
Effects of an Exercise Program Combining Aerobic and Resistance Training on Protein Expressions of Neurotrophic Factors in Obese Rats Injected with Beta-Amyloid



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
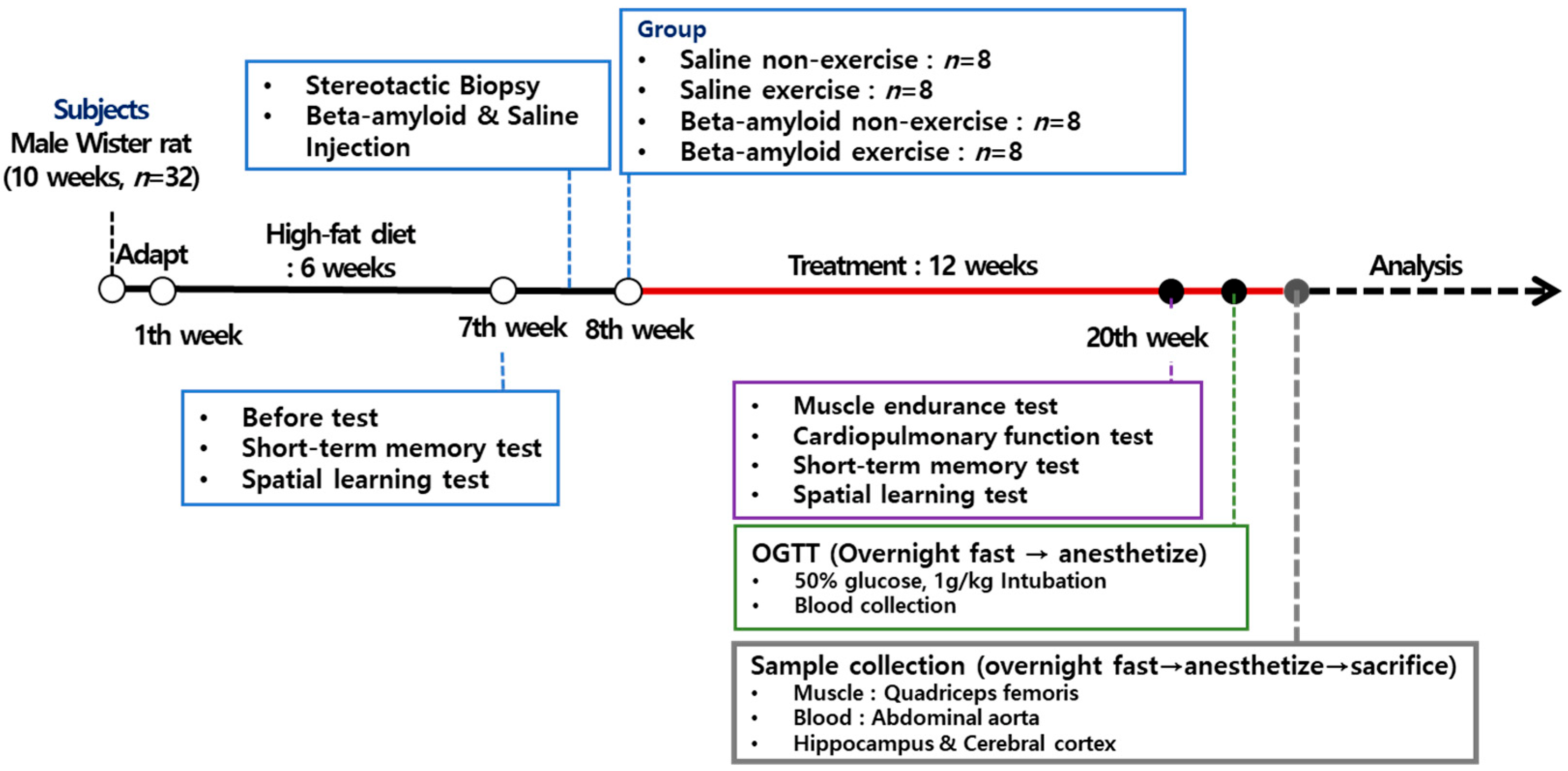
2.2. Experimental Procedures
2.3. Dietary Composition
2.4. Exercise Program
2.5. Stereotactic Biopsy for βA25-35 Injection
2.6. Experimental Variables and Methods
3. Results
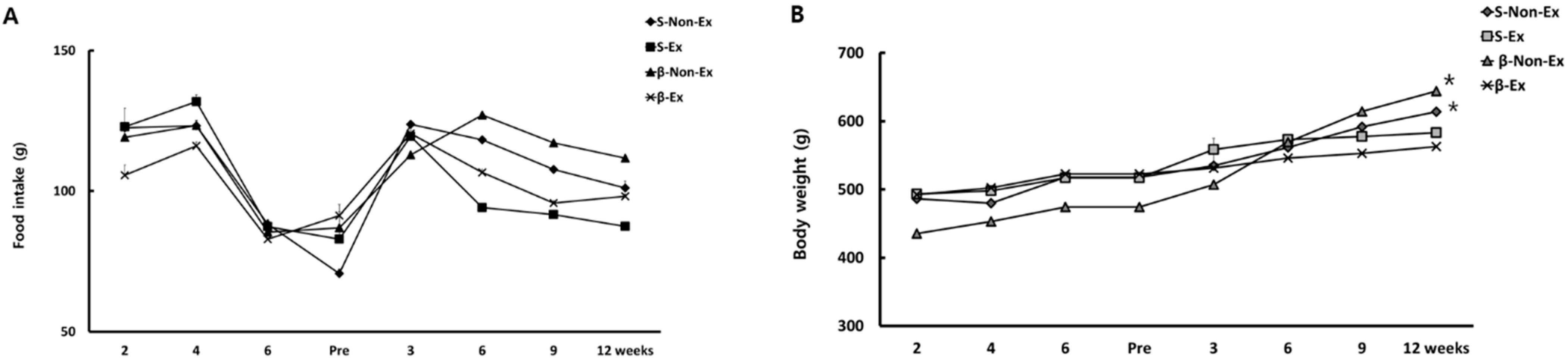
3.1. Changes in Food Intake and Body Weight
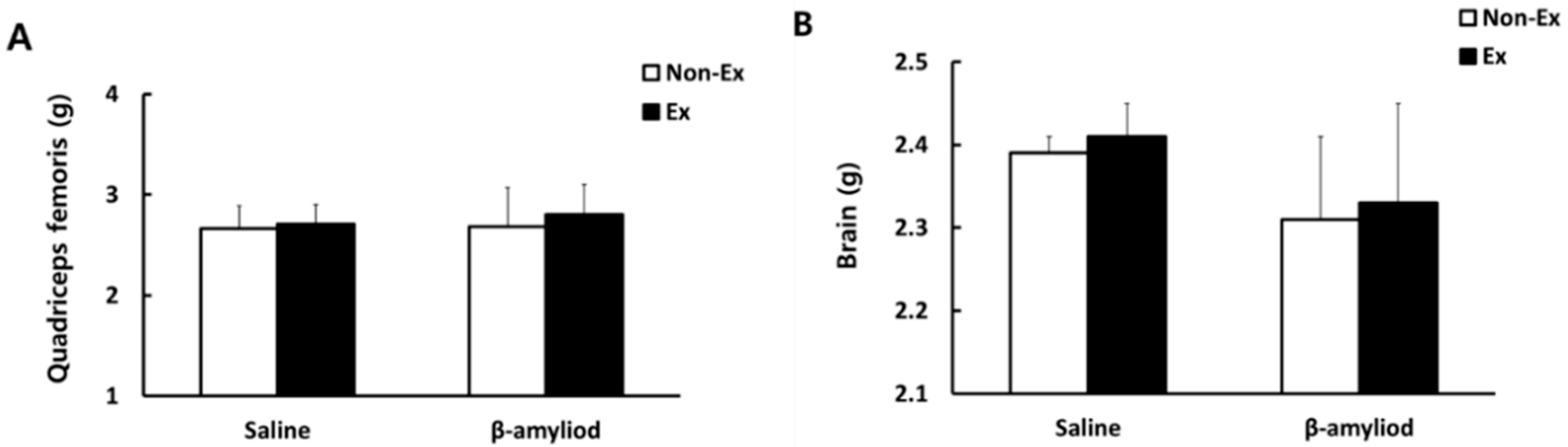
3.2. Tissue Weight
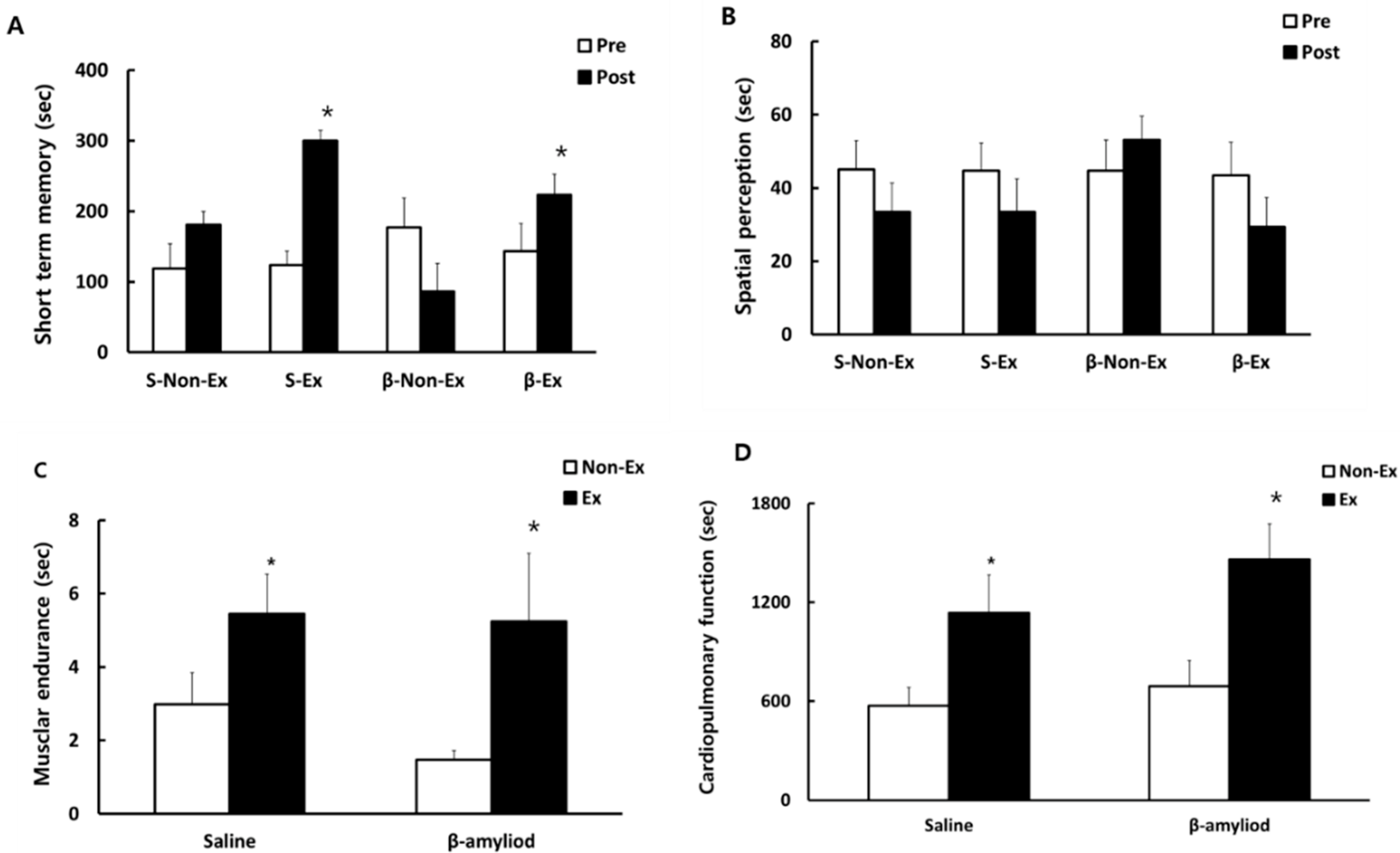
3.3. Comparison of Cognitive Function and Physical Fitness
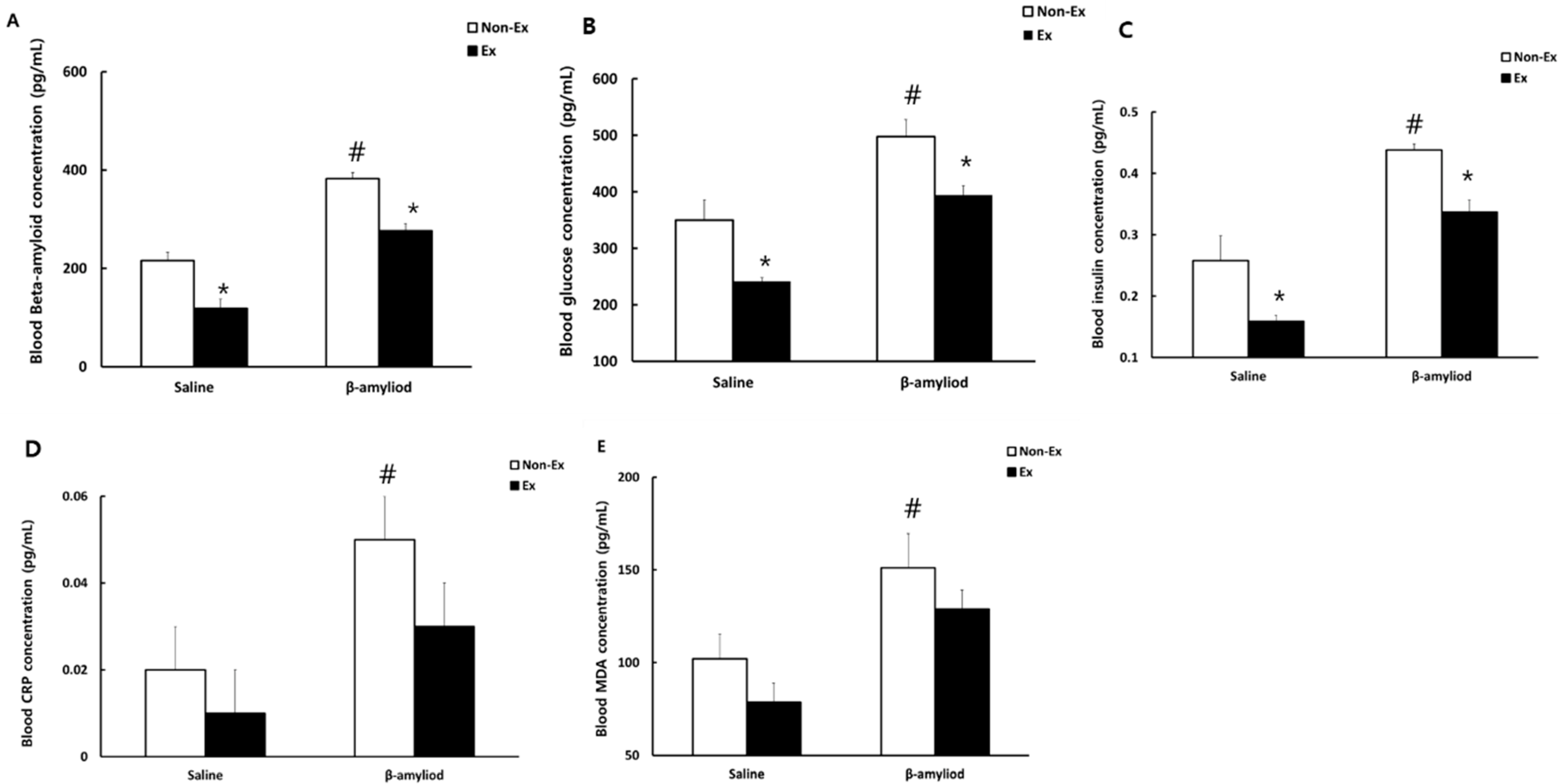
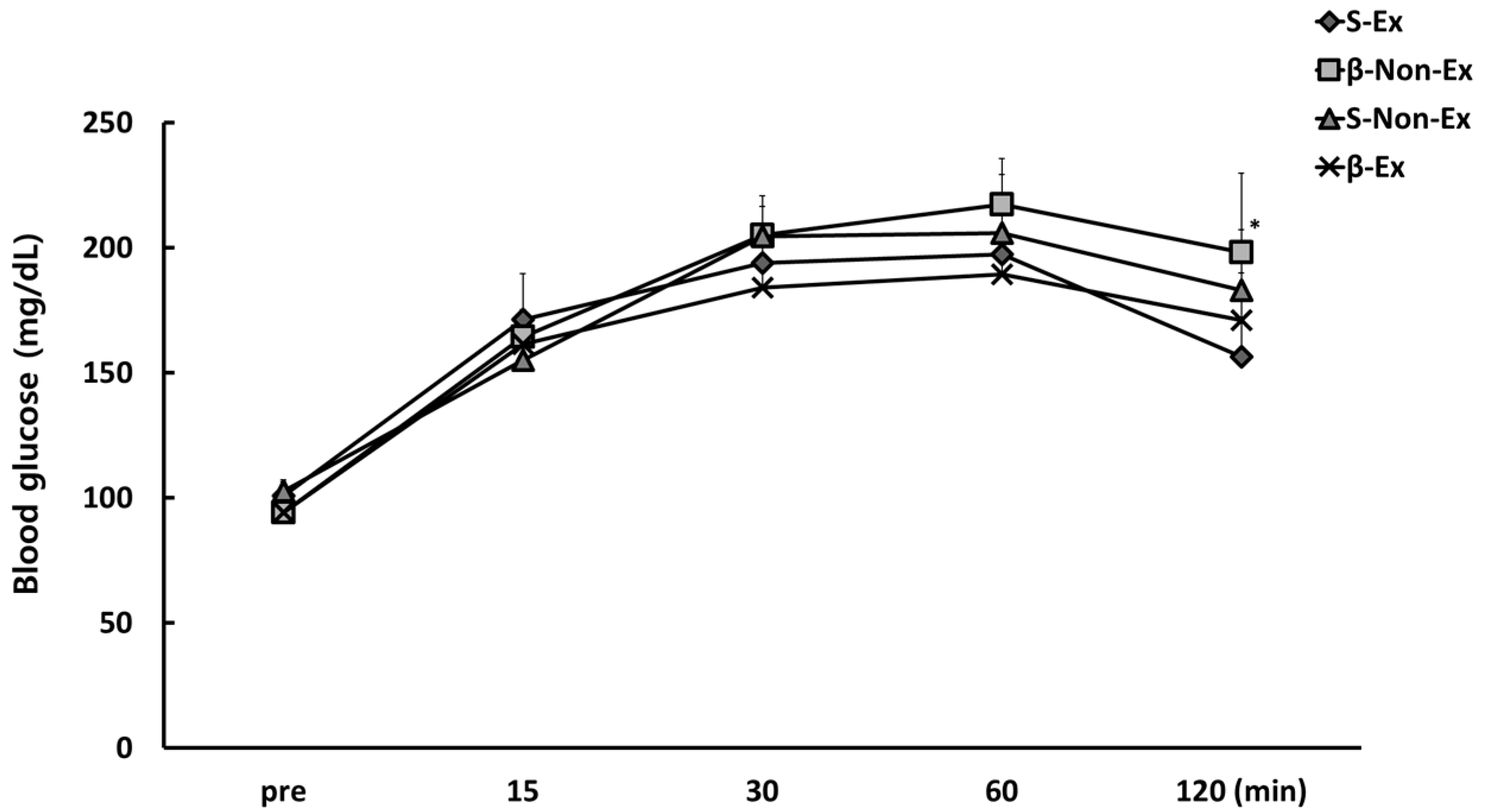
3.4. Changes in Blood Indicators
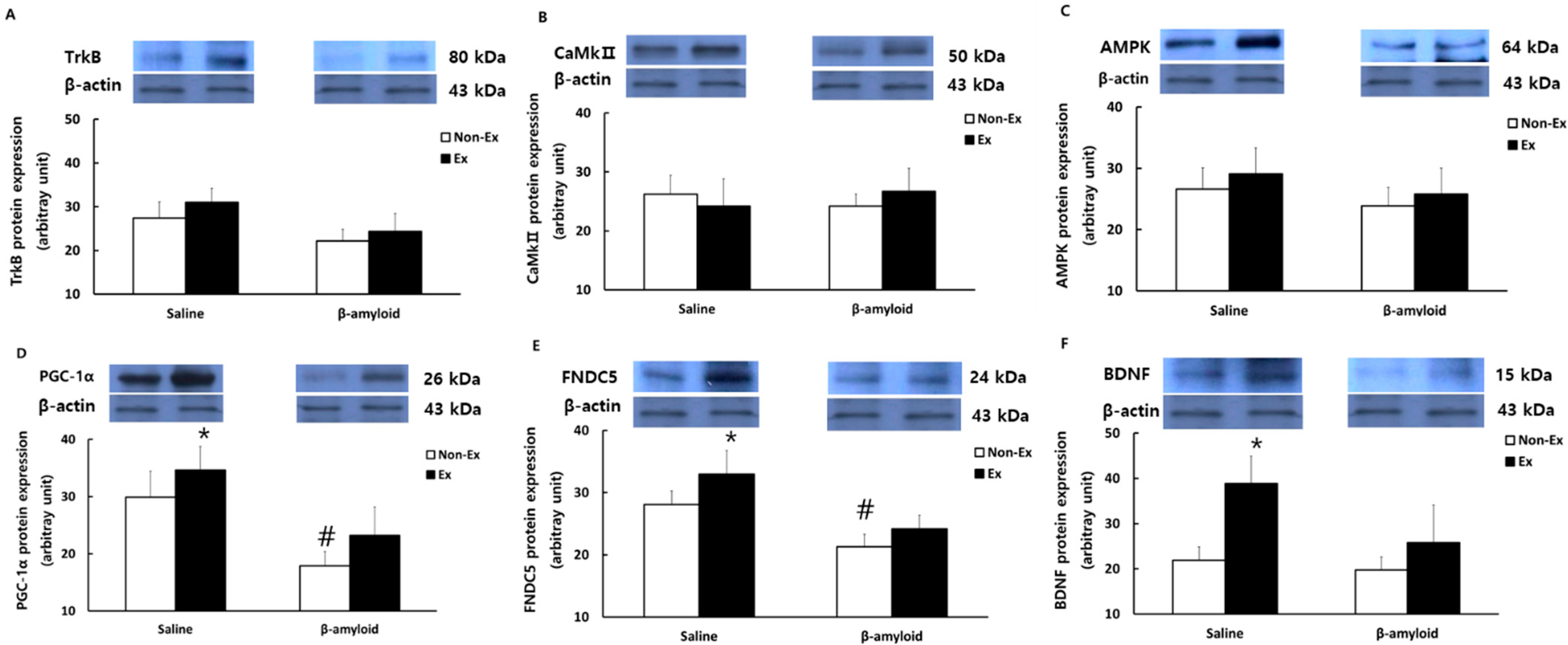
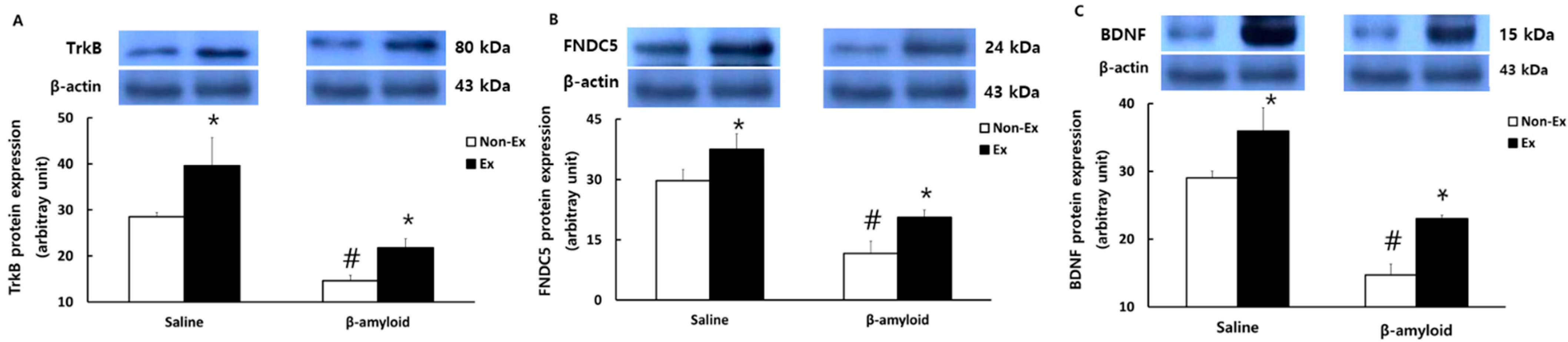
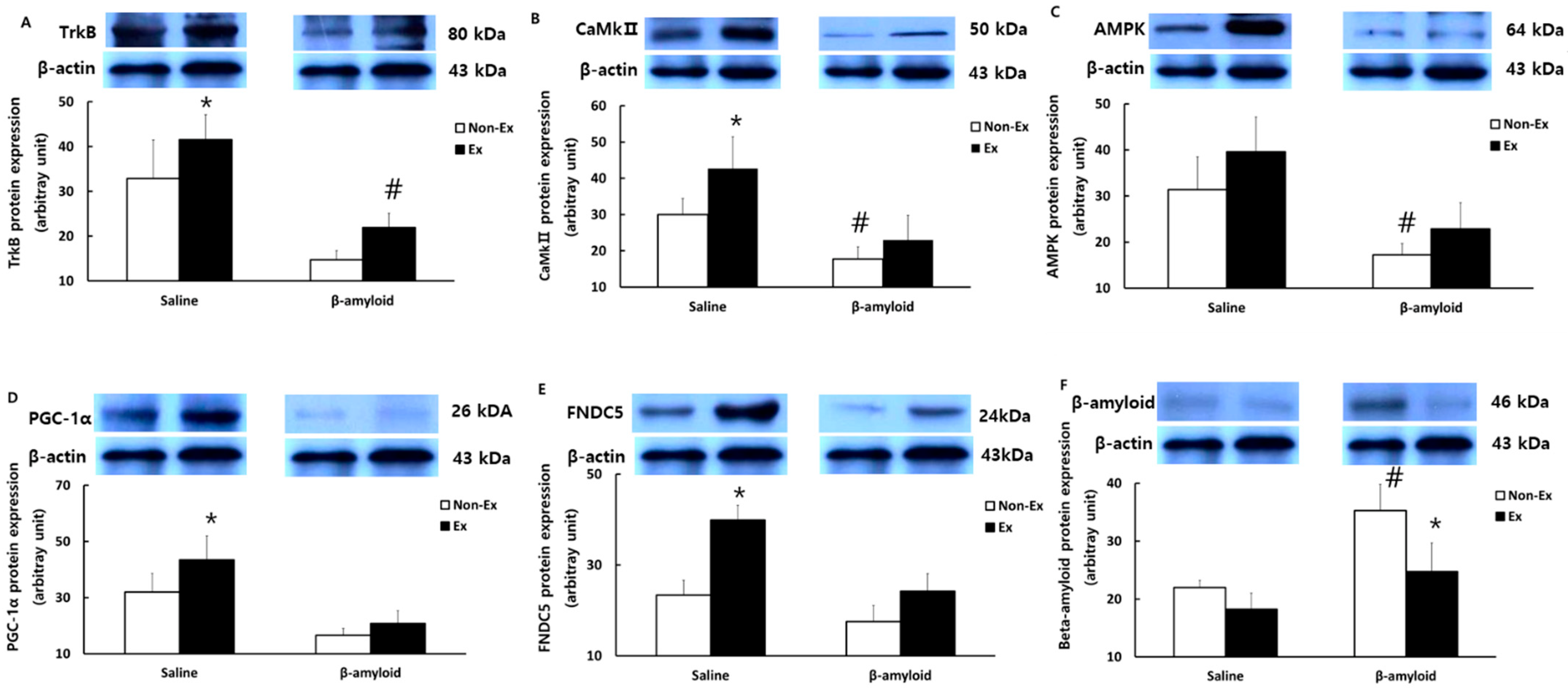
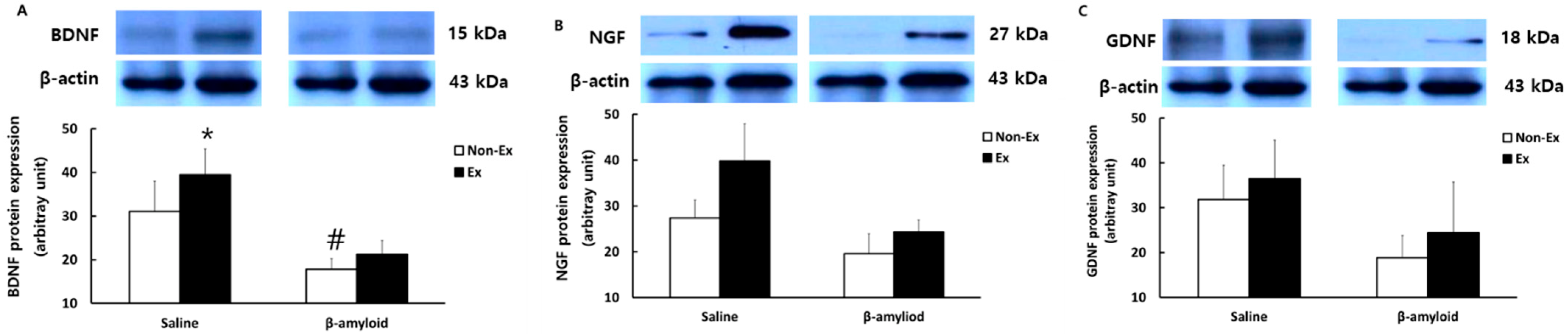
3.5. Changes in Protein Expressions of Cytokines in Different Tissues
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eduardo, P.M.M.; Poliane, G.T.V.; Renata, S.; Edurardo, L.; Renato, S.M.J.; Sergio, M.; Andrea, C.D. Aging process, cognitive decline and alzheimer’s disease: Can strength training modulate these responses? CNS Neurol. Disord. Drug Targets 2015, 14, 1209–1213. [Google Scholar]
- Wang, Y. Alzheimer disease: Lessons from immunotherapy for Alzheimer disease. Nat. Rev. Neurol. 2014, 10, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Hengstler, J.G.; Heimerdinger, C.K.; Schiffer, I.B.; Gebhard, S.; Sagemüller, J.; Tanner, B.; Bolt, H.M.; Oesch, F. Dietary topoisomerase II–poisons: Contribution of soy products to infant leukemia? EXCLI J. 2002, 1, 8–14. [Google Scholar]
- Bianchi, V.E.; Rizzi, L.; Bresciani, E.; Omeljaniuk, R.J.; Torsello, A. Androgen therapy in neurodegenerative diseases. J. Endocr. Soc. 2020, 4, bvaa120. [Google Scholar] [CrossRef] [PubMed]
- Vegeto, E.; Villa, A.; Torre, S.D.; Crippa, V.; Rusmini, P.; Cristofani, R.; Galbiati, M.; Maggi, A.; Poletti, A. The role of sex and sex hormones in neurodegenerative diseases. Endocr Rev. 2020, 41, 273–319. [Google Scholar] [CrossRef] [PubMed]
- Ozbeyli, D.; Sari, G.; Ozkan, N.; Karademir, B.; Yuksel, M.; Cilingir, K.O.T.; Kasimay, C.O. Protective effects of different exercise modalities in an Alzheimer’s disease-like model. Behav. Brain Res. 2017, 328, 159–177. [Google Scholar] [CrossRef]
- Zhang, L.; Chao, F.; Luo, Y.M.; Xiao, Q.; Jiang, L.; Zhou, C.; Zhang, Y.; Lv, F.L.; Ma, J.Q.; Tang, Y. Exercise prevents cognitive function decline and demyelination in the white matter of APP/PS1 transgenic AD mice. Curr. Alzheimer Res. 2017, 14, 645–655. [Google Scholar] [CrossRef]
- Shehab, M.A.E.; Osama, H.A.J. Aerobic exercise improves quality of life, psychological well-being and systemic inflammation in subjects with Alzheimer’s disease. Afr. Health Sci. 2016, 16, 1045–1055. [Google Scholar]
- Jaroslaw, M.; Ewa, Z.; Joanna, M.; Katarzyna, K.; Joanna, M.; Jerzy, A.; Zoladz, A.J.; Anna, J. Interval training-induced alleviation of rigidity and hypertonia in patients with Parkinson’s disease is accompanied by increased basal serum brain-derived neurotrophic factor. J. Rehabil. Med. 2015, 47, 372–375. [Google Scholar]
- Bostrom, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; James, L.; Rasbach, K.; Bostrom, E.A.; Choi, J.H.; Long, J.Z.; Kajimura, S.; et al. A PGC-1α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–471. [Google Scholar] [CrossRef]
- Sweeney, G.; Song, J. The association between PGC-1α and Alzheimer’s disease. Anat. Cell Biol. 2016, 49, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Garland, T.J.; Schutz, T.; Chappell, H.; Keeney, M.A.; Meek, B.K.; Copes, T.H.; Acosta, L.E.; Drenowatz, W.; Maciel, C.; Dijk, G. The biological control of voluntary exercise, spontaneous physical and daily energy expenditure in relation to obesity: Human and rodent perspectives. J. Exp. Biol. 2011, 214, 206–235. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, J.S.; Jonathan, P.L.; Andis, K. Physical activity and exercise attenuate neuroinflammation in neurological diseases. Brain Res. Bull. 2016, 125, 19–29. [Google Scholar]
- Weinberg, L.; Hasni, A.; Shinohara, M.; Duarte, A. A single bout of resistance exercise can enhance episodic memory performance. Acta Psychol. 2014, 153, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Naoki, S.; Hidenori, A.; Takashi, S.; Kenji, T. Frailty and sarcopenia: A new bridge to dementia. Jpn. J. Clin. Med. 2016, 74, 505–509. [Google Scholar]
- De la Rosa, A.; Olaso-Gonzalez, G.; Arc-Chagnaud, C.; Millan, F.; Salvador-Pascual, A.; García-Lucerga, C.; Blasco-Lafarga, C.; Garcia-Dominguez, E.; Carretero, A.; Correas, A.G.; et al. Physical exercise in the prevention and treatment of Alzheimer’s disease. J. Sport Health Sci. 2020, 9, 394–404. [Google Scholar] [CrossRef]
- Northey, J.M.; Cherbuin, N.; Pumpa, K.L.; Smee, D.J.; Rattray, B. Exercise interventions for cognitive function in adults older than 50: A systematic review with meta-analysis. Br. J. Sports Med. 2018, 52, 154–160. [Google Scholar] [CrossRef]
- Liang, Y.; Su, Q.; Sheng, Z.; Weng, Q.; Niu, Y.; Zhou, H.; Liu, C. Effectiveness of physical activity interventions on cognition, neuropsychiatric symptoms, and quality of life of Alzheimer’s disease: An update of a systematic review and meta-analysis. Front. Aging Neurosci. 2022, 14, 830824. [Google Scholar] [CrossRef]
- Lee, G.; Kim, K. Effects of resistance exercise and fermented soybean ingestion on the appearance of inflammatory cytokine, metabolic risk factors, and the body composition in obesity-induced white rats. Int. J. Crisis Saf. 2020, 5, 55–65. [Google Scholar] [CrossRef]
- Kim, K.; Jung, S.; Ahn, N.; Park, J.; Ju, Y.; Kim, S.; Lee, G.; Kim, M.; Jeong, Y. Effects of long-term endurance exercise and Salvia miltiorrhiza vinegar on body composition and insulin resistance in high fat diet-induced obese rats. Korean J. Food Preserv. 2017, 24, 666–672. [Google Scholar] [CrossRef]
- Ghasemi, R.; Zarifkar, A.; Rastegar, K.; Maghsoudi, M.; Moosavi, M. Repeated intra-hippocampal injection of beta-amyloid 25–35 induces a reproducible impairment of learning and memory: Considering caspase-3 and MAPKs activity. Eur. J. Pharmacol. 2014, 726, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Bermudez, R.F.; Introini, C.I.; Coleman, M.K.; McGaugh, J.L. Insular cortex and amygdala lesions induced after aversive training impair retention: Effects of degree of training. Neurobiol. Learn. Mem. 1997, 67, 57–63. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Cechetti, F.; Worm, P.V.; Elsner, V.R.; Bertoldi, K.; Sanches, E.; Ben, J.; Siqueira, I.R.; Siqueira, I.R.; Netto, C.A. Forced treadmill exercise prevents oxidative stress and memory deficits following chronic cerebral hypoperfusion in the rat. Neurobiol. Learn. Mem. 2012, 97, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Ahn, N.; Kim, K. Effects of aerobic and resistance exercise on myokines in high fat diet-induced middle-aged obese rats. Int. J. Environ. Res. Public Health 2020, 17, 2685. [Google Scholar] [CrossRef]
- Wu, C.; Yang, L.; Yong, L.; Dong, Y.; Yang, B.; Tucker, L.D.; Zong, X.; Zhang, Q. Effects of exercise training on anxious–depressive-like behavior in Alzheimer rat. Med. Sci. Sports Exerc. 2020, 52, 1456–1469. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Yang, L.; Tucker, D.; Dong, Y.; Zhu, L.; Duan, R.; Liu, T.C.; Zhang, Q. Beneficial effects of exercise pretreatment in a sporadic Alzheimer’s rat model. Med. Sci. Sports Exerc. 2018, 50, 945–956. [Google Scholar] [CrossRef]
- Alkadhi, K.A.; Dao, A.T. Exercise decreases BACE and APP levels in the hippocampus of a rat model of Alzheimer’s disease. Mol. Cell. Neurosci. 2018, 86, 25–29. [Google Scholar] [CrossRef]
- Carek, P.J.; Laibstain, S.E.; Carek, S.M. Exercise for the treatment of depression and anxiety. Int. J. Psychiatry Med. 2011, 41, 15–28. [Google Scholar] [CrossRef]
- Mee-inta, O.; Zhao, Z.; Kuo, Y. Physical exercise inhibits inflammation and microglial activation. Cells 2019, 8, 691. [Google Scholar] [CrossRef]
- Wrann, C.D.; White, J.P.; Salogiannnis, J.; Laznik-Bogoslavski, D.; Wu, J.; Ma, D.; Lin, J.D.; Greenberg, M.E.; Spiegelman, B.M. Exercise induces hippocampal BDNF through a PGC-1alpha/FNDC5 pathway. Cell Metab. 2013, 18, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Fairey, A.S.; Courneya, K.S.; Field, C.J.; Bell, G.J.; Jones, L.W.; Mackey, J.R. Randomized controlled trial of exercise and blood immune function in postmenopausal breast cancer survivors. J. Appl. Physiol. 2005, 98, 1534–1540. [Google Scholar] [CrossRef] [PubMed]
- Sellami, M.; Gasmi, M.; Denham, J.; Hayes, L.D.; Stratton, D.; Padulo, J.; Bragazzi, N. Effects of acute and chronic exercise on immunological parameters in the elderly aged: Can physical activity counteract the effects of aging? Front. Immunol. 2018, 9, 2187. [Google Scholar] [CrossRef] [PubMed]
- Herrup, K. The case for rejecting the amyloid cascade hypothesis. Nat. Neurosci. 2015, 18, 794–799. [Google Scholar] [CrossRef]
- Choi, S.H.; Bylykbashi, E.; Chatila, Z.K.; Lee, S.W.; Pulli, B.; Clemenson, G.D.; Kim, E.; Rompala, A.; Oram, M.K.; Asselin, C.; et al. Combined adult neurogenesis and BDNF mimic exercise effects on cognition in an Alzheimer’s mouse model. Science 2018, 361, eaan8821. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Chen, J.; Mao, K.; She, H.; Ren, Y.; Gui, C.; Wu, X.; Zou, F.; Li, W. Mitochondrial calcium dysfunction contributes to autophagic cell death induced by MPP+via AMPK pathway. Biochem. Biophys. Commun. 2019, 509, 390–394. [Google Scholar] [CrossRef]
- Dao, A.T.; Zagaar, M.A.; Levine, A.T.; Salim, S.; Eriksen, J.L.; Alkadhi, K.A. Treadmill exercise prevents learning and memory impairment in Alzheimer’s disease-like pathology. J. Med. Food 2013, 10, 507–515. [Google Scholar] [CrossRef]
- Wright, D.C.; Geiger, P.C.; Han, D.H.; Jones, T.E.; Holloszy, J.O. Calcium induces increases in peroxisome proliferator-activated receptor gamma coactivator-1alpha and mitochondrial biogenesis by a pathway leading to p38 mitogen-activated protein kinase activation. J. Biol. Chem. 2007, 282, 18793–18802. [Google Scholar] [CrossRef]
- Souza, L.C.; Jesse, C.R.; Del, F.L.; Gomes, M.G.; Goes, A.T.R.; Filho, C.B.; Luchese, C.; Pereira, A.A.M.; Boeira, S.P. Swimming exercise prevents behavioural disturbances induced by an intracerebroventricular injection of amyloid-β1-42 peptide through modulation of cytokine/NF-kappaB pathway and indolemine-2,3-dioxygenase in mouse brain. Behav. Brain Res. 2017, 331, 1–13. [Google Scholar] [CrossRef]
- Rezaee, Z.; Marandi, S.; Alaei, H.; Esfarjani, F. The effect of preventive exercise on the neuroprotection in 6-hydroxydopamine-lesioned rat brain. Appl. Physiol. Nutr. Metab. 2019, 44, 1267–1275. [Google Scholar] [CrossRef]
- Saket, M.N.; Shaohua, X.; Joanna, S.K.; Krisztina, M.; Lennart, B.; Mark, P.M. Exercise and BDNF reduce Aβ production by enhancing α-secretase processing of APP. J. Neurochem. 2017, 142, 286–296. [Google Scholar]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, G.; Kim, Y.; Jang, J.-H.; Lee, C.; Yoon, J.; Ahn, N.; Kim, K. Effects of an Exercise Program Combining Aerobic and Resistance Training on Protein Expressions of Neurotrophic Factors in Obese Rats Injected with Beta-Amyloid. Int. J. Environ. Res. Public Health 2022, 19, 7921. https://doi.org/10.3390/ijerph19137921
Lee G, Kim Y, Jang J-H, Lee C, Yoon J, Ahn N, Kim K. Effects of an Exercise Program Combining Aerobic and Resistance Training on Protein Expressions of Neurotrophic Factors in Obese Rats Injected with Beta-Amyloid. International Journal of Environmental Research and Public Health. 2022; 19(13):7921. https://doi.org/10.3390/ijerph19137921
Chicago/Turabian StyleLee, Gyuho, Yunwook Kim, Jung-Hee Jang, Chan Lee, Jaewoo Yoon, Nayoung Ahn, and Kijin Kim. 2022. "Effects of an Exercise Program Combining Aerobic and Resistance Training on Protein Expressions of Neurotrophic Factors in Obese Rats Injected with Beta-Amyloid" International Journal of Environmental Research and Public Health 19, no. 13: 7921. https://doi.org/10.3390/ijerph19137921
APA StyleLee, G., Kim, Y., Jang, J.-H., Lee, C., Yoon, J., Ahn, N., & Kim, K. (2022). Effects of an Exercise Program Combining Aerobic and Resistance Training on Protein Expressions of Neurotrophic Factors in Obese Rats Injected with Beta-Amyloid. International Journal of Environmental Research and Public Health, 19(13), 7921. https://doi.org/10.3390/ijerph19137921