

Complement Modulation Mitigates Inflammation-Mediated Preterm Birth and Fetal Neural Inflammation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Study Design
2.2. Animal Husbandry and Care
2.3. Model of Preterm Birth
2.4. Tissue Processing
2.5. Immunofluorescence Staining and Imaging
2.6. Recombinant Protein
2.7. Cytokine Assays
2.8. Statistical Analysis
3. Results
3.1. Complement Activation Increases over Time in Model of Preterm Birth
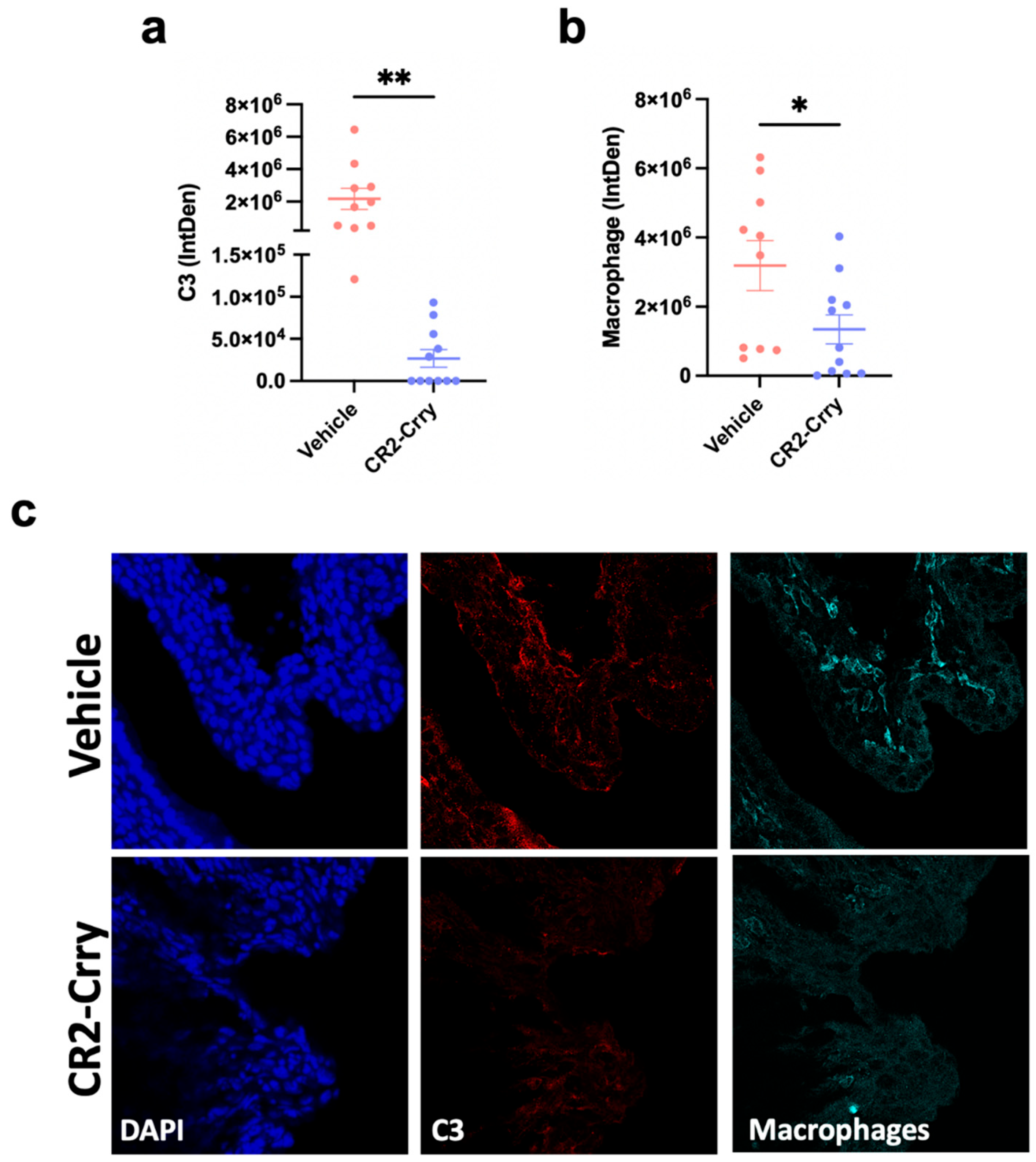
3.2. Effect of CR2-Crry on Preterm Birth Delivery Outcomes and Cellular Changes
3.3. Mitigation of Pro-Inflammatory Maternal and Fetal Response in Model of Preterm Birth
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galinsky, R.; Polglase, G.R.; Hooper, S.B.; Black, M.J.; Moss, T.J.M. The consequences of chorioamnionitis: Preterm birth and effects on development. J. Pregnancy 2013, 2013, 412831. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.A.; Michelle, J.K.; Osterman, M.H.S. Shifts in the Distribution of Births by Gestational Age: United States, 2014–2022. Natl. Vital Stat. Rep. 2024, 73, 1. [Google Scholar]
- Kim, C.J.; Romero, R.; Chaemsaithong, P.; Chaiyasit, N.; Yoon, B.H.; Kim, Y.M. Acute chorioamnionitis and funisitis: Definition, pathologic features, and clinical significance. Am. J. Obstet. Gynecol. 2015, 213, S29–S52. [Google Scholar] [PubMed]
- Bastek, J.A.; Gómez, L.M.; Elovitz, M.A. The role of inflammation and infection in preterm birth. Clin. Perinatol. 2011, 38, 385–406. [Google Scholar] [CrossRef] [PubMed]
- Van Hoeven, K.H.; Anyaegbunam, A.; Whitty, J.E.; Distant, J.; Crawford, C. Clinical significance of increasing histologic severity of acute inflammation in the fetal membranes and umbilical cord. Pediatr. Pathol. Lab. Med. 1996, 16, 731–744. [Google Scholar]
- Srinivas, S.K.; Ma, Y.; Sammel, M.D.; Chou, D.; McGrath, C.; Parry, S.; Elovitz, M.A. Placental inflammation and viral infection are implicated in second trimester pregnancy loss. Am. J. Obstet. Gynecol. 2006, 195, 797–802. [Google Scholar] [CrossRef]
- Lee, S.M.; Park, J.W.; Kim, B.J.; Park, C.-W.; Park, J.S.; Jun, J.K.; Yoon, B.H.; Baud, O. Acute histologic chorioamnionitis is a risk factor for adverse neonatal outcome in late preterm birth after preterm premature rupture of membranes. PLoS ONE 2013, 8, e79941. [Google Scholar]
- Prediction and Prevention of Spontaneous Preterm Birth: ACOG Practice Bulletin, Number 234. Obstet. Gynecol. 2021, 138, e65–e90. [CrossRef]
- Girardi, G.; Lingo, J.J.; Fleming, S.D.; Regal, J.F. Essential Role of Complement in Pregnancy: From Implantation to Parturition and Beyond. Front. Immunol. 2020, 11, 1681. [Google Scholar]
- Elimian, A.; Figueroa, R.; Canterino, J.; Verma, U.; Aguero-Rosenfeld, M.; Tejani, N. Amniotic fluid complement c3 as a marker of intra-amniotic infection. Obstet. Gynecol. Surv. 1998, 92, 72–76. [Google Scholar] [CrossRef]
- Gonzalez, J.M.; Dong, Z.; Romero, R.; Girardi, G. Cervical remodeling/ripening at term and preterm delivery: The same mechanism initiated by different mediators and different effector cells. PLoS ONE 2011, 6, e26877. [Google Scholar] [CrossRef]
- Gonzalez, J.M.; Franzke, C.W.; Yang, F.; Romero, R.; Girardi, G. Complement activation triggers metalloproteinases release inducing cervical remodeling and preterm birth in mice. Am. J. Pathol. 2011, 179, 838–849. [Google Scholar]
- Pedroni, S.M.A.; Gonzalez, J.M.; Wade, J.; Jansen, M.A.; Serio, A.; Marshall, I.; Lennen, R.J.; Girardi, G. Complement inhibition and statins prevent fetal brain cortical abnormalities in a mouse model of preterm birth. Biochim. Biophys. Acta—Mol. Basis Dis. 2014, 1842, 107–115. [Google Scholar]
- Tomlinson, S.; Thurman, J. Tissue-Targeted Complement Therapeutics. Mol Immunol 2018, 102, 120–128. [Google Scholar] [PubMed]
- Atkinson, C.; Song, H.; Lu, B.; Qiao, F.; Burns, T.A.; Holers, V.M.; Tsokos, G.C.; Tomlinson, S. Targeted complement inhibition by C3d recognition ameliorates tissue injury without apparent increase in susceptibility to infection. J. Clin. Investig. 2005, 115, 2444–2453. [Google Scholar]
- McDougall, A.R.A.; Hastie, R.; Goldstein, M.; Tuttle, A.; Ammerdorffer, A.; Gülmezoglu, A.M.; Vogel, J.P. New medicines for spontaneous preterm birth prevention and preterm labour management: Landscape analysis of the medicine development pipeline. BMC Pregnancy Childbirth 2023, 23, 525. [Google Scholar] [CrossRef]
- Liu, F.; Ryan, S.T.; Fahnoe, K.C.; Morgan, J.G.; Cheung, A.E.; Storek, M.J.; Best, A.; Chen, H.A.; Locatelli, M.; Xu, S.; et al. C3d-Targeted factor H inhibits tissue complement in disease models and reduces glomerular injury without affecting circulating complement. Mol. Ther. 2024, 32, 1061–1079. [Google Scholar] [CrossRef]
- Hatchell, D.; Alshareef, M.; Vasas, T.; Guglietta, S.; Borucki, D.; Guo, C.; Mallah, K.; Eskandari, R.; Tomlinson, S. A role for P-selectin and complement in the pathological sequelae of germinal matrix hemorrhage. J. Neuroinflammation 2023, 20, 143. [Google Scholar] [CrossRef]
- Elovitz, M.A.; Wang, Z.; Chien, E.K.; Rychlik, D.F.; Phillippe, M. A New Model for Inflammation-Induced Preterm Birth: The Role of Platelet-Activating Factor and Toll-Like Receptor-4. Am. J. Pathol. 2003, 163, 2103–2111. [Google Scholar]
- Weber, E.M.; Algers, B.; Hultgren, J.; Olsson, I.A.S. Pup mortality in laboratory mice--infanticide or not? Acta Vet Scand. 2013, 55, 83. [Google Scholar] [CrossRef]
- Alawieh, A.; Langley, E.F.; Weber, S.; Adkins, D.; Tomlinson, S. Identifying the role of complement in triggering neuroinflammation after traumatic brain injury. J. Neurosci. 2018, 38, 2519–2532. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Qiao, F.; Atkinson, C.; Holers, V.M.; Tomlinson, S. A Novel Targeted Inhibitor of the Alternative Pathway of Complement and Its Therapeutic Application in Ischemia/Reperfusion Injury. J. Immunol. 2008, 181, 8068–8076. [Google Scholar] [CrossRef]
- Gonzalez, J.M.; Pedroni, S.M.A.; Girardi, G. Statins prevent cervical remodeling, myometrial contractions and preterm labor through amechanism that involves hemoxygenase-1 and complement inhibition. Mol. Hum. Reprod. 2014, 20, 579–589. [Google Scholar]
- Galindo-Sevilla, N.; Reyes-Arroyo, F.; Mancilla-Ramírez, J. The role of complement in preterm birth and prematurity. J. Perinat. Med. 2019, 47, 793–803. [Google Scholar]
- Choi, G.B.; Yim, Y.S.; Wong, H.; Kim, S.; Kim, H.; Kim, S.V.; Hoeffer, C.A.; Littman, D.R.; Huh, J.R. The maternal interleukin-17a pathway in mice promotes autism-like phenotypes in offspring. Science 2016, 351, 933–939. [Google Scholar] [CrossRef] [PubMed]
- MacDorman, M.F.; Gregory, E.C.W. Fetal and perinatal mortality: United States, 2013. Natl. Vital Stat. Rep. 2015, 64, 1–24. [Google Scholar] [PubMed]
- Goldenberg, R.L.; Culhane, J.F.; Iams, J.D.; Romero, R. Epidemiology and causes of preterm birth. Lancet 2008, 371, 75–84. [Google Scholar] [CrossRef]
- Kovács, K.; Kovács, Ő.Z.; Bajzát, D.; Imrei, M.; Nagy, R.; Németh, D.; Kói, T.; Szabó, M.; Fintha, A.; Hegyi, P.; et al. The histologic fetal inflammatory response and neonatal outcomes: Systematic review and meta-analysis. Am. J. Obstet. Gynecol. 2024, 230, 493–511.e3. [Google Scholar] [CrossRef]
- Yoon, B.H.; Romero, R.; Park, J.S.; Kim, C.J.; Kim, S.H.; Choi, J.-H.; Han, T.R. Fetal exposure to an intra-amniotic inflammation and the development of cerebral palsy at the age of three years. Am. J. Obstet. Gynecol. 2000, 182, 675–681. [Google Scholar]
- Goldenberg, R.L.; Hauth, J.C.; Andrews, W.W. Intrauterine Infection and Preterm Delivery. N. Engl. J. Med. 2000, 342, 1500–1507. [Google Scholar] [CrossRef]
- Stygar, D.; Wang, H.; Vladic, Y.S.; Ekman, G.; Eriksson, H.; Sahlin, L. Increased level of matrix metalloproteinases 2 and 9 in the ripening process of the human cervix. Biol. Reprod. 2002, 67, 889–894. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-J.; Jung, K.-L.; Oh, S.-Y.; Kim, J.-H.; Roh, C.-R. Cervicovaginal matrix metalloproteinase-9 and cervical ripening in human term parturition. Eur. J. Obstet. Gynecol. Reprod. Biol. 2009, 142, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Dubicke, A.; Ekman-Ordeberg, G.; Mazurek, P.; Miller, L.; Yellon, S.M. Density of Stromal Cells and Macrophages Associated with Collagen Remodeling in the Human Cervix in Preterm and Term Birth. Reprod. Sci. 2016, 23, 595–603. [Google Scholar] [CrossRef]
- Girardi, G.; Yarilin, D.; Thurman, J.M.; Holers, V.M.; Salmon, J.E. Complement activation induces dysregulation of angiogenic factors and causes fetal rejection and growth restriction. J. Exp. Med. 2006, 203, 2165–2175. [Google Scholar] [CrossRef]
- Yoon, B.H.; Jun, J.K.; Romero, R.; Park, K.H.; Gomez, R.; Choi, J.-H.; Kim, I.-O. Amniotic fluid inflammatory cytokines (interleukin-6, interleukin-1β, and tumor necrosis factor-α), neonatal brain white matter lesions, and cerebral palsy. Am. J. Obstet. Gynecol. 1997, 177, 19–26. [Google Scholar] [PubMed]
- Yoon, B.H.; Romero, R.; Kim, C.J.; Jun, J.K.; Gomez, R.; Choi, J.-H.; Syu, H.C. Amniotic fluid interleukin-6: A sensitive test for antenatal diagnosis of acute inflammatory lesions of preterm placenta and prediction of perinatal morbidity. Am. J. Obstet. Gynecol. 1995, 172, 960–970. [Google Scholar]
- Romero, R.; Miranda, J.; Chaiworapongsa, T.; Chaemsaithong, P.; Gotsch, F.; Dong, Z.; Ahmed, A.I.; Yoon, B.H.; Hassan, S.S.; Kim, C.J.; et al. A Novel Molecular Microbiologic Technique for the Rapid Diagnosis of Microbial Invasion of the Amniotic Cavity and Intra-Amniotic Infection in Preterm Labor with Intact Membranes. Am. J. Reprod. Immunol. 2014, 71, 330–358. [Google Scholar] [CrossRef]
- Pataky, R.; Howie, F.A.; Girardi, G.; Boardman, J.P. Complement C5a is present in CSF of human newborns and is elevated in association with preterm birth. J. Matern. Fetal Neonatal Med. 2017, 30, 2413–2416. [Google Scholar] [CrossRef]
- Yoon, B.H.; Kim, C.J.; Romero, R.; Jun, J.K.; Park, K.H.; Choi, S.T.; Chi, J.G. Experimentally induced intrauterine infection causes fetal brain white matter lesions in rabbits. Am. J. Obstet. Gynecol. 1997, 177, 797–802. [Google Scholar] [CrossRef]
- Alshareef, M.; Mallah, K.; Vasas, T.; Alawieh, A.; Borucki, D.; Couch, C.; Cutrone, J.; Shope, C.; Eskandari, R.; Tomlinson, S. A Role of Complement in the Pathogenic Sequelae of Mouse Neonatal Germinal Matrix Hemorrhage. Int. J. Mol. Sci. 2022, 23, 2943. [Google Scholar] [CrossRef]
- Alshareef, M.; Hatchell, D.; Vasas, T.; Mallah, K.; Shingala, A.; Cutrone, J.; Alawieh, A.; Guo, C.; Tomlinson, S.; Eskandari, R. Complement Drives Chronic Inflammation and Progressive Hydrocephalus in Murine Neonatal Germinal Matrix Hemorrhage. Int. J. Mol. Sci. 2023, 24, 10171. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McElwee, E.R.; Hatchell, D.; Alshareef, M.; Mallah, K.; Hall, H.; Robinson, H.; Eskandari, R.; Chang, E.; Sullivan, S.; Tomlinson, S. Complement Modulation Mitigates Inflammation-Mediated Preterm Birth and Fetal Neural Inflammation. Cells 2025, 14, 1045. https://doi.org/10.3390/cells14141045
McElwee ER, Hatchell D, Alshareef M, Mallah K, Hall H, Robinson H, Eskandari R, Chang E, Sullivan S, Tomlinson S. Complement Modulation Mitigates Inflammation-Mediated Preterm Birth and Fetal Neural Inflammation. Cells. 2025; 14(14):1045. https://doi.org/10.3390/cells14141045
Chicago/Turabian StyleMcElwee, Eliza R., Devin Hatchell, Mohammed Alshareef, Khalil Mallah, Harriet Hall, Hannah Robinson, Ramin Eskandari, Eugene Chang, Scott Sullivan, and Stephen Tomlinson. 2025. "Complement Modulation Mitigates Inflammation-Mediated Preterm Birth and Fetal Neural Inflammation" Cells 14, no. 14: 1045. https://doi.org/10.3390/cells14141045
APA StyleMcElwee, E. R., Hatchell, D., Alshareef, M., Mallah, K., Hall, H., Robinson, H., Eskandari, R., Chang, E., Sullivan, S., & Tomlinson, S. (2025). Complement Modulation Mitigates Inflammation-Mediated Preterm Birth and Fetal Neural Inflammation. Cells, 14(14), 1045. https://doi.org/10.3390/cells14141045