Abstract
Agriculture impacts both human welfare and biodiversity at the same time. Still, social and ecological assessments have commonly analyzed the relevance of agroecosystems separately. We evaluated the human and avian feeding use of the biocultural landscape in Jardín, Colombia, using a socioecological approach. Together with farmers, we identified the main socioecological units of the landscape (i.e., fincas, grazing lands, town, forests) and determined the use of each unit in terms of food foraging from forests, crop cultivation, cattle grazing, food commercialization, and food industrialization. We compared the richness of the food resources produced among finca sections (i.e., gardens, coffee–banana plantations, grazing lands). Then, we surveyed avian behavior to contrast the richness of bird species, feeding use and intensity, and food types consumed by birds among the units. Fincas were shown to play a pivotal role in feeding both humans and birds. Gardens provide food for people as well as nectarivore and frugivore birds. Coffee–banana plantations are economically relevant, but their food provision is limited and could be enhanced by increasing the diversity of the food crops within them. The town supports commerce and granivorous birds, whereas grazing lands have limited feeding importance. Forests are used by birds to capture invertebrates but do not supply much food for the people. Our approach fosters the identification of key socioecological units, demonstrating that studying both humans and wildlife enhances the comprehension of biocultural landscapes.
1. Introduction
For thousands of years, food production systems, including agriculture, have been the most important activities for the nourishment, development, and welfare of human societies [1,2,3]. At present, 38% of the Earth’s surface is covered with agricultural lands, nearly 25% of it in the Americas [4]. More than 60 million people across Latin America are small farmers and rural families who perform agricultural practices that sustain the population of the region [5]. Agroecosystems throughout this region are tightly linked to the biological, economical, historical, and cultural heritage of rural communities [6], which converge in the biocultural landscape [7]; moreover, they exert a significant impact on biodiversity [5]. The effects of community-based agriculture on wildlife are variable in Latin America; whereas some productive systems (e.g., agroforestry systems) might maintain or increase diversity compared to natural ecosystems [8,9,10,11], others (e.g., monocultures) may represent a serious threat to the conservation of biodiversity [12,13]. Still, social and ecological research on the agriculture–wildlife relationship in Latin American biocultural landscapes is scarce, limiting our ability to document, understand, compare, and manage farming activities [6,14]. Given that biocultural landscapes may be barely understood by separate evaluations of their cultural and ecological components [15,16], it is imperative to promote their study from a comprehensive socioecological approach.
Ethological research fosters ecological and evolutionary knowledge of animals, given that behavior determines survival and adaptation processes [17]. Because the first response of animals to anthropogenic disturbances might be associated with behavior [18], studies about ethology are needed to understand the effects of productive practices on wildlife and guide biodiversity conservation initiatives. In this context, analyzing feeding habits might be particularly useful, as this behavior is crucial for the survival of any zoological group and determines several aspects of their life history. Although research on feeding imposes methodological constraints regarding animal groups that exhibit cryptic habits, birds are ideal study models because they display a vast array of foraging habits and preferences, adjust their feeding patterns to anthropogenic disturbances, and are relatively easy to survey [19]. Moreover, birds are usually strongly bonded to rural communities, which provide provisioning, regulating, cultural, and support services within biocultural landscapes [20].
To our knowledge, there are no studies that integrate ethological and social research working with analytical scopes and processes for evaluating the feeding relevance of human-dominated landscapes in Latin America. Thus, in this study we analyzed the feeding use of a biocultural landscape in the Colombian Andes by humans and birds with a socioecological approach. First, together with people from Jardín, Antioquia, we identified the main socioecological units of the biocultural landscape. Then, we determined the human feeding use of these units in terms of forest, crop cultivation, cattle grazing, food commercialization, and food industrialization. Furthermore, we estimated the richness of food resources produced in the fincas, which are small farms generally composed of a garden, a house, a coffee–banana plantation, and occasionally a small plot for cattle grazing.
We expected that higher proportions of human use would be indicative of greater social relevance of specific socioecological units in terms of food production and distribution. Consequently, we anticipated a greater presence of crop cultivation areas within fincas, increased proportions of cattle feeding on grazing lands, and low values of human feeding use of forests. As urban settlements usually function as centers of trade and interchange [21], we supposed that commercial and industrial activities would occur in the town. Given that previous studies found that gardens have a pivotal land-use function for local food production [22], we presumed that we would find an important number of food resources cultivated in such units.
Along with our social analysis, we surveyed bird behavior to compare the proportions of feeding use by the avian community within the socioecological units of the biocultural landscape. We also estimated the number of feeding bird species, the intensity of avian feeding, and the food types consumed by birds across the study site. We expected to record a higher proportion of bird feeding use, a larger number of species, higher feeding intensity, and a greater frequency of food types consumed by birds in the socioecological units that benefit their different guilds. As suggested by previous studies [23,24], we anticipated that forests would satisfy the feeding needs of the greatest number of bird species, mainly those consuming invertebrates. We expected that gardens would provide important feeding resources for frugivorous and nectarivorous birds [25,26], whereas coffee–banana plantations and grazing lands would benefit insectivores [27,28]. We predicted that the town would feed a lower number of species but a higher number of individuals of those species, mainly granivorous, as shown by other studies in the region [26]. Finally, we intended to identify the socioecological units that were key for both human and bird feeding across the biocultural landscape. The relevance was expected to be evident through high levels of feeding use by and food provision for the rural population of Jardín, together with important levels of foraging use by several bird species that consume different food types.
Our research highlights the productive relevance of the biocultural landscape to local rural communities, as well as its influence on the feeding behavior of birds. Given that both humans and wild animals depend on the food supplied by socioecological units, it is important to perform this type of integral assessment in order to gain comprehensive knowledge about biocultural landscapes, maintaining biological and cultural diversity, and sustainable management of farming lands. Moreover, our evaluation fosters the identification of key socioecological units that meet the feeding needs of both rural communities and wildlife.
2. Materials and Methods
2.1. Study Landscape
We performed our research in Jardín, at the Department of Antioquia, Colombia. The site is located in the western range of the Colombian Andes at an altitude of ~1700 m. It comprises a biocultural landscape that is culturally relevant because of local agricultural traditions, which are mainly associated with coffee production [29,30]. In addition, the site is embedded within a richly biodiverse region that is widely recognized for its bird and plant diversity [31]. The studied biocultural landscape can be broadly divided into a rural area and an urban area. The rural area (~19,991.64 ha) [32] is inhabited by farmers who mainly produce coffee, bananas, and cattle products for commercialization. The urban area (~98.6 ha) [32] is highly commercial and residential, and offers well-organized and serious ecotourism (e.g., sightseeing, birdwatching, hiking) and agritourism activities. The region includes a lower montane wet forest zone [33], with temperatures ranging from 12–18 °C and rainfall of 2000–4000 mm per year [34]. Large patches of original forests are no longer found across the biocultural landscape because of anthropogenic transformation [35].
2.2. Ethnographic Research
Due to COVID-19 social restrictions, we avoided working with groups of people to gather information for our research. Instead, we utilized unstructured interviews and participant observations to learn about the local feeding use of the biocultural landscape. Such qualitative methods might require a reduced sample size because they demand skill, care, and time to interpret the information obtained, but in turn they can produce meaningful, representative, locally contextualized, and high-quality results [36]. Moreover, both methods enabled us to identify research ambiguities, revise the authority of respondents regarding the study topic, perform a critical evaluation of the gathered evidence, and gain a deeper understanding of the human feeding use of the landscape [36].
We interacted with 25 community members in the rural area of Jardín during September 2021. The main topics of the conversations were the general territorial use and the feeding use of the biocultural landscape by the community, as well as the food resources produced within the fincas. First, we identified the main socioecological units of the biocultural landscape, which are represented by areas of the territory set aside for specific uses by the community [37], including the following: fincas, which are generally composed of a garden, a house, and a coffee–banana plantation, and occasionally a small plot for cattle grazing; grazing lands, representing large plots of induced grasslands exclusively used for pasture feeding of cows; the town, referring to the urban area of Jardín; and forests, encompassing small, disperse, and scarce patches of secondary vegetation found on abandoned coffee plantations or grazing lands, forest remnants, and riparian habitats (Figure 1).

Figure 1.
Distribution of point-count stations where we surveyed feeding birds across the biocultural landscape in Jardín, Antioquia. Location of study site shown on a map of Colombia. Geometric shapes indicate point-count stations based on evaluated socioecological units: triangles—gardens; circles—coffee–banana plantations; stars—town; diamonds—forests; squares—grazing lands. Gardens were often covered with lawn and ornamental flowering herbs, short fruiting trees, and bird feeders. Coffee–banana plantations were dominated by short banana trees and coffee shrubs. Town largely lacked vegetation and was mostly covered with pavement and small buildings. Tree and shrub strata were well developed in forests, with different plant species present. Grazing lands represent plots of induced grasslands with occasional guava trees and an absence of shrubs.
2.3. Human Feeding Uses of Socioecological Units
We identified four general categories of activities related to food production and distribution that people performed in each socioecological unit of the biocultural landscape: crop cultivation, cattle grazing, food commercialization, and food industrialization. With this information, we determined the human feeding use of each socioecological unit by modeling the proportion (θ) of times that each type of activity was performed in fincas, grazing lands, the town, and forests, as compared to the total number of recorded activities for the same socioecological unit. We performed this analysis with a hierarchical binomial model [38]
where x represents the number of times an activity was observed in socioecological unit i, and n is the total number of activities observed for the same socioecological unit. θ was initially modeled by using an uninformative beta prior with a = b = 1. The estimated proportions were compared across socioecological units to estimate the difference in probability of each activity occurring in each unit. The probability of a given activity was significantly different between two socioecological units when 95% of the highest density intervals of the posterior distribution of the estimated difference excluded 0. We used Markov chain Monte Carlo with three Markov chains and 5000 iterations to perform Bayesian inference; the R-hat statistic was employed to assess convergence [39]. Given that using just one Markov chain would have limited us to determine if posterior distributions were being sampled appropriately, we used multiple Markov chains to identify convergence. Still, we avoided using more than three chains to reduce computation times. Analyses were performed with the Bayesian First Aid package [40], which was implemented with JAGS [41] in the R statistical programming language [42]. JAGS evaluates Bayesian graphical models with a Gibbs sampler [41].
xi ∼ binomial (θi, ni)
2.4. Richness of Food Resources Produced in the Fincas
We interrogated community members about the edible plants and animals that they produced and/or obtained within their fincas. Additionally, we determined the particular sections of fincas (i.e., coffee–banana plantations, grazing lands, gardens) where these food resources were harvested. We used the incidence frequency of food resources produced in each section of the fincas to estimate and compare the richness of food resources among sections with the “iNext” package [43]. Specifically, we computed sample-size-based rarefaction (interpolated estimation) and extrapolation (predictive estimation) sampling curves for food resource richness, which allowed us to quantify and compare richness among finca sections with a unified standardization method [43].
2.5. Bird Surveys
We surveyed birds at 147 unlimited-radius point-count stations during the same period that we performed the ethnographic research (September 2021). All stations were examined once. We decided to use unlimited-radius point-count stations in order to increase the amount of bird behavioral data. Most of the records were gathered within a confined area, as we performed detailed observations regarding avian activity. This was evidenced by the average distance to all observed birds (23.6 m), which was measured in the field using a rangefinder. Additionally, records associated with each socioecological unit were collected from a mean distance of around 30 m or less (grazing lands: 30.2 m; town: 28.5 m; coffee–banana plantations: 24.5 m; forests: 18.1 m; gardens: 16.9 m). The stations were located in the socioecological units where food was produced (coffee–banana plantations, gardens, and grazing lands), as well as in the town and forests for comparison purposes. The number of point-count stations per socioecological unit was decided according to the area they covered across the biocultural landscape. In this way, we surveyed 47 stations in coffee–banana plantations, 31 in grazing lands, 30 in gardens, 26 in forests, and 13 in the town (Figure 1). Point-count stations were separated from each other by at least 200 m. Because most of our bird observations were collected at distances closer than 50 m (93% of the records), the separation between stations was satisfactory to provide independent bird data [44,45]. We determined the locations of stations with the aid of the Global Positioning System.
Surveys consisted of visually identifying and counting the number of bird individuals present at each station during a 10 min period of observation. For each bird, we recorded the behavior it displayed and the food type it consumed. The kind of behavior was determined during the first 5 s after detection, in order to avoid an observer’s effect and autocorrelation among the records [46,47]. Observed behavior was classified as feeding—when birds were actively foraging or consuming food, or other—when birds were performing any other activity than feeding. We categorized the food types consumed by birds as invertebrates, fruits, nectar, seeds, vertebrates, and other plant material (e.g., leaves, roots, sprouts, flowers). Surveys were performed during the 4 h after daybreak to cover peak bird activity.
2.6. Avian Feeding Use of the Biocultural Landscape
We modeled avian feeding use of the biocultural landscape by following the same approach we used to assess human feeding use of the socioecological units. In this case, we evaluated avian feeding use by modeling the proportion (θ) of times that birds fed in coffee–banana plantations, grazing lands, gardens, forests, and the town, compared to the total number of recorded behaviors for the same socioecological unit. The estimated proportions were compared across socioecological units to estimate the difference in probability of feeding occurring in each unit.
2.7. Bird Species Richness and Feeding Intensity
We estimated and compared the number of bird species that fed in coffee–banana plantations, grazing lands, gardens, forests, and the town, by following the same procedure that we used to estimate the richness of food resources produced in the fincas. However, in this case, we used the observed abundance of species that fed in each socioecological unit to estimate species richness [43]. We evaluated bird feeding intensity across the biocultural landscape by using ANOVA to compare the average number of feeding birds in coffee–banana plantations, grazing lands, gardens, forests, and the town. Significant statistical differences in bird feeding intensity among socioecological units were assumed when the 95% confidence intervals did not overlap.
2.8. Food Types Consumed by Birds
We modeled food types consumed by birds through the same approach that we used to assess the human feeding use of socioecological units. We analyzed the food types that birds consumed across the landscape by modeling the proportion (θ) of times that birds fed on each type (invertebrates, fruits, nectar, and seeds) in coffee–banana plantations, grazing lands, gardens, forests, and the town, compared to the total number of types of food ingested for the same unit. The estimated proportions were compared across socioecological units to estimate the difference in probability of the food types consumed by the birds in each unit. Appendix A depicts a flowchart that summarizes the methodology that we used throughout this research.
3. Results
3.1. Human Feeding Use of Socioecological Units
Food production and distribution varied among the socioecological units of the biocultural landscape (Table 1, Figure 2). Fincas were mainly allocated to the cultivation of human food resources, followed by grazing lands (Figure 2a). Grazing activities were heavily performed within grazing lands, but some fincas were also used for cattle ranging (Figure 2b). The town was a pivotal unit for the commercialization of food resources, followed by fincas and grazing lands (Figure 2c). Industrialization activities were scarce throughout the biocultural landscape, but were present in grazing lands, fincas, and the town (Figure 2d). Forests were not reported to be used for any feeding activity by local residents. This might be influenced by the fact that hunting is forbidden by local authorities, and some wild fruits consumed by residents are maintained in home gardens, where they occasionally obtain these products.

Table 1.
Number of occasions when activities related to food production and distribution were performed in socioecological units of biocultural landscape.
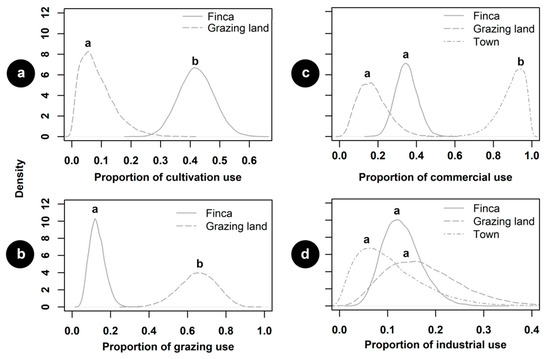
Figure 2.
Estimated proportions of human feeding use of socioecological units in biocultural landscape: (a) crop cultivation; (b) cattle grazing; (c) food commercialization; and (d) food industrialization. Letters above curves indicate statistical differences between units. The probability of a given use was significantly different between two socioecological units when 95% of the highest density intervals of the posterior distribution of the estimated difference excluded 0.
3.2. Richness of Food Resources Produced in the Fincas
We recorded a total of 50 food resources that were produced within the fincas (Appendix B). According to our estimations, the richness of food resources peaked in gardens (Figure 3, Table S1). Coffee–banana plantations and grazing lands had similar food richness values, and estimates for these units were significantly lower than those for gardens (Figure 3, Table S1).
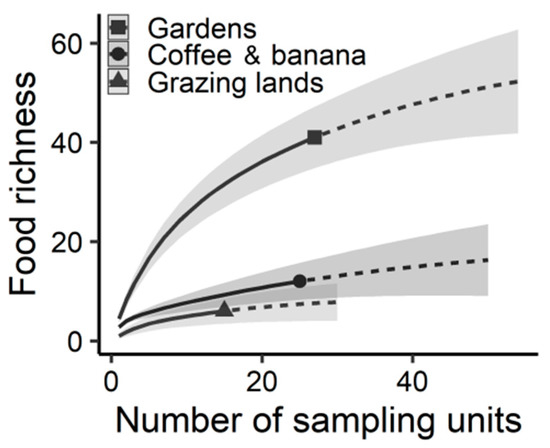
Figure 3.
Estimated richness of food resources produced by farmers of Jardín in different sections of their fincas. Food resources include edible plants and animals. Interpolated estimations are indicated by continuous lines, extrapolated estimates by dotted lines, and 95% confidence intervals by shaded areas.
3.3. Avian Feeding Use of the Biocultural Landscape
Most events of bird feeding were recorded in coffee–banana plantations, followed by gardens, the town, forests, and grazing lands (Appendix C). The proportion of bird feeding varied among the socioecological units of the biocultural landscape (Figure 4). The main patterns suggest that gardens, the town, and forests were more used by foraging birds than grazing lands. Moreover, forests exhibited higher bird feeding proportions than coffee–banana plantations.
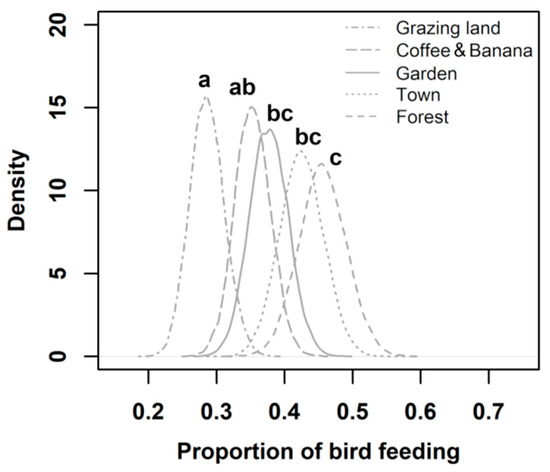
Figure 4.
Estimated proportions of avian feeding use in grazing lands, coffee–banana plantations, gardens, the town, and forests. Letters above curves indicate statistical differences between units. The probability of bird feeding was significantly different between two socioecological units when 95% of the highest density intervals of the posterior distribution of the estimated difference excluded 0.
3.4. Bird Species Richness and Feeding Intensity
We recorded a total of 89 avian species foraging across the biocultural landscape (Appendix D). In our analysis, we estimated that the species richness of feeding birds was similar between gardens, grazing lands, coffee–banana plantations, and forests (Figure 5, Table S2). The town was markedly different to the rest of the socioecological units, containing the lowest estimated species richness (Table S2). Avian feeding intensity was also similar throughout the biocultural landscape (Figure 6). Differences in the number of feeding birds were only evident between coffee–banana plantations and the town, with the former having lower values than the latter.
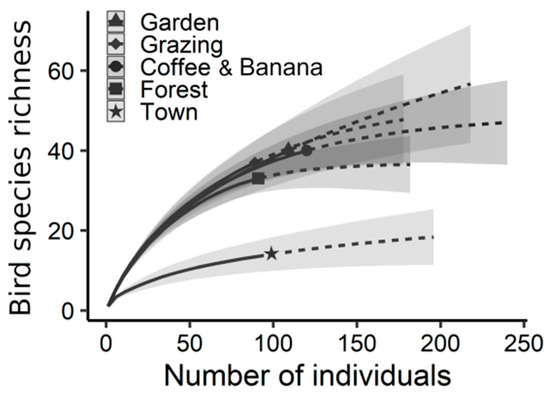
Figure 5.
Bird species richness estimated in gardens, grazing lands, coffee–banana plantations, forests, and the town. Interpolated estimations are indicated by continuous lines, extrapolated estimations by dotted lines, and 95% confidence intervals by shaded areas.
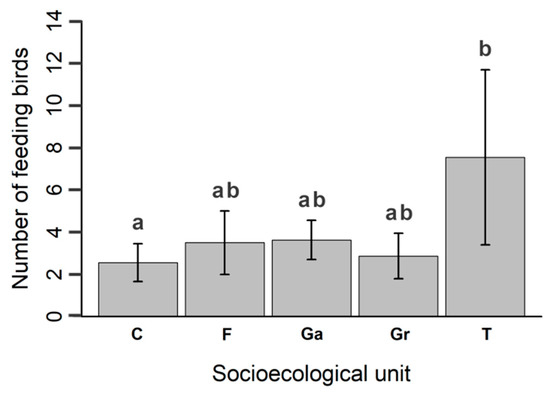
Figure 6.
Bird feeding intensity of socioecological units, represented by average number of feeding birds in coffee–banana plantations (C), forests (F), gardens (Ga), grazing lands (Gr), and the town (T). Letters above bars depict statistical differences between units.
3.5. Food Type Consumed by Birds
Most of the recorded birds were feeding on invertebrates, followed by fruits, nectar, and seeds (Appendix C). We observed few birds foraging on vertebrates (1 individual) and other plant materials (2 individuals); thus, we were not able to consider these types of food consumed by birds in our estimations. The proportion of invertebrates consumed by birds was greater in forests than in coffee–banana plantations, the town, and gardens (Figure 7a). Birds foraged proportionally more on fruit in gardens than in forests, grazing lands, and the town (Figure 7b). The proportion of nectar consumed by birds was higher in gardens than in forests and the town (Figure 7c). Finally, birds ate seeds proportionally more in the town than in the gardens and grazing lands (Figure 7d).
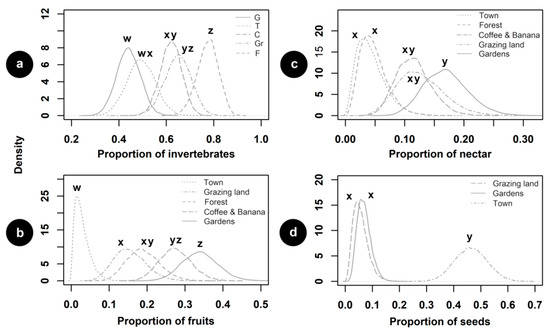
Figure 7.
Estimated proportions of food types consumed by birds in socioecological units of biocultural landscape: (a) invertebrates; (b) fruits; (c) nectar; and (d) seeds. Letters above curves indicate statistical differences between units. G—gardens; T—the town; C—coffee–banana plantations; Gr—grazing lands; F—forests.
4. Discussion
4.1. Human Feeding Use of Socioecological Units
As expected, we observed that fincas were a fundamental unit across the biocultural landscape in terms of human feeding. Most food was cultivated there, not only for local consumption, but also for exportation (e.g., coffee, purple passion fruit Passiflora pinnatistipula) [29,48]. Hence, fincas play a key role in the food security of the community [49] and simultaneously provide incomes for local families, which they invest in non-food resources (e.g., textiles, medicines, construction assets) and daily food items that are not produced within fincas (e.g., rice, chocolate).
Although we supposed that the town would be the main commercial unit where local and foreign people trade food, we noted that the finca was also an important socioecological unit for the exchange of cultivated food (e.g., banana, coffee, avocado) and industrialized food (e.g., cheese, unrefined brown sugar). Additionally, fincas play a role in cattle ranging, because some families still rear cows for their milk, even when this tradition has been replaced by extending the land destined for coffee and banana plantations. Despite grazing lands contributing to cultivation purposes, most of the food items grown there were intended for feeding cattle (i.e., guava), and very little for the direct feeding of local families. Forests were evidently not used as feeding units by the community, given that old poaching practices have been banned and are punishable by local regulations. Moreover, we were not able to detect any tradition of gathering edible plants in forests by the community, as food is mainly cultivated, grown, or purchased across this biocultural landscape.
4.2. Richness of Food Resources Produced in the Fincas
The estimated richness of food resources, including both edible plants and animals, varied among the finca sections. As described by other studies [22], gardens play a key role in producing different food types and are mainly allocated to nourishing local families. An increased variety in food resources might aid the local population by enhancing the diversification of their diet [50]. Moreover, expanding the type of food resources that they produce in gardens would have a positive impact on the farmers’ economy because they would avoid having to buy such items, whereas the surplus in production could be sold to generate extra revenue [51]. In sum, gardens not only provide leisure services to the local population [52], but also have a tangible benefit in terms of their diet and economy [50].
The estimated food richness for coffee–banana plantations and grazing lands was low. Many of the food items produced in these socioecological units are intended for sale (e.g., coffee, bananas, avocados, milk). Thus, their production might not be regulated by local feeding needs, but by external pressure from buyers, international markets, commercial regulations, and consumers [53]. In this way, food produced within coffee–banana plantations and grazing lands have a fundamental positive effect on the economy of local families but might have a reduced direct impact on their diet.
4.3. Avian Feeding Use of the Biocultural Landscape
Birds exhibited a higher proportion of feeding activity within forests, the town, and gardens, than in grazing lands. Although forests have been severely fragmented and disturbed across the biocultural landscape, they could still serve as key avian habitats [54,55], particularly for foraging [56]. In contrast, the town and gardens function as important anthropogenic feeding grounds for the local avifauna. Urban settlements are known to indirectly provide feeding resources (e.g., food spills) for those birds that tolerate high disturbance levels [57,58], whereas humans directly supply food for birds within gardens as a leisure activity [59].
Grazing lands provided fewer avian feeding opportunities than other socioecological units. These lands might have a reduced avian feeding proportion given that they represent simplified habitats that are mainly covered with grasses, which reduces the availability of feeding substrates and resources for birds [60,61,62]. Furthermore, coffee–banana plantations exhibited lower bird feeding proportions than forests. This pattern might be related to the poor microhabitat heterogeneity of local plantations [63,64], given that their shrub strata were usually occupied by a unique plant species (i.e., coffee), whereas the tree strata were typically dominated by bananas.
4.4. Bird Species Richness and Feeding Intensity
Gardens, grazing lands, coffee–banana plantations, and forests supported similar estimated bird species richness. This result was unexpected, because forests usually encompass a greater number of bird species than anthropogenic habitats [63,65]. We suspect that high disturbance levels and the lack of connectivity among wooded patches might have led to a decrease in the ecological conditions of the forests, reducing the number of bird species that are able to feed within them [66,67]. At the same time, it may be plausible that a homogenization process is occurring across the biocultural landscape, driven by anthropogenic disturbances [68], suggesting that similar numbers of bird species feed among the different socioecological units. The processes of ecological filtering and homogenization operating on the avian community might be more evident for the town, given that this unit exhibited the least bird species compared to other urban areas of Latin America [26]. Further research about the composition of the avian community might help to confirm such processes [67], as well as identify vulnerable groups of birds that are being excluded from feeding across the biocultural landscape, such as forest specialist species [69].
According to our feeding intensity analysis, the number of foraging birds differed only between coffee–banana plantations and the town. We presumed that the town would support a higher number of feeding birds, given that there is usually high predictability in terms of food availability in urban areas [70], which could be exploited by a reduced number of flocking species, including exotic birds (e.g., rock pigeon) [71] and aerial insectivores (e.g., swallows) [72]. The coffee–banana plantations had reduced vegetation complexity relative to their structure and composition, which might lead to fewer feeding opportunities, thus restricting the number of feeding birds [62,73].
4.5. Food Types Consumed by Birds
The number of feeding events suggested that birds intensively foraged on invertebrates across the biocultural landscape. Invertebrates are key resources for birds, which represent a source of nutrients that impact on breeding, fitness-related traits, diet quality, and offspring development [74,75]. In fact, many birds feed on invertebrates even if they specialize in consuming other types of food (e.g., nectar, fruit) [76,77]. As expected, we observed that invertebrates were particularly ingested within forests, possibly because forests provide adequate microhabitats that increase the diversity and availability of such animals [78,79,80]. Moreover, the negative effect of pesticides on invertebrates might be lower in forests than in gardens and plantations. Our analyses revealed similar proportions of invertebrate consumption between forests and grazing lands; however, aerial insectivores that forage in large flocks (e.g., swallows) might bias such similarity, as these birds are heavily associated with grassy open habitats [27].
Gardens were found to be an important source of fruits and nectar. In these socioecological units, fruits that are cultivated for human use can also be available for birds. Furthermore, as in other regions of the world, local people enjoy providing supplementary feeding for wild birds by maintaining feeders [59]. Specifically, bananas produced within fincas were mainly available for birds, providing access to fruit resources that might be scarce in other units, including the town. Ornamental plants, which were abundant within local gardens, might be fundamental for nectarivorous birds [25], as we noted that during the season when we conducted the study, the presence of flowers was rare in forests and the town. Our results support previous research highlighting the importance of urban settlements for granivorous birds [26,56], given that seeds were mostly consumed in the town. Seeds produced by grasses and herbs [81,82], as well as crops (e.g., rice) that local people provide for urban bird species (e.g., rock pigeon), might be responsible for such a pattern.
5. Conclusions
Our results demonstrate that fincas play a pivotal role in the feeding welfare of both humans and birds across the biocultural landscape. They represent multi-purpose socioecological units that impact the local economy, human nutrition, bird foraging activities, society–nature relationships, local identities, and cultural preservation. In particular, gardens are notable sections for the fincas because they are direct and heterogeneous sources of nourishment for the rural population of Jardín, representing the first line of protection against food insecurity and dependency. For example, we observed more than 40 types of food resources produced by farmers in the gardens of the biocultural landscape. Additionally, gardens provide fruit and nectar for birds, whereas these foraging items might be less predictable and abundant in natural habitats [83,84]. The close human–avian relationship might be particularly evident in gardens, as that is where the food that feeds both of them is cultivated, while the human pastime of bird feeding strengthens this bond. As a result, gardens might be seen as key units for feeding the population and avifauna of Jardín and for tailoring local conservation actions.
Although coffee–banana plantations are important for the economy of families, their feeding relevance could be enhanced by increasing the diversity of the food that is cultivated within them. For instance, in these particular sections of the fincas, we observed about 12 types of food resources produced by local farmers. Such a strategy could support the diversification of feeding resources of both local residents and birds. Among all of the socioecological units, grazing lands were shown to have a poor human and avian feeding relationship. To give an example, we only detected 6 types of food resources produced by farmers in such units. Promoting silvopastoral management within grazing lands could improve their positive effect on human and bird communities. Even though the feeding importance of forests was not as evident as that of gardens, we believe that the former deserves particular attention because they are heavily used by birds as foraging grounds, especially for capturing invertebrates. Invertebrates are indispensable food items for birds [74,75], and the avian regulation of their populations is fundamental for humans in order to avoid potential damage to crops [85]. Moreover, forests represent attractive socioecological units for carrying out birdwatching activities, which could increase local interest in biological conservation and could provide extra revenue to reduce the local dependency on coffee production. This recreational use of the landscape is present in Jardín, but it is usually offered by few people and for observing target birds (e.g., Andean cock-of-the-rock, Rupicola peruvianus), reducing the site’s potential in terms of bird diversity and touristic popularity.
We showed that studying both humans and wildlife promotes a more comprehensive understanding of biocultural landscapes, particularly when evaluating common vital needs, such as feeding. Additionally, our socioecological approach enabled us to identify key units for nourishing birds and the population of Jardín, including gardens. Making a positive impact on sustainability, such as by increasing local residents’ appreciation of biodiversity, promoting informed decision-making on resource use, coordinating management measures, and facilitating the production–conservation dialogue [86,87,88], could be fostered by using socioecological approaches to benefit human and animal communities in this and other regions of the globe. Further research should include surveys over a broader time frame in order to detect and comprehend inter- and intra-annual variations in the patterns that we observed.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/su14084789/s1. Table S1: Estimated richness of food resources produced by farmers of Jardín in (a) coffee–banana plantations, (b) gardens, and (c) grazing lands. Table S2: Estimated richness of bird species in (a) forests, (b) coffee–banana plantations, (c) gardens, (d) grazing lands, and (e) town.
Author Contributions
Conceptualization, R.O.-Á. and A.C.; methodology, R.O.-Á. and A.C.; formal analysis, R.O.-Á.; investigation, R.O.-Á.; resources, A.C.; writing—original draft preparation, R.O.-Á. and A.C.; writing—review and editing, R.O.-Á. and A.C.; supervision, A.C.; project administration, A.C.; funding acquisition, A.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work is part of the PAPIIT, DGAPA, UNAM (IN206520), and CONACYT (A1-S-14306) projects, Mexico, which provided financial support. R.O.-Á gratefully acknowledges the support from the DGAPA-UNAM Postdoctoral Fellowship 2021–2022.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We are indebted to all the community members of Jardín who made this study possible, in particular Mary Luz Sánchez, Juan Castaño, Carolina Marín, Angela Gallego, Alejandro Vázquez, Teresita, familia Marín, familia Vázquez, familia Vargas, and familia Gallego. We thank Fernando Cediel, José Castaño, Alejandro Suárez, and four anonymous reviewers for useful discussions and comments that enriched the content of the manuscript. We appreciate the advice of Orin J. Robinson regarding hierarchical binomial modeling.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Graphical representation of the methodology used in our study. The research was composed by surveys on both (1) ethnographic and (2) bird aspects. We employed unstructured in-depth interviews and participant observation (1.1) to determine the human feeding use of the biocultural landscape. We identified the socioecological units of the studied landscape, estimated the richness of food resources produced by local farmers in each unit, and estimated the proportion of human feeding uses per unit (1.2). In addition, we identified four main socioecological units across the biocultural landscape, including fincas, forests, the town, and grazing lands (1.3). Sections of the fincas were represented by gardens, coffee–banana plantations, and cattle grazing plots (1.4). We identified four categories of activities related to food production and distribution, which were performed in each socioecological unit, including crop cultivation, cattle grazing, food commercialization, and food industrialization (1.5). Point-count stations (2.1) were used to gather behavioral observations and determine the avian feeding use of the biocultural landscape. Specifically, we estimated the proportion of avian feeding use, bird species richness, feeding intensity, and food type consumed by birds in each socioecological unit of the biocultural landscape (2.2). Food types consumed by birds included invertebrates, nectar, fruits, and seeds (2.3). R = estimations obtained through sample-size-based rarefaction and extrapolation sampling curves; H = estimations obtained through hierarchical binomial models.

Appendix B
Incidence frequency of food resources produced in each section of the fincas, including gardens, coffee–banana plantations, and grazing lands.
| Food Resource | Section of Fincas | ||
| Scientific Name (Common Name) | Garden | Coffee–Banana | Grazing |
| Allium schoenoprasum (Chives) | 1 | 0 | 0 |
| Allium cepa (Onion) | 5 | 0 | 0 |
| Ananas comosus (Pineapple) | 2 | 0 | 0 |
| Annona cherimola (Custard apple) | 1 | 0 | 0 |
| Arracacia xanthorrhiza (Arracacha) | 1 | 0 | 0 |
| Banana leaf | 0 | 1 | 0 |
| Capsicum sp. (Chili, variety 1) | 0 | 1 | 0 |
| Capsicum sp. (Chili, variety 2) | 0 | 1 | 0 |
| Carica papaya (Papaya) | 4 | 0 | 0 |
| Cheese | 0 | 0 | 3 |
| Chili sauce | 3 | 0 | 0 |
| Citrus aurantifolia (Key lime) | 2 | 0 | 0 |
| Citrus latifolia (Persian lime) | 3 | 0 | 0 |
| Citrus cinensis (Orange) | 6 | 0 | 0 |
| Citrus reticulata (Mandarin orange) | 6 | 0 | 0 |
| Coffea spp. (Coffee) | 2 | 20 | 0 |
| Coriandrum sativum (Cilantro) | 1 | 0 | 0 |
| Cucurbita sp. (Ahuyama) | 1 | 0 | 0 |
| Eggs | 2 | 0 | 0 |
| Eriobotrya japonica (Loquat) | 1 | 0 | 0 |
| Honey | 3 | 0 | 0 |
| Mangifera indica (Mango) | 3 | 0 | 0 |
| Manihot esculenta (Yuca) | 5 | 0 | 0 |
| Milk (goat) | 1 | 0 | 1 |
| Milk (cow) | 0 | 0 | 6 |
| Musa sp. (Banana, variety 1) | 2 | 18 | 0 |
| Musa sp. (Banana, variety 2) | 0 | 2 | 0 |
| Musa sp. (Banana, variety 3) | 0 | 1 | 0 |
| Musa sp. (Banana, variety 4) | 0 | 2 | 0 |
| Musa sp. (Banana, variety 5) | 3 | 14 | 0 |
| Passiflora ligularis (Sweet granadilla) | 0 | 0 | 1 |
| Persea americana (Avocado) | 8 | 7 | 0 |
| Phaseolus vulgaris (Bean) | 8 | 0 | 0 |
| Phaseolus sp. (String bean) | 3 | 0 | 0 |
| Pork | 2 | 0 | 0 |
| Poultry | 2 | 0 | 0 |
| Psidium sp. (Guava, variety 1) | 10 | 1 | 3 |
| Psidium sp. (Guava, variety 2) | 1 | 0 | 0 |
| Psidium sp. (Guava, variety 3) | 1 | 0 | 0 |
| Psidium sp. (Guava, variety 4) | 1 | 0 | 0 |
| Psidium sp. (Guava, variety 5) | 1 | 0 | 0 |
| Psidium sp. (Guava, variety 6) | 1 | 0 | 0 |
| Psidium sp. (Guava, variety 7) | 1 | 0 | 0 |
| Psidium sp. (Guava, variety 8) | 1 | 0 | 0 |
| Psidium sp. (Guava, variety 9) | 1 | 0 | 0 |
| Psidium sp. (Guava, variety 10) | 1 | 0 | 0 |
| Solanum tuberosum (Potato) | 1 | 0 | 0 |
| Solanum lycopersicum (Tomato) | 6 | 0 | 0 |
| Spondias mombin (Yellow mombin) | 4 | 1 | 1 |
| Zea mays (Corn) | 9 | 0 | 0 |
Appendix C
Sample sizes of bird foraging observations per socioecological unit of the biocultural landscape. The number of foraging events for the whole community and for birds that foraged on invertebrates, fruits, nectar, and seeds, are given.
| Group | Forests | Gardens | Coffee-Banana Plantations | Grazing Lands | Town | Total |
| Whole community | 91 | 109 | 120 | 89 | 98 | 507 |
| Invertebrates | 64 | 43 | 69 | 45 | 33 | 254 |
| Fruits | 15 | 33 | 30 | 10 | 1 | 89 |
| Nectar | 3 | 16 | 12 | 8 | 2 | 41 |
| Seeds | 0 | 6 | 0 | 3 | 31 | 40 |
Appendix D
Observed abundance of avian species that fed in each socioecological unit of the biocultural landscape. The nomenclature and classification of species follows that proposed by the American Ornithological Society.
| Family | Socioecological Unit | ||||
| Species | Forest | Coffee | Garden | Grazing | Town |
| Cracidae | |||||
| Ortalis columbiana | 0 | 0 | 1 | 0 | 0 |
| Columbidae | |||||
| Columba livia | 0 | 0 | 0 | 0 | 42 |
| Patagioenas cayennensis | 0 | 1 | 0 | 0 | 0 |
| Columbina talpacoti | 0 | 0 | 0 | 1 | 1 |
| Cuculidae | |||||
| Piaya cayana | 0 | 1 | 0 | 0 | 0 |
| Apodidae | |||||
| Streptoprocne zonaris | 15 | 0 | 0 | 0 | 0 |
| Trochilidae | |||||
| Phaethornis guy | 2 | 2 | 1 | 0 | 0 |
| Anthracothorax nigricollis | 0 | 0 | 1 | 0 | 0 |
| Coeligena coeligena | 1 | 0 | 0 | 0 | 0 |
| Chlorostilbon melanorhynchus | 2 | 3 | 4 | 0 | 0 |
| Saucerottia saucerottei | 0 | 1 | 3 | 0 | 2 |
| Amazilia tzacatl | 0 | 2 | 4 | 1 | 0 |
| Uranomitra franciae | 1 | 0 | 0 | 1 | 0 |
| Trochilidae sp. | 0 | 0 | 1 | 0 | 0 |
| Rallidae | |||||
| Pardirallus nigricans | 0 | 0 | 0 | 2 | 0 |
| Charadriidae | |||||
| Vanellus chilensis | 0 | 0 | 0 | 5 | 0 |
| Ardeidae | |||||
| Bubulcus ibis | 0 | 1 | 0 | 1 | 0 |
| Cathartidae | |||||
| Cathartes aura | 0 | 2 | 0 | 1 | 0 |
| Accipitridae | |||||
| Rupornis magnirostris | 0 | 0 | 0 | 1 | 0 |
| Momotidae | |||||
| Momotus aequatorialis | 2 | 1 | 2 | 0 | 0 |
| Capitonidae | |||||
| Eubucco bourcierii | 2 | 0 | 0 | 0 | 0 |
| Ramphastidae | |||||
| Aulacorhynchus haematopygus | 3 | 0 | 1 | 0 | 0 |
| Picidae | |||||
| Melanerpes formicivorus | 0 | 0 | 1 | 0 | 0 |
| Melanerpes rubricapillus | 0 | 0 | 0 | 1 | 0 |
| Dryobates fumigatus | 2 | 0 | 0 | 0 | 0 |
| Dryocopus lineatus | 1 | 0 | 1 | 0 | 0 |
| Colaptes rubiginosus | 0 | 2 | 0 | 0 | 0 |
| Psittacidae | |||||
| Pionus chalcopterus | 0 | 3 | 0 | 0 | 0 |
| Thamnophilidae | |||||
| Thamnophilus multistriatus | 1 | 0 | 0 | 0 | 0 |
| Furnariidae | |||||
| Lepidocolaptes souleyetii | 3 | 0 | 0 | 0 | 0 |
| Tyrannidae | |||||
| Mionectes oleagineus | 0 | 1 | 0 | 0 | 0 |
| Leptopogon superciliaris | 3 | 2 | 0 | 0 | 0 |
| Tolmomyias sulphurescens | 0 | 1 | 0 | 0 | 0 |
| Todirostrum cinereum | 0 | 1 | 1 | 0 | 0 |
| Zimmerius chrysops | 2 | 2 | 2 | 0 | 0 |
| Camptostoma obsoletum | 1 | 0 | 1 | 1 | 0 |
| Elaenia flavogaster | 0 | 0 | 0 | 1 | 0 |
| Serpophaga cinerea | 0 | 0 | 1 | 0 | 0 |
| Machetornis rixosa | 0 | 0 | 0 | 1 | 0 |
| Myiodynastes chrysocephalus | 0 | 1 | 0 | 0 | 0 |
| Myiozetetes cayanensis | 0 | 0 | 1 | 3 | 0 |
| Tyrannus melancholicus | 0 | 1 | 0 | 6 | 2 |
| Myiarchus sp. | 0 | 1 | 0 | 0 | 0 |
| Sayornis nigricans | 0 | 0 | 2 | 2 | 0 |
| Contopus cooperi | 1 | 0 | 0 | 0 | 0 |
| Contopus cinereus | 0 | 1 | 0 | 1 | 0 |
| Vireonidae | |||||
| Vireo leucophrys | 2 | 0 | 0 | 0 | 0 |
| Vireo chivi | 0 | 1 | 0 | 0 | 0 |
| Corvidae | |||||
| Cyanocorax affinis | 1 | 0 | 1 | 1 | 0 |
| Hirundinidae | |||||
| Pygochelidon cyanoleuca | 0 | 0 | 2 | 9 | 30 |
| Stelgidopteryx ruficollis | 0 | 8 | 9 | 2 | 0 |
| Troglodytidae | |||||
| Troglodytes aedon | 0 | 1 | 1 | 1 | 1 |
| Turdidae | |||||
| Myadestes ralloides | 2 | 0 | 0 | 0 | 0 |
| Turdus ignobilis | 0 | 6 | 1 | 0 | 0 |
| Mimidae | |||||
| Mimus gilvus | 0 | 0 | 0 | 1 | 0 |
| Fringillidae | |||||
| Euphonia laniirostris | 0 | 0 | 5 | 0 | 1 |
| Passerellidae | |||||
| Zonotrichia capensis | 0 | 6 | 4 | 1 | 4 |
| Atlapetes albinucha | 2 | 2 | 0 | 2 | 0 |
| Icteridae | |||||
| Psarocolius angustifrons | 2 | 0 | 0 | 0 | 0 |
| Icterus chrysater | 0 | 2 | 1 | 0 | 0 |
| Quiscalus lugubris | 0 | 0 | 0 | 0 | 6 |
| Quiscalus mexicanus | 0 | 0 | 0 | 0 | 1 |
| Hypopyrrhus pyrohypogaster | 0 | 0 | 1 | 0 | 0 |
| Parulidae | |||||
| Mniotilta varia | 1 | 0 | 0 | 0 | 0 |
| Setophaga ruticilla | 1 | 0 | 0 | 0 | 0 |
| Setophaga fusca | 3 | 2 | 0 | 1 | 0 |
| Setophaga petechia | 0 | 0 | 1 | 0 | 0 |
| Cardellina canadensis | 1 | 0 | 0 | 0 | 0 |
| Myioborus miniatus | 9 | 2 | 1 | 6 | 0 |
| Cardinalidae | |||||
| Piranga flava | 0 | 0 | 0 | 2 | 0 |
| Thraupidae | |||||
| Chlorophanes spiza | 3 | 0 | 1 | 0 | 0 |
| Hemithraupis guira | 0 | 0 | 1 | 0 | 0 |
| Sicalis flaveola | 0 | 0 | 0 | 5 | 0 |
| Volatinia jacarina | 0 | 2 | 0 | 0 | 0 |
| Tachyphonus rufus | 0 | 3 | 0 | 0 | 0 |
| Ramphocelus dimidiatus | 0 | 0 | 1 | 0 | 0 |
| Ramphocelus flammigerus | 0 | 6 | 6 | 2 | 0 |
| Sporophila nigricollis | 0 | 0 | 5 | 0 | 0 |
| Coereba flaveola | 0 | 3 | 5 | 1 | 1 |
| Tiaris olivaceus | 0 | 0 | 0 | 1 | 0 |
| Anisognathus somptuosus | 2 | 0 | 0 | 0 | 0 |
| Stilpnia heinei | 6 | 3 | 3 | 5 | 0 |
| Stilpnia vitriolina | 0 | 8 | 9 | 8 | 1 |
| Stilpnia cyanicollis | 0 | 4 | 4 | 2 | 0 |
| Tangara labradorides | 2 | 0 | 0 | 0 | 0 |
| Tangara gyrola | 7 | 10 | 2 | 2 | 0 |
| Tangara arthus | 3 | 3 | 0 | 2 | 0 |
| Thraupis episcopus | 0 | 8 | 5 | 4 | 3 |
| Thraupis palmarum | 2 | 10 | 12 | 2 | 3 |
References
- Zeder, M.A. The origins of agriculture in the Near East. Curr. Anthropol. 2011, 52, S221–S235. [Google Scholar] [CrossRef]
- Zeder, M.A. Core questions in domestication research. Proc. Natl. Acad. Sci. USA 2015, 112, 3191–3198. [Google Scholar] [CrossRef] [PubMed]
- Clement, C.R.; Casas, A.; Parra-Rondinel, F.A.; Levis, C.; Peroni, N.; Hanazaki, N.; Cortés-Zárraga, L.; Rangel-Landa, S.; Alves, R.P.; Ferreira, M.J.; et al. Disentangling domestication from food production systems in the Neotropics. Quaternay 2021, 4, 4. [Google Scholar] [CrossRef]
- FAO. Uso de la Tierra en la Agricultura Según las Cifras; Organización de las Naciones Unidas Para la Alimentación y la Agricultura: Santiago, Chile, 2020. [Google Scholar]
- Salcedo, S.; Guzmán, L. Agricultura Familiar en América Latina y el Caribe: Recomendaciones de Política; Organización de las Naciones Unidas para la Alimentación y la Agricultura: Santiago, Chile, 2014. [Google Scholar]
- von Wehrden, H.; Abson, D.J.; Beckmann, M.; Cord, A.F.; Klotz, S.; Seppelt, R. Realigning the land-sharing/land-sparing debate to match conservation needs: Considering diversity scales and land-use history. Landsc. Ecol. 2014, 29, 941–948. [Google Scholar] [CrossRef]
- Bezaury-Creel, J.; Graf-Montero, S.; Barcklay-Briseño, K.; de la Maza-Hernández, R.; Machado-Macías, J.S.; Rodríguez-Martínez del Sobral, E.; Rojas-González de Castilla, S.; Ruíz-Barranco, H. Los Paisajes Bioculturales, un Instrumento Para el Desarrollo Rural y la Conservación del Patrimonio Natural y Cultural; Comisión Nacional de Áreas Naturales Protegidas, Agencia Francesa de Desarrollo: Ciudad de México, México, 2015. [Google Scholar]
- Moreno-Calles, A.; Casas, A.; Blancas, J.; Torres, I.; Peréz-Negrón, E.; Caballero, J.; Masera, O.; García-Barrios, L. Agroforestry systems and biodiversity conservation in arid zones: The case of the Tehuacán-Cuicatlán Valley, Central Mexico. Agrofor. Syst. 2010, 80, 315–331. [Google Scholar] [CrossRef]
- Vallejo, M.; Casas, A.; Blancas, J.; Moreno-Calles, A.I.; Solís, L.; Rangel-Landa, S.; Dávila, P.; Téllez, O. Agroforestry systems in the highlands of the Tehuacán Valley, Mexico: Indigenous culture and biodiversity conservation. Agrofor. Syst. 2014, 88, 125–140. [Google Scholar] [CrossRef]
- Vallejo, M.; Ramírez, I.; Reyes-González, A.; Moreno-Calles, A.I.; Casas, A. Agroforestry systems of the Tehuacán Valley: Changes and maintenance of their distribution at regional level. Land 2019, 8, 1–26. [Google Scholar]
- Rendón-Sandoval, F.J.; Casas, A.; Moreno-Calles, A.I.; Torres-García, I.; García Frapolli, E. Traditional agroforestry systems and conservation of native plant diversity of seasonally dry tropical forests. Sustainability 2020, 12, 4600. [Google Scholar] [CrossRef]
- Chappell, M.J.; LaValle, L.A. Food security and biodiversity: Can we have both? An agroecological analysis. Agric. Hum. Values 2011, 28, 3–26. [Google Scholar] [CrossRef]
- Freudmann, A.; Mollik, P.; Tschapka, M.; Schulze, C.H. Impacts of oil palm agriculture on phyllostomid bat assemblages. Biodivers. Conserv. 2015, 24, 3583–3599. [Google Scholar] [CrossRef]
- Casas, A.; Parra, F.; Blancas, J.; Rangel-Landa, S.; Vallejo, M.; Figueredo, C.J.; Moreno-Calles, A.I. Origen de la domesticación y la agricultura: Cómo y por qué. In Domesticación en el Continente Americano. Volumen 1. Manejo de Biodiversidad y Evolución Dirigida por las Culturas del Nuevo Mundo; Casas, A., Torres-Guevara, J., Parra, F., Eds.; UNAM-UNALM: Morelia, México, 2016; pp. 189–224. [Google Scholar]
- Boege, E. El Patrimonio Biocultural de los Pueblos Indígenas de México; Instituto Nacional de Antropología e Historia, Comisión Nacional Para el Desarrollo de los Pueblos Indígenas: Ciudad de México, México, 2008. [Google Scholar]
- Toledo, V.M. El paradigma biocultural: Crisis ecológica, modernidad y culturas tradicionales. Soc. Ambiente 2013, 1, 50–60. [Google Scholar] [CrossRef]
- Lemon, W.C. Fitness consequences of foraging behaviour in the zebra finch. Nature 1991, 352, 153–155. [Google Scholar] [CrossRef]
- Wong, B.B.M.; Candolin, U. Behavioral responses to changing environments. Behav. Ecol. 2015, 26, 665–673. [Google Scholar] [CrossRef]
- Chambers, S.A. Birds as Environmental Indicators: Review of Literature; Parks Victoria Technical Series No. 55; Parks Victoria: Melbourne, Australia, 2008. [Google Scholar]
- Tidemann, S.; Gosler, A.G. Ethno-Ornithology: Birds, Indigenous Peoples, Culture and Society; Earthscan: London, UK, 2010. [Google Scholar]
- Harris, C.D.; Ullman, E.L. The nature of cities. Ann. Am. Acad. Polit. Soc. Sci. 1945, 242, 7–17. [Google Scholar] [CrossRef]
- Galhena, D.H.; Freed, R.; Maredia, K.M. Home gardens: A promising approach to enhance household food security and wellbeing. Agric. Food Secur. 2013, 2, 1–13. [Google Scholar] [CrossRef]
- Seress, G.; Hammer, T.; Bókony, V.; Vincze, E.; Preiszner, B.; Pipoly, I.; Sinkovics, C.; Evans, K.L.; Liker, A. Impact of urbanization on abundance and phenology of caterpillars and consequences for breeding in an insectivorous bird. Ecol. Appl. 2018, 28, 1143–1156. [Google Scholar] [CrossRef]
- Lim, H.C.; Sodhi, N.S. Responses of avian guilds to urbanisation in a tropical city. Landsc. Urban. Plan. 2004, 66, 199–215. [Google Scholar] [CrossRef]
- Davis, A.; Major, R.E.; Taylor, C.E. The association between nectar availability and nectarivore density in urban and natural environments. Urban. Ecosyst. 2015, 18, 503–515. [Google Scholar] [CrossRef]
- Ortega-Álvarez, R.; MacGregor-Fors, I. Dusting-off the file: A review of knowledge on urban ornithology in Latin America. Landsc. Urban. Plan. 2011, 101, 1–10. [Google Scholar] [CrossRef]
- Ikin, K.; Knight, E.; Lindenmayer, D.B.; Fischer, J.; Manning, A.D. Linking bird species traits to vegetation characteristics in a future urban development zone: Implications for urban planning. Urban. Ecosyst. 2012, 15, 961–977. [Google Scholar] [CrossRef]
- Narango, D.L.; Tallamy, D.W.; Snyder, K.J.; Rice, R.A. Canopy tree preference by insectivorous birds in shade-coffee farms: Implications for migratory bird conservation. Biotropica 2019, 51, 387–398. [Google Scholar] [CrossRef]
- Cardona Sánchez, J.F. Cafés Especiales: Situación Actual y Aplicación del Programa de Calidad de Café Nespresso AAA, en el Municipio de Jardín Antioquia. Ph.D. Thesis, Corporación Universitaria Lasallista, Caldas, Antioquia, 2010. [Google Scholar]
- Marín Duque, L.J. Prácticas Culturales y Culinarias Alrededor del Turismo Cafetero: Municipio de Jardín de Antioquia. Ph.D. Thesis, Universidad de Antioquia, Andes, Antioquia, 2021. [Google Scholar]
- Orejuela-Gartner, J. Orquídeas de los bosques de niebla del suroccidente de Colombia y oportunidades para su conservación. Lankesteriana 2013, 13, 152. [Google Scholar] [CrossRef][Green Version]
- Municipio de Jardín. Acuerdo N°16. Por Medio del Cual se Adopta la Revisión y Ajuste de Largo Plazo, del Esquema de Ordenamiento Territorial del Municipio de Jardín; Municipio de Jardín, Departamento de Antioquia, Colombia: Jardín, Colombia, 2018. [Google Scholar]
- Holdridge, L.R. Determination of world plant formations from simple climatic data. Science 1947, 105, 367–368. [Google Scholar] [CrossRef] [PubMed]
- Corantioquia. Plan de Manejo Ambiental. Parte alta de la Microcuenca La Linda. Municipio de Jardín, Antioquia; Corporación Autónoma Regional del Centro de Antioquia, CORANTIOQUIA: Medellín, Colombia, 2002. [Google Scholar]
- Toro Murillo, J.L. Árboles de las Montañas de Antioquia; Corporación Autónoma Regional del Centro de Antioquia, CORANTIOQUIA: Medellín, Colombia, 2012. [Google Scholar]
- Drury, R.; Homewood, K.; Randall, S. Less is more: The potential of qualitative approaches in conservation research. Anim. Conserv. 2011, 14, 18–24. [Google Scholar] [CrossRef]
- Boege, E. Protegiendo lo Nuestro: Manual Para la Gestión Ambiental Comunitaria, Uso y Conservación de la Biodiversidad de los Campesinos Indígenas de América Latina; Programas de las Naciones Unidas para el Medio Ambiente; Fondo para el Desarrollo de los Pueblos Indígenas de América Latina y el Caribe: Ciudad de México, Mexico, 2005. [Google Scholar]
- Kruschke, J.K. Doing Bayesian Data Analysis: A Tutorial with R, JAGS, and STAN, 2nd ed.; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Gelman, A.; Rubin, D.B. Inference from iterative simulation using multiple sequences. Stat. Sci. 1992, 7, 457–511. [Google Scholar] [CrossRef]
- Baath, R. Bayesian First Aid: A package that implements Bayesian alternatives to the classical *.test functions in R. In Proceedings of the UseR! 2014—The International R User Conference, Los Angeles, CA, USA, 14–18 July 2014. [Google Scholar]
- Plummer, M. JAGS: A Program for Analysis of Bayesian Graphical Models Using Gibbs Sampling. In Proceedings of the 3rd International Workshop on Distributed Statistical Computing, Vienna, Austria, 20–22 March 2003. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.r-project.org/ (accessed on 1 December 2021).
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Bibby, C.; Burgess, N.; Hill, D.; Mustoe, S. Bird Census Techniques; Academic Press: London, UK, 2000. [Google Scholar]
- Ruiz Gutiérrez, V.; Berlanga García, H.A.; Calderón-Parra, R.; Savarino Drago, A.; Aguilar-Gómez, M.Á.; Rodríguez-Contreras, V. Manual Ilustrado para el Monitoreo de Aves. PROALAS: Programa de América Latina Para las Aves Silvestres; Comisión Nacional Para el Conocimiento y Uso de la Biodiversidad/Iniciativa Para la Conservación de las Aves de Norte América, Laboratorio de Ornitología de Cornell: Ciudad de México, Mexico, 2020. [Google Scholar]
- Wagner, J.L. Visibility and bias in avian foraging data. Condor 1981, 83, 263–264. [Google Scholar] [CrossRef]
- Acevedo, M.A. Bird feeding behavior as a measure of restoration success in a Caribbean Forested Wetland. Ornitol. Neotrop. 2007, 18, 305–310. [Google Scholar]
- Ocampo, J.; Wyckhuys, K. Tecnología Para el Cultivo de la Gulupa (Passiflora edulis f. edulis Sims) en Colombia; Centro de Bio-Sistemas de la Universidad Jorge Tadeo Lozano, Centro Internacional de Agricultura Tropical, Ministerio de Agricultura y Desarrollo Rural: Bogotá, Colombia, 2012. [Google Scholar]
- FAO. El estado Mundial de la Agricultura y la Alimentación; Organización de las Naciones Unidas Para la Alimentación y la Agricultura: Roma, Italy, 2016. [Google Scholar]
- Talukder, A.; Kiess, L.; Huq, N.; de Pee, S.; Darnton-Hill, I.; Bloem, M.W. Increasing the production and consumption of vitamin A–rich fruits and vegetables: Lessons learned in taking the Bangladesh homestead gardening programme to a national scale. Food Nutr. Bull. 2000, 21, 165–172. [Google Scholar] [CrossRef]
- Calvet-Mir, L.; Gómez-Bagetthun, E.; Reyes-García, V. Beyond food production: Home gardens ecosystem services. A case study in Vall Fosca, Catalan Pyrenees, Northeastern Spain. Ecol. Econ. 2012, 74, 153–160. [Google Scholar] [CrossRef]
- Sontia, N.F.; Svendsen, E.S. Why garden? Personal and abiding motivations for community gardening in New York City. Soc. Nat. Resour. 2018, 31, 1189–1205. [Google Scholar] [CrossRef]
- Goulart, F.F.; Carvalho-Ribeiro, S.; Soares-Filho, B. Farming-biodiversity segregation or integration? Revisiting land sparing versus land sharing debate. J. Environ. Prot. 2016, 7, 1016–1032. [Google Scholar] [CrossRef]
- Suarez-Rubio, M.; Thomlinson, J.R. Landscape and patch-level factors influence bird communities in an urbanized tropical island. Biol. Conserv. 2009, 142, 1311–1321. [Google Scholar] [CrossRef]
- Morante-Filho, J.C.; Benchimol, M.; Faria, D. Landscape composition is the strongest determinant of bird occupancy patterns in tropical forest patches. Landsc. Ecol. 2021, 36, 105–117. [Google Scholar] [CrossRef]
- Ortega-Álvarez, R.; Ruiz-Gutiérrez, V.; Robinson, O.J.; Berrones Benítez, E.; Medina Mena, I.; Zúñiga-Vega, J.J. Beyond incidence data: Assessing bird habitat use in indigenous working landscapes through the analysis of behavioral variation among land uses. Landsc. Urban. Plan. 2021, 211, 104100. [Google Scholar] [CrossRef]
- Auman, H.J.; Meathrel, C.E.; Richardson, A. Supersize me: Does anthropogenic food change the body condition of Silver Gulls? A comparison between urbanized and remote, non-urbanized areas. Waterbirds 2008, 31, 122–126. [Google Scholar] [CrossRef]
- Leveau, L.M. Artificial Light at Night (ALAN) is the main driver of nocturnal Feral Pigeon (Columba livia f. domestica) foraging in urban areas. Animals 2020, 10, 554. [Google Scholar] [CrossRef]
- Jones, D.N.; James Reynolds, S. Feeding birds in our towns and cities: A global research opportunity. J. Avian Biol. 2008, 39, 265–271. [Google Scholar] [CrossRef]
- Maron, M.; Lill, A. The influence of livestock grazing and weed invasion on habitat use by birds in grassy woodland remnants. Biol. Conserv. 2005, 124, 439–450. [Google Scholar] [CrossRef]
- Dennis, P.; Skartveit, J.; McCracken, D.I.; Pakeman, R.J.; Beaton, K.; Kunaver, A.; Evans, D.M. The effects of livestock grazing on foliar arthropods associated with bird diet in upland grasslands of Scotland. J. Appl. Ecol. 2008, 45, 279–287. [Google Scholar] [CrossRef]
- Castaño-Villa, G.J.; Ramos-Valencia, S.A.; Fontúrbel, F.E. Fine-scale habitat structure complexity determines insectivorous bird diversity in a tropical forest. Acta Oecol. 2014, 61, 19–23. [Google Scholar] [CrossRef]
- Komar, O. Ecology and conservation of birds in coffee plantations: A critical review. Bird Conserv. Int. 2006, 16, 1–23. [Google Scholar] [CrossRef]
- Leyequién, E.; de Boer, W.F.; Toledo, V.M. Bird community composition in a shaded coffee agro-ecological matrix in Puebla, Mexico: The effects of landscape heterogeneity at multiple spatial scales. Biotropica 2010, 42, 236–245. [Google Scholar] [CrossRef]
- Mastrangelo, M.E.; Gavin, M.C. Impacts of agricultural intensification on avian richness at multiple scales in Dry Chaco forests. Biol. Conserv. 2014, 179, 63–71. [Google Scholar] [CrossRef]
- Silva, C.P.; Sepúlveda, R.D.; Barbosa, O. Nonrandom filtering effect on birds: Species and guilds response to urbanization. Ecol. Evol. 2016, 6, 3711–3720. [Google Scholar] [CrossRef]
- Vázquez-Reyes, L.D.; Arizmendi, M.D.C.; Godínez-Álvarez, H.O.; Navarro-Sigüenza, A.G. Directional effects of biotic homogenization of bird communities in Mexican seasonal forests. Condor 2017, 119, 275–288. [Google Scholar] [CrossRef]
- McKinney, M.L.; Lockwood, J.L. Biotic homogenization: A few winners replacing many losers in the next mass extinction. Trends Ecol. Evol. 1999, 14, 450–453. [Google Scholar] [CrossRef]
- Devictor, V.; Julliard, R.; Clavel, J.; Jiguet, F.; Lee, A.; Couvet, D. Functional biotic homogenization of bird communities in disturbed landscapes. Glob. Ecol. Biogeogr. 2008, 17, 252–261. [Google Scholar] [CrossRef]
- Shochat, E. Credit or debit? Resource input changes population dynamics of city-slicker birds. Oikos 2004, 106, 622–626. [Google Scholar] [CrossRef]
- Leveau, L.M.; Zuria, I. Flocking the city: Avian demography and population dynamics in urban Latin America. In Avian Ecology in Latin American Cityscapes; MacGregor-Fors, I., Escobar-Ibáñez, J.F., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 57–77. [Google Scholar]
- Cristaldi, M.A.; Giraudo, A.R.; Arzamendia, V.; Bellini, G.P.; Claus, J. Urbanization impacts on the trophic guild composition of bird communities. J. Nat. Hist. 2017, 51, 2385–2404. [Google Scholar] [CrossRef]
- Ghadiri Khanaposhtani, M.; Kaboli, M.; Karami, M.; Etemad, V. Effect of habitat complexity on richness, abundance and distributional pattern of forest birds. Environ. Manag. 2012, 50, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Arnold, K.E.; Ramsay, S.L.; Donaldson, C.; Adam, A. Parental prey selection affects risk-taking behaviour and spatial learning in avian offspring. Proc. R. Soc. B Biol. Sci. 2007, 274, 2563–2569. [Google Scholar] [CrossRef] [PubMed]
- Arnold, K.E.; Ramsay, S.L.; Henderson, L.; Larcombe, S.D. Seasonal variation in diet quality: Antioxidants, invertebrates and blue tits Cyanistes caeruleus. Biol. J. Linn. Soc. 2010, 99, 708–717. [Google Scholar] [CrossRef]
- Durães, R.; Marini, M.Â. A quantitative assessment of bird diets in the Brazilian Atlantic forest, with recommendations for future diet studies. Ornitol. Neotrop. 2005, 16, 65–83. [Google Scholar]
- Moran, A.J.; Prosser, S.W.J.; Moran, J.A. DNA metabarcoding allows non-invasive identification of arthropod prey provisioned to nestling Rufous hummingbirds (Selasphorus rufus). PeerJ 2019, 2019, 1–21. [Google Scholar] [CrossRef]
- Maleque, M.A.; Maeto, K.; Ishii, H.T. Arthropods as bioindicators of sustainable forest management, with a focus on plantation forests. Appl. Entomol. Zool. 2009, 44, 1–11. [Google Scholar] [CrossRef]
- González, G.; Lodge, D.J. Soil biology research across latitude, elevation and disturbance gradients: A review of forest studies from Puerto Rico during the past 25 years. Forests 2017, 8, 178. [Google Scholar] [CrossRef]
- Fenoglio, M.S.; Rossetti, M.R.; Videla, M. Negative effects of urbanization on terrestrial arthropod communities: A meta-analysis. Glob. Ecol. Biogeogr. 2020, 29, 1412–1429. [Google Scholar] [CrossRef]
- Lill, A. Food resources and urban colonisation by Lorikeets and Parrots. Vic. Nat. 2009, 126, 70–72. [Google Scholar]
- Li, N.; Yang, W.; Fang, S.; Li, X.; Liu, Z.; Leng, X.; An, S. Dispersal of invasive Phytolacca americana seeds by birds in an urban garden in China. Integr. Zool. 2017, 12, 26–31. [Google Scholar] [CrossRef]
- Federman, S.; Sinnott-Armstrong, M.; Baden, A.L.; Chapman, C.A.; Daly, D.C.; Richard, A.R.; Valenta, K.; Donoghue, M.J. The paucity of frugivores in Madagascar may not be due to unpredictable temperatures or fruit resources. PLoS ONE 2017, 12, e0168943. [Google Scholar] [CrossRef] [PubMed]
- von Helversen, D.; von Helversen, O. Object recognition by echolocation: A nectar-feeding bat exploiting the flowers of a rain forest vine. J. Comp. Physiol. A 2003, 189, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Maas, B.; Karp, D.S.; Bumrungsri, S.; Darras, K.; Gonthier, D.; Huang, J.C.-C.; Lindell, C.A.; Maine, J.J.; Mestre, L.; Michel, N.L.; et al. Bird and bat predation services in tropical forests and agroforestry landscapes. Biol. Rev. 2016, 91, 1081–1101. [Google Scholar] [CrossRef] [PubMed]
- Duchelle, A. Observations on natural resource use and conservation by the Shuar in Ecuador’s Cordillera del Cóndor. Ethnobot. Res Appl. 2007, 5, 5–23. [Google Scholar] [CrossRef]
- Jernigan, K.; Dauphiné, N. Aguaruna knowledge of bird foraging ecology: A comparison with scientific data. Ethnobot. Res. Appl. 2008, 6, 93–106. [Google Scholar] [CrossRef][Green Version]
- Hohbein, R.R.; Nibbelink, N.P.; Cooper, R.J. Non-governmental organizations improve the social-ecological fit of institutions conserving the Andean bear in Colombia. Ecol. Soc. 2021, 26, 13. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).