- Wolf, C.; Hennig, M.; Romanovicz, D.; Steinebrunner, I. Developmental defects and seedling lethality in apyrase AtAPY1 and AtAPY2 double knockout mutants. Plant Mol. Biol. 2007, 64, 657–672. [Google Scholar] [CrossRef] [PubMed]
- McAlvin, C.B.; Stacey, G. Transgenic expression of the soybean apyrase in Lotus japonicus enhances nodulation. Plant Physiol. 2005, 137, 1456–1462. [Google Scholar] [CrossRef] [PubMed]
- Govindarajulu, M.; Kim, S.Y.; Libault, M.; Berg, R.H.; Tanaka, K.; Stacey, G.; Taylor, C.G. GS52 Ecto-Apyrase Plays a Critical Role during Soybean Nodulation. Plant Physiol. 2009, 149, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- Riewe, D.; Grosman, L.; Fernie, A.R.; Wucke, C.; Geigenberger, P. The potato-specific apyrase is apoplastically localized and has influence on gene expression, growth, and development. Plant Physiol. 2008, 147, 1092–1109. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, K.; Yasunaga, E.; Niwa, M.; Ohwatari, Y.; Nakashima, A.; Inagaki, Y.; Ichinose, Y.; Shiraishi, T. H2O2 production by copper amine oxidase, a component of the ecto-apyrase (ATPase)-containing protein complex(es) in the pea cell wall, is regulated by an elicitor and a suppressor from Mycosphaerella pinodes. J. Gen. Plant Pathol. 2012, 78, 311–315. [Google Scholar] [CrossRef]
- Wu, S.; Peiffer, M.; Luthe, D.S.; Felton, G.W. ATP Hydrolyzing Salivary Enzymes of Caterpillars Suppress Plant Defenses. PLoS ONE 2012, 7, e41947. [Google Scholar] [CrossRef] [PubMed]
- Detoni, M.D.; Faria-Pinto, P.; Quellis, L.R.; Rust, N.M.; Tavares, L.S.; Santos, M.D.; Isaias, R.M.D.; Santos, J.C.; Fernandes, G.W.; Soares, G.L.G.; et al. Galls from Calliandra brevipes BENTH (Fabaceae: Mimosoidae): Evidence of apyrase activity contribution in a plant-insect interaction. Aust. J. Bot. 2012, 60, 559–567. [Google Scholar] [CrossRef]
- Guiguet, A.; Dubreuil, G.; Harris, M.O.; Appel, H.M.; Schultz, J.C.; Pereira, M.H.; Giron, D. Shared weapons of blood- and plant-feeding insects: Surprising commonalities for manipulating hosts. J. Insect Physiol. 2016, 84, 4–21. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.R.; Roux, S.J. Characterization of nucleoside triphosphatase-activity in isolated pea nuclei and its photoreversible regulation by light. Plant Physiol. 1986, 81, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.L.; Song, C.J.; Roux, S.J. Regulation of a recombinant pea nuclear apyrase by calmodulin and casein kinase II. Biochim. Biophys. Acta Gene Struct. Expr. 2000, 1494, 248–255. [Google Scholar] [CrossRef]
- Shibata, K.; Morita, Y.; Abe, S.; Stankovic, B.; Davies, E. Apyrase from pea stems: Isolation, purification, characterization and identification of a NTPase from the cytoskeleton fraction of pea stem tissue. Plant Physiol. Biochem. 1999, 37, 881–888. [Google Scholar] [CrossRef]
- Shibata, K.; Abe, S.; Davies, E. Structure of the coding region and mRNA variants of the apyrase gene from pea (Pisum sativum). Acta Physiol. Plant. 2001, 23, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Tong, C.G.; Dauwalder, M.; Clawson, G.A.; Hatem, C.L.; Roux, S.J. The major nucleoside triphosphatase in pea (Pisum-sativum L.) nuclei and in rat-liver nuclei share common epitopes also present in nuclear lamins. Plant Physiol. 1993, 101, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Tang, J.; Li, H.B. The presence and biological significance of extracellular calmodulin in cells. Chin. Sci. Bull. 1995, 40, 1153–1159. [Google Scholar]
- Biro, R.L.; Daye, S.; Serlin, B.S.; Terry, M.E.; Datta, N.; Sopory, S.K.; Roux, S.J. Characterization of oat calmodulin and radioimmunoassay of its subcellular-distribution. Plant Physiol. 1984, 75, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Hepler, P.K.; Kunkel, J.G.; Rounds, C.M.; Winship, L.J. Calcium entry into pollen tubes. Trends Plant Sci. 2012, 17, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Loro, G.; Drago, I.; Pozzan, T.; Lo Schiavo, F.; Zottini, M.; Costa, A. Targeting of Cameleons to various subcellular compartments reveals a strict cytoplasmic/mitochondrial Ca2+ handling relationship in plant cells. Plant J. 2012, 71, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.H.G.; Lioutas, A.; Le Dily, F.; Soronellas, D.; Pohl, A.; Bonet, J.; Nacht, A.S.; Samino, S.; Font-Mateu, J.; Vicent, G.P.; et al. ADP-ribose-derived nuclear ATP synthesis by NUDIX5 is required for chromatin remodeling. Science 2016, 352, 1221–1225. [Google Scholar] [CrossRef] [PubMed]
- Ali, G.S.; Reddy, A.S.N. ATP, phosphorylation and transcription regulate the mobility of plant splicing factors. J. Cell Sci. 2006, 119, 3527–3538. [Google Scholar] [CrossRef] [PubMed]
- Massalski, C.; Bloch, J.; Zebisch, M.; Steinebrunner, I. The Biochemical Properties of the Arabidopsis Ecto-Nucleoside Triphosphate Diphosphohydrolase AtAPY1 Contradict a Direct Role in Purinergic Signaling. PLoS ONE 2015, 10, e0115832. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Y.; Li, G.S.; Liang, S.F. Several Affinity Tags Commonly Used in Chromatographic Purification. J. Anal. Methods Chem. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clark, G.; Roux, S.J. Role of Ca2+ in Mediating Plant Responses to Extracellular ATP and ADP. Int. J. Mol. Sci. 2018, 19, 3590. https://doi.org/10.3390/ijms19113590
Clark G, Roux SJ. Role of Ca2+ in Mediating Plant Responses to Extracellular ATP and ADP. International Journal of Molecular Sciences. 2018; 19(11):3590. https://doi.org/10.3390/ijms19113590
Chicago/Turabian StyleClark, Greg, and Stanley J. Roux. 2018. "Role of Ca2+ in Mediating Plant Responses to Extracellular ATP and ADP" International Journal of Molecular Sciences 19, no. 11: 3590. https://doi.org/10.3390/ijms19113590
APA StyleClark, G., & Roux, S. J. (2018). Role of Ca2+ in Mediating Plant Responses to Extracellular ATP and ADP. International Journal of Molecular Sciences, 19(11), 3590. https://doi.org/10.3390/ijms19113590