
Cross-Talk of NADPH Oxidases and Inflammation in Obesity

 , ,
, ,  , ,
, , 
Abstract
1. Introduction
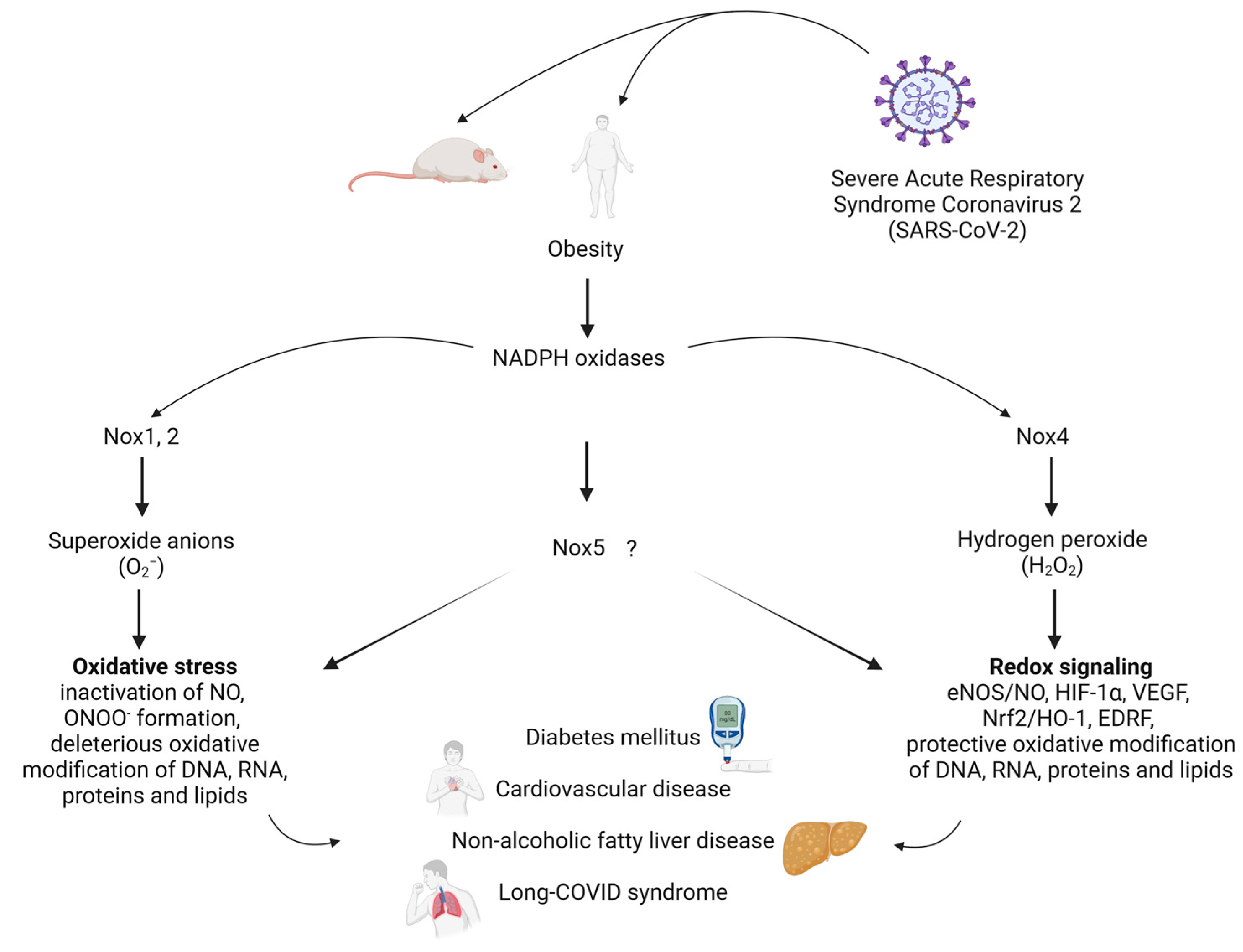
2. NADPH Oxidases and Obesity
2.1. NOX1
2.2. NOX2
2.3. NOX4
2.4. NOX5
3. Cross-Talk of NADPH Oxidases, Inflammation, Hypercholesterolemia and Obesity
4. Oxidative Stress and NADPH Oxidases in Non-Alcoholic Fatty Liver Disease
5. COVID-19, Obesity, Inflammation and Oxidative Stress in Cardiometabolic Disorders
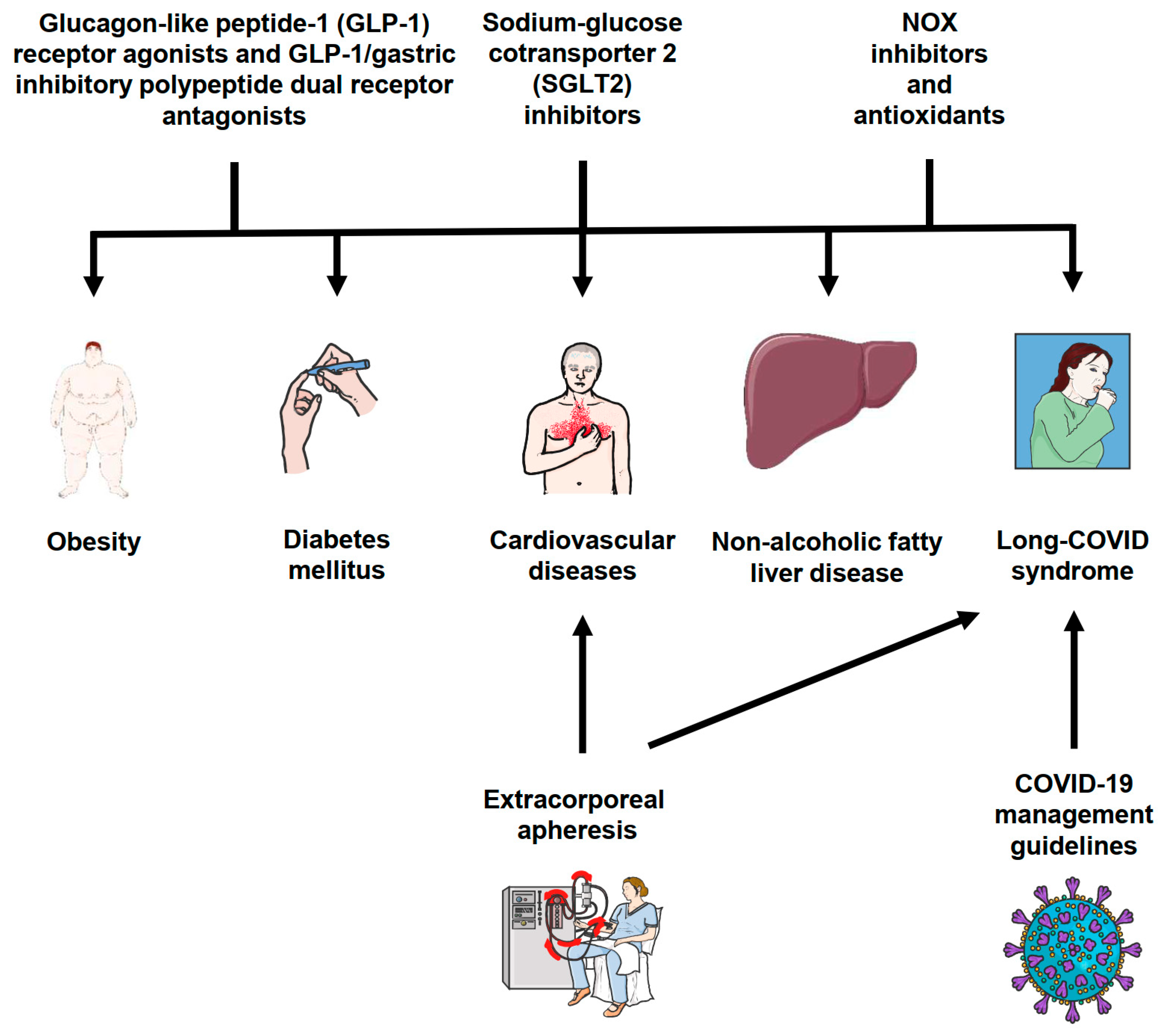
6. Clinical Implications
7. Open Questions
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Global Health Estimates 2020: Deaths by Cause, Age, Sex, by Country and by Region, 2000–2019; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Powell-Wiley, T.M.; Poirier, P.; Burke, L.E.; Despres, J.P.; Gordon-Larsen, P.; Lavie, C.J.; Lear, S.A.; Ndumele, C.E.; Neeland, I.J.; Sanders, P.; et al. Obesity and Cardiovascular Disease: A Scientific Statement From the American Heart Association. Circulation 2021, 143, e984–e1010. [Google Scholar] [CrossRef]
- Bornstein, S.R.; Ehrhart-Bornstein, M.; Wong, M.L.; Licinio, J. Is the worldwide epidemic of obesity a communicable feature of globalization? Exp. Clin. Endocrinol. Diabetes 2008, 116 (Suppl. S1), S30–S32. [Google Scholar] [CrossRef]
- Loffler, M.C.; Betz, M.J.; Blondin, D.P.; Augustin, R.; Sharma, A.K.; Tseng, Y.H.; Scheele, C.; Zimdahl, H.; Mark, M.; Hennige, A.M.; et al. Challenges in tackling energy expenditure as obesity therapy: From preclinical models to clinical application. Mol. Metab. 2021, 51, 101237. [Google Scholar] [CrossRef]
- Fernandez-Sanchez, A.; Madrigal-Santillan, E.; Bautista, M.; Esquivel-Soto, J.; Morales-Gonzalez, A.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sanchez-Rivera, G.; Valadez-Vega, C.; Morales-Gonzalez, J.A. Inflammation, oxidative stress, and obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef]
- Amponsah-Offeh, M.; Diaba-Nuhoho, P.; Speier, S.; Morawietz, H. Oxidative Stress, Antioxidants and Hypertension. Antioxidants 2023, 12, 281. [Google Scholar] [CrossRef]
- Rotariu, D.; Babes, E.E.; Tit, D.M.; Moisi, M.; Bustea, C.; Stoicescu, M.; Radu, A.F.; Vesa, C.M.; Behl, T.; Bungau, A.F.; et al. Oxidative stress—Complex pathological issues concerning the hallmark of cardiovascular and metabolic disorders. Biomed. Pharmacother. 2022, 152, 113238. [Google Scholar] [CrossRef]
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C. Oxidative stress and metabolic disorders: Pathogenesis and therapeutic strategies. Life Sci. 2016, 148, 183–193. [Google Scholar] [CrossRef]
- Steven, S.; Frenis, K.; Oelze, M.; Kalinovic, S.; Kuntic, M.; Bayo Jimenez, M.T.; Vujacic-Mirski, K.; Helmstadter, J.; Kroller-Schon, S.; Munzel, T.; et al. Vascular Inflammation and Oxidative Stress: Major Triggers for Cardiovascular Disease. Oxid. Med. Cell Longev. 2019, 2019, 7092151. [Google Scholar] [CrossRef]
- Grandl, G.; Wolfrum, C. Hemostasis, endothelial stress, inflammation, and the metabolic syndrome. Semin. Immunopathol. 2018, 40, 215–224. [Google Scholar] [CrossRef]
- Muller, G.; Morawietz, H. Nitric oxide, NAD(P)H oxidase, and atherosclerosis. Antioxid. Redox Signal. 2009, 11, 1711–1731. [Google Scholar] [CrossRef]
- Brandes, R.P.; Weissmann, N.; Schroder, K. NADPH oxidases in cardiovascular disease. Free Radic. Biol. Med. 2010, 49, 687–706. [Google Scholar]
- Cave, A.C.; Brewer, A.C.; Narayanapanicker, A.; Ray, R.; Grieve, D.J.; Walker, S.; Shah, A.M. NADPH oxidases in cardiovascular health and disease. Antioxid. Redox Signal. 2006, 8, 691–728. [Google Scholar] [CrossRef]
- Lassegue, B.; Griendling, K.K. NADPH oxidases: Functions and pathologies in the vasculature. Arter. Thromb. Vasc. Biol. 2010, 30, 653–661. [Google Scholar]
- Rueckschloss, U.; Duerrschmidt, N.; Morawietz, H. NADPH oxidase in endothelial cells: Impact on atherosclerosis. Antioxid. Redox Signal. 2003, 5, 171–180. [Google Scholar] [CrossRef]
- Schroder, K. NADPH oxidases: Current aspects and tools. Redox Biol. 2020, 34, 101512. [Google Scholar] [CrossRef]
- Drummond, G.R.; Selemidis, S.; Griendling, K.K.; Sobey, C.G. Combating oxidative stress in vascular disease: NADPH oxidases as therapeutic targets. Nat. Rev. Drug Discov. 2011, 10, 453–471. [Google Scholar] [CrossRef]
- Lambeth, J.D. NOX enzymes and the biology of reactive oxygen. Nat. Rev. Immunol. 2004, 4, 181–189. [Google Scholar]
- Lambeth, J.D.; Cheng, G.; Arnold, R.S.; Edens, W.A. Novel homologs of gp91phox. Trends Biochem. Sci. 2000, 25, 459–461. [Google Scholar]
- El-Benna, J.; Dang, P.M.; Gougerot-Pocidalo, M.A. Priming of the neutrophil NADPH oxidase activation: Role of p47phox phosphorylation and NOX2 mobilization to the plasma membrane. Semin. Immunopathol. 2008, 30, 279–289. [Google Scholar] [CrossRef]
- Nauseef, W.M. The phagocyte NOX2 NADPH oxidase in microbial killing and cell signaling. Curr. Opin. Immunol. 2019, 60, 130–140. [Google Scholar] [CrossRef]
- Drummond, G.R.; Sobey, C.G. Endothelial NADPH oxidases: Which NOX to target in vascular disease? Trends Endocrinol. Metab. 2014, 25, 452–463. [Google Scholar] [CrossRef]
- Muller, G.; Morawietz, H. NAD(P)H oxidase and endothelial dysfunction. Horm. Metab. Res. 2009, 41, 152–158. [Google Scholar] [CrossRef]
- Morawietz, H. Cardiovascular protection by Nox4. Cardiovasc. Res. 2018, 114, 353–355. [Google Scholar] [CrossRef]
- Targher, G.; Day, C.P.; Bonora, E. Risk of cardiovascular disease in patients with nonalcoholic fatty liver disease. N. Engl. J. Med. 2010, 363, 1341–1350. [Google Scholar] [CrossRef]
- Peiseler, M.; Schwabe, R.; Hampe, J.; Kubes, P.; Heikenwalder, M.; Tacke, F. Immune mechanisms linking metabolic injury to inflammation and fibrosis in fatty liver disease—Novel insights into cellular communication circuits. J. Hepatol. 2022, 77, 1136–1160. [Google Scholar] [CrossRef]
- Gabbia, D.; Cannella, L.; De Martin, S. The Role of Oxidative Stress in NAFLD-NASH-HCC Transition-Focus on NADPH Oxidases. Biomedicines 2021, 9, 687. [Google Scholar] [CrossRef]
- Nasce, A.; Gariani, K.; Jornayvaz, F.R.; Szanto, I. NADPH Oxidases Connecting Fatty Liver Disease, Insulin Resistance and Type 2 Diabetes: Current Knowledge and Therapeutic Outlook. Antioxidants 2022, 11, 1131. [Google Scholar] [CrossRef]
- Perakakis, N.; Barthel, A.; Bornstein, S.R. The Role of Endocrine and Metabolic System in COVID-19 Disease—The Transcampus Experience and Review of Evidence From International Collaborating Groups. Horm. Metab. Res. 2022, 54, 494–495. [Google Scholar] [CrossRef]
- Steenblock, C.; Schwarz, P.E.H.; Ludwig, B.; Linkermann, A.; Zimmet, P.; Kulebyakin, K.; Tkachuk, V.A.; Markov, A.G.; Lehnert, H.; de Angelis, M.H.; et al. COVID-19 and metabolic disease: Mechanisms and clinical management. Lancet Diabetes Endocrinol. 2021, 9, 786–798. [Google Scholar] [CrossRef]
- Morawietz, H.; Julius, U.; Bornstein, S.R. Cardiovascular diseases, lipid-lowering therapies and European registries in the COVID-19 pandemic. Cardiovasc. Res. 2020, 116, e122–e125. [Google Scholar] [CrossRef]
- Moreno-Fernandez, J.; Ochoa, J.; Ojeda, M.L.; Nogales, F.; Carreras, O.; Diaz-Castro, J. Inflammation and oxidative stress, the links between obesity and COVID-19: A narrative review. J. Physiol. Biochem. 2022, 78, 581–591. [Google Scholar] [CrossRef]
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive Oxygen Species in Metabolic and Inflammatory Signaling. Circ. Res. 2018, 122, 877–902. [Google Scholar] [CrossRef]
- Jiang, F.; Lim, H.K.; Morris, M.J.; Prior, L.; Velkoska, E.; Wu, X.; Dusting, G.J. Systemic upregulation of NADPH oxidase in diet-induced obesity in rats. Redox Rep. 2011, 16, 223–229. [Google Scholar] [CrossRef]
- Youn, J.Y.; Siu, K.L.; Lob, H.E.; Itani, H.; Harrison, D.G.; Cai, H. Role of vascular oxidative stress in obesity and metabolic syndrome. Diabetes 2014, 63, 2344–2355. [Google Scholar] [CrossRef]
- Xu, L.; Balzarolo, M.; Robinson, E.L.; Lorenz, V.; Della Verde, G.; Joray, L.; Mochizuki, M.; Kaufmann, B.A.; Valstar, G.; de Jager, S.C.A.; et al. NOX1 mediates metabolic heart disease in mice and is upregulated in monocytes of humans with diastolic dysfunction. Cardiovasc. Res. 2022, 118, 2973–2984. [Google Scholar] [CrossRef]
- Thompson, J.A.; Larion, S.; Mintz, J.D.; Belin de Chantemele, E.J.; Fulton, D.J.; Stepp, D.W. Genetic Deletion of NADPH Oxidase 1 Rescues Microvascular Function in Mice with Metabolic Disease. Circ. Res. 2017, 121, 502–511. [Google Scholar] [CrossRef]
- Sheehan, A.L.; Carrell, S.; Johnson, B.; Stanic, B.; Banfi, B.; Miller, F.J., Jr. Role for Nox1 NADPH oxidase in atherosclerosis. Atherosclerosis 2011, 216, 321–326. [Google Scholar] [CrossRef]
- Sela, M.; Tirza, G.; Ravid, O.; Volovitz, I.; Solodeev, I.; Friedman, O.; Zipori, D.; Gur, E.; Krelin, Y.; Shani, N. NOX1-induced accumulation of reactive oxygen species in abdominal fat-derived mesenchymal stromal cells impinges on long-term proliferation. Cell Death Dis. 2015, 6, e1728. [Google Scholar] [CrossRef]
- Saenz-Medina, J.; Munoz, M.; Sanchez, A.; Rodriguez, C.; Jorge, E.; Corbacho, C.; Izquierdo, D.; Santos, M.; Donoso, E.; Virumbrales, E.; et al. Nox1-derived oxidative stress as a common pathogenic link between obesity and hyperoxaluria-related kidney injury. Urolithiasis 2020, 48, 481–492. [Google Scholar] [CrossRef]
- Violi, F.; Sanguigni, V.; Carnevale, R.; Plebani, A.; Rossi, P.; Finocchi, A.; Pignata, C.; De Mattia, D.; Martire, B.; Pietrogrande, M.C.; et al. Hereditary deficiency of gp91(phox) is associated with enhanced arterial dilatation: Results of a multicenter study. Circulation 2009, 120, 1616–1622. [Google Scholar] [CrossRef]
- Loffredo, L.; Martino, F.; Carnevale, R.; Pignatelli, P.; Catasca, E.; Perri, L.; Calabrese, C.M.; Palumbo, M.M.; Baratta, F.; Del Ben, M.; et al. Obesity and hypercholesterolemia are associated with NOX2 generated oxidative stress and arterial dysfunction. J. Pediatr. 2012, 161, 1004–1009. [Google Scholar] [CrossRef]
- DeVallance, E.; Branyan, K.W.; Lemaster, K.; Olfert, I.M.; Smith, D.M.; Pistilli, E.E.; Frisbee, J.C.; Chantler, P.D. Aortic dysfunction in metabolic syndrome mediated by perivascular adipose tissue TNFalpha- and NOX2-dependent pathway. Exp. Physiol. 2018, 103, 590–603. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Ali, M.M.; Miranda, E.R.; Mey, J.T.; Blackburn, B.K.; Haus, J.M.; Phillips, S.A. Nox2 contributes to hyperinsulinemia-induced redox imbalance and impaired vascular function. Redox Biol. 2017, 13, 288–300. [Google Scholar] [CrossRef]
- Du, J.; Fan, L.M.; Mai, A.; Li, J.M. Crucial roles of Nox2-derived oxidative stress in deteriorating the function of insulin receptors and endothelium in dietary obesity of middle-aged mice. Br. J. Pharmacol. 2013, 170, 1064–1077. [Google Scholar] [CrossRef]
- Rahman, M.M.; El Jamali, A.; Halade, G.V.; Ouhtit, A.; Abou-Saleh, H.; Pintus, G. Nox2 Activity Is Required in Obesity-Mediated Alteration of Bone Remodeling. Oxid. Med. Cell Longev. 2018, 2018, 6054361. [Google Scholar] [CrossRef]
- Hafstad, A.D.; Hansen, S.S.; Lund, J.; Santos, C.X.C.; Boardman, N.T.; Shah, A.M.; Aasum, E. NADPH Oxidase 2 Mediates Myocardial Oxygen Wasting in Obesity. Antioxidants 2020, 9, 171. [Google Scholar] [CrossRef]
- Pepping, J.K.; Vandanmagsar, B.; Fernandez-Kim, S.O.; Zhang, J.; Mynatt, R.L.; Bruce-Keller, A.J. Myeloid-specific deletion of NOX2 prevents the metabolic and neurologic consequences of high fat diet. PLoS ONE 2017, 12, e0181500. [Google Scholar] [CrossRef]
- Schroder, K.; Wandzioch, K.; Helmcke, I.; Brandes, R.P. Nox4 acts as a switch between differentiation and proliferation in preadipocytes. Arter. Thromb. Vasc. Biol. 2009, 29, 239–245. [Google Scholar] [CrossRef]
- Den Hartigh, L.J.; Omer, M.; Goodspeed, L.; Wang, S.; Wietecha, T.; O’Brien, K.D.; Han, C.Y. Adipocyte-Specific Deficiency of NADPH Oxidase 4 Delays the Onset of Insulin Resistance and Attenuates Adipose Tissue Inflammation in Obesity. Arter. Thromb. Vasc. Biol. 2017, 37, 466–475. [Google Scholar] [CrossRef]
- Hua, H.; Wu, M.; Wu, T.; Ji, Y.; Jin, L.; Du, Y.; Zhang, Y.; Huang, S.; Zhang, A.; Ding, G.; et al. Reduction of NADPH oxidase 4 in adipocytes contributes to the anti-obesity effect of dihydroartemisinin. Heliyon 2023, 9, e14028. [Google Scholar] [CrossRef]
- Li, Y.; Mouche, S.; Sajic, T.; Veyrat-Durebex, C.; Supale, R.; Pierroz, D.; Ferrari, S.; Negro, F.; Hasler, U.; Feraille, E.; et al. Deficiency in the NADPH oxidase 4 predisposes towards diet-induced obesity. Int. J. Obes. 2012, 36, 1503–1513. [Google Scholar] [CrossRef]
- Ray, R.; Murdoch, C.E.; Wang, M.; Santos, C.X.; Zhang, M.; Alom-Ruiz, S.; Anilkumar, N.; Ouattara, A.; Cave, A.C.; Walker, S.J.; et al. Endothelial Nox4 NADPH oxidase enhances vasodilatation and reduces blood pressure in vivo. Arter. Thromb. Vasc. Biol. 2011, 31, 1368–1376. [Google Scholar] [CrossRef]
- Zhang, M.; Mongue-Din, H.; Martin, D.; Catibog, N.; Smyrnias, I.; Zhang, X.; Yu, B.; Wang, M.; Brandes, R.P.; Schroder, K.; et al. Both cardiomyocyte and endothelial cell Nox4 mediate protection against hemodynamic overload-induced remodelling. Cardiovasc. Res. 2018, 114, 401–408. [Google Scholar] [CrossRef]
- Schroder, K.; Zhang, M.; Benkhoff, S.; Mieth, A.; Pliquett, R.; Kosowski, J.; Kruse, C.; Luedike, P.; Michaelis, U.R.; Weissmann, N.; et al. Nox4 is a protective reactive oxygen species generating vascular NADPH oxidase. Circ. Res. 2012, 110, 1217–1225. [Google Scholar] [CrossRef]
- Schurmann, C.; Rezende, F.; Kruse, C.; Yasar, Y.; Lowe, O.; Fork, C.; van de Sluis, B.; Bremer, R.; Weissmann, N.; Shah, A.M.; et al. The NADPH oxidase Nox4 has anti-atherosclerotic functions. Eur. Heart J. 2015, 36, 3447–3456. [Google Scholar] [CrossRef]
- Langbein, H.; Brunssen, C.; Hofmann, A.; Cimalla, P.; Brux, M.; Bornstein, S.R.; Deussen, A.; Koch, E.; Morawietz, H. NADPH oxidase 4 protects against development of endothelial dysfunction and atherosclerosis in LDL receptor deficient mice. Eur. Heart J. 2016, 37, 1753–1761. [Google Scholar] [CrossRef]
- Brendel, H.; Shahid, A.; Hofmann, A.; Mittag, J.; Bornstein, S.R.; Morawietz, H.; Brunssen, C. NADPH oxidase 4 mediates the protective effects of physical activity against obesity-induced vascular dysfunction. Cardiovasc. Res. 2020, 116, 1767–1778. [Google Scholar] [CrossRef]
- Touyz, R.M.; Anagnostopoulou, A.; Camargo, L.L.; Rios, F.J.; Montezano, A.C. Vascular Biology of Superoxide-Generating NADPH Oxidase 5-Implications in Hypertension and Cardiovascular Disease. Antioxid. Redox Signal. 2019, 30, 1027–1040. [Google Scholar] [CrossRef]
- Guzik, T.J.; Chen, W.; Gongora, M.C.; Guzik, B.; Lob, H.E.; Mangalat, D.; Hoch, N.; Dikalov, S.; Rudzinski, P.; Kapelak, B.; et al. Calcium-dependent NOX5 nicotinamide adenine dinucleotide phosphate oxidase contributes to vascular oxidative stress in human coronary artery disease. J. Am. Coll. Cardiol. 2008, 52, 1803–1809. [Google Scholar] [CrossRef]
- Garcia, J.G.; Ansorena, E.; Milagro, F.I.; Zalba, G.; de Miguel, C. Endothelial Nox5 Expression Modulates Glucose Uptake and Lipid Accumulation in Mice Fed a High-Fat Diet and 3T3-L1 Adipocytes Treated with Glucose and Palmitic Acid. Int. J. Mol. Sci. 2021, 22, 2729. [Google Scholar] [CrossRef]
- Garcia, J.G.; de Miguel, C.; Milagro, F.I.; Zalba, G.; Ansorena, E. Endothelial NOX5 Expression Modulates Thermogenesis and Lipolysis in Mice Fed with a High-Fat Diet and 3T3-L1 Adipocytes through an Interleukin-6 Dependent Mechanism. Antioxidants 2021, 11, 30. [Google Scholar] [CrossRef]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef]
- Morawietz, H. Identification of a novel redox switch between metabolism and cardiac function using HyPer power. Pflug. Arch. 2023, 475, 907–909. [Google Scholar] [CrossRef]
- Ghanim, H.; Garg, R.; Aljada, A.; Mohanty, P.; Kumbkarni, Y.; Assian, E.; Hamouda, W.; Dandona, P. Suppression of nuclear factor-kappaB and stimulation of inhibitor kappaB by troglitazone: Evidence for an anti-inflammatory effect and a potential antiatherosclerotic effect in the obese. J. Clin. Endocrinol. Metab. 2001, 86, 1306–1312. [Google Scholar] [CrossRef][Green Version]
- Belambri, S.A.; Rolas, L.; Raad, H.; Hurtado-Nedelec, M.; Dang, P.M.; El-Benna, J. NADPH oxidase activation in neutrophils: Role of the phosphorylation of its subunits. Eur. J. Clin. Investig. 2018, 48 (Suppl. S2), e12951. [Google Scholar] [CrossRef]
- Bouraoui, A.; Louzada, R.A.; Aimeur, S.; Waeytens, J.; Wien, F.; My-Chan Dang, P.; Bizouarn, T.; Dupuy, C.; Baciou, L. New insights in the molecular regulation of the NADPH oxidase 2 activity: Negative modulation by Poldip2. Free Radic. Biol. Med. 2023, 199, 113–125. [Google Scholar] [CrossRef]
- Rivera, J.; Sobey, C.G.; Walduck, A.K.; Drummond, G.R. Nox isoforms in vascular pathophysiology: Insights from transgenic and knockout mouse models. Redox Rep. 2010, 15, 50–63. [Google Scholar] [CrossRef]
- Xue, S.; Tang, H.; Zhao, G.; Fang, C.; Shen, Y.; Yan, D.; Yuan, Y.; Fu, W.; Shi, Z.; Tang, X.; et al. C-C motif ligand 8 promotes atherosclerosis via NADPH oxidase 2/reactive oxygen species-induced endothelial permeability increase. Free Radic. Biol. Med. 2021, 167, 181–192. [Google Scholar] [CrossRef]
- Buchmann, G.K.; Schurmann, C.; Warwick, T.; Schulz, M.H.; Spaeth, M.; Muller, O.J.; Schroder, K.; Jo, H.; Weissmann, N.; Brandes, R.P. Deletion of NoxO1 limits atherosclerosis development in female mice. Redox Biol. 2020, 37, 101713. [Google Scholar] [CrossRef]
- Mima, A.; Qi, W.; Hiraoka-Yamomoto, J.; Park, K.; Matsumoto, M.; Kitada, M.; Li, Q.; Mizutani, K.; Yu, E.; Shimada, T.; et al. Retinal not systemic oxidative and inflammatory stress correlated with VEGF expression in rodent models of insulin resistance and diabetes. Investig. Ophthalmol. Vis. Sci. 2012, 53, 8424–8432. [Google Scholar] [CrossRef]
- Peng, Y.; Li, J.Z.; You, M.; Murr, M.M. Roux-en-Y gastric bypass improves glucose homeostasis, reduces oxidative stress and inflammation in livers of obese rats and in Kupffer cells via an AMPK-dependent pathway. Surgery 2017, 162, 59–67. [Google Scholar] [CrossRef]
- Najjar, R.S.; Knapp, D.; Wanders, D.; Feresin, R.G. Raspberry and blackberry act in a synergistic manner to improve cardiac redox proteins and reduce NF-kappaB and SAPK/JNK in mice fed a high-fat, high-sucrose diet. Nutr. Metab. Cardiovasc. Dis. NMCD 2022, 32, 1784–1796. [Google Scholar] [CrossRef]
- Hofmann, A.; Brunssen, C.; Morawietz, H. Contribution of lectin-like oxidized low-density lipoprotein receptor-1 and LOX-1 modulating compounds to vascular diseases. Vasc. Pharmacol. 2018, 107, 1–11. [Google Scholar] [CrossRef]
- Sawamura, T.; Kume, N.; Aoyama, T.; Moriwaki, H.; Hoshikawa, H.; Aiba, Y.; Tanaka, T.; Miwa, S.; Katsura, Y.; Kita, T.; et al. An endothelial receptor for oxidized low-density lipoprotein. Nature 1997, 386, 73–77. [Google Scholar]
- Morawietz, H.; Rueckschloss, U.; Niemann, B.; Duerrschmidt, N.; Galle, J.; Hakim, K.; Zerkowski, H.R.; Sawamura, T.; Holtz, J. Angiotensin II induces LOX-1, the human endothelial receptor for oxidized low-density lipoprotein. Circulation 1999, 100, 899–902. [Google Scholar] [CrossRef]
- Morawietz, H.; Duerrschmidt, N.; Niemann, B.; Galle, J.; Sawamura, T.; Holtz, J. Induction of the oxLDL receptor LOX-1 by endothelin-1 in human endothelial cells. Biochem. Biophys. Res. Commun. 2001, 284, 961–965. [Google Scholar] [CrossRef]
- Catar, R.; Chen, L.; Zhao, H.; Wu, D.; Kamhieh-Milz, J.; Lucht, C.; Zickler, D.; Krug, A.W.; Ziegler, C.G.; Morawietz, H.; et al. Native and Oxidized Low-Density Lipoproteins Increase the Expression of the LDL Receptor and the LOX-1 Receptor, Respectively, in Arterial Endothelial Cells. Cells 2022, 11, 204. [Google Scholar] [CrossRef]
- Kamhieh-Milz, J.; Chen, L.; Goettsch, C.; Pfefferkorn, A.M.; Hofmann, A.; Brunssen, C.; Muller, G.; Walther, T.; Ashraf, M.I.; Moll, G.; et al. Ang II Promotes ET-1 Production by Regulating NOX2 Activity Through Transcription Factor Oct-1. Arter. Thromb. Vasc. Biol. 2023, 43, 1429–1440. [Google Scholar] [CrossRef]
- Riazi, K.; Azhari, H.; Charette, J.H.; Underwood, F.E.; King, J.A.; Afshar, E.E.; Swain, M.G.; Congly, S.E.; Kaplan, G.G.; Shaheen, A.A. The prevalence and incidence of NAFLD worldwide: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2022, 7, 851–861. [Google Scholar] [CrossRef]
- Loomba, R.; Friedman, S.L.; Shulman, G.I. Mechanisms and disease consequences of nonalcoholic fatty liver disease. Cell 2021, 184, 2537–2564. [Google Scholar] [CrossRef]
- Samuel, V.T.; Shulman, G.I. Nonalcoholic Fatty Liver Disease as a Nexus of Metabolic and Hepatic Diseases. Cell Metab. 2018, 27, 22–41. [Google Scholar] [CrossRef]
- Perakakis, N.; Stefanakis, K.; Mantzoros, C.S. The role of omics in the pathophysiology, diagnosis and treatment of non-alcoholic fatty liver disease. Metabolism 2020, 111S, 154320. [Google Scholar] [CrossRef]
- Polyzos, S.A.; Perakakis, N.; Boutari, C.; Kountouras, J.; Ghaly, W.; Anastasilakis, A.D.; Karagiannis, A.; Mantzoros, C.S. Targeted Analysis of Three Hormonal Systems Identifies Molecules Associated with the Presence and Severity of NAFLD. J. Clin. Endocrinol. Metab. 2020, 105, e390–e400. [Google Scholar] [CrossRef]
- Arroyave-Ospina, J.C.; Wu, Z.; Geng, Y.; Moshage, H. Role of Oxidative Stress in the Pathogenesis of Non-Alcoholic Fatty Liver Disease: Implications for Prevention and Therapy. Antioxidants 2021, 10, 174. [Google Scholar] [CrossRef]
- Matsumoto, M.; Zhang, J.; Zhang, X.; Liu, J.; Jiang, J.X.; Yamaguchi, K.; Taruno, A.; Katsuyama, M.; Iwata, K.; Ibi, M.; et al. The NOX1 isoform of NADPH oxidase is involved in dysfunction of liver sinusoids in nonalcoholic fatty liver disease. Free Radic. Biol. Med. 2018, 115, 412–420. [Google Scholar] [CrossRef]
- Liang, S.; Ma, H.Y.; Zhong, Z.; Dhar, D.; Liu, X.; Xu, J.; Koyama, Y.; Nishio, T.; Karin, D.; Karin, G.; et al. NADPH Oxidase 1 in Liver Macrophages Promotes Inflammation and Tumor Development in Mice. Gastroenterology 2019, 156, 1156–1172.e6. [Google Scholar] [CrossRef]
- Cui, W.; Matsuno, K.; Iwata, K.; Ibi, M.; Matsumoto, M.; Zhang, J.; Zhu, K.; Katsuyama, M.; Torok, N.J.; Yabe-Nishimura, C. NOX1/nicotinamide adenine dinucleotide phosphate, reduced form (NADPH) oxidase promotes proliferation of stellate cells and aggravates liver fibrosis induced by bile duct ligation. Hepatology 2011, 54, 949–958. [Google Scholar] [CrossRef]
- Luo, X.; Bai, Y.; He, S.; Sun, S.; Jiang, X.; Yang, Z.; Lu, D.; Wei, P.; Liang, Y.; Peng, C.; et al. Sirtuin 1 ameliorates defenestration in hepatic sinusoidal endothelial cells during liver fibrosis via inhibiting stress-induced premature senescence. Cell Prolif. 2021, 54, e12991. [Google Scholar] [CrossRef]
- Kim, S.Y.; Jeong, J.M.; Kim, S.J.; Seo, W.; Kim, M.H.; Choi, W.M.; Yoo, W.; Lee, J.H.; Shim, Y.R.; Yi, H.S.; et al. Pro-inflammatory hepatic macrophages generate ROS through NADPH oxidase 2 via endocytosis of monomeric TLR4-MD2 complex. Nat. Commun. 2017, 8, 2247. [Google Scholar] [CrossRef]
- Shiau, D.J.; Kuo, W.T.; Davuluri, G.V.N.; Shieh, C.C.; Tsai, P.J.; Chen, C.C.; Lin, Y.S.; Wu, Y.Z.; Hsiao, Y.P.; Chang, C.P. Hepatocellular carcinoma-derived high mobility group box 1 triggers M2 macrophage polarization via a TLR2/NOX2/autophagy axis. Sci. Rep. 2020, 10, 13582. [Google Scholar] [CrossRef]
- Coats, B.R.; Schoenfelt, K.Q.; Barbosa-Lorenzi, V.C.; Peris, E.; Cui, C.; Hoffman, A.; Zhou, G.; Fernandez, S.; Zhai, L.; Hall, B.A.; et al. Metabolically Activated Adipose Tissue Macrophages Perform Detrimental and Beneficial Functions during Diet-Induced Obesity. Cell Rep. 2017, 20, 3149–3161. [Google Scholar] [CrossRef]
- Pepping, J.K.; Freeman, L.R.; Gupta, S.; Keller, J.N.; Bruce-Keller, A.J. NOX2 deficiency attenuates markers of adiposopathy and brain injury induced by high-fat diet. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E392–E404. [Google Scholar] [CrossRef]
- Bettaieb, A.; Jiang, J.X.; Sasaki, Y.; Chao, T.I.; Kiss, Z.; Chen, X.; Tian, J.; Katsuyama, M.; Yabe-Nishimura, C.; Xi, Y.; et al. Hepatocyte Nicotinamide Adenine Dinucleotide Phosphate Reduced Oxidase 4 Regulates Stress Signaling, Fibrosis, and Insulin Sensitivity during Development of Steatohepatitis in Mice. Gastroenterology 2015, 149, 468–480.e10. [Google Scholar] [CrossRef]
- Eun, H.S.; Chun, K.; Song, I.S.; Oh, C.H.; Seong, I.O.; Yeo, M.K.; Kim, K.H. High nuclear NADPH oxidase 4 expression levels are correlated with cancer development and poor prognosis in hepatocellular carcinoma. Pathology 2019, 51, 579–585. [Google Scholar] [CrossRef]
- Carmona-Cuenca, I.; Roncero, C.; Sancho, P.; Caja, L.; Fausto, N.; Fernandez, M.; Fabregat, I. Upregulation of the NADPH oxidase NOX4 by TGF-beta in hepatocytes is required for its pro-apoptotic activity. J. Hepatol. 2008, 49, 965–976. [Google Scholar] [CrossRef]
- Yoshida, T.; Murayama, H.; Kawashima, M.; Nagahara, R.; Kangawa, Y.; Mizukami, S.; Kimura, M.; Abe, H.; Hayashi, S.M.; Shibutani, M. Apocynin and enzymatically modified isoquercitrin suppress the expression of a NADPH oxidase subunit p22phox in steatosis-related preneoplastic liver foci of rats. Exp. Toxicol. Pathol. 2017, 69, 9–16. [Google Scholar] [CrossRef]
- Owada, S.; Endo, H.; Okada, C.; Yoshida, K.; Shida, Y.; Tatemichi, M. Setanaxib as a Potent Hypoxia-specific Therapeutic Agent Against Liver Cancer. Anticancer Res. 2020, 40, 5071–5079. [Google Scholar] [CrossRef]
- Leisman, D.E.; Ronner, L.; Pinotti, R.; Taylor, M.D.; Sinha, P.; Calfee, C.S.; Hirayama, A.V.; Mastroiani, F.; Turtle, C.J.; Harhay, M.O.; et al. Cytokine elevation in severe and critical COVID-19: A rapid systematic review, meta-analysis, and comparison with other inflammatory syndromes. Lancet Respir. Med. 2020, 8, 1233–1244. [Google Scholar] [CrossRef]
- Bornstein, S.R.; Dalan, R.; Hopkins, D.; Mingrone, G.; Boehm, B.O. Endocrine and metabolic link to coronavirus infection. Nat. Rev. Endocrinol. 2020, 16, 297–298. [Google Scholar] [CrossRef]
- Hollstein, T.; Schulte, D.M.; Schulz, J.; Gluck, A.; Ziegler, A.G.; Bonifacio, E.; Wendorff, M.; Franke, A.; Schreiber, S.; Bornstein, S.R.; et al. Autoantibody-negative insulin-dependent diabetes mellitus after SARS-CoV-2 infection: A case report. Nat. Metab. 2020, 2, 1021–1024. [Google Scholar] [CrossRef]
- Rubino, F.; Amiel, S.A.; Zimmet, P.; Alberti, G.; Bornstein, S.; Eckel, R.H.; Mingrone, G.; Boehm, B.; Cooper, M.E.; Chai, Z.; et al. New-Onset Diabetes in COVID-19. N. Engl. J. Med. 2020, 383, 789–790. [Google Scholar] [CrossRef]
- Steenblock, C.; Richter, S.; Berger, I.; Barovic, M.; Schmid, J.; Schubert, U.; Jarzebska, N.; von Massenhausen, A.; Linkermann, A.; Schurmann, A.; et al. Viral infiltration of pancreatic islets in patients with COVID-19. Nat. Commun. 2021, 12, 3534. [Google Scholar] [CrossRef]
- Steenblock, C.; Hassanein, M.; Khan, E.G.; Yaman, M.; Kamel, M.; Barbir, M.; Lorke, D.E.; Rock, J.A.; Everett, D.; Bejtullah, S.; et al. Diabetes and COVID-19: Short- and Long-Term Consequences. Horm. Metab. Res. 2022, 54, 503–509. [Google Scholar] [CrossRef]
- Kanczkowski, W.; Evert, K.; Stadtmuller, M.; Haberecker, M.; Laks, L.; Chen, L.S.; Frontzek, K.; Pablik, J.; Hantel, C.; Beuschlein, F.; et al. COVID-19 targets human adrenal glands. Lancet Diabetes Endocrinol. 2022, 10, 13–16. [Google Scholar] [CrossRef]
- Kanczkowski, W.; Beuschlein, F.; Bornstein, S.R. Is there a role for the adrenal glands in long COVID? Nat. Rev. Endocrinol. 2022, 18, 451–452. [Google Scholar] [CrossRef]
- Tadayon Najafabadi, B.; Rayner, D.G.; Shokraee, K.; Shokraie, K.; Panahi, P.; Rastgou, P.; Seirafianpour, F.; Momeni Landi, F.; Alinia, P.; Parnianfard, N.; et al. Obesity as an independent risk factor for COVID-19 severity and mortality. Cochrane Database Syst. Rev. 2023, 5, CD015201. [Google Scholar] [CrossRef]
- Hoong, C.W.S.; Hussain, I.; Aravamudan, V.M.; Phyu, E.E.; Lin, J.H.X.; Koh, H. Obesity is Associated with Poor COVID-19 Outcomes: A Systematic Review and Meta-Analysis. Horm. Metab. Res. 2021, 53, 85–93. [Google Scholar] [CrossRef]
- Ayres, J.S. A metabolic handbook for the COVID-19 pandemic. Nat. Metab. 2020, 2, 572–585. [Google Scholar] [CrossRef]
- Russo, L.; Lumeng, C.N. Properties and functions of adipose tissue macrophages in obesity. Immunology 2018, 155, 407–417. [Google Scholar] [CrossRef]
- Hornung, F.; Rogal, J.; Loskill, P.; Loffler, B.; Deinhardt-Emmer, S. The Inflammatory Profile of Obesity and the Role on Pulmonary Bacterial and Viral Infections. Int. J. Mol. Sci. 2021, 22, 3456. [Google Scholar] [CrossRef]
- Steenblock, C.; Hassanein, M.; Khan, E.G.; Yaman, M.; Kamel, M.; Barbir, M.; Lorke, D.E.; Everett, D.; Bejtullah, S.; Lohmann, T.; et al. Obesity and COVID-19: What are the Consequences? Horm. Metab. Res. 2022, 54, 496–502. [Google Scholar] [CrossRef]
- Julius, U.; Schatz, U.; Tselmin, S.; Morawietz, H. COVID-19 and Lipid Disorders. Horm. Metab. Res. 2022, 54, 514–521. [Google Scholar] [CrossRef]
- Steenblock, C.; Walther, R.; Tselmin, S.; Jarzebska, N.; Voit-Bak, K.; Toepfner, N.; Siepmann, T.; Passauer, J.; Hugo, C.; Wintermann, G.; et al. Post COVID and Apheresis—Where are we Standing? Horm. Metab. Res. 2022, 54, 715–720. [Google Scholar] [CrossRef]
- Achleitner, M.; Steenblock, C.; Danhardt, J.; Jarzebska, N.; Kardashi, R.; Kanczkowski, W.; Straube, R.; Rodionov, R.N.; Bornstein, N.; Tselmin, S.; et al. Clinical improvement of Long-COVID is associated with reduction in autoantibodies, lipids, and inflammation following therapeutic apheresis. Mol. Psychiatry, 2023; online ahead of print. [Google Scholar] [CrossRef]
- Hoffmann, C.; Gerber, P.A.; Cavelti-Weder, C.; Licht, L.; Kotb, R.; Al Dweik, R.; Cherfane, M.; Bornstein, S.R.; Perakakis, N. Liver, NAFLD and COVID-19. Horm. Metab. Res. 2022, 54, 522–531. [Google Scholar] [CrossRef]
- Loffredo, L.; Violi, F. COVID-19 and cardiovascular injury: A role for oxidative stress and antioxidant treatment? Int. J. Cardiol. 2020, 312, 136. [Google Scholar] [CrossRef]
- Violi, F.; Oliva, A.; Cangemi, R.; Ceccarelli, G.; Pignatelli, P.; Carnevale, R.; Cammisotto, V.; Lichtner, M.; Alessandri, F.; De Angelis, M.; et al. Nox2 activation in COVID-19. Redox Biol. 2020, 36, 101655. [Google Scholar] [CrossRef]
- Taylor, J.P.; Tse, H.M. The role of NADPH oxidases in infectious and inflammatory diseases. Redox Biol. 2021, 48, 102159. [Google Scholar] [CrossRef]
- Youn, J.Y.; Zhang, Y.; Wu, Y.; Cannesson, M.; Cai, H. Therapeutic application of estrogen for COVID-19: Attenuation of SARS-CoV-2 spike protein and IL-6 stimulated, ACE2-dependent NOX2 activation, ROS production and MCP-1 upregulation in endothelial cells. Redox Biol. 2021, 46, 102099. [Google Scholar] [CrossRef]
- Jiang, Z.; Wu, L.; van der Leeden, B.; van Rossum, A.C.; Niessen, H.W.M.; Krijnen, P.A.J. NOX2 and NOX5 are increased in cardiac microvascular endothelium of deceased COVID-19 patients. Int. J. Cardiol. 2023, 370, 454–462. [Google Scholar] [CrossRef]
- Ciacci, P.; Paraninfi, A.; Orlando, F.; Rella, S.; Maggio, E.; Oliva, A.; Cangemi, R.; Carnevale, R.; Bartimoccia, S.; Cammisotto, V.; et al. Endothelial dysfunction, oxidative stress and low-grade endotoxemia in COVID-19 patients hospitalised in medical wards. Microvasc. Res. 2023, 149, 104557. [Google Scholar] [CrossRef]
- Damiano, S.; Sozio, C.; La Rosa, G.; Santillo, M. NOX-Dependent Signaling Dysregulation in Severe COVID-19: Clues to Effective Treatments. Front. Cell Infect. Microbiol. 2020, 10, 608435. [Google Scholar] [CrossRef]
- Shiravi, A.A.; Saadatkish, M.; Abdollahi, Z.; Miar, P.; Khanahmad, H.; Zeinalian, M. Vitamin D can be effective on the prevention of COVID-19 complications: A narrative review on molecular aspects. Int. J. Vitam. Nutr. Res. 2022, 92, 134–146. [Google Scholar] [CrossRef]
- Shojaeefar, E.; Malih, N.; Rezaei, N. The possible double-edged sword effects of vitamin D on COVID-19: A hypothesis. Cell Biol. Int. 2021, 45, 54–57. [Google Scholar] [CrossRef]
- Yousefian, M.; Shakour, N.; Hosseinzadeh, H.; Hayes, A.W.; Hadizadeh, F.; Karimi, G. The natural phenolic compounds as modulators of NADPH oxidases in hypertension. Phytomedicine 2019, 55, 200–213. [Google Scholar] [CrossRef]
- Youn, J.Y.; Wang, J.; Li, Q.; Huang, K.; Cai, H. Robust therapeutic effects on COVID-19 of novel small molecules: Alleviation of SARS-CoV-2 S protein induction of ACE2/TMPRSS2, NOX2/ROS, and MCP-1. Front. Cardiovasc. Med. 2022, 9, 957340. [Google Scholar] [CrossRef]
- Paul, A.K.; Hossain, M.K.; Mahboob, T.; Nissapatorn, V.; Wilairatana, P.; Jahan, R.; Jannat, K.; Bondhon, T.A.; Hasan, A.; de Lourdes Pereira, M.; et al. Does Oxidative Stress Management Help Alleviation of COVID-19 Symptoms in Patients Experiencing Diabetes? Nutrients 2022, 14, 321. [Google Scholar] [CrossRef]
- Mokra, D.; Adamcakova, J.; Mokry, J. Green Tea Polyphenol (−)-Epigallocatechin-3-Gallate (EGCG): A Time for a New Player in the Treatment of Respiratory Diseases? Antioxidants 2022, 11, 1566. [Google Scholar] [CrossRef]
- Mingrone, G.; Castagneto-Gissey, L.; Bornstein, S.R. New Horizons: Emerging Antidiabetic Medications. J. Clin. Endocrinol. Metab. 2022, 107, e4333–e4340. [Google Scholar] [CrossRef]
- Mingrone, G.; Bornstein, S.; Le Roux, C.W. Optimisation of follow-up after metabolic surgery. Lancet Diabetes Endocrinol. 2018, 6, 487–499. [Google Scholar] [CrossRef]
- Sano, M. A new class of drugs for heart failure: SGLT2 inhibitors reduce sympathetic overactivity. J. Cardiol. 2018, 71, 471–476. [Google Scholar] [CrossRef]
- Packer, M.; Anker, S.D.; Butler, J.; Filippatos, G.; Pocock, S.J.; Carson, P.; Januzzi, J.; Verma, S.; Tsutsui, H.; Brueckmann, M.; et al. Cardiovascular and Renal Outcomes with Empagliflozin in Heart Failure. N. Engl. J. Med. 2020, 383, 1413–1424. [Google Scholar] [CrossRef]
- Anker, S.D.; Butler, J.; Filippatos, G.; Ferreira, J.P.; Bocchi, E.; Bohm, M.; Brunner-La Rocca, H.P.; Choi, D.J.; Chopra, V.; Chuquiure-Valenzuela, E.; et al. Empagliflozin in Heart Failure with a Preserved Ejection Fraction. N. Engl. J. Med. 2021, 385, 1451–1461. [Google Scholar] [CrossRef]
- Wharton, S.; Blevins, T.; Connery, L.; Rosenstock, J.; Raha, S.; Liu, R.; Ma, X.; Mather, K.J.; Haupt, A.; Robins, D.; et al. Daily Oral GLP-1 Receptor Agonist Orforglipron for Adults with Obesity. N. Engl. J. Med. 2023; online ahead of print. [Google Scholar] [CrossRef]
- Jastreboff, A.M.; Kaplan, L.M.; Frias, J.P.; Wu, Q.; Du, Y.; Gurbuz, S.; Coskun, T.; Haupt, A.; Milicevic, Z.; Hartman, M.L.; et al. Triple-Hormone-Receptor Agonist Retatrutide for Obesity—A Phase 2 Trial. N. Engl. J. Med. 2023; online ahead of print. [Google Scholar] [CrossRef]
- Bornstein, S.R.; Rubino, F.; Khunti, K.; Mingrone, G.; Hopkins, D.; Birkenfeld, A.L.; Boehm, B.; Amiel, S.; Holt, R.I.; Skyler, J.S.; et al. Practical recommendations for the management of diabetes in patients with COVID-19. Lancet Diabetes Endocrinol. 2020, 8, 546–550. [Google Scholar] [CrossRef]
- Bornstein, S.R.; Rubino, F.; Ludwig, B.; Rietzsch, H.; Schwarz, P.E.H.; Rodionov, R.N.; Khunti, K.; Hopkins, D.; Birkenfeld, A.L.; Boehm, B.; et al. Consequences of the COVID-19 pandemic for patients with metabolic diseases. Nat. Metab. 2021, 3, 289–292. [Google Scholar] [CrossRef]
- Bornstein, S.R.; Voit-Bak, K.; Donate, T.; Rodionov, R.N.; Gainetdinov, R.R.; Tselmin, S.; Kanczkowski, W.; Muller, G.M.; Achleitner, M.; Wang, J.; et al. Chronic post-COVID-19 syndrome and chronic fatigue syndrome: Is there a role for extracorporeal apheresis? Mol. Psychiatry 2022, 27, 34–37. [Google Scholar] [CrossRef]
- Mangione, C.M.; Barry, M.J.; Nicholson, W.K.; Cabana, M.; Chelmow, D.; Coker, T.R.; Davis, E.M.; Donahue, K.E.; Doubeni, C.A.; Jaén, C.R.; et al. Vitamin, Mineral, and Multivitamin Supplementation to Prevent Cardiovascular Disease and Cancer: US Preventive Services Task Force Recommendation Statement. JAMA 2022, 327, 2326–2333. [Google Scholar] [CrossRef]
- Helm, M.M.; Alaba, T.; Klimis-Zacas, D.; Izuora, K.; Basu, A. Effect of Dietary Berry Supplementation on Antioxidant Biomarkers in Adults with Cardiometabolic Risks: A Systematic Review of Clinical Trials. Antioxidants 2023, 12, 1182. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| NOX Isoform | Role in Obesity and Cardiometabolic Diseases | References |
|---|---|---|
| NOX1 | Mediates endothelial activation, inflammation and cardiac hypertrophy; promotes microvascular dysfunction in db/db mice; increases superoxide anion formation in abdominal fat-derived mesenchymal stromal cells and renal cortex; acts as a link between obesity and kidney injury. | [36,37,38,39,40] |
| NOX2 | Higher expression in obese patients; promotes endothelial dysfunction in obese rats; NOX2 deletion reduces obesity-induced effects; NOX2 inhibition improves vascular function. | [41,42,43,44,45,46,47,48] |
| NOX4 | Anti-atherosclerotic, vaso- and cardioprotective role; involved in insulin-induced adipocyte differentiation; NOX4 inhibition reduces adipocyte differentiation and ROS generation; adipocyte-specific deletion delays diet-induced inflammation and insulin resistance; deletion causes adipose tissue accumulation and insulin resistance by high-fat diet. | [49,50,51,52,53,54,55,56,57,58] |
| NOX5 | Elevated NOX5 expression and activity in coronary arteries of patients with coronary artery disease; limited knowledge about role in obesity, but reduced body weight gain and adipose tissue mass in humanized NOX5 knock-in mice suggest an additional protective role in obesity. | [59,60,61,62] |
| Therapeutic Approach | Targeted Conditions/Effects | References |
|---|---|---|
| Glucagon-like peptide-1 receptor agonists (GLP-1 RAs) and GLP-1/gastric inhibitory polypeptide dual receptor antagonists | Hyperglycemia, insulin resistance, obesity | [130,131,135,136] |
| Sodium-glucose cotransporter 2 (SGLT2) inhibitors | Type 2 diabetes, hyperglycemia, diabetic nephropathy, cardiovascular diseases, heart failure with reduced ejection fraction | [130,132,133,134] |
| COVID-19 management guidelines | Guidance for diabetes management during the pandemic, post-pandemic complications, and risk reduction for SARS-CoV-2 infections | [30,137] |
| Extracorporeal apheresis | Potential therapeutic option for patients with COVID-19 and post-COVID-19 syndrome, targeting inflammatory and autoimmune pathogenesis by reducing proinflammatory mediators and ß-adrenergic and muscarinic receptor autoantibodies | [31,113,139] |
| NOX inhibitors and antioxidants | Selective inhibitors of NOX1 and 2 and antioxidants could reduce superoxide anion formation, oxidative stress, increase NO availability and reduce the deleterious oxidative modification of biomolecules | [6,16,17,22,28] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morawietz, H.; Brendel, H.; Diaba-Nuhoho, P.; Catar, R.; Perakakis, N.; Wolfrum, C.; Bornstein, S.R. Cross-Talk of NADPH Oxidases and Inflammation in Obesity. Antioxidants 2023, 12, 1589. https://doi.org/10.3390/antiox12081589
Morawietz H, Brendel H, Diaba-Nuhoho P, Catar R, Perakakis N, Wolfrum C, Bornstein SR. Cross-Talk of NADPH Oxidases and Inflammation in Obesity. Antioxidants. 2023; 12(8):1589. https://doi.org/10.3390/antiox12081589
Chicago/Turabian StyleMorawietz, Henning, Heike Brendel, Patrick Diaba-Nuhoho, Rusan Catar, Nikolaos Perakakis, Christian Wolfrum, and Stefan R. Bornstein. 2023. "Cross-Talk of NADPH Oxidases and Inflammation in Obesity" Antioxidants 12, no. 8: 1589. https://doi.org/10.3390/antiox12081589
APA StyleMorawietz, H., Brendel, H., Diaba-Nuhoho, P., Catar, R., Perakakis, N., Wolfrum, C., & Bornstein, S. R. (2023). Cross-Talk of NADPH Oxidases and Inflammation in Obesity. Antioxidants, 12(8), 1589. https://doi.org/10.3390/antiox12081589