
Recent Clinical Advances on Long Non-Coding RNAs in Triple-Negative Breast Cancer


1
Amity Institute of Biotechnology, Amity University Rajasthan, Jaipur 303002, India
2
Department of Food and Nutrition, College of Bionano Technology, Gachon University, Seongnam-si 13120, Republic of Korea
3
Arontier Co., Seoul 06735, Republic of Korea
*
Authors to whom correspondence should be addressed.
Cells 2023, 12(4), 674; https://doi.org/10.3390/cells12040674
Submission received: 21 December 2022
/
Revised: 13 February 2023
/
Accepted: 17 February 2023
/
Published: 20 February 2023
(This article belongs to the Special Issue Long Non-coding RNAs in Cancer Metastasis)
Abstract
:Triple-negative breast cancer (TNBC) is a more aggressive type of breast cancer due to its heterogeneity and complex molecular mechanisms. TNBC has a high risk for metastasis, and it is difficult to manage clinical conditions of the patients. Various investigations are being conducted to overcome these challenges using RNA, DNA, and proteins for early diagnosis and treatment. Recently, long non-coding RNAs (lncRNAs) have emerged as a novel target to treat the multistep process of TNBC. LncRNAs regulate epigenetic expression levels, cell proliferation and apoptosis, and tumour invasiveness and metastasis. Thus, lncRNA-based early diagnosis and treatment options could be helpful, especially for patients with severe TNBC. lncRNAs are expressed in a highly specific manner in cells and tissues and are involved in TNBC progression and development. lncRNAs could be used as sensitive and specific targets for diagnosis, treatment, and monitoring of patients with TNBC. Therefore, the exploration of novel diagnostic and prognostic biomarkers is of extreme importance. Here, we discuss the molecular advances on lncRNA regulation of TNBC and lncRNA-based early diagnosis, treatment, and drug resistance.
1. Introduction
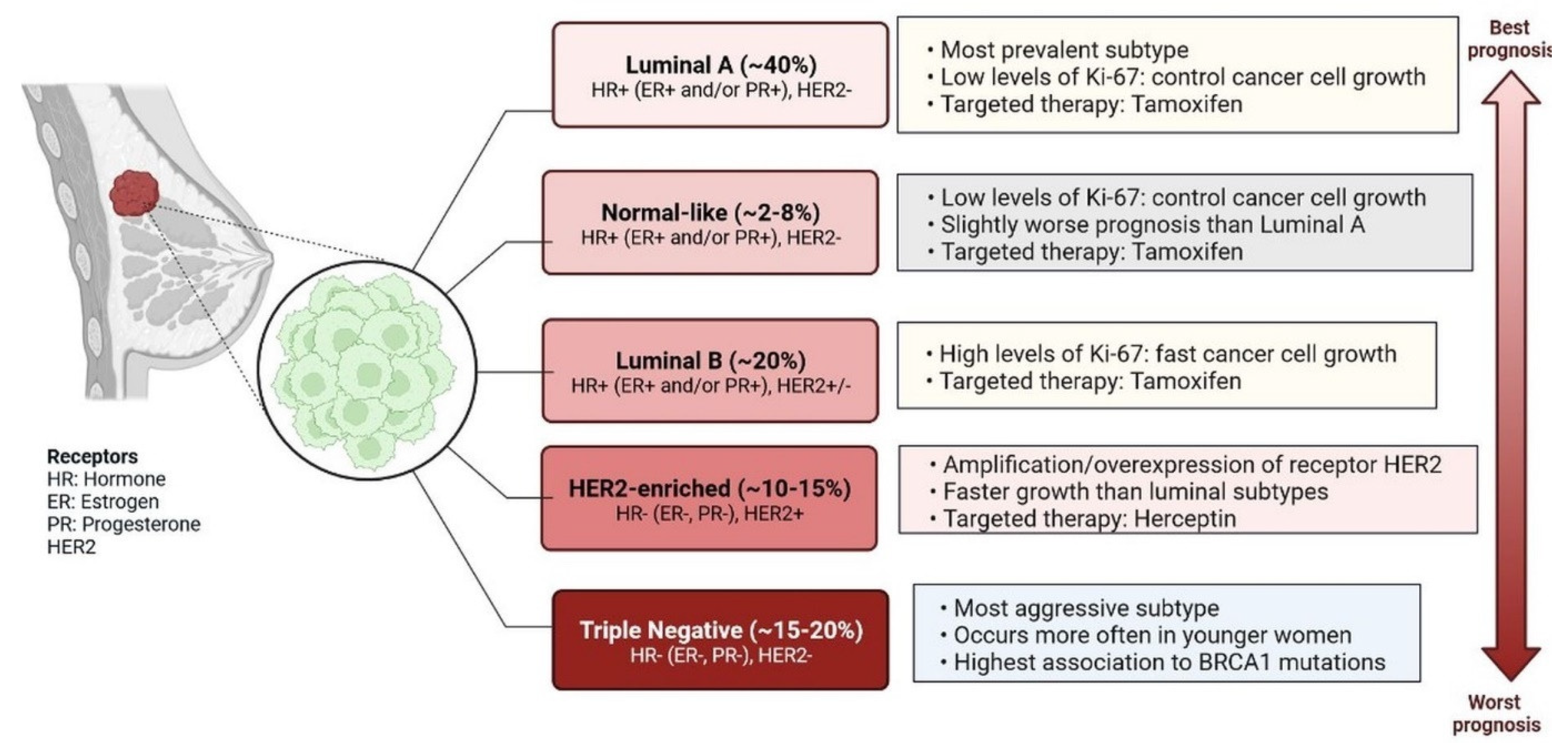
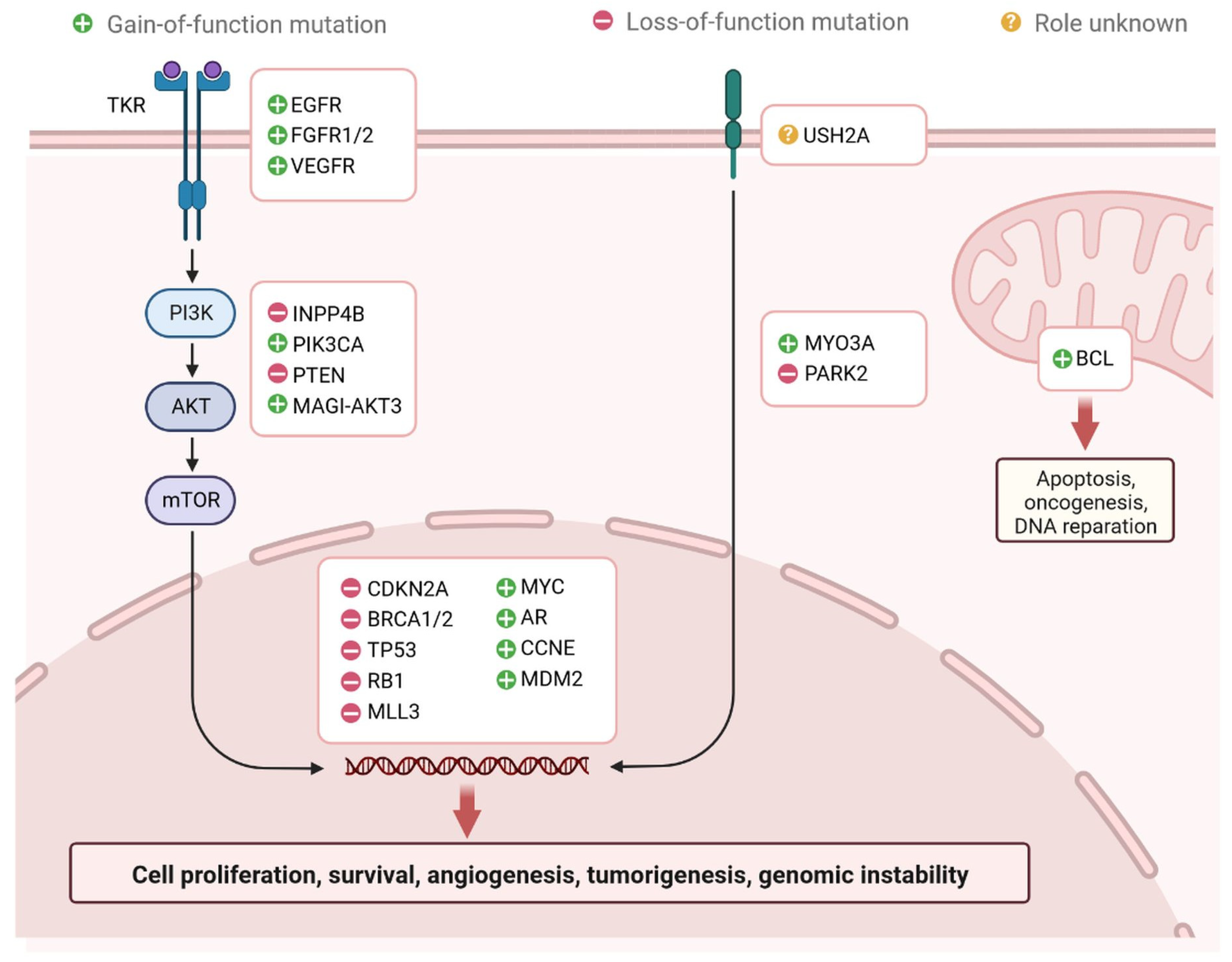
Breast cancer (BC) is caused by epigenetic modifications and is a highly heterogeneous disease. It exhibits various histological and clinical features [1]. There are five main intrinsic molecular subtypes of BC. To my knowledge, the five intrinsic molecular subtypes are: Luminal A, Luminal B, HER2-enriched, Triple-negative/Basal- like, and Claudin-low. Among these, Luminal A breast cancers are HER2-negative (HR+/HER2−) and include ER+/PR+, ER+/PR−, and ER−/PR+ status. (Figure 1) [2]. Triple-negative breast cancer (TNBC) is the most aggressive molecular clinical subtype of all invasive cancers. Various genetic markers are expressed during the development of TNBC (Figure 2) [3], which are involved in a gain or loss of function. Genetic markers involved in gain of function include EGFR, EGFR1/2, VEGFR, MYC, AR, CCNE, MDM2, PIK3CA, MAGI-AKT3, MYO3A, PARK2, and genetic markers involved in loss of function are INPP4B, PTEN, CDKN2A, BRCA1/2, TP53, RB1, and MLL3 (Figure 2) [4]. Advances in genetic heterogeneity research have revealed prognostic and therapeutic targets in TNBC [1,2,3]. TNBC has been divided into four distinct subtypes using gene expression analysis: basal-like immune-activated (BLIA), basal-like immune-suppressed (BLIS), mesenchymal (MES), and luminal androgen receptor (LAR). The classification of TNBC subtypes lacks a standardised system, however. Basal-like (BL) tumours make up around 80% of TNBC tumours, and because BL tumours cluster physiologically apart from the other BC subtypes, intrinsic subtyping is less effective for significant diagnosis and classification than it is for the other clinical subtypes. Multiple initiatives to investigate DNA, RNA, microRNA, and protein expression patterns through cross-platform research such as the Cancer Genome Atlas have provided more comprehensive evidence of BC heterogeneity. TNBC subtyping is useful for classifying patients for individualised care. However, research is ongoing, and no laboratory approach for classifying TNBC subtypes has yet been used in clinical settings. The overall malignancy indicated above was used to evaluate the initial and modified versions of the Lehman classifier, and the results indicated that non-cancer cells strongly influence the gene expression profiles that affect treatment response and prognosis in TNBC. The interrelations between cancer cells and the immune system, originally known as “immune surveillance”, provide an excellent example of the significance of each element of the tumour microenvironment on behaviour and prognosis. Not all patients respond well to immunotherapy, even though TNBC is the most immune-activated subtype of all BC, as shown by immune gene expression as well as levels of intra- and stromal tumour-infiltrating lymphocytes (TILs). The appropriate way to define immune activation is still being worked out, but current methods include looking for TILs, producing the protein programmed death ligand 1 (PDL1), immune gene signatures, individual immune gene RNA expression, and immune cell clonality (T cells and B cells) investigations. The TNBC subtype is still requires further investigations to improve treatment responder binding proteins are highly conserved, and binds with coding and non-coding RNAs through their RNA binding domain and metabolic process such as RNA splicing, polyadenylation, localization, translation, and destruction. Various types of non-coding RNAs (ncRNAs) have been investigated, including microRNAs (miRNAs), ribosomal RNAs (rRNAs), transfer RNAs (tRNAs), small interfering RNAs (siRNAs), small nuclear RNAs (snRNAs), extracellular RNAs (exRNAs), small Cajal body-specific RNAs, circular RNAs (circRNAs), and long non-coding RNAs (lncRNAs) [5,6,7,8,9,10,11,12]. ncRNAs are seen in a variety of malignancies, which are actively involved in cell proliferation and inhibition of tumour development [8]. RBPs are actively involved in regulations of TNBC at the transcriptional and post-transcriptional level [8]. The hnRNP E1 RBPs interact with PNUTS pre-RNA and suppress the splicing of lncRNA-PNUTS and regulates EMT (epithelial–mesenchymal transition) and promotes tumour development via interacting with miR-205 as a competitive sponge [8,9]. Additionally, N6-methyladenosine (m6A) readers, writers, and erasers are all RBPs that vary the functionality of lncRNAs by changing their expression levels. RBPs have different mechanisms to regulate the expression of lncRNAs. Breast cancer prognostic markers for autophagy, aerobic glycolysis, stemness, and immune-related lncRNA have been established [8,9]. Transcribed genomes can produce thousands of lncRNAs, which contain more than 200 nucleotides. The first lncRNA was identified in 1990 in a mouse model [6]. lncRNAs are found in cytosolic or nuclear regions as interspersed, overlapping regions of coding and non-coding transcripts. They have different molecular functions and roles, including molecular signalling, scaffolding, acting as decoys, integrating developmental signals, clarifying the cellular context, guiding gene expression, or responding to different stimuli [7]. Understanding of scaffolding complexes would provide novel strategies for the implementation of specific signalling components to alter molecular processes [7,8]. Sense, antisense, and bidirectional as well as intronic and intergenic lncRNAs have been observed, and they all participate in various cellular processes. lncRNAs, including NRON, HEIH, HCP5, LINC00096, growth-stasis-specific transcript 5 (GAS5), NEAT1, AWPPH, LUCAT1, HAND2-AS1, POU3F3, MALAT1, and ANRIL, are actively involved in TNBC apoptosis and proliferation [10,11,12,13,14,15,16]. All these lncRNAs could be potential targets for diagnosis and drug development against TNBC cells. Several studies have reported that lncRNAs play important roles in TNBC disease progression through various gene regulatory mechanisms and the induction of intramolecular interactions. The aberrant expression of lncRNAs is involved in TNBC initiation, progression, and metastasis, and affects various biomarkers. Therefore, lncRNAs are important for early diagnosis and the clinical management of patients.
2. LncRNAs
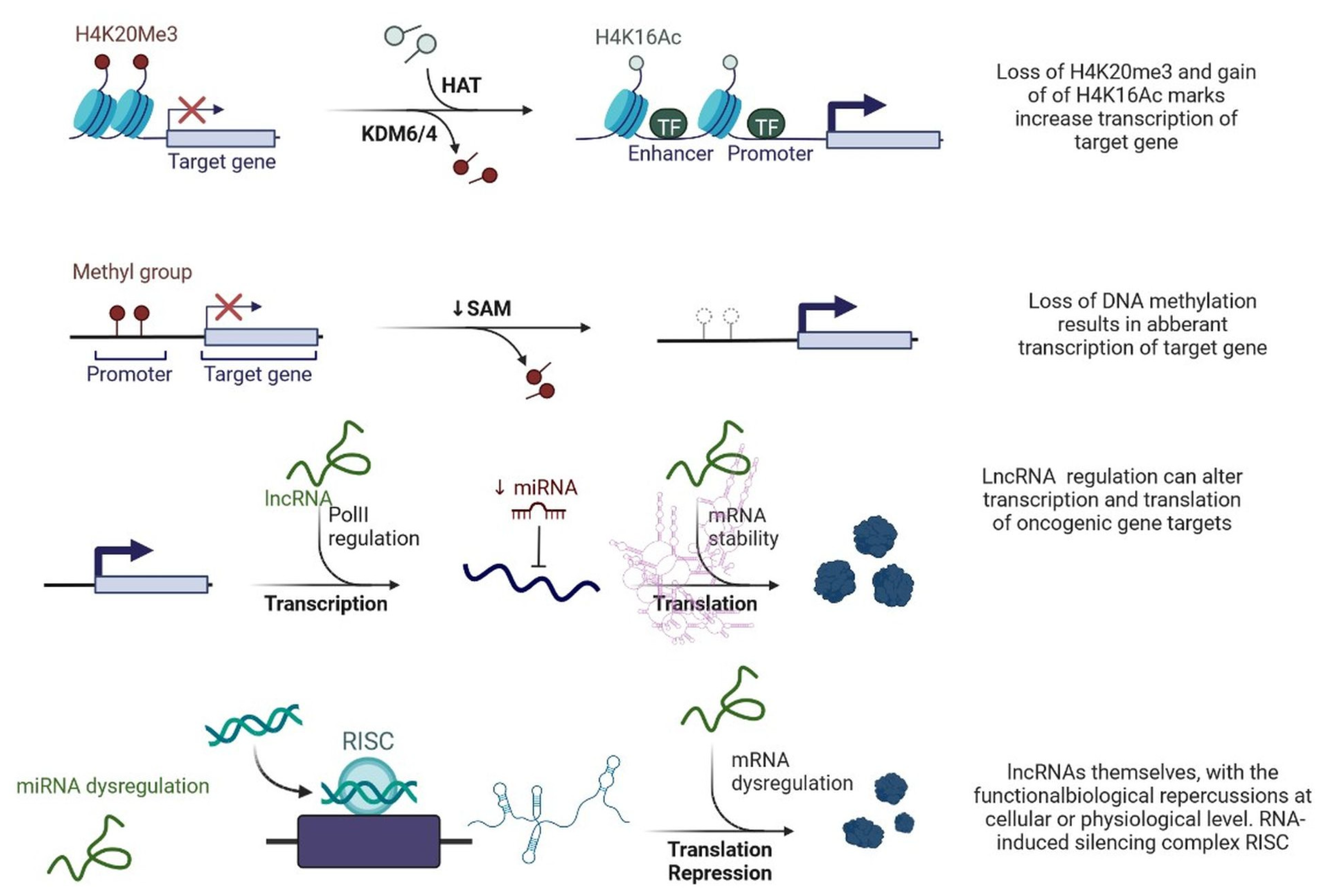
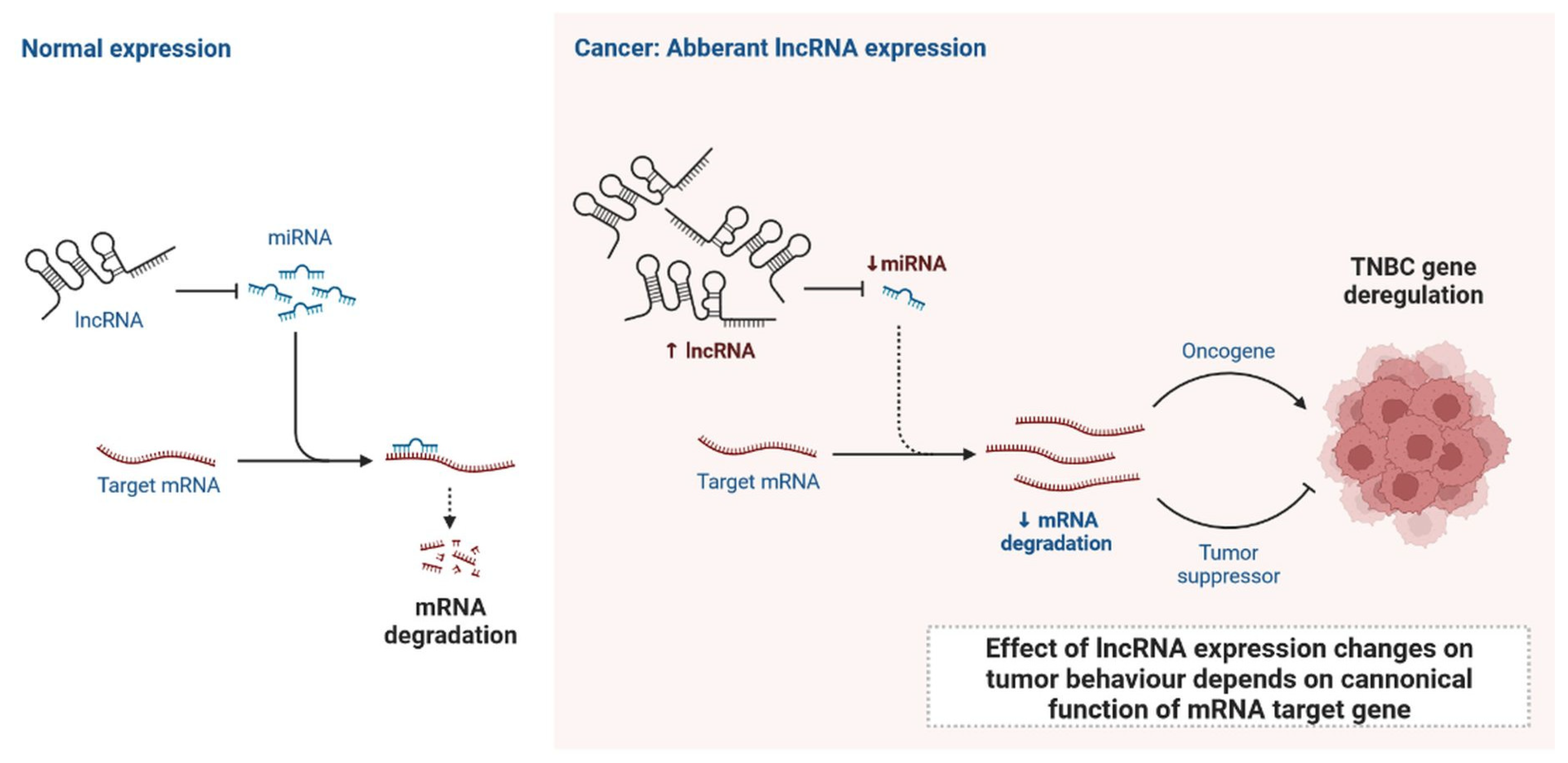
lncRNAs are actively involved in gene expression, epigenetic deregulation, chromatin remodelling, DNA methylation, translation of oncogenic gene targets, and biogenesis (Figure 3). They are transcribed by RNA polymerase II, after which most transcripts are spliced, and are mainly found in the nucleus and chromatin, being expressed in cells and tissues in a specific manner [6,9,17]. Transcriptional regulation and various molecular processes in the cytoplasm are controlled by lncRNAs; various circulating lncRNAs are transmitted via exosomes and bind to various transcription factors, chromatin-regulated complexes, RNA-binding proteins, nascent RNA transcripts, and chromatin [17]. The normal expression of lncRNAs and the effect of their expression changes on tumour behaviour depends on the canonical function of the mRNA target genes (Figure 4). lncRNAs can bind to the active site of proteins and regulate molecular processes at the post-transcriptional level. They are involved in functional biological processes at the cellular or physiological levels. RNA-induced silencing complexes (RISCs) are formed with the help of lysine-specific demethylase 5B (KDM5B, also known as histone demethylase JARID1B), trimethylation of lysine 4 on the histone H3 protein subunit (H3K4me3), monomethylation of lysine 4 on the histone H3 protein subunit (H3K4me1), hsa-miR-448 (also known as miRNA448), breast cancer 1/2 (BRCA1/2), retinoblastoma protein (pRB), caveolin-1 (CAV-1), Homeobox protein Hox-A5 (HOXA5), Stratifin (SFN), methyl groups (CH3), and Ras homolog gene family, member A (RhoA) (Figure 3 and Figure 5) [18]. In 2019, it was found that the lncRNA MIR100HG regulates proliferation in TNBC and the expression of the p27 gene after formation of an RNA–DNA triplex at the promoter [19]. Moreover, MIR100HG silencing leads to reduced transcription and translation of p27 [19,20]. Three triplex-forming oligonucleotides (TFOs) have been observed on the lncRNA of p27, which binds to the triplex-targeting ability (TTA) site at the 5’UTR; this event has been observed in TNBC cell lysates [21]. The binding of TFO1 and TTA is a unique mechanism by which MIR100HG regulates the transcription factors at the promoter region of p27 [21,22]. Plasmacytoma variant translocation 1 (PVT1) is another type of lncRNA that is transcribed by a gene situated at the 8q24 chromosomal region and plays and important role in TNBC development. It contains 12 exons that when spliced generate lncRNAs [23]. PVT1 binds to Krüppel-like factor 5 (KLF5) and generates a BAP1 deubiquitinase that induces TNBC via beta-catenin upregulation. Furthermore, the PVT1 promoter also acts as a regulator of the expression of the MYC proto-oncogene and BHLH transcription factor (c-MYC) [24]. These findings show that lncRNAs also mediate regulation at the transcriptional level.
3. Clinical Updates on lncRNAs in TNBC
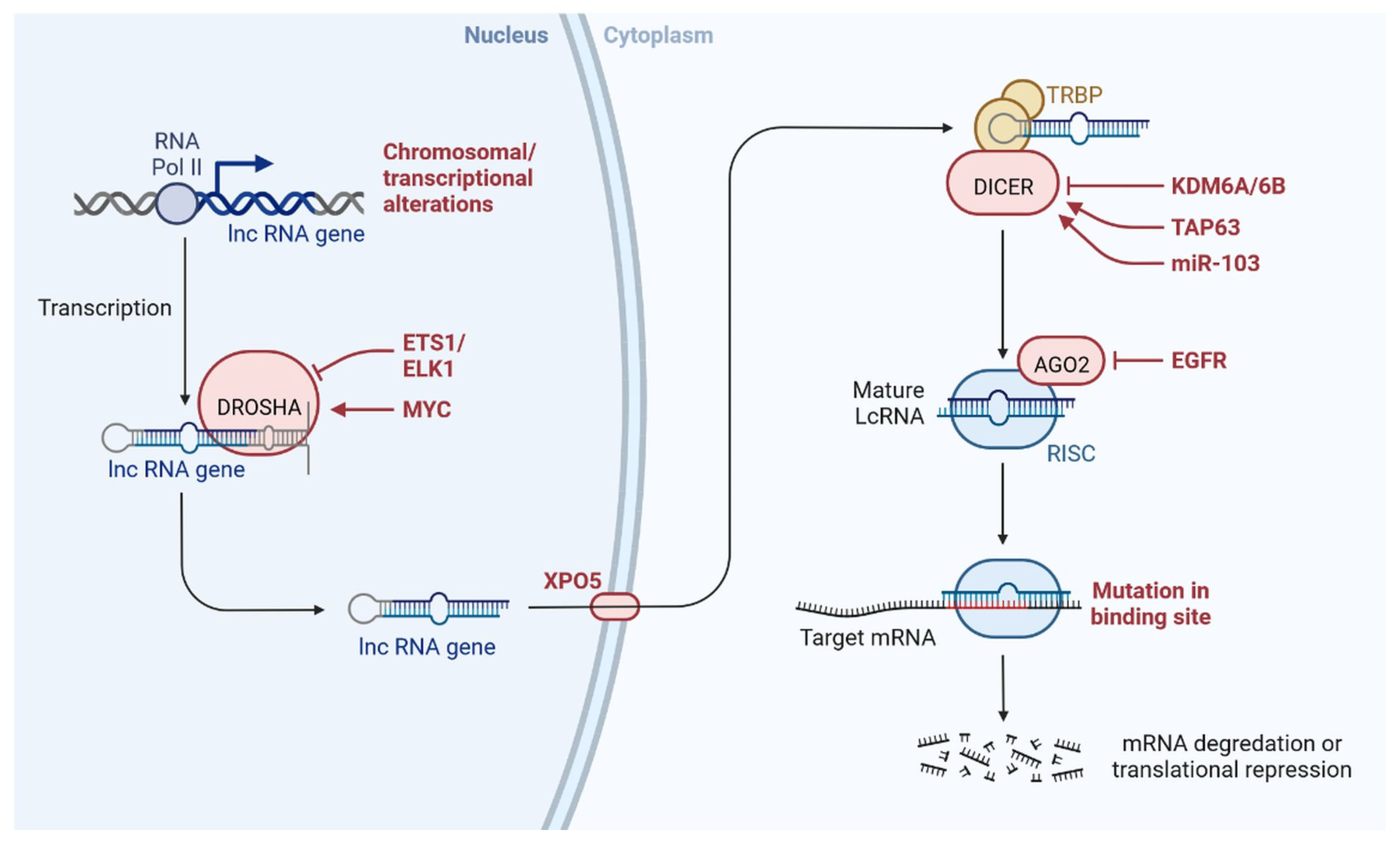
Recently, lncRNA expression in patients with TNBC was investigated; 1034 lncRNAs were identified using NGS technologies and microarrays, out of which, 537 lncRNAs regulate 451 protein-coding genes [14]. These genes are also detected in TNBC cells and are involved in cell signalling pathways such as the MAPK and PI3K-Akt pathways, which may lead to heterogeneity [14,24]. lncRNAs also act as miRNAs, binding to miRNA-targeted mRNAs and dysregulated miRNAs [25]. This crosstalk forms a complex post-transcriptional regulatory network including mRNAs and lncRNAs that is called the competing endogenous RNA (ceRNA) network [26]. ceRNA-mediated regulatory mechanisms constitute an important pathway in lncRNA-modulated post-transcriptional regulation in TNBC [27]. A microarray-based ceRNA network analysis revealed that 4852 lncRNAs are related to the diagnosis and treatment outcome of TNBC [28]. Another study using the TCGA database found that 150 lncRNAs are expressed at the tissue level and 823 in serum and these lncRNAs could act as prognostic factors in TNBC [29]. Furthermore, the study found that the lncRNA OSTN-AS1 is a novel immune-related prognostic marker [29]. An integrated ceRNA network involving three miRNAs (CHRDL1, FCGR1A, and RSAD2) and two lncRNAs (HIF1A-AS2 and AK124454) was developed using microarray analysis [30]. These findings demonstrate that lncRNAs play major roles in the regulation of cell signalling, genetic heterogeneity, TNBC development, and pathological features (Figure 6) shown in Table 1.
3.1. Importance of lncRNAs in Tumour Invasiveness and Metastasis
Tumour invasion and metastasis explain the severity and mortality rate in patients with TNBC (Figure 6) [78,79]. GAS5 overexpression induces the expression of miR-196a-5p, which activates the FOXO1/PI3K/Akt signalling pathway [80]. TROJAN is a drug that reduces the metastasis burden. Degradation of TROJAN is regulated by ZMYND8, and the ubiquitin–proteasome pathway is involved in this process [81]. CCAT1 activates the migration of TNBC cells via miR-218/ZFX signalling [40]. Various ncRNAs are involved in cell migration and invasion via specific regulatory pathways, including MIR503HG through the miR-103/OLFM4 axis [60], CCAT1 through the dysregulation of the miR-218/ZFX axis [40], AFAP1-AS1 through the activation of Wnt/β-catenin signalling [82], miR-34a through the activation of EMT-associated signalling pathways [83], PAPAS through miR-34a.83 downregulation [52], sONE through sONE/NOS3/NO signalling activation [53], LINC-ZNF469-3 by activating the miR-574-5p/ZEB1 axis [71,78], ZEB2 through the activation of PI3K/Akt/GSK3β/ZEB2 signalling [45], PVT1 by regulating p21 and KLF5/β-catenin signalling [24], ARNILA by mimicking ceRNA for miR-204, AIRN by downregulating Wnt/β-catenin/mTOR/PI3K signalling [36], RMST by downregulating Wnt/β-catenin/mTOR/PI3K signalling [67], and MALAT1 by upregulating miR-129-5p and miR-1/Slug expression [84]. Furthermore, miR-448 and some other lncRNAs play very important roles in invasion and metastasis, including SKAI1BC, HULC, HOTAIR, SNHG12, SNAR, WT1-AS, LINC01096, DANCR, NEF, HIF1A-AS2, LncKLHDC7B, and ROR [30,31,32,38,48,55,58,59,60,61,62,63,64,65,66,67,68,69,85,86].
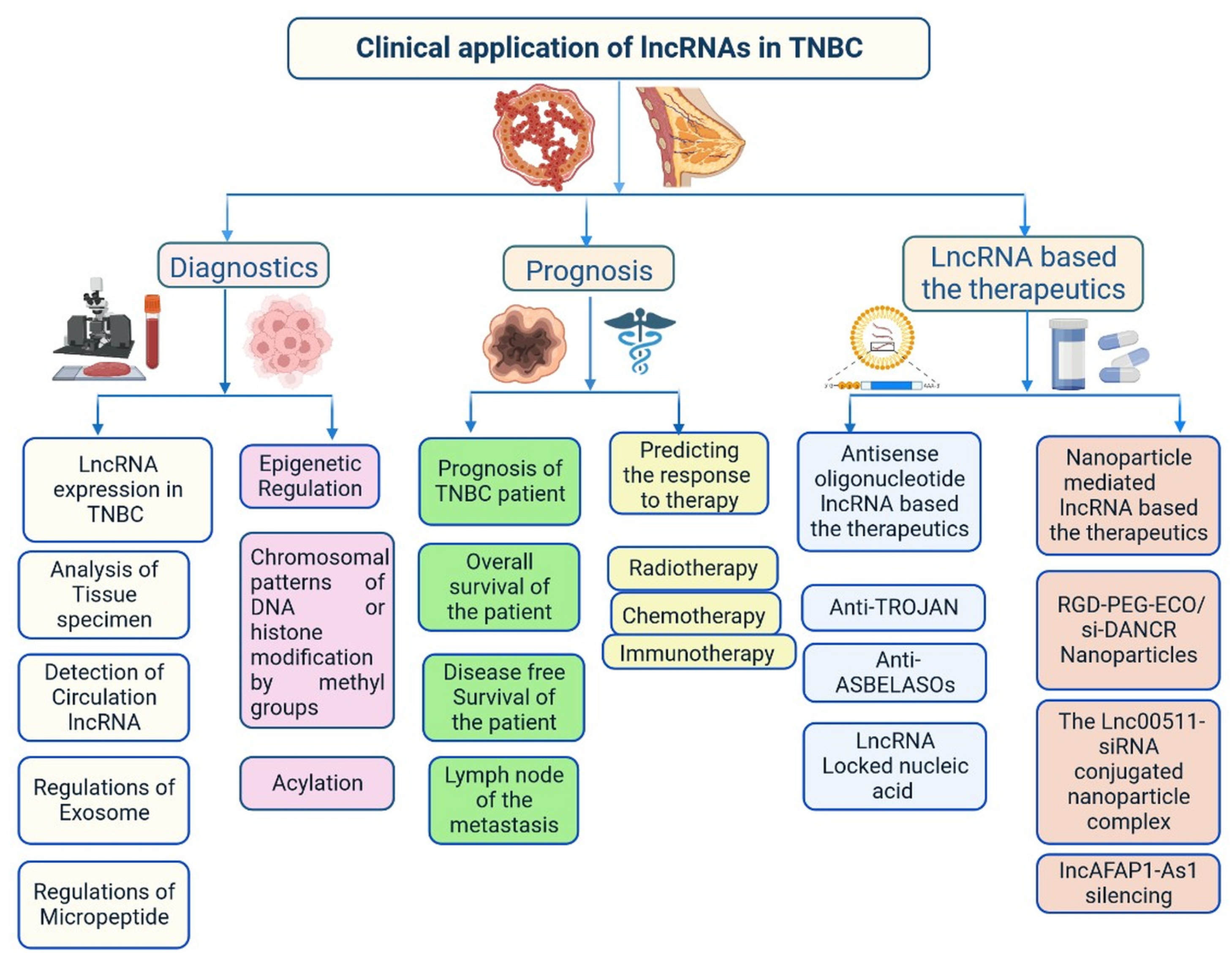
3.2. Importance of lncRNAs in Clinical Diagnosis
Several studies have found that lncRNAs are involved in the regulation of various transcription factors, epigenetic changes, chromatin remodelling, DNA methylation patterns, alternative splicing, post-translational modifications, and interaction with small peptides. All these events have great importance in the early diagnosis and treatment of patients with TNBC [14,86]. lncRNA expression levels in the blood and tissues of patients with TNBC at different stages has been investigated [14]. Based on reverse transcription quantitative PCR analysis data, the lncRNAs HIF1A-AS2, UCA1, and ANRIL can be used for TNBC detection, with areas under the curve in the range of 0.827–0.840, and a diagnostic accuracy of 0.962 for ANRIL [87]. ANRIL, SOX2OT, and ANRASSF1 are used to differentiate between healthy and TNBC cells. TINCR expression is used to differentiate various histological subtypes of BC, as it is highly expressed in TNBC cells [88]. UCA1 is associated with TNBC, acting as a specific marker for TNBC diagnosis. EZH2 is highly expressed in TNBC tissues and prevents apoptosis by activating the miR-4458/SOCS1 axis [89]. LINC00299 expression is increased in TNBC. Several lncRNAs bind to mRNAs, protecting them and increasing their stability. The oncogenic transcription factor SOX9 is activated by LINC02095 [90]. DANCR interacts with RXRA and activates PI3K/Akt signalling in TNBC [58]. LINC00152 enhances NEDD4-1-facilitated ubiquitination and dysregulation of PTEN protein in TNBC [91]. Cell cycle arrest at the G1 phase is induced by MIR100HG, with p27 binding to RNA–DNA; p27 is a cyclin-dependent kinase (CDK) inhibitor. Cell cycle arrest at the G0/G1 phase is induced by LINC00339 and RMST in TNBC through the miR-377-3p/HOXC6 signalling pathway [19,20,77,92]. GAS5 is actively involved in the inhibition of TNBC cells through its action on miR-196a-5p and miR-378a-5p/SUFU signalling [93]. Further understanding of the roles of all these lncRNAs in TNBC is needed to improve early diagnosis and clinical management of patients. Various genes are targeted by ncRNAs, including LARP7, CDKN1A, KLF2, TIA1, DDX3X, CDK, and QKI [94,95,96,97,98]. An analysis of the TCGA database showed that 1097 lncRNAs are expressed in BC, with 1510 differentially expressed lncRNAs in TNBC cells, 35 plasma lncRNAs in TNBC, and 672 in non-TNBC cells [14]. Some lncRNAs are directly linked to prognosis in TNBC, including FOXCUT, LINC00299, AP000924.1, AC091043.1, AL354793.1, AC010343.3, and FGF10-AS1 [14]. Plasma-specific lncRNAs are also used for diagnosis of TNBC, such as UCA1, ANRIL, and HIF1A-AS2 [30]. lncRNAs associated with lymph node metastasis, such as LINC000173, LINC00096, ZEB2-AS1, HIF1A-AS2, HULC, LUCAT1, SNHG12, MALAT1, HOTAIR, HIF1A-AS2, LINC00096, ADPGK-AS1, and ZEB2-AS1, have also shown importance in diagnosis and prognosis [11,14,30,49].
3.3. Importance of lncRNAs in Treatment
lncRNAs affect the response to treatments such as chemotherapy, immunotherapy, and radiotherapy [99]. H19 is expressed in patients with TNBC during neoadjuvant chemotherapy and is related to effective clinical outcomes. LINK-A expression is linked to response to pembrolizumab treatment in patients with TNBC because its decreased expression reduces CD8+ T-cell infiltration [59]. These lncRNAs act as biomarkers for treatment response in patients with TNBC. LncAFAP1-AS1 expression has been observed in patients with TNBC who received radiotherapy after surgery, and this lncRNA acts as biomarker for radiotherapy [82]. Moreover, lncRNAs are involved in angiogenesis. LINC01133 expression is induced by mesenchymal stem/stromal cells that adjoin TNBC cells [33]. lncRNAs are actively involved in the regulation of cell proliferation and apoptosis as well as drug resistance in TNBC [16,44,47,61,99]. DRHC and HOTAIR inhibit TNBC growth and development [31]. HOTAIR plays a role in the invasion and migration of TNBC cells and is used as a biomarker for TNBC metastasis in circulation and tissues, indicating poor survival and response [31,32]. DRHC inhibits TNBC cell proliferation by downregulating the expression of HOTAIR, whereas HOTAIR does not affect the expression level of DRHC. H19 expression is reduced in TNBC cells, whereas PTCSC3 expression is not altered by H19 overexpression [61]. HIST2H2BC and SNRPEP4 were identified in 165 frozen tissue samples by transcriptome microarrays; these lncRNAs are involved in taxane chemotherapy in patients with TNBC. Increased miR-377-3p expression delays TNBC progression by regulating the inc00339/miR-377-3p/HOXC6 axis and inhibits TNBC proliferation and apoptosis. Therefore, it is used as therapeutic target. HIF1A-AS2 expression is upregulated in TNBC mammary tissue, which is linked to overall survival. HOTAIR is closely associated with androgen receptor expression and used as a therapeutic strategy to prevent metastasis. The miR-199a/FOXP2 pathway is induced by LINC01133 and triggers the proliferation of TNBC cells. Various lncRNAs act as stem cell markers, such as DANCR, LINC01638, LINC-ZNF469-3, NEAT1, NRAD1, and ASRPS [75,87]. Some lncRNAs promote vasculogenic mimicry, providing growth supplementation for tumour formation in TNBC. TP73-AS1, which is activated by the miR-490-3p/TWIST1 pathway, is one example. LINK-A alters glycolysis by mediating HIF1α phosphorylation at Tyr565 and Ser7 [3,16,44,47]. MANCR inhibits DNA damage and prevents disease progression [66]. AWPPH is involved in the prevention of tumourigenesis upon treatment with carboplatin; AWPPH small interfering RNA (siRNA) silencing leads to increased chemosensitivity in TNBC [10,56]. TUG1 induces the expression of miR-197, reduces the activation of WNT signalling, and enhances TNBC cell sensitivity to cisplatin [75]. These findings demonstrate the importance of lncRNAs in the prevention of tumourigenesis. More studies are required to explore lncRNA treatment options. Early studies showed that HOTAIR recruits the polycomb repressive complex 2 to its target genes through the CoREST/REST H3K4 demethylase complex [75].
4. Nanoparticle-Based Targeted Therapy with ncRNAs for TNBC
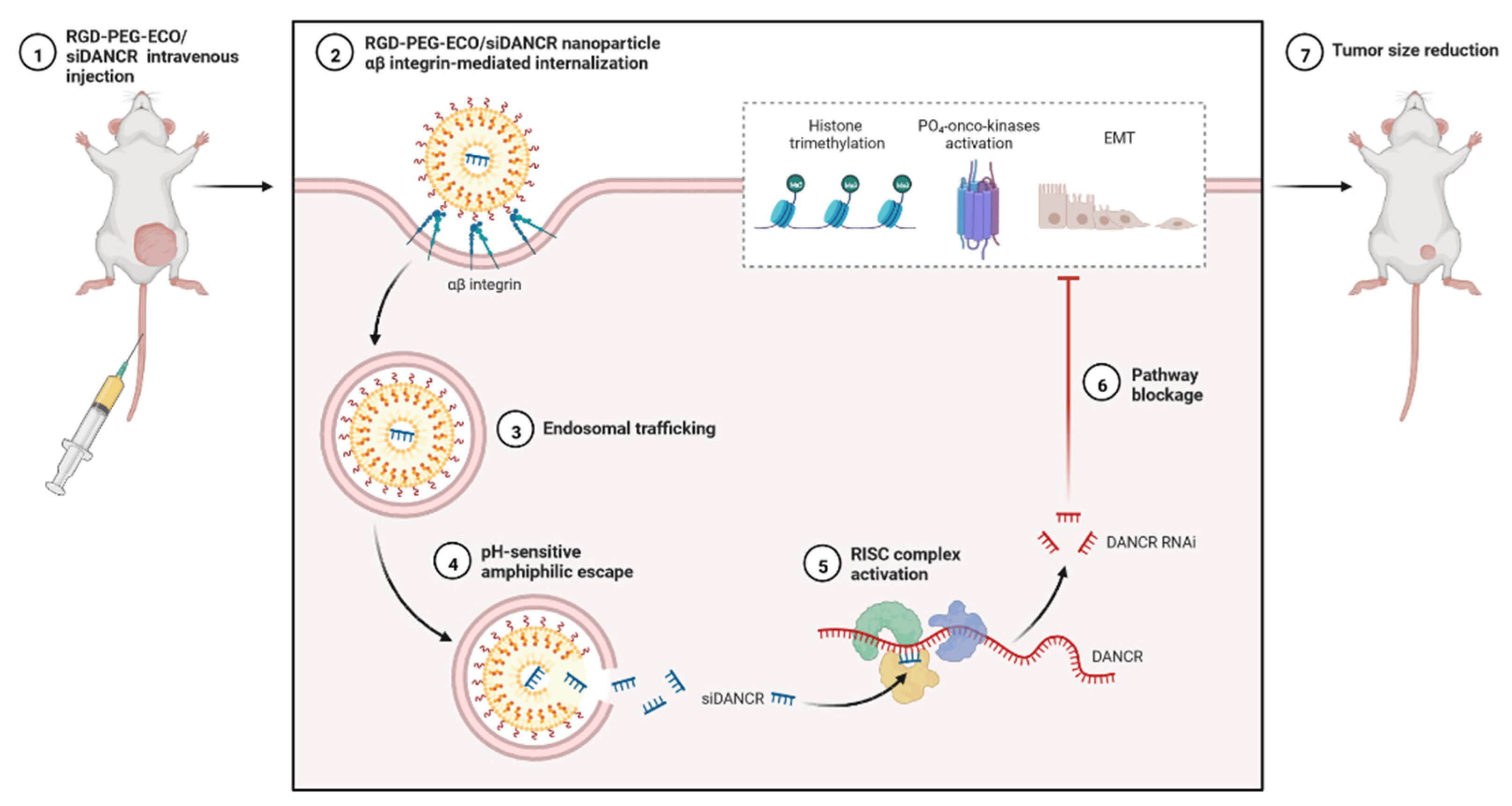
lncRNAs are versatile, able to exert multilevel gene regulation, and have emerged as therapeutic targets for clinically complicated TNBC cases (Figure 6) [25]. DANCR may potentially be used to reduce the limitations of monotherapy in TNBC networks and to lower the risk of side effects in healthy tissues [58]. DANCR is targeted by RNA interference (RNAi) (Figure 5 and Figure 6). The main challenge of RNAi therapy is targeted delivery; a non-viral siRNA-based delivery system has shown limited efficacy and temporary expression [94]. Amino acid-based lipid carriers have shown promising results in siRNA, nucleic acids, and CRISPR/Cas-based approaches [95]. DANCR overexpression was established in TNBC using RGD-PEG-ECO/siDANCR nanoparticles for effective cytosolic delivery of siDANCR [96]. Injection of the RGD-PEG-ECO/siDANCR nanoparticles led to reduce the progression of disease severity (Figure 7) [97]. In another study, LINC00511-siRNA was used to deliver siRNA in patients with TNBC [97]. Nanoengineered platforms were used to deliver lncAFAP1-AS1 siRNA (siAFAP1-AS1) to reverse radioresistance and increase the efficacy in TNBC tumour models [98]. These findings demonstrate that RNA nanoparticle-based targeted therapy can be more effective in TNBC. In conclusion, various lncRNAs are abnormally expressed and used in the treatment of TNBC, including ASOs, LNA, or RNA nanotechnology targeting lncRNAs.
5. LncRNAs Involved in The Regulation of Drug Resistance
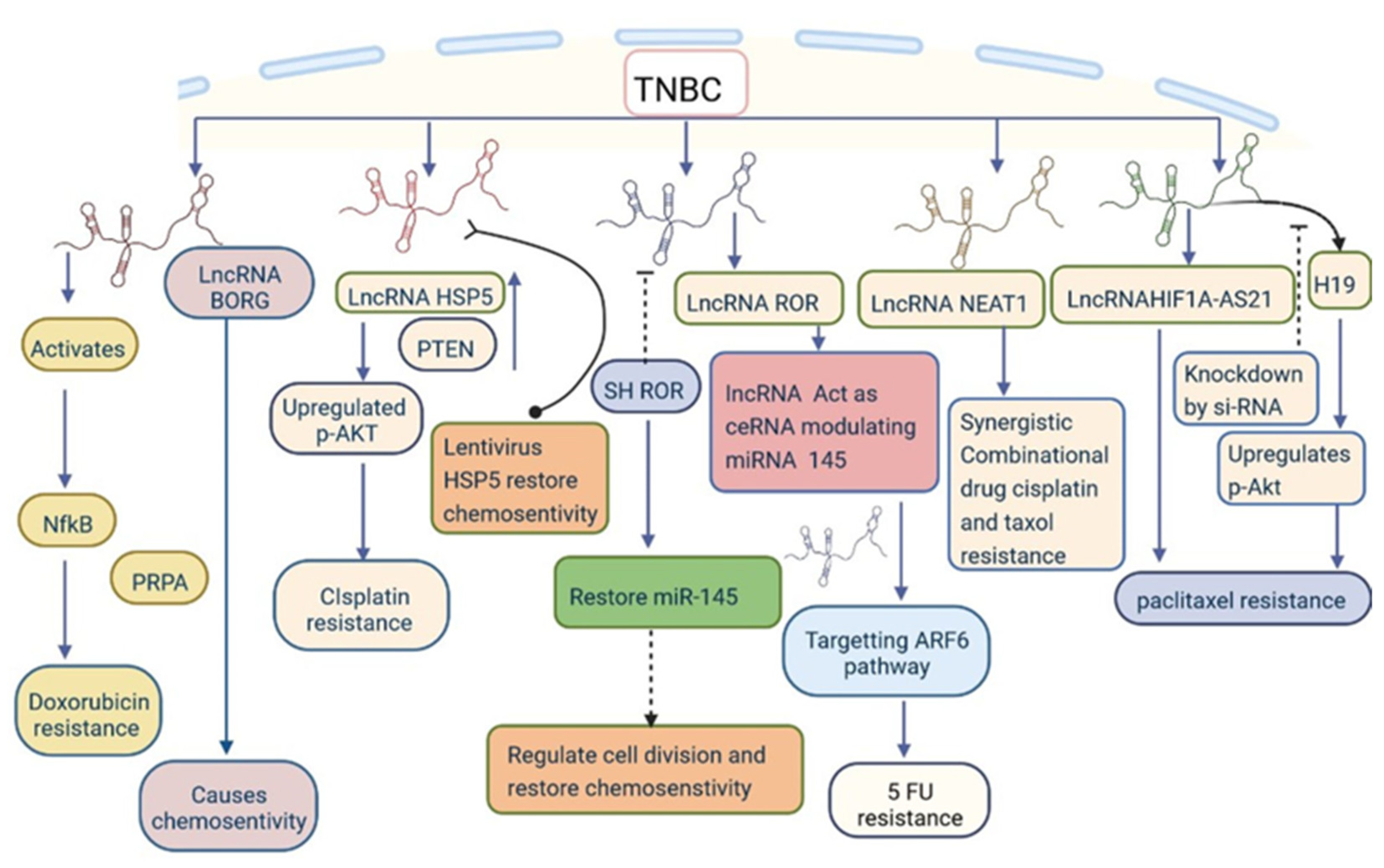
lncRNAs can alter the genetic regulation that may lead to the development of drug resistance. GAS5 promotes drug resistance to adriamycin, paclitaxel, and cisplatin [3,16,44,47]. Furthermore, GAS5 expression is reduced by mTORC1/mTORC2 (AZD8055) and PI3K/mTOR (BEZ235) inhibitors [99]. HOTAIR expression is controlled by EGFR/HER-2 inhibitor-based treatment such as lapatinib or the c-ABL inhibitor imatinib. Suppression of HOTAIR expression by the action of β-catenin on the HOTAIR promoter at the LEF1/TCF4-binding site increases drug resistance to combined therapy [100]. LINC01139 binding to the pleckstrin homology domain of AKT leads to hyperactivation and causes drug resistance to AKT inhibitors, which are commonly used for treatment of patients with TNBC [101]. Another treatment option for TNBC is immunotherapy-based treatments. LINK-A causes drug resistance by activating LINK-A–PKA–TRIM71 signalling, reducing the efficacy of immune checkpoint inhibitor-based treatments [102]. HIF1A-AS2 and AK12 4454 also cause drug resistance in TNBC cells [30]. H19 and NEAT1 induce resistance to paclitaxel through the AKT signalling pathway [16,61]. BORG causes resistance to doxorubicin through NF-κB signalling (Figure 8) [4,74]. DNA damage and repair is a complex process, and various signalling pathways are involved; impairment of this process leads to tumour development [103]. LINP1 participates in DNA double-strand break repair mechanisms using a scaffold linked up with Ku80 and DNA-PKcs, which prevents resistance to ionizing radiation [104]. INP1 prevents resistance to radiotherapy in TNBC [105]. The knockout of PCAT6 enhances the radiosensitivity of TNBC cells via the miR-185-5p/TPD52 axis [50]. These findings demonstrate the potential use of lncRNAs for the regulation of drug resistance in patients with TNBC (Table 2 and Figure 8). Further studies are required to investigate the roles of other lncRNAs in drug resistance.
6. Future Perspectives of lncRNAs as Potential Diagnosis and Pharmacological Tools/Targets
Different aspects of lncRNAs remain unclear, including their expression patterns in TNBC cells and their role in the modulation of mRNA coding genes [14,25]. Understanding their molecular heterogeneity would be helpful for developing novel drugs [113]. Single-stranded oligonucleotide antagonists targeting ASBEL have been designed to improve the half-life of the lncRNA in the serum. lncRNA expression is also regulated by tyrosine kinase receptors (TKRs) and non-TKRs through the simultaneous action of multiple genes; more studies are required to identify unknown mechanisms for the simultaneous targeting of multiple genes [114]. Moreover, studies focusing on molecular mechanisms are needed to improve our understanding of how FDA-approved chemotherapeutic agents for malignant neoplasms exert their regulatory action through epigenetic mechanisms on TNBC. The expression level of lncRNAs is dysregulated by chromosomes 1 and 10 via an unknown mechanism, which also needs to be explored [112]. The co-localization of lncRNAs plays a major role in TNBC progression and endocrine-based resistance therapy; we need to consider co-expressed lncRNAs to identify possible strategies for better diagnosis and treatment options [115]. Some lncRNAs are used as biomarkers; high-throughput analysis of lncRNAs by next generation sequencing in TNBC cells should be conducted using cell lines and animal models to identify abnormally expressed lncRNAs in TNBC. lncRNA replacement therapy could potentially be used to restore tumour-suppressive lncRNAs [115]. A CRISPR/Cas9 (Clustered Regularly Interspaced Short Palindromic Repeats/CRISPR-associated protein 9)-based approach can be used to reprogram transcription regulatory network and immune regulations of lncRNAs [116,117,118] The co-expression patterns of lncRNAs with transcription and translation need to be further explored to identify genetic heterogeneity pathophysiology mechanisms for early diagnosis, drug discovery, and understanding the treatment response and drug resistance mechanisms in patients with TNBC.
7. Conclusions
Clinical management of patients with TNBC is difficult owing to aggressive tumour behaviour and histological heterogeneity. The biological behaviour of TNBC cells, including genetic and epigenetic regulation, is not fully understood. We need to discover novel molecular biomarkers and therapeutic targets for better treatment efficacy. Personalized therapy for TNBC patients is required to reduce TNBC progression. lncRNAs may prove to be very useful, as they play important roles in TNBC development and response to follow-up treatment. Considering the challenges of in vivo experimental designs, lncRNAs continue to be promising as biomarkers and potential therapeutic targets. Most lncRNAs exhibit low sequence conservation, which may limit the scope of efficacy. For instance, most lncRNAs are not common in humans or mice; therefore, loss-of-function experiments in mice are not feasible and positive clinical outcomes using this approach have remained limited. Accordingly, with the significant advances in the genetic study of lncRNAs, novel methods should be explored for diagnosis, therapy, and prognosis, but the potential clinical applications of lncRNAs are significant. RNAi is the most widely applied and efficient technology for targeting lncRNAs. To use lncRNAs in therapeutic settings, however, different technologies must be investigated, and more study is required. Compared to esiRNAs (endonuclease-made siRNA), conventional siRNAs show more off-target effects. Short hairpin RNA (shRNA), which is produced inside of cells, is another type of RNAi. When compared to esiRNAs, shRNAs have a significantly greater off-target effects and produce a silent response that may be temporary or sustained. Plasmid vectors containing shRNA or siRNA are used to transfect tumour cells. Therefore, toxicity and off-target effects are other limitations of the lncRNA delivery system. To overcome the limitations of current diagnosis and treatment strategies, additional research is required for the use of lncRNAs as diagnostic biomarkers and therapeutic targets in TNBC.
Author Contributions
D.D.S.: Conceptualization, validation, methodology, formal analysis, writing—original draft; D.K.Y.: Conceptualization, validation, methodology, formal analysis, writing—original draft, investigation, software; H.-J.L.: funding. All authors have read and agreed to the published version of the manuscript.
Funding
The authors acknowledge the Cooperative Research Program for Agriculture Science and Technology Development (project no. PJ01701902) Rural Development Administration, Republic of Korea.
Acknowledgments
D.D.S. thanks the Amity Institute of Biotechnology, Amity University Rajasthan, Jaipur, India, and D.K.Y. thanks Gachon University, Republic of Korea, for providing the necessary computational and journal subscriptions for the needed literature search. The authors appreciate Biorender’s graphics assistance.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
Long non-coding RNA: lncRNA; Triple-negative breast cancer: TNBC; Breast cancer: BC; Luminal A: LA; Hormone receptor positive: HR+; Oestrogen receptor-positive: ER+; Human epidermal growth factor receptor 2: HER2; Human epidermal growth factor receptor 2 positive: HER2+; Epidermal growth factor receptor: EGFR; Vascular endothelial growth factor: VEGFR; Master regulator of cell cycle entry and proliferative metabolism:- c-Myc (MYC); Androgen receptor: AR; Cyclin E: CCNE; Mouse double minute 2 homolog: MDM2; Phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit alpha: PIK3CA; Membrane-associated guanylate kinase WW and PDZ domain containing I: MAGI; Serine/threonine kinase 3: AKT3; Myosin IIIA: MYO3A; Parkin RBR E3 ubiquitin protein ligase: PRKN; Inositol polyphosphate-4-phosphatase type II B: INPP4B; Phosphatase and tensin homolog: PTEN; Cyclin-dependent kinase inhibitor 2A: CDKN2A; Breast cancer gene 1: BRCA1; Breast cancer gene 2: BRCA2; Tumour protein p53: TP53; RB transcriptional corepressor 1: RB1; Myeloid/lymphoid or mixed-lineage leukaemia 3: MLL3; Ribonucleic acid: RNA; Deoxyribonucleic acid: DNA; Non-coding RNA: ncRNA; microRNAs: miRNAs; Ribosomal ribonucleic acid: rRNA; Transfer RNAs: tRNAs; Small interfering RNAs: siRNA; Small nuclear RNAs: snRNAs; Extracellular RNAs: exRNAs; Circular RNAs: circRNAs; Non-coding repressor of NFAT: NRON; Hepatocellular carcinoma up-regulated EZH2-associated long non-coding RNA: HEIH; HLA complex P5: HCP5; one of the most upregulated lncRNA: LINC00096; Nuclear paraspeckle assembly transcript 1: NEAT1; Associated with poor prognosis of hepatocellular carcinoma: AWPPH; Lung cancer-associated transcript 1: LUCAT1; Heart and neural crest derivatives expressed transcript 2 antisense RNA 1: HAND2-AS1; POU domain, class 3, transcription factor: POU3F3; Metastasis-associated lung adenocarcinoma transcript 1: MALAT1; Antisense non-coding RNA in the INK4 locus: ANRIL; Protein kinase B: AKT; Mitogen-activated protein kinases: MAPKs; Competing endogenous RNAs: ceRNA; Homo sapiens OSTN antisense RNA 1: OSTN-AS1; Competing endogenous RNA: ceRNA; Chordin-like 1: CHRDL1; High affinity immunoglobulin gamma Fc receptor IA: FCGR1A; Radical S-adenosyl methionine domain-containing 2: RSAD2; HIF1A antisense RNA 2: HIF1A-AS2; HOX antisense intergenic RNA: HOTAIR; Down-regulated in human cancers: DRHC; Long intergenic non-protein coding RNA 1133: LINC01133; Long intergenic non-protein coding RNA 1096: LINC01096; Long non-coding RNA high expression in hepatocellular carcinoma: lncRNA HEIH; Androgen receptor negatively regulated lncRNA: ARNILA; Long intergenic non-protein coding RNA02095: LINC02095; Wilms tumour protein antisense RNA: WT1-AS; Growth arrest specific 5: GAS5; Colon cancer-associated transcript 1: CCAT1; A small regulatory peptide of STAT3: ASRPS; Radical S-adenosyl methionine domain-containing protein 2: AND2-AS1; RNA-induced silencing complex: RISC; Lysine-specific demethylase 5B: KDM5B; Histone demethylase 1B: JARID1B; Trimethylation of lysine 4 on the histone H3 protein subunit: H3K4me3; Monomethylation of lysine 4 on the histone H3 protein subunit: H3K4me1; retinoblastoma protein: pRB; caveolin 1: CAV1; Homeobox protein Hox-A5: HOXA5; Stratifin: SFN; methyl group: CH3; Ras homolog gene family, member A: RhoA; Cyclin-dependent kinase inhibitor 1B: p27; Mir-100-Let-7a-2-Mir-125b-1 cluster host gene: MIR100HG; Triplex-forming oligonucleotides: TFO; Triplex-targeting ability: TTA; Plasmacytoma variant translocation 1: PVT1; Krüppel-like factors: KLF5; BRCA1-associated protein-1: BAP1; Basic helix–loop–helix: bHLH; Next-generation sequencing: NGS; Epidermal growth factor receptor family: ErbB; Phosphatidylinositol-3-kinase: PI3K; Mammalian target of rapamycin: mTOR; POU domain, class 3, transcription factor 3: POU3F3; Negative regulatory factor: NEF; Zinc finger E-box-binding homeobox 2 antisense RNA1: ZEB2 – AS1; Small nucleolar RNA host gene 12: SNHG12; Prostate cancer-associated transcript 6: CAT6; Hepatocellular carcinoma up-regulated long non-coding RNA: HULC; Pap fimbrial major pilin protein: papA; lncRNA human histocompatibility leukocyte antigen (HLA), complex P5: HCP5; non-coding RNA in the aldehyde dehydrogenase 1A pathway: NRAD1; Small NF90 (ILF3)-associated RNA I: SNAR-I; DNA and RNA binding protein: SON; Differentiation antagonizing non-protein coding RNA: DANCR; Long intergenic non-coding RNA for kinase activation: LINK-A; MIR503 host gene: MIR503HG; Long non-coding RNA (lncRNA) nuclear enriched abundant transcript 1: NEAT1; Papillary thyroid carcinoma susceptibility candidate 3: PTCSC3; Nicotinamide phosphoribosyl transferase: NAMPT; Mitotically associated long non-coding RNA: MANCR; Rhabdomyosarcoma 2-associated transcript: RMST; Small conductance Ca2+-activated K+ (SK) ATP binding cassette subfamily A member 1: SK AI1BC; Receptor tyrosine kinase-like orphan receptor: ROR; Antisense of IGF2R non-protein coding RNA: AIRN; Long intergenic non-protein coding zinc finger protein 469: LINC ZNF469; Programmed cell death 4 antisense RNA 1: PDCD4-AS1; Human ovarian cancer-specific transcript 2: HOST2; BMP/OP-responsive gene: BORG; H19 imprinted maternally expressed transcript: H19; lncRNA P73 antisense RNA 1: TP73-AS1; Taurine up-regulated 1: TUG1; Mir-100-Let-7a-2-Mir-125b-1 cluster host gene: MIR100HG; Homo sapiens zinc finger MYND-type containing 8: ZMYND8; Zinc finger protein x-linked: ZFX; Human olfactomedin 4: OLFM4; Actin filament-associated protein 1: AFAP1; Epithelial–mesenchymal transition: EMT; Studied nitric oxide synthase 3: NOS3; Glycogen synthase kinase-3 β: GSK3β; Krueppel-like factor 5/β: KLF5/β; Wingless-related integration site: Wnt; Rhabdomyosarcoma 2 associated transcript: RMST; Spindle and kinetochore associated complex subunit 1: SKA1; LIM homeobox: Lhx; Kelch domain containing 1: KLHDC1; Hypoxia inducible factor 1 subunit alpha: HIF1A; HIF1A antisense RNA 2: HIF1A-AS2; Urothelial cancer-associated 1: UCA1; Area under curve: AUC; SOX2 overlapping transcript: SOX2OT; SRY-box transcription factor 2: SOX2; Ras association domain family member 1: RASSF1; Enhancer of Zeste 2 polycomb repressive complex 2 subunit: EZH2; Suppressor of cytokine signalling 1: SOCS1; Adenosine diphosphate-dependent glucokinase antisense RNA 1: ADPGK-AS1; Retinoid X receptor: RXR; Neural precursor cell expressed developmentally down-regulated protein 4-1: NEDD4-1; Phosphatase and tensin homolog: PTEN; Cyclin-dependent kinases: CDKs; Homeobox C cluster 6: HOXC6; SUFU negative regulator of hedgehog signalling: SUFU; La ribonucleoprotein 7- Transcriptional Regulator: LARP7; Cyclin Dependent Kinase inhibitor 1A: CDKN1A; T-cell intracellular antigen 1:TIA1; DEAD-box helicase 3 X-Linked: DDX3X; QKI, KH domain containing RNA binding: QKI; Fibroblast growth factor 10: FGF10; FGF10 antisense RNA 1: FGF10-AS1; Neoadjuvant chemotherapy: NAC; Mesenchymal stem/stromal cells: MSCs; Papillary thyroid carcinoma susceptibility candidate 3: PTCSC3; Human putative histone H2B type 2-C: HIST2H2BC; Small nuclear ribonucleoprotein polypeptide E pseudogene 4: SNRPEP4; Overall survival: OS; Androgen receptor: AR; Twist family BHLH transcription factor 1: TWIST1; Hypoxia-inducible factor 1-alpha: HIF1α; Mitotically associated long non-coding RNA: MANCR; Associated with poor prognosis of hepatocellular carcinoma: AWPPH; Nemo-like kinase: NLK; Polycomb repressive complex 2: PRC2; Repressor element-1 silencing transcription factor: REST; Co-element-1 silencing transcription factor: CoREST; RNA interference: RNAi; Clustered regularly interspaced short palindromic repeats: CRISPR; CRISPR-associated protein 9: Cas 9; Ras association domain family member 1: RASSF1; Terminal differentiation–induced non-coding RNA: TINCR; Urothelial cancer-associated 1: UCA1; Polycomb repressive complex 2: PRC2; Antisense oligonucleotides: ASOs; Locked nucleic acid: LNA; Growth-stasis-specific transcript 5: GAS 5; Pan-class I PI3K and mTOR kinase inhibitor (Dactolisib): BEZ235; Lymphoid enhancer binding factor 1: LEF1; Transcription factor 4: TCF4; Tripartite motif-containing 71: RIM71; DNA double-strand break: DSB; Tyrosine kinase receptors: TKRs.
References
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast Cancer Cell Line Classification and Its Relevance with Breast Tumor Subtyping. J. Cancer 2017, 8, 3131–3141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onitilo, A.A.; Engel, J.M.; Greenlee, R.T.; Mukesh, B.N. Breasreastct Cancer Subtypes Based on ER/PR and Her2 Expression: Comparison of Clinicopathologic Features and Survival. Clin. Med. Res. 2009, 7, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Marra, A.; Trapani, D.; Viale, G.; Criscitiello, C.; Curigliano, G. Practical Classification of Triple-Negative Breast Cancer: Intratumoral Heterogeneity, Mechanisms of Drug Resistance, and Novel Therapies. NPJ Breast Cancer 2020, 6, 54. [Google Scholar] [CrossRef] [PubMed]
- Aine, M.; Boyaci, C.; Hartman, J.; Häkkinen, J.; Mitra, S.; Campos, A.B.; Nimeus, E.; Ehinger, A.; Vallon-Christersson, J.; Borg, Å.; et al. Molecular Analyses of Triple-Negative Breast Cancer in the Young and Elderly. Breast Cancer Res. 2021, 23, 20. [Google Scholar] [CrossRef] [PubMed]
- Zagami, P.; Carey, L.A. Triple negative breast cancer: Pitfalls and progress. NPJ Breast Cancer 2022, 8, 95. [Google Scholar] [CrossRef]
- Bissanum, R.; Chaichulee, S.; Kamolphiwong, R.; Navakanitworakul, R.; Kanokwiroon, K. Molecular Classification Models for Triple Negative Breast Cancer Subtype Using Machine Learning. J. Pers. Med. 2021, 11, 881. [Google Scholar] [CrossRef] [PubMed]
- Volovat, S.R.; Volovat, C.; Hordila, I.; Hordila, D.-A.; Mirestean, C.C.; Miron, O.T.; Lungulescu, C.; Scripcariu, D.V.; Stolniceanu, C.R.; Konsoulova-Kirova, A.A.; et al. MiRNA and LncRNA as Potential Biomarkers in Triple-Negative Breast Cancer: A Review. Front. Oncol. 2020, 10, 526850. [Google Scholar] [CrossRef]
- Lee, J.; Park, H.Y.; Kim, W.W.; Lee, S.J.; Jeong, J.H.; Kang, S.H.; Jung, J.H.; Chae, Y.S. Biological function of long noncoding RNA snaR in HER2-positive breast cancer cells. Tumour Biol. 2017, 39, 1010428317707374. [Google Scholar] [CrossRef] [Green Version]
- Melone, V.; Salvati, A.; Brusco, N.; Alexandrova, E.; D’Agostino, Y.; Palumbo, D.; Palo, L.; Terenzi, I.; Nassa, G.; Rizzo, F.; et al. Functional Relationships between Long Non-Coding RNAs and Estrogen Receptor Alpha: A New Frontier in Hormone-Responsive Breast Cancer Management. Int. J. Mol. Sci. 2023, 24, 1145. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, Q.; Xing, W.; Wang, W. Role of MicroRNA/LncRNA Intertwined with the Wnt/β-Catenin Axis in Regulating the Pathogenesis of Triple-Negative Breast Cancer. Front. Pharmacol. 2022, 13, 814971. [Google Scholar] [CrossRef]
- Mou, E.; Wang, H. LncRNA LUCAT1 Facilitates Tumorigenesis and Metastasis of Triple-Negative Breast Cancer through Modulating MiR-5702. Biosci. Rep. 2019, 39, BSR20190489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, L.; Xiao, X.; Zhao, Y.; Yin, L.; Fu, M.; Zhang, X.; Jiang, P. The Functional Roles of Long Noncoding RNA DANCR in Human Cancers. J. Cancer 2020, 11, 6970–6981. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Liu, L.; Wang, Z. Long Non-coding RNA Heart and Neural Crest Derivatives Expressed 2-antisense RNA 1 Overexpression Inhibits the Proliferation of Cancer Cells by Reducing RUNX2 Expression in Triple-negative Breast Cancer. Oncol. Lett. 2019, 18, 6775–6780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhang, N.; Liu, Y.; Su, P.; Liang, Y.; Li, Y.; Wang, X.; Chen, T.; Song, X.; Sang, Y.; et al. Epigenetic Regulation of NAMPT by NAMPT-AS Drives Metastatic Progression in Triple-Negative Breast Cancer. Cancer Res. 2019, 79, 3347–3359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di, S.; Bai, R.; Lu, D.; Chen, C.; Ma, T.; Zou, Z.; Zhang, Z. Long Non-Coding RNA MAFG-AS1 Promotes Proliferation and Metastasis of Breast Cancer by Modulating STC2 Pathway. Cell Death Discov. 2022, 8, 249. [Google Scholar] [CrossRef]
- Du, T.; Shi, Y.; Xu, S.; Wan, X.; Sun, H.; Liu, B. Long Non-Coding RNAs in Drug Resistance of Breast Cancer. OncoTargets Ther. 2020, 13, 7075–7087. [Google Scholar] [CrossRef]
- Mei, J.; Hao, L.; Wang, H.; Xu, R.; Liu, Y.; Zhu, Y.; Liu, C. Systematic Characterization of Non-coding RNAs in Triple-negative Breast Cancer. Cell Prolif. 2020, 53, e12801. [Google Scholar] [CrossRef] [Green Version]
- Richard, J.L.C.; Eichhorn, P.J.A. Deciphering the Roles of LncRNAs in Breast Development and Disease. Oncotarget 2018, 9, 20179–20212. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Ke, H.; Zhang, H.; Ma, Y.; Ao, L.; Zou, L.; Yang, Q.; Zhu, H.; Nie, J.; Wu, C.; et al. LncRNA MIR100HG Promotes Cell Proliferation in Triple-Negative Breast Cancer through Triplex Formation with P27 Loci. Cell Death Discov. 2018, 9, 805. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wang, Z.; Yu, S.; Liu, D.; Sun, L. LncmiRHG-MIR100HG: A New Budding Star in Cancer. Front. Oncol. 2022, 12, 997532. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, N.; Song, P.; Fu, Y.; Ren, Y.; Li, Z.; Wang, J. LncRNA GATA3-AS1 Facilitates Tumour Progression and Immune Escape in Triple-negative Breast Cancer through Destabilization of GATA3 but Stabilization of PD-L1. Cell Prolif. 2020, 53, e12855. [Google Scholar] [CrossRef]
- Qi, F.; Qin, W.; Zang, Y. Molecular Mechanism of Triple-negative Breast Cancer-associated BRCA1 and the Identification of Signaling Pathways. Oncol. Lett. 2019, 17, 2905–2914. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Tan, Y.; Wang, H.; Xu, M.; Xu, L. Long Non-Coding RNA Plasmacytoma Variant Translocation 1 (PVT1) Enhances Proliferation, Migration, and Epithelial-Mesenchymal Transition (EMT) of Pituitary Adenoma Cells by Activating β-Catenin, c-Myc, and Cyclin D1 Expression. Med. Sci. Monit. 2019, 25, 7652–7659. [Google Scholar] [CrossRef]
- Tang, J.; Li, Y.; Sang, Y.; Yu, B.; Lv, D.; Zhang, W.; Feng, H. LncRNA PVT1 Regulates Triple-Negative Breast Cancer through KLF5/Beta-Catenin Signaling. Oncogene 2018, 37, 4723–4734. [Google Scholar] [CrossRef]
- Zhang, W.; Guan, X.; Tang, J. The Long Non-coding RNA Landscape in Triple-negative Breast Cancer. Cell Prolif. 2021, 54, e12966. [Google Scholar] [CrossRef]
- López-Urrutia, E.; Bustamante Montes, L.P.; Ladrón de Guevara Cervantes, D.; Pérez-Plasencia, C.; Campos-Parra, A.D. Crosstalk between Long Non-Coding RNAs, Micro-RNAs and MRNAs: Deciphering Molecular Mechanisms of Master Regulators in Cancer. Front. Oncol. 2019, 9, 669. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Song, G.; Li, M.; Hao, X.; Huang, Y.; Lan, J.; Yang, S.; Zhang, Z.; Zhang, G.; Mu, J. Construction and Comprehensive Analysis of a CeRNA Network to Reveal Potential Novel Biomarkers for Triple-Negative Breast Cancer. Cancer Manag. Res. 2020, 12, 7061–7075. [Google Scholar] [CrossRef] [PubMed]
- Le, K.; Guo, H.; Zhang, Q.; Huang, X.; Xu, M.; Huang, Z.; Yi, P. Gene and LncRNA Co-Expression Network Analysis Reveals Novel CeRNA Network for Triple-Negative Breast Cancer. Sci. Rep. 2019, 9, 15122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Mi, M.; Li, X.; Zheng, X.; Wu, G.; Zhang, L. LncRNA OSTN-AS1 May Represent a Novel Immune-Related Prognostic Marker for Triple-Negative Breast Cancer Based on Integrated Analysis of a CeRNA Network. Front. Genet. 2019, 10, 850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, G.; Han, J. HIF1A-AS2 Predicts Poor Prognosis and Regulates Cell Migration and Invasion in Triple-negative Breast Cancer. J. Cell. Biochem. 2019, 120, 10513–10518. [Google Scholar] [CrossRef]
- Zhao, W.; Geng, D.; Li, S.; Chen, Z.; Sun, M. LncRNA HOTAIR Influences Cell Growth, Migration, Invasion, and Apoptosis via the MiR-20a-5p/HMGA2 Axis in Breast Cancer. Cancer Med. 2018, 7, 842–855. [Google Scholar] [CrossRef]
- Yu, F.; Wang, L.; Zhang, B. Long Non-coding RNA DRHC Inhibits the Proliferation of Cancer Cells in Triple Negative Breast Cancer by Downregulating Long Non-coding RNA HOTAIR. Oncol. Lett. 2019, 18, 3817–3822. [Google Scholar] [CrossRef] [Green Version]
- Tu, Z.; Schmöllerl, J.; Cuiffo, B.G.; Karnoub, A.E. Microenvironmental Regulation of Long Noncoding RNA LINC01133 Promotes Cancer Stem Cell-Like Phenotypic Traits in Triple-Negative Breast Cancers. Stem Cells 2019, 37, 1281–1292. [Google Scholar] [CrossRef]
- Zhao, M.; Zhang, M.; Tao, Z.; Cao, J.; Wang, L.; Hu, X. MiR-331-3p Suppresses Cell Proliferation in TNBC Cells by Downregulating NRP2. Technol. Cancer Res. Treat. 2020, 19, 153303382090582. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Zhou, B.; Lv, Y.; Qian, Q. LncRNA HEIH Regulates Cell Proliferation and Apoptosis through MiR-4458/SOCS1 Axis in Triple-Negative Breast Cancer. Human Cell 2019, 32, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Shen, Y.; Zhang, W.; Jin, J.; Huang, D.; Fang, H.; Ji, W.; Shi, Y.; Tang, L.; Chen, W.; et al. An Androgen Receptor Negatively Induced Long Non-Coding RNA ARNILA Binding to MiR-204 Promotes the Invasion and Metastasis of Triple-Negative Breast Cancer. Cell Death Differ. 2018, 25, 2209–2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tariq, A.; Hao, Q.; Sun, Q.; Singh, D.K.; Jadaliha, M.; Zhang, Y.; Chetlangia, N.; Ma, J.; Holton, S.E.; Bhargava, R.; et al. LncRNA-Mediated Regulation of SOX9 Expression in Basal Subtype Breast Cancer Cells. RNA 2020, 26, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xi, C.; Yang, X.; Lu, X.; Yu, K.; Zhang, Y.; Gao, R. LncRNA WT1-AS Inhibits Triple-Negative Breast Cancer Cell Migration and Invasion by Downregulating Transforming Growth Factor Β1. Cancer Biother. Radiopharm. 2019, 34, 671–675. [Google Scholar] [CrossRef]
- Filippova, E.A.; Fridman, M.V.; Burdennyy, A.M.; Loginov, V.I.; Pronina, I.V.; Lukina, S.S.; Dmitriev, A.A.; Braga, E.A. Long Noncoding RNA GAS5 in Breast Cancer: Epigenetic Mechanisms and Biological Functions. Int. J. Mol. Sci. 2021, 22, 6810. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Li, X.; Fan, Q.; Liu, G.; Yin, J. CCAT1 Promotes Triple-Negative Breast Cancer Progression by Suppressing MiR-218/ZFX Signaling. Aging 2019, 11, 4858–4875. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, S.; Zhu, X.; Zhang, L.; Deng, J.; Li, F.; Guo, B.; Zhang, S.; Wu, R.; Zhang, Z.; et al. LncRNA-Encoded Polypeptide ASRPS Inhibits Triple-Negative Breast Cancer Angiogenesis. J. Exp. Med. 2020, 217, e20190950. [Google Scholar] [CrossRef]
- Tang, M.; Liu, Y.; Zhang, Q.; Zhang, P.; Wu, J.; Wang, J.; Ruan, Y.; Huang, Y. Antitumor Efficacy of the Runx2-dendritic Cell Vaccine in Triple-negative Breast Cancer In Vitro. Oncol. Lett. 2018, 16, 2813–2822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Meng, X.; Yu, Y.; Pan, L.; Zheng, Q.; Lin, W. LncRNA POU3F3 Promotes Proliferation and Inhibits Apoptosis of Cancer Cells in Triple-Negative Breast Cancer by Inactivating Caspase 9. Biosci. Biotechnol. Biochem. 2019, 83, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wu, X.; Liang, Y.; Ye, G.; Che, Y.; Wu, X.; Zhu, X.; Fan, H.; Fan, X.; Xu, J. MiR-155 Increases Stemness and Decitabine Resistance in Triple-negative Breast Cancer Cells by Inhibiting TSPAN5. Mol. Carcinog. 2020, 59, 447–461. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Deng, L.; Wang, Y.-D. Roles and Mechanisms of Long Non-Coding RNAs in Breast Cancer. Int. J. Mol. Sci. 2023, 24, 89. [Google Scholar] [CrossRef]
- Tian, Y.; Xia, S.; Ma, M.; Zuo, Y. LINC00096 Promotes the Proliferation and Invasion by Sponging MiR-383-5p and Regulating RBM3 Expression in Triple-Negative Breast Cancer. OncoTargets Ther. 2019, 12, 10569–10578. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Zhao, W.; Zhang, H.; Chu, Z.; Liu, H.; Fang, X.; Tang, D. Long Non-Coding RNA ANRIL Promotes Chemoresistance in Triple-Negative Breast Cancer via Enhancing Aerobic Glycolysis. Life Sci. 2022, 306, 120810. [Google Scholar] [CrossRef]
- Tamang, S.; Acharya, V.; Roy, D.; Sharma, R.; Aryaa, A.; Sharma, U.; Khandelwal, A.; Prakash, H.; Vasquez, K.M.; Jain, A. SNHG12: An LncRNA as a Potential Therapeutic Target and Biomarker for Human Cancer. Front. Oncol. 2019, 9, 901. [Google Scholar] [CrossRef]
- Xia, L.; Wang, H. LncRNA LUCAT1/ELAVL1/LIN28B/SOX2 Positive Feedback Loop Promotes Cell Stemness in Triple-Negative Breast Cancer. Breast J. 2022, 2022, 1–12. [Google Scholar] [CrossRef]
- Shi, R.; Wu, P.; Liu, M.; Chen, B.; Cong, L. Knockdown of LncRNA PCAT6 Enhances Radiosensitivity in Triple-Negative Breast Cancer Cells by Regulating MiR-185-5p/TPD52 Axis. OncoTargets Ther. 2020, 13, 3025–3037. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.-X.; Chen, S.; Huang, L.; Shao, Z.-M. Predictive and Prognostic Value of Matrix Metalloproteinase (MMP)-9 in Neoadjuvant Chemotherapy for Triple-Negative Breast Cancer Patients. BMC Cancer 2018, 18, 909. [Google Scholar] [CrossRef]
- Tokumaru, Y.; Katsuta, E.; Oshi, M.; Sporn, J.C.; Yan, L.; Le, L.; Matsuhashi, N.; Futamura, M.; Akao, Y.; Yoshida, K.; et al. High Expression of miR-34a Associated with Less Aggressive Cancer Biology but Not with Survival in Breast Cancer. Int. J. Mol. Sci. 2020, 21, 3045. [Google Scholar] [CrossRef]
- Wang, L.; Luan, T.; Zhou, S.; Lin, J.; Yang, Y.; Liu, W.; Tong, X.; Jiang, W. LncRNA HCP5 Promotes Triple Negative Breast Cancer Progression as a CeRNA to Regulate BIRC3 by Sponging MiR-219a-5p. Cancer Med. 2019, 8, 4389–4403. [Google Scholar] [CrossRef]
- Han, C.; Fu, Y.; Zeng, N.; Yin, J.; Li, Q. LncRNA FAM83H-AS1 Promotes Triple-Negative Breast Cancer Progression by Regulating the MiR-136-5p/Metadherin Axis. Aging 2020, 12, 3594–3616. [Google Scholar] [CrossRef]
- Niu, L.; Fan, Q.; Yan, M.; Wang, L. LncRNA NRON Down-Regulates LncRNA SnaR and Inhibits Cancer Cell Proliferation in TNBC. Biosci. Rep. 2019, 39, BSR20190468. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Qu, H.; Gong, W.; Xiang, J.; Yang, M.; Zhang, W. LncRNA AWPPH and MiRNA-21 Regulates Cancer Cell Proliferation and Chemosensitivity in Triple-negative Breast Cancer by Interacting with Each Other. J. Cell. Biochem. 2019, 120, 14860–14866. [Google Scholar] [CrossRef]
- Youness, R.A.; Hafez, H.M.; Khallaf, E.; Assal, R.A.; Abdel Motaal, A.; Gad, M.Z. The Long Noncoding RNA SONE Represses Triple-negative Breast Cancer Aggressiveness through Inducing the Expression of MiR-34a, MiR-15a, MiR-16, and Let-7a. J. Cell. Physiol. 2019, 234, 20286–20297. [Google Scholar] [CrossRef]
- Jin, S.-J.; Jin, M.-Z.; Xia, B.-R.; Jin, W.-L. Long Non-Coding RNA DANCR as an Emerging Therapeutic Target in Human Cancers. Front. Oncol. 2019, 9, 1225. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Li, C.; Xing, Z.; Hu, Q.; Liang, K.; Han, L.; Wang, C.; Hawke, D.H.; Wang, S.; Zhang, Y.; et al. The LINK-A LncRNA Activates Normoxic HIF1α Signalling in Triple-Negative Breast Cancer. Nat. Cell. Biol. 2016, 18, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Dong, G.; Shi, H.; Zhang, J.; Ning, Z.; Bao, X.; Liu, C.; Hu, J.; Liu, M.; Xiong, B. LncRNA MIR503HG Inhibits Cell Migration and Invasion via MiR-103/OLFM4 Axis in Triple Negative Breast Cancer. J. Cell. Mol. Med. 2019, 23, 4738–4745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kansara, S.; Pandey, V.; Lobie, P.E.; Sethi, G.; Garg, M.; Pandey, A.K. Mechanistic Involvement of Long Non-Coding RNAs in Oncotherapeutics Resistance in Triple-Negative Breast Cancer. Cells 2020, 9, 1511. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Hou, M.; Zhan, Y.; Sheng, X. LncRNA PTCSC3 Inhibits Triple-negative Breast Cancer Cell Proliferation by Downregulating LncRNA H19. J. Cell. Biochem. 2019, 120, 15083–15088. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Xiong, Z.; Li, R.; Liang, H.; Jia, C.; Deng, M. Long Non-Coding RNA NRON Is Downregulated in HCC and Suppresses Tumour Cell Proliferation and Metastasis. Biomed. Pharmacother. 2018, 104, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Goh, C.Y.; Wyse, C.; Ho, M.; O’Beirne, E.; Howard, J.; Lindsay, S.; Kelly, P.; Higgins, M.; McCann, A. Exosomes in Triple Negative Breast Cancer: Garbage Disposals or Trojan Horses? Cancer Lett. 2020, 473, 90–97. [Google Scholar] [CrossRef]
- Tracy, K.M.; Tye, C.E.; Ghule, P.N.; Malaby, H.L.H.; Stumpff, J.; Stein, J.L.; Stein, G.S.; Lian, J.B. Mitotically-Associated LncRNA (MANCR) Affects Genomic Stability and Cell Division in Aggressive Breast Cancer. Mol. Cancer Res. 2018, 16, 587–598. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Liu, D.; Wu, X.; Zeng, Y.; Li, L.; Hou, Y.; Li, W.; Liu, Z. Long Non-coding RNA (LncRNA) RMST in Triple-negative Breast Cancer (TNBC): Expression Analysis and Biological Roles Research. J. Cell. Physiol. 2018, 233, 6603–6612. [Google Scholar] [CrossRef]
- Ferraro, D.A.; Patella, F.; Zanivan, S.; Donato, C.; Aceto, N.; Giannotta, M.; Dejana, E.; Diepenbruck, M.; Christofori, G.; Buess, M. Endothelial Cell-Derived Nidogen-1 Inhibits Migration of SK-BR-3 Breast Cancer Cells. BMC Cancer 2019, 19, 312. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Kim, Y.-J.; Ji, Z.; Kang, J.M.; Wirianto, M.; Paudel, K.R.; Smith, J.A.; Ono, K.; Kim, J.-A.; Eckel-Mahan, K.; et al. ROR Activation by Nobiletin Enhances Antitumor Efficacy via Suppression of IκB/NF-ΚB Signaling in Triple-Negative Breast Cancer. Cell Death Dis. 2022, 13, 374. [Google Scholar] [CrossRef]
- Merikhian, P.; Eisavand, M.R.; Farahmand, L. Triple-Negative Breast Cancer: Understanding Wnt Signaling in Drug Resistance. Cancer Cell Int. 2021, 21, 419. [Google Scholar] [CrossRef]
- Wang, P.-S.; Chou, C.-H.; Lin, C.-H.; Yao, Y.-C.; Cheng, H.-C.; Li, H.-Y.; Chuang, Y.-C.; Yang, C.-N.; Ger, L.-P.; Chen, Y.-C.; et al. A Novel Long Non-Coding RNA Linc-ZNF469-3 Promotes Lung Metastasis through MiR-574-5p-ZEB1 Axis in Triple Negative Breast Cancer. Oncogene 2018, 37, 4662–4678. [Google Scholar] [CrossRef]
- Wang, D.; Wang, Z.; Zhang, L.; Sun, S. LncRNA PDCD4-AS1 Alleviates Triple Negative Breast Cancer by Increasing Expression of IQGAP2 via MiR-10b-5p. Transl. Oncol. 2021, 14, 100958. [Google Scholar] [CrossRef] [PubMed]
- Hua, K.; Deng, X.; Hu, J.; Ji, C.; Yu, Y.; Li, J.; Wang, X.; Fang, L. Long Noncoding RNA HOST2, Working as a Competitive Endogenous RNA, Promotes STAT3-Mediated Cell Proliferation and Migration via Decoying of Let-7b in Triple-Negative Breast Cancer. J. Exp. Clin. Cancer Res. 2020, 39, 58. [Google Scholar] [CrossRef] [PubMed]
- Jahangiri, L.; Ishola, T. Dormancy in Breast Cancer, the Role of Autophagy, lncRNAs, miRNAs and Exosomes. Int. J. Mol. Sci. 2022, 23, 5271. [Google Scholar] [CrossRef]
- Tao, W.; Sun, W.; Zhu, H.; Zhang, J. Knockdown of Long Non-Coding RNA TP73-AS1 Suppresses Triple Negative Breast Cancer Cell Vasculogenic Mimicry by Targeting MiR-490-3p/TWIST1 Axis. Biochem. Biophys. Res. Commun. 2018, 504, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Sun, L.; Wan, F. Molecular Mechanisms of TUG1 in the Proliferation, Apoptosis, Migration and Invasion of Cancer Cells (Review). Oncol. Lett. 2019, 18, 4393–4402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.-Y.; Zhou, Z.-Y.; Zhang, K.-J.; Pang, J.; Wang, S.-M. Long Non-Coding RNA MIR100HG Promotes the Migration, Invasion and Proliferation of Triple-Negative Breast Cancer Cells by Targeting the MiR-5590-3p/OTX1 Axis. Cancer Cell Int. 2020, 20, 508. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Tang, H.; Ling, L.; Li, N.; Jia, X.; Zhang, Z.; Wang, X.; Shi, L.; Yin, J.; Qiu, N.; et al. LINC01638 LncRNA Activates MTDH-Twist1 Signaling by Preventing SPOP-Mediated c-Myc Degradation in Triple-Negative Breast Cancer. Oncogene 2018, 37, 6166–6179. [Google Scholar] [CrossRef]
- Al-Mahmood, S.; Sapiezynski, J.; Garbuzenko, O.B.; Minko, T. Metastatic and Triple-Negative Breast Cancer: Challenges and Treatment Options. Drug Deliv. Transl. Res. 2018, 8, 1483–1507. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhou, J.; Wang, Z.; Wang, P.; Gao, X.; Wang, Y. Long Noncoding RNA GAS5 Suppresses Triple Negative Breast Cancer Progression through Inhibition of Proliferation and Invasion by Competitively Binding MiR-196a-5p. Biomed. Pharmacother. 2018, 104, 451–457. [Google Scholar] [CrossRef]
- Jin, X.; Xu, X.-E.; Jiang, Y.-Z.; Liu, Y.-R.; Sun, W.; Guo, Y.-J.; Ren, Y.-X.; Zuo, W.-J.; Hu, X.; Huang, S.-L.; et al. The Endogenous Retrovirus-Derived Long Noncoding RNA TROJAN Promotes Triple-Negative Breast Cancer Progression via ZMYND8 Degradation. Sci. Adv. 2019, 5, eaat9820. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Liu, P.; Tang, H.; Xie, X.; Kong, Y.; Song, C.; Qiu, X.; Xiao, X. AFAP1-AS1 Promotes Epithelial-Mesenchymal Transition and Tumorigenesis Through Wnt/β-Catenin Signaling Pathway in Triple-Negative Breast Cancer. Front. Pharmacol. 2018, 9, 1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imani, S.; Wei, C.; Cheng, J.; Khan, A.; Fu, S.; Yang, L.; Tania, M.; Zhang, X.; Xiao, X.; Zhang, X.; et al. MicroRNA-34a Targets Epithelial to Mesenchymal Transition-Inducing Transcription Factors (EMT-TFs) and Inhibits Breast Cancer Cell Migration and Invasion. Oncotarget 2017, 8, 21362–21379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, Y.; Geng, C.; Dong, Q. LncRNA PAPAS May Promote Triple-Negative Breast Cancer by Downregulating MiR-34a. J. Int. Med. Res. 2019, 47, 3709–3718. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Li, H.; Sun, R.; Li, P.; Yang, Z.; Liu, Y.; Wang, Z.; Yang, Y.; Yin, C. Long Non-coding RNA ZEB2-AS1 Promotes the Proliferation, Metastasis and Epithelial Mesenchymal Transition in Triple-negative Breast Cancer by Epigenetically Activating ZEB2. J. Cell. Mol. Med. 2019, 23, 3271–3279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, Y.; Li, Y.; Zhou, Z.; Ma, M.; Fu, K. Long Non-Coding RNA MALAT1 Promotes Proliferation and Invasion via Targeting MiR-129-5p in Triple-Negative Breast Cancer. Biomed. Pharmacother. 2017, 95, 922–928. [Google Scholar] [CrossRef] [PubMed]
- Mofed, D.; Omran, J.I.; Sabet, S.; Baiomy, A.A.; Emara, M.; Salem, T.Z. The Regulatory Role of Long Non-Coding RNAs as a Novel Controller of Immune Response against Cancer Cells. Mol. Biol. Rep. 2022, 49, 11775–11793. [Google Scholar] [CrossRef]
- Zuo, K.; Yuan, X.; Liang, X.; Sun, X.; Liu, S.; Connell, P.P.; Li, X.; Yang, W. QRT-PCR-Based DNA Homologous Recombination-Associated 4-Gene Score Predicts Pathologic Complete Response to Platinum-Based Neoadjuvant Chemotherapy in Triple-Negative Breast Cancer. Breast Cancer Res. Treat. 2022, 191, 335–344. [Google Scholar] [CrossRef]
- Zhang, W.; Yang, S.; Chen, D.; Yuwen, D.; Zhang, J.; Wei, X.; Han, X.; Guan, X. SOX2-OT Induced by PAI-1 Promotes Triple-Negative Breast Cancer Cells Metastasis by Sponging MiR-942-5p and Activating PI3K/Akt Signaling. Cell. Mol. Life Sci. 2022, 79, 59. [Google Scholar] [CrossRef]
- Zhou, Y.; Meng, X.; Chen, S.; Li, W.; Li, D.; Singer, R.; Gu, W. IMP1 Regulates UCA1-Mediated Cell Invasion through Facilitating UCA1 Decay and Decreasing the Sponge Effect of UCA1 for MiR-122-5p. Breast Cancer Res. 2018, 20, 32. [Google Scholar] [CrossRef]
- Manoochehri, M.; Jones, M.; Tomczyk, K.; Fletcher, O.; Schoemaker, M.J.; Swerdlow, A.J.; Borhani, N.; Hamann, U. DNA Methylation of the Long Intergenic Noncoding RNA 299 Gene in Triple-Negative Breast Cancer: Results from a Prospective Study. Sci. Rep. 2020, 10, 11762. [Google Scholar] [CrossRef]
- Zheng, S.; Li, M.; Miao, K.; Xu, H. LncRNA GAS5-promoted Apoptosis in Triple-negative Breast Cancer by Targeting MiR-378a-5p/SUFU Signaling. J. Cell. Biochem. 2020, 121, 2225–2235. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Zhu, Q.; Lu, H.; Chang, A.; Gao, C.; Zhou, Q.; Luo, K. HEXIM1 Controls P-TEFb Processing and Regulates Drug Sensitivity in Triple-Negative Breast Cancer. Mol. Biol. Cell 2020, 31, 1867–1878. [Google Scholar] [CrossRef] [PubMed]
- Aranza-Martínez, A.; Sánchez-Pérez, J.; Brito-Elias, L.; López-Camarillo, C.; Cantú de León, D.; Pérez-Plasencia, C.; López-Urrutia, E. Non-Coding RNAs Associated With Radioresistance in Triple-Negative Breast Cancer. Front. Oncol. 2021, 11, 752270. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Mangala, L.S.; Rodriguez-Aguayo, C.; Kong, X.; Lopez-Berestein, G.; Sood, A.K. RNA Interference-Based Therapy and Its Delivery Systems. Cancer Metastasis. Rev. 2018, 37, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Hattab, D.; Bakhtiar, A. Bioengineered SiRNA-Based Nanoplatforms Targeting Molecular Signaling Pathways for the Treatment of Triple Negative Breast Cancer: Preclinical and Clinical Advancements. Pharmaceutics 2020, 12, 929. [Google Scholar] [CrossRef]
- Vaidya, A.M.; Sun, Z.; Ayat, N.; Schilb, A.; Liu, X.; Jiang, H.; Sun, D.; Scheidt, J.; Qian, V.; He, S.; et al. Systemic Delivery of Tumor-Targeting SiRNA Nanoparticles against an Oncogenic LncRNA Facilitates Effective Triple-Negative Breast Cancer Therapy. Bioconjugate Chem. 2019, 30, 907–919. [Google Scholar] [CrossRef]
- Wu, B.; Yuan, Y.; Han, X.; Wang, Q.; Shang, H.; Liang, X.; Jing, H.; Cheng, W. Structure of LINC00511-siRNA-conjugated Nanobubbles and Improvement of Cisplatin Sensitivity on Triple Negative Breast Cancer. FASEB J. 2020, 34, 9713–9726. [Google Scholar] [CrossRef]
- Bi, Z.; Li, Q.; Dinglin, X.; Xu, Y.; You, K.; Hong, H.; Hu, Q.; Zhang, W.; Li, C.; Tan, Y.; et al. Nanoparticles (NPs)-Meditated LncRNA AFAP1-AS1 Silencing to Block Wnt/β -Catenin Signaling Pathway for Synergistic Reversal of Radioresistance and Effective Cancer Radiotherapy. Adv. Sci. 2020, 7, 2000915. [Google Scholar] [CrossRef]
- Jordan, N.J.; Dutkowski, C.M.; Barrow, D.; Mottram, H.J.; Hutcheson, I.R.; Nicholson, R.I.; Guichard, S.M.; Gee, J.M. Impact of Dual MTORC1/2 MTOR Kinase Inhibitor AZD8055 on Acquired Endocrine Resistance in Breast Cancer in Vitro. Breast Cancer Res. 2014, 16, R12. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-L.; Overstreet, A.-M.; Chen, M.-S.; Wang, J.; Zhao, H.-J.; Ho, P.-C.; Smith, M.; Wang, S.-C. Combined Inhibition of EGFR and C-ABL Suppresses the Growth of Triple-Negative Breast Cancer Growth through Inhibition of HOTAIR. Oncotarget 2015, 6, 11150–11161. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.; Hu, Q.; Li, C.; Xing, Z.; Ma, G.; Wang, C.; Li, J.; Ye, Y.; Yao, J.; Liang, K.; et al. The LINK-A LncRNA Interacts with PtdIns(3,4,5)P3 to Hyperactivate AKT and Confer Resistance to AKT Inhibitors. Nat. Cell Biol. 2017, 19, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Ye, Y.; Chan, L.-C.; Li, Y.; Liang, K.; Lin, A.; Egranov, S.D.; Zhang, Y.; Xia, W.; Gong, J.; et al. Oncogenic LncRNA Downregulates Cancer Cell Antigen Presentation and Intrinsic Tumor Suppression. Nat. Immunol. 2019, 20, 835–851. [Google Scholar] [CrossRef] [PubMed]
- Shin, V.Y.; Chen, J.; Cheuk, I.W.-Y.; Siu, M.-T.; Ho, C.-W.; Wang, X.; Jin, H.; Kwong, A. Long Non-Coding RNA NEAT1 Confers Oncogenic Role in Triple-Negative Breast Cancer through Modulating Chemoresistance and Cancer Stemness. Cell Death Dis. 2019, 10, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gooding, A.J.; Parker, K.A.; Valadkhan, S.; Schiemann, W.P. The LncRNA BORG: A Novel Inducer of TNBC Metastasis, Chemoresistance, and Disease Recurrence. J. Cancer Metastasis Treat. 2019, 5, 41. [Google Scholar] [CrossRef]
- Zhang, Y.; He, Q.; Hu, Z.; Feng, Y.; Fan, L.; Tang, Z.; Yuan, J.; Shan, W.; Li, C.; Hu, X.; et al. Long Noncoding RNA LINP1 Regulates Repair of DNA Double-Strand Breaks in Triple-Negative Breast Cancer. Nat. Struct. Mol. Biol. 2016, 23, 522–530. [Google Scholar] [CrossRef]
- Eades, G.; Wolfson, B.; Zhang, Y.; Li, Q.; Yao, Y.; Zhou, Q. LincRNA-RoR and MiR-145 Regulate Invasion in Triple-Negative Breast Cancer via Targeting ARF6. Mol. Cancer Res. 2015, 13, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Mitobe, Y.; Ikeda, K.; Suzuki, T.; Takagi, K.; Kawabata, H.; Horie-Inoue, K.; Inoue, S. ESR1-Stabilizing Long Noncoding RNA TMPO-AS1 Promotes Hormone-Refractory Breast Cancer Progression. Mol. Cell. Biol. 2019, 39, e00261-19. [Google Scholar] [CrossRef]
- Tang, L.; Chen, Y.; Chen, H.; Jiang, P.; Yan, L.; Mo, D.; Tang, X.; Yan, F. DCST1-AS1 Promotes TGF-β-Induced Epithelial–Mesenchymal Transition and Enhances Chemoresistance in Triple-Negative Breast Cancer Cells via ANXA1. Front. Oncol. 2020, 10, 280. [Google Scholar] [CrossRef]
- Wang, X.; Li, S.; Xiao, H.; Deng, X. Serum LncRNA TINCR Serve as a Novel Biomarker for Predicting the Prognosis in Triple-Negative Breast Cancer. Technol. Cancer Res. Treat. 2020, 19, 153303382096557. [Google Scholar] [CrossRef]
- Wu, M.; Wen, L.; Zhou, Y.; Wu, W. Role of LncRNA AGAP2-AS1 in Breast Cancer Cell Resistance to Apoptosis by the Regulation of MTA1 Promoter Activity. Technol. Cancer Res. Treat. 2022, 21, 153303382210853. [Google Scholar] [CrossRef]
- Hsu, M.-Y.; Hsieh, C.-H.; Huang, Y.-T.; Chu, S.-Y.; Chen, C.-M.; Lee, W.-J.; Liu, S.-J. Enhanced Paclitaxel Efficacy to Suppress Triple-Negative Breast Cancer Progression Using Metronomic Chemotherapy with a Controlled Release System of Electrospun Poly-d-l-Lactide-Co-Glycolide (PLGA) Nanofibers. Cancers 2021, 13, 3350. [Google Scholar] [CrossRef]
- Narui, K.; Ishikawa, T.; Shimizu, D.; Yamada, A.; Tanabe, M.; Sasaki, T.; Oba, M.S.; Morita, S.; Nawata, S.; Kida, K.; et al. Anthracycline Could Be Essential for Triple-Negative Breast Cancer: A Randomised Phase II Study by the Kanagawa Breast Oncology Group (KBOG) 1101. Breast 2019, 47, 1–9. [Google Scholar] [CrossRef]
- Xu, J.; Wu, K.; Jia, Q.; Ding, X. Roles of MiRNA and IncRNA in Triple-Negative Breast Cancer. J. Zhejiang Univ. Sci. B 2020, 21, 673–689. [Google Scholar] [CrossRef]
- Pucci, C.; Martinelli, C.; Ciofani, G. Innovative Approaches for Cancer Treatment: Current Perspectives and New Challenges. Ecancermedicalscience 2019, 13, 961. [Google Scholar] [CrossRef]
- Huang, L.; Liang, G.; Zhang, Q.; Zhao, W. The Role of Long Noncoding RNAs in Antiestrogen Resistance in Breast Cancer: An Overview and Update. J. Breast Cancer 2020, 23, 129. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.D.; Verma, R.; Tripathi, S.K.; Sahu, R.; Trivedi, P.; Yadav, D.K. Breast Cancer Transcriptional Regulatory Network Reprogramming by using the CRISPR/Cas9 System: An Oncogenetics Perspective. Curr. Top Med. Chem. 2021, 21, 2800–2813. [Google Scholar] [CrossRef]
- Singh, D.D.; Hawkins, R.D.; Lahesmaa, R.; Tripathi, S.K. CRISPR/Cas9 guided genome and epigenome engineering and its therapeutic applications in immune mediated diseases. Semin. Cell Dev. Biol. 2019, 96, 32–43. [Google Scholar] [CrossRef]
- Chen, B.; Dragomir, M.P.; Yang, C.; Li, Q.; Horst, D.; Calin, G.A. Targeting Non-Coding RNAs to Overcome Cancer Therapy Resistance. Sig. Transduct. Target. Ther. 2022, 7, 121. [Google Scholar] [CrossRef]
Figure 1.
Main intrinsic or molecular subtypes of breast cancer. There are five main intrinsic or molecular subtypes of BC such as Luminal A (~40%) [HR+ (ER+ and/or PR+), HER2-], Normal-like (~2–8%) [HR+ (ER+ and/or PR+), HER2-], Luminal B (~20%) [HR+ (ER+ and/or PR+), HER2+/−], HER2-enriched (~10–15%) [HR− (ER−, PR−), HER2+], and Triple-negative (~15–20%) [HR- (ER−, PR−) HER2].
Figure 1.
Main intrinsic or molecular subtypes of breast cancer. There are five main intrinsic or molecular subtypes of BC such as Luminal A (~40%) [HR+ (ER+ and/or PR+), HER2-], Normal-like (~2–8%) [HR+ (ER+ and/or PR+), HER2-], Luminal B (~20%) [HR+ (ER+ and/or PR+), HER2+/−], HER2-enriched (~10–15%) [HR− (ER−, PR−), HER2+], and Triple-negative (~15–20%) [HR- (ER−, PR−) HER2].
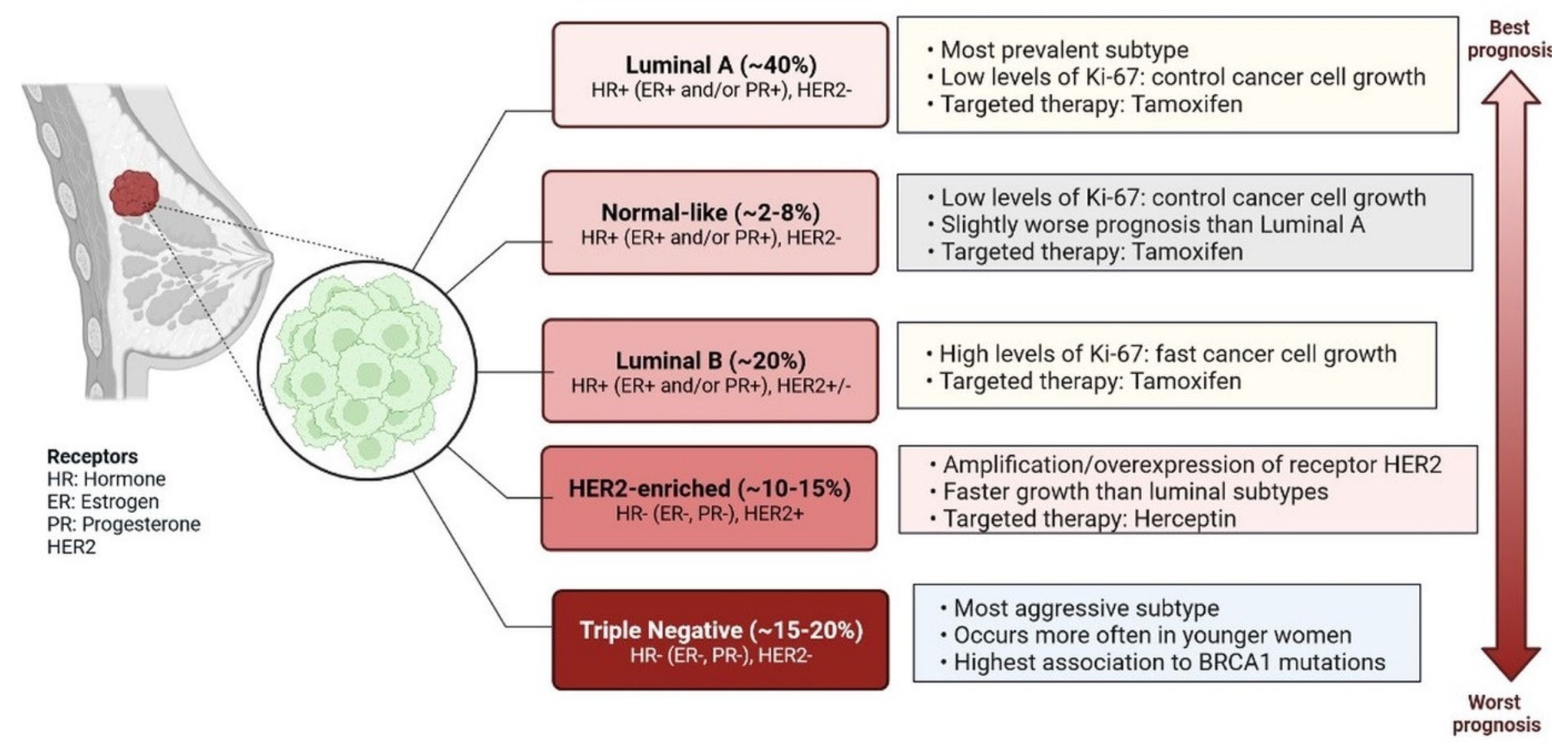
Figure 2.
Regulation of genetic markers associated with TNBC. Genetic markers associated with gain of function: EGFR, EGFR1/2, VEGFR, MYC, AR, CCNE, MDM2, PIK3CA, MAGI-AKT3, MYO3A, and PARK2. Genetic markers associated with Loss of function: INPP4B, PTEN, CDKN2A, BRCA1/2, TP53, RB1, and MLL3.
Figure 2.
Regulation of genetic markers associated with TNBC. Genetic markers associated with gain of function: EGFR, EGFR1/2, VEGFR, MYC, AR, CCNE, MDM2, PIK3CA, MAGI-AKT3, MYO3A, and PARK2. Genetic markers associated with Loss of function: INPP4B, PTEN, CDKN2A, BRCA1/2, TP53, RB1, and MLL3.

Figure 3.
Epigenetic deregulation in cancer including chromatin remodelling, DNA methylation, and non-coding RNA regulation that alters transcription and translation of oncogenic gene targets.
Figure 3.
Epigenetic deregulation in cancer including chromatin remodelling, DNA methylation, and non-coding RNA regulation that alters transcription and translation of oncogenic gene targets.
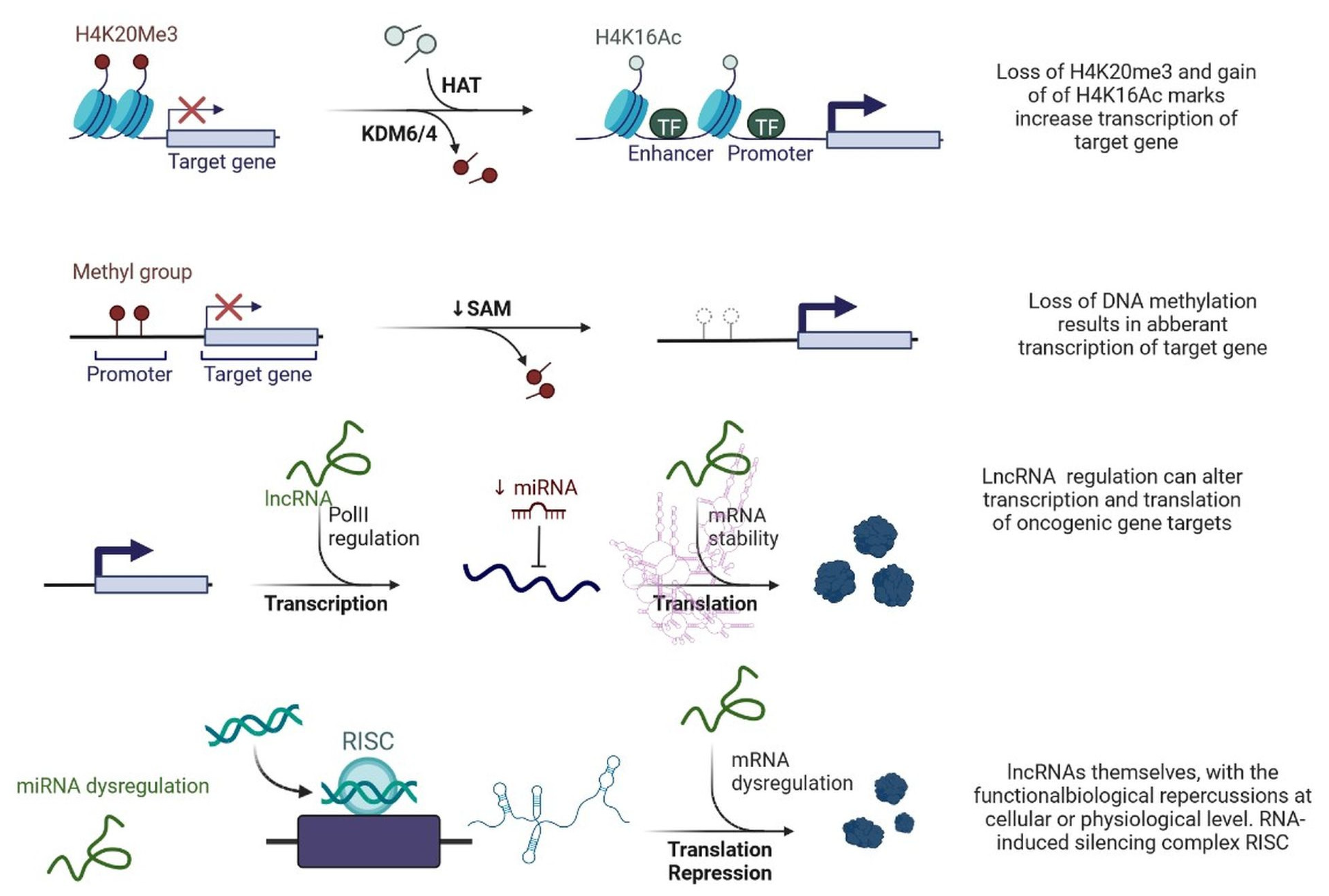
Figure 4.
Normal expression of lncRNA and effect of lncRNA expression changes on tumour behaviour depends on canonical function of mRNA target gene.
Figure 4.
Normal expression of lncRNA and effect of lncRNA expression changes on tumour behaviour depends on canonical function of mRNA target gene.
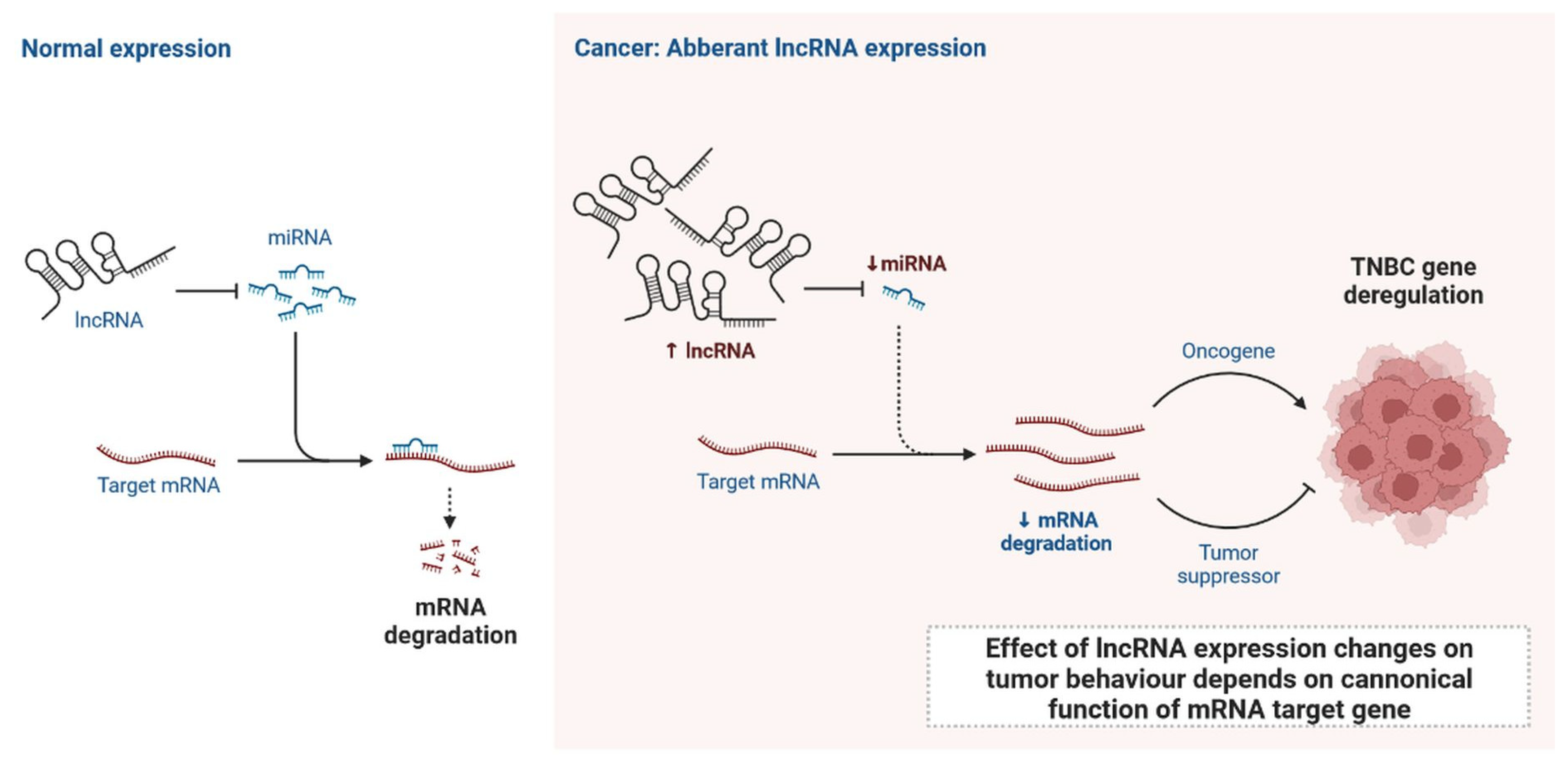
Figure 5.
lncRNAs are involved with the functional repercussions at the cellular and physiological level. RNA-induced silencing complex (RISC): KDM5B (lysine-specific demethylase 5B also known as histone demethylase JARID1B), H3K4me3 (trimethylation of lysine 4 on the histone H3 protein subunit), H3K4me1 (monomethylation of lysine 4 on the histone H3 protein subunit), hsa-miR-448 (also known miRNA448), BRCA1/2 (breast cancer 1/2), pRB (retinoblastoma protein), CAV 1 (caveolin 1), HOXA5 (Homeobox protein Hox-A5), SFN (Stratifin), CH3 (methyl group), and RhoA (Ras homolog gene family, member A).
Figure 5.
lncRNAs are involved with the functional repercussions at the cellular and physiological level. RNA-induced silencing complex (RISC): KDM5B (lysine-specific demethylase 5B also known as histone demethylase JARID1B), H3K4me3 (trimethylation of lysine 4 on the histone H3 protein subunit), H3K4me1 (monomethylation of lysine 4 on the histone H3 protein subunit), hsa-miR-448 (also known miRNA448), BRCA1/2 (breast cancer 1/2), pRB (retinoblastoma protein), CAV 1 (caveolin 1), HOXA5 (Homeobox protein Hox-A5), SFN (Stratifin), CH3 (methyl group), and RhoA (Ras homolog gene family, member A).
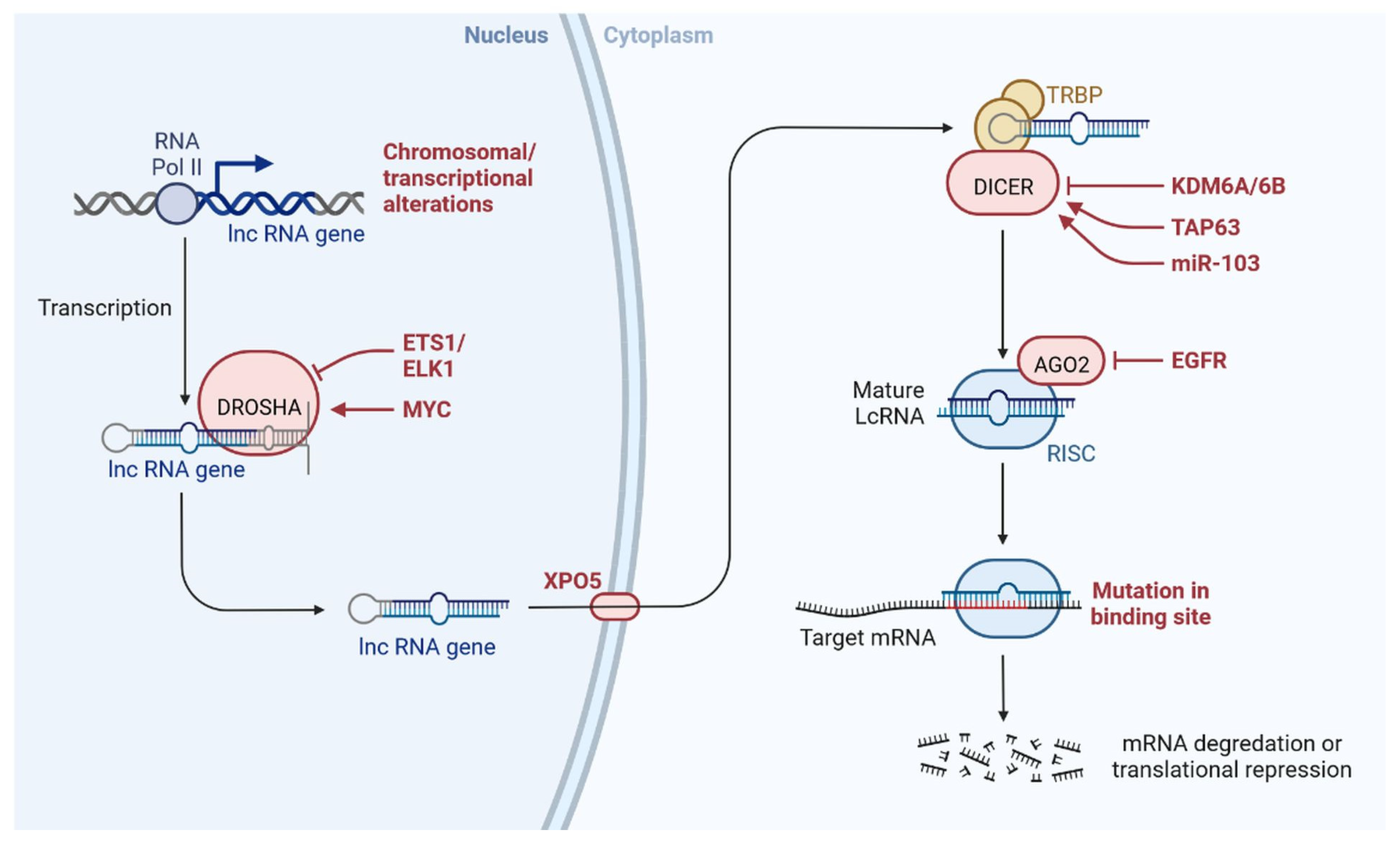
Figure 6.
Clinical importance of lncRNA in triple-negative breast cancer.
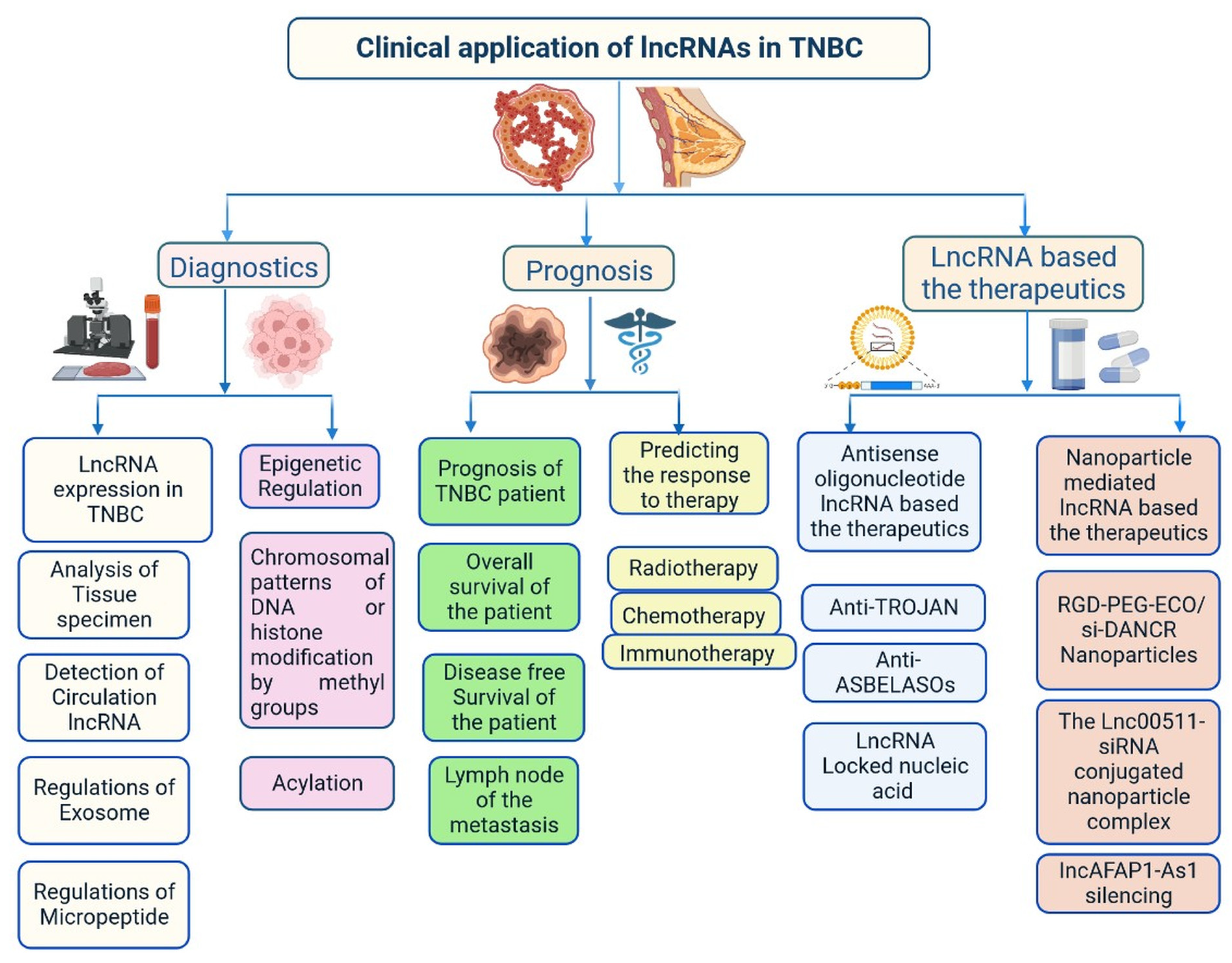
Figure 7.
Expression analysis of RGD-PEG-ECO/siDANCR nanoparticles in an animal model of TNBC.
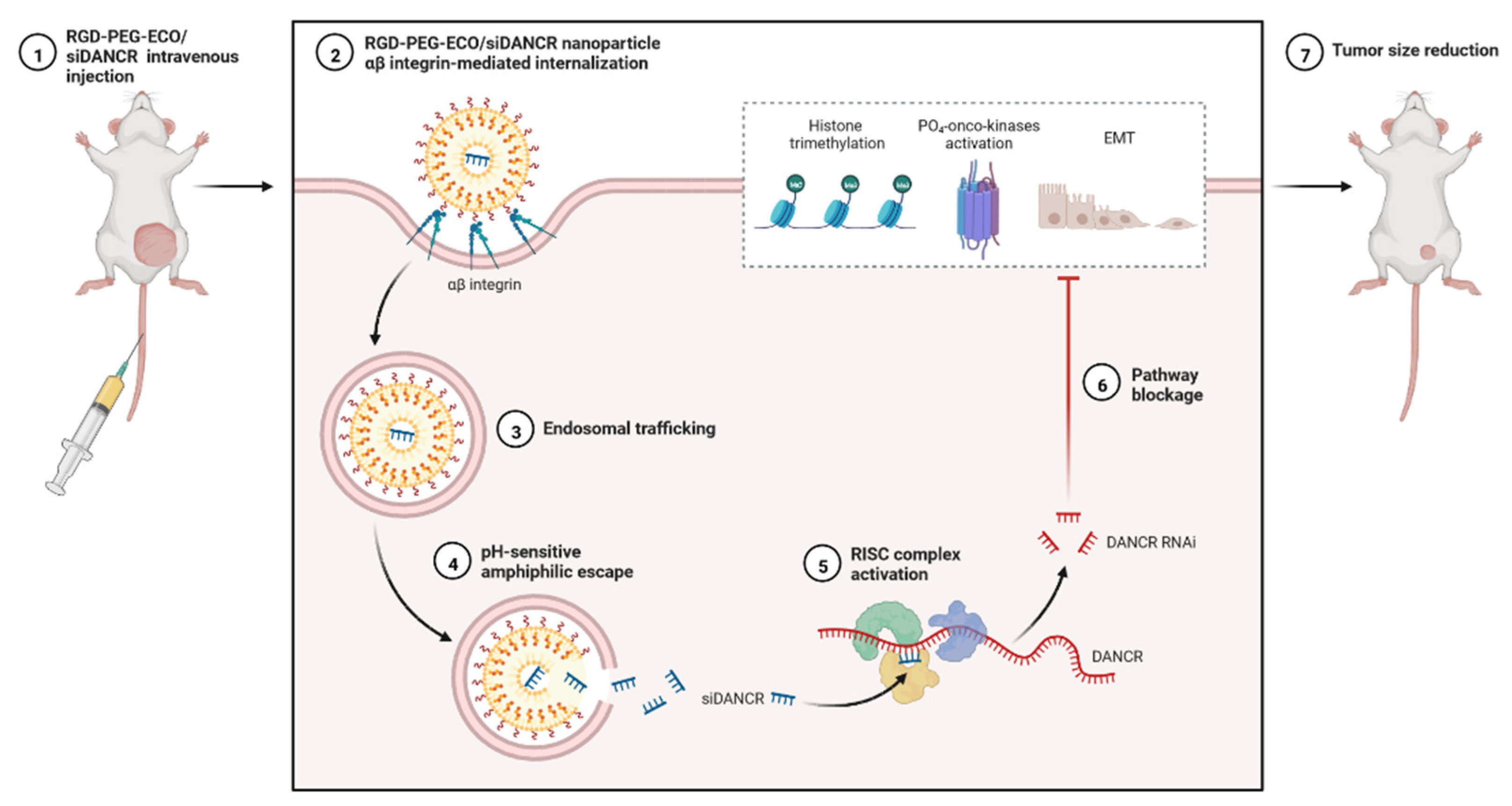
Figure 8.
lncRNAs are involved in drug resistance through various mechanisms.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Important lncRNAs associated with triple-negative breast cancer.
| S. N. | lncRNAs | Regulation of Expression | Clinical Importance | Potential Targets | Reference |
|---|---|---|---|---|---|
| 1 | HOTAIR | Upregulation | Increase cell invasion and migration | LEF1/TCF4 | [31] |
| 2 | DRHC | Downregulation | Reduce cell proliferation | HOTAIR | [32] |
| 3 | LINC01133 | Upregulation | Promote phenotypic features like cell stem cells (CSCs) | KLF4 | [33] |
| 4 | LINC01096 | Upregulation | Encourage cell invasion | miR-3130-3p | [34] |
| 5 | HEIH | Upregulation | Increase cell proliferation and prevent cell death | miR-4458/SOCS1 | [35] |
| 6 | ARNILA | Downregulation | Invasion and metastasis | miR-204/SOX4 | [36] |
| 7 | LINC02095 | Upregulation | Promote cell proliferation | SOX9 | [37] |
| 8 | WT1-AS | Downregulation | Inhibit cell migration and invasion | TGF-β1 | [38] |
| 9 | GAS5 | Downregulation | Promote cell apoptosis | miR-378a-5p/SUFU | [39] |
| 10 | CCAT1 | Upregulation | Encourage cell division | miR-218/ZFX | [40] |
| 11 | ASRPS | Downregulation | Inhibit angiogenesis and cell proliferation | STAT3 | [41] |
| 12 | AND2-AS1 | Downregulation | Inhibit angiogenesis inhibit cell division | RUNX2 | [42] |
| 13 | POU3F3 | Upregulation | Promote cell proliferation and inhibit cell apoptosis | Caspase-9 | [43] |
| 14 | NEF | Downregulation | Inhibit cell migration and invasion | miR-155 | [44] |
| 15 | ZEB2-AS1 | Upregulation | Promote cell proliferation, metastasis, and EMT | ZEB2 | [45] |
| 16 | LINC0009 | Upregulation | Increase cell proliferation and invasion | miR-383-5p/RBM3 | [46] |
| 17 | ANRIL | Upregulation | Increase cell proliferation and apoptosis | miR-448/KDM5B | [47] |
| 18 | SNHG12 | Upregulation | Induce cell proliferation, migration, and apoptosis | MMP13 | [48] |
| 19 | LUCAT1 | Upregulation | Encourage cell division, movement, and invasion | miR-5702 | [49] |
| 20 | PCAT6 | Upregulation | Radiotherapy resistance | miR-185-5p/TPD52 | [50] |
| 22 | HULC | Upregulation | Promote metastasis | MMP-2, MMP-9 | [51] |
| 23 | PAPAS | Upregulation | Induce cell migration and invasion | miR-34a | [52] |
| 24 | HCP5 | Upregulation | Increase cell proliferation; reduce cell apoptosis | miR-219a-5p/BIRC3 | [53] |
| 25 | NRAD1 | Upregulation | Stimulate cell proliferation and CSC-like phenotypic traits | miR-219a-5p/BIRC3 | [54] |
| 26 | SNAR | Upregulation | Stimulate cell division | [55] | |
| 27 | AWPPH | Upregulation | Activate cell proliferation | miR-21; FZD7 | [56] |
| 28 | sONE | Downregulation | Prevent cell proliferation | TP53/c-Myc | [57] |
| 29 | DANCR | Upregulation | Promote cell proliferation and invasion | miR-216a-5p | [58] |
| 30 | LINK-A | Upregulation | Increase resistance to immunotherapy, AKT inhibitors, and glycolysis reprogramming | PI3K/GPCR | [59] |
| 31 | MIR503HG | Downregulation | Reduce cell migration and invasion | miR-103/OLFM4 | [60] |
| 32 | NEAT1 | Upregulation | Increase cell apoptosis | [61] | |
| 33 | PTCSC3 | Downregulation | Prevent cell proliferation | H19 | [62] |
| 34 | NRON | Downregulation | Inhibit cell proliferation | snaR | [63] |
| 35 | TROJAN | Upregulation | Promote cell proliferation and invasion | ZMYND8 | [64] |
| 36 | NAMPT-AS | Upregulation | Increase cell metastasis | miR-548b-3p/NAMPT | [14] |
| 37 | MANCR | Upregulation | Promote cell proliferation; inhibit DNA damage | [65] | |
| 38 | RMST | Downregulation | Prevent cell proliferation | [66] | |
| 39 | SK AI1BC | Upregulation | Increase cell migration and invasion | K AI1 | [67] |
| 40 | ROR | Upregulation | Promote cell invasion and metastasis | miR-145/ARF6 | [68] |
| 41 | AIRN | Downregulation | Inhibit cell migration and invasion | Wnt/β-catenin/mTOR/PI3K | [69] |
| 42 | LINC-ZNF469-3 | Upregulation | Promote cell invasion | miR-574-5p/ZEB1 | [70] |
| 43 | PDCD4-AS1 | Downregulation | Inhibit cell proliferation and migration | PDCD4 | [71] |
| 44 | HOST2 | Downregulation | Inhibit cell proliferation | et-7 b/CDK6 | [72] |
| 45 | BORG | Upregulation | Promote doxorubicin resistance | RPA1 | [73] |
| 46 | PVT1 | Upregulation | Promote cell proliferation and migration, and EMT | p21, KLF5/β-catenin | [24] |
| 47 | H19 | Upregulation | Promote paclitaxel resistance and CSC-like phenotypic traits | Akt | [62] |
| 48 | TP73-AS1 | Downregulation | Promote cell vasculogenic mimicry | miR-490-3p/TWIST1 | [74] |
| 49 | TUG1 | Downregulation | Enhance cisplatin sensitivity | miR-197/NLK | [75] |
| 50 | MIR100HG | Upregulation | Promote cell proliferation | p27 | [76] |
| 51 | LINC01638 | Upregulation | Promote cell proliferation | c-Myc | [77] |
Table 2.
lncRNAs participate in the drug resistance of TNBC treatment.
| S.N. | lncRNA | Expression Patterns | Pathway/Target | Expression Pattern Drugs | Reference |
|---|---|---|---|---|---|
| 1 | H19 | Upregulation | H19/Let-7/LIN28 axis | Anthracycline, paclitaxel, fulverstrant, doxorubicin tamoxifen | [19] |
| 2 | NEAT1 | Upregulation | miR-211/HMGA2 | Paclitaxel, 5-FU, cisplatin | [61] |
| 3 | GAS5 | Downregulation | miR-21/mTOR/ | Adriamycin, trastuzumab, tamoxifen, paclitaxel | [91] |
| 4 | LINK-A | Upregulation | PIP3/GPCR/cAMP/PKA/ TRIM71/PLC | Immune checkpoint blockers | [101] |
| 5 | UCA1 | Upregulation | Wnt/b-catenin signalling | Trastuzumab, tamoxifen | [89] |
| 6 | LINP1 | Upregulation | Caspase-9/Bax | Doxorubicin, 5-FU, tamoxifen | [105] |
| 7 | Linc-ROR | Upregulation | miR-194-3p/MECP2 | Tamoxifen, paclitaxel, 5-FU | [106] |
| 8 | TMPO-AS1 | Upregulation | ER | Endocrine therapy | [107] |
| 9 | DCST1-AS1 | Upregulation | ANXA1 | Doxorubicin, paclitaxel | [108] |
| 10 | TINCR | Upregulation | miR-125b/ERBB2 | Trastuzumab | [109] |
| 11 | HOTAIR | Upregulation | ER | Tamoxifen, doxorubicin, trastuzumab, | [100] |
| 12 | AFAP1-AS1 | Upregulation | AUF1/ERBB2 | Trastuzumab | [98] |
| 13 | AGAP2-AS1 | Upregulation | CBP/ MyD88/H3K27/NF-kB | Trastuzumab | [110] |
| 14 | AK124454 | Upregulation | Paclitaxel | [111] | |
| 15 | NONHSAT057282 | Upregulation | ELF1 and E2F1 | Anthracycline | [112] |
| 16 | NONHSAG023333 | Upregulation | ELF1 and E2F1 | Anthracycline | [112] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Singh, D.D.; Lee, H.-J.; Yadav, D.K. Recent Clinical Advances on Long Non-Coding RNAs in Triple-Negative Breast Cancer. Cells 2023, 12, 674. https://doi.org/10.3390/cells12040674
AMA Style
Singh DD, Lee H-J, Yadav DK. Recent Clinical Advances on Long Non-Coding RNAs in Triple-Negative Breast Cancer. Cells. 2023; 12(4):674. https://doi.org/10.3390/cells12040674
Chicago/Turabian StyleSingh, Desh Deepak, Hae-Jeung Lee, and Dharmendra Kumar Yadav. 2023. "Recent Clinical Advances on Long Non-Coding RNAs in Triple-Negative Breast Cancer" Cells 12, no. 4: 674. https://doi.org/10.3390/cells12040674
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.