Rcs Phosphorelay Responses to Truncated Lipopolysaccharide-Induced Cell Envelope Stress in Yersinia enterocolitica



Abstract

1. Introduction
2. Results
2.1. waaF Gene Knockout in Y. enterocolitica Constructed a Truncated LPS Structure
2.2. Truncated LPS-Induced Envelope Stress in ΔwaaF Strain
2.3. Deletion of waaF Gene Increased Susceptibility of Y. enterocolitica to Environmental Stress
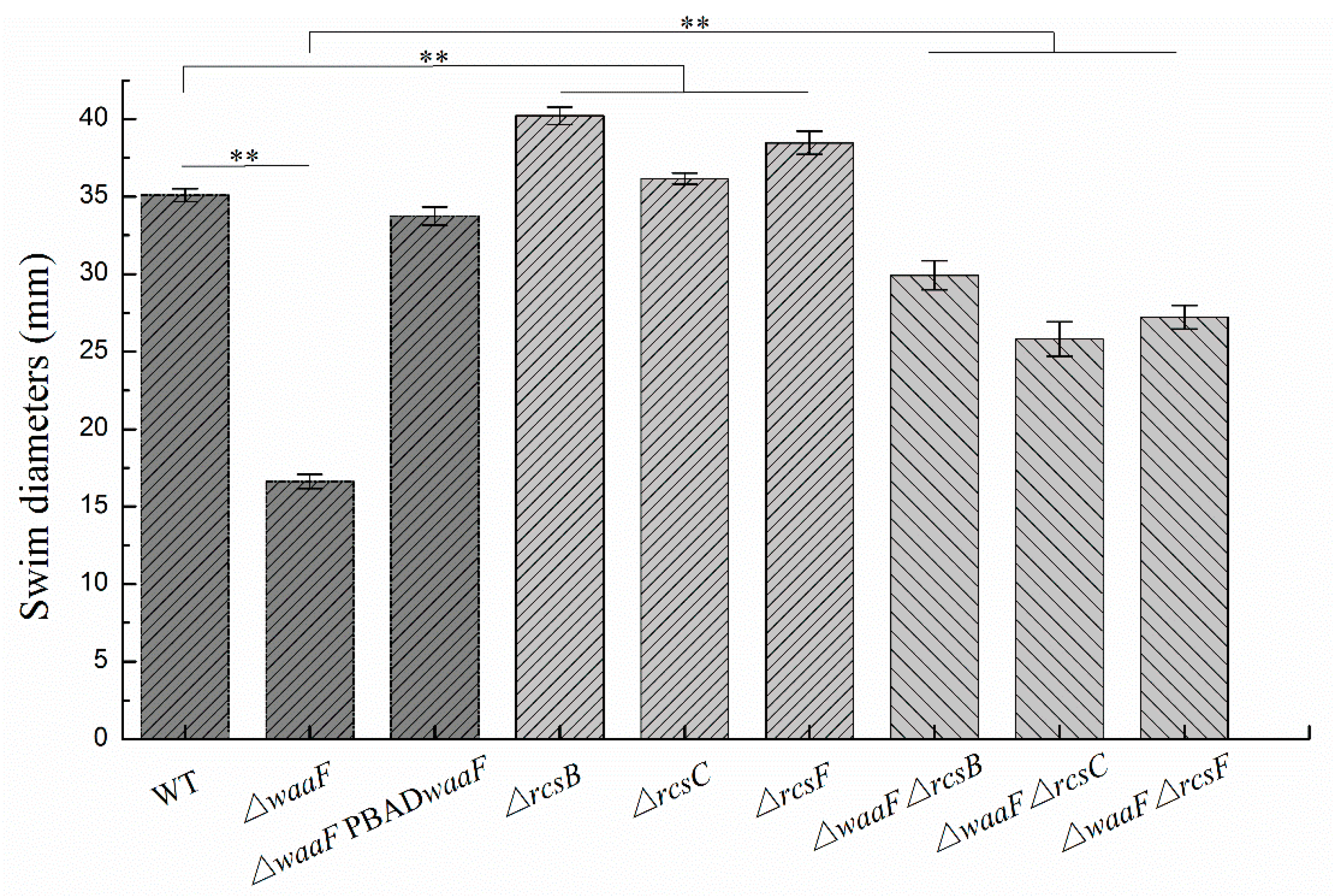
2.4. Disruption of the Rcs Phosphorelay Reversed the Decreased Motility and Biofilm Formation in ΔwaaF Strain
2.5. LPS Truncation Caused by waaF Deletion Activated the Rcs Phosphorelay in Y. enterocolitica
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Plasmids, and Culture Conditions
4.2. Plasmids and Strains Construction
4.3. Observation of Colony Morphology
4.4. LPS Isolation and Silver Stained SDS-PAGE
4.5. Hydrophobicity Test
4.6. NPN Uptake Assays
4.7. Stress Survival Assays
4.8. Motility Assays
4.9. Biofilm Assays
4.10. Electron Microscopy of Cell Morphology
4.11. RNA Isolation and Quantitative Real-Time PCR (RT-qPCR)
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huovinen, E.; Sihvonen, L.M.; Virtanen, M.J.; Haukka, K.; Siitonen, A.; Kuusi, M. Symptoms and sources of Yersinia enterocolitica-infection: A case-control study. BMC Infect. Dis. 2010, 10, 122. [Google Scholar] [CrossRef] [PubMed]
- Fabrega, A.; Vila, J. Yersinia enterocolitica: Pathogenesis, virulence and antimicrobial resistance. Enferm. Infecc. Microbiol. Clín. 2012, 30, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Zadernowska, A.; Chajecka-Wierzchowska, W.; Laniewska-Trokenheim, L. Yersinia enterocolitica: A dangerous, but often ignored, foodborne pathogen. Food Rev. Int. 2014, 30, 53–70. [Google Scholar] [CrossRef]
- European Food Safety Authority and European Centre for Disease Prevention and Control. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, 5500. [Google Scholar]
- Annamalai, T.; Venkitanarayanan, K. Expression of major cold shock proteins and genes by Yersinia enterocolitica in synthetic medium and foods. J. Food Prot. 2005, 68, 2454–2458. [Google Scholar] [CrossRef]
- Bari, M.L.; Hossain, M.A.; Isshiki, K.; Ukuku, D. Behavior of Yersinia enterocolitica in foods. J. Pathog. 2011, 2011, 420732. [Google Scholar] [CrossRef]
- Rahman, A.; Bonny, T.S.; Stonsaovapak, S.; Ananchaipattana, C. Yersinia enterocolitica: Epidemiological studies and outbreaks. J. Pathog. 2011, 2011, 239391. [Google Scholar] [CrossRef]
- Meng, J.; Bai, J.Q.; Chen, J.Y. Transcriptomic analysis reveals the role of RcsB in suppressing bacterial chemotaxis, flagellar assembly and infection in Yersinia enterocolitica. Curr. Genet. 2020, 66, 971–988. [Google Scholar] [CrossRef]
- Brzostek, K.; Skorek, K.; Raczkowska, A. OmpR, a central integrator of several cellular responses in Yersinia enterocolitica. Adv. Exp. Med. Biol. 2012, 954, 325–334. [Google Scholar]
- Meng, J.; Huang, C.; Huang, X.N.; Liu, D.Y.; Han, B.Z.; Chen, J.Y. Osmoregulated periplasmic glucans transmit external signals through Rcs phosphorelay pathway in Yersinia enterocolitica. Front. Microbiol. 2020, 11, 122. [Google Scholar] [CrossRef]
- Guo, X.P.; Sun, Y.C. New insights into the non-orthodox two component Rcs phosphorelay system. Front. Microbiol. 2017, 8, 2014. [Google Scholar] [CrossRef] [PubMed]
- Wall, E.; Majdalani, N.; Gottesman, S. The complex Rcs regulatory cascade. Annu. Rev. Microbiol. 2018, 72, 111–139. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.M.; Silhavy, T.J. Envelope stress responses: Balancing damage repair and toxicity. Nat. Rev. Microbiol. 2019, 17, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Castanie-Cornet, M.P.; Cam, K.; Jacq, A. RcsF is an outer membrane lipoprotein involved in the RcsCDB phosphorelay signaling pathway in Escherichia coli. J. Bacteriol. 2006, 188, 4264–4270. [Google Scholar] [CrossRef]
- Cho, S.H.; Szewczyk, J.; Pesavento, C.; Zietek, M.; Banzhaf, M.; Roszczenko, P.; Asmar, A.; Laloux, G.; Hov, A.K.; Leverrier, P.; et al. Detecting envelope stress by monitoring β-barrel assembly. Cell 2014, 159, 1652–1664. [Google Scholar] [CrossRef]
- Pristovsek, P.; Sengupta, K.; Lohr, F.; Schafer, B.; von Trebra, M.W.; Ruterjans, H.; Bernhard, F. Structural analysis of the DNA-binding domain of the Erwinia amylovora RcsB protein and its interaction with the RcsAB box. J. Biol. Chem. 2003, 278, 17752–17759. [Google Scholar] [CrossRef]
- Venkatesh, G.R.; Kembou Koungni, F.C.; Paukner, A.; Stratmann, T.; Blissenbach, B.; Schnetz, K. BglJ-RcsB Heterodimers Relieve Repression of the Escherichia coli bgl Operon by H-NS. J. Bacteriol. 2010, 192, 6456–6464. [Google Scholar] [CrossRef]
- Pannen, D.; Fabisch, M.; Gausling, L.; Schnetz, K. Interaction of the RcsB response regulator with auxiliary transcription regulators in Escherichia coli. J. Biol. Chem. 2016, 291, 2357–2370. [Google Scholar] [CrossRef]
- Bialas, N.; Kasperkiewicz, K.; Radziejewska-Lebrecht, J.; Skurnik, M. Bacterial cell surface structures in Yersinia enterocolitica. Arch. Immunol. Ther. Exp. 2012, 60, 199–209. [Google Scholar] [CrossRef]
- Wang, X.; Quinn, P.J. Lipopolysaccharide: Biosynthetic pathway and structure modification. Prog. Lipid Res. 2010, 49, 97–107. [Google Scholar] [CrossRef]
- Rosenfeld, Y.; Shai, Y. Lipopolysaccharide (Endotoxin)-host defense antibacterial peptides interactions: Role in bacterial resistance and prevention of sepsis. Biochim. Biophys. Acta 2006, 1758, 1513–1522. [Google Scholar] [CrossRef] [PubMed]
- Leskinen, K.; Varjosalo, M.; Li, Z.L.; Li, C.M.; Skurnik, M. Expression of the Yersinia enterocolitica O:3 LPS O-antigen and outer core gene clusters is RfaH-dependent. Microbiology 2015, 161, 1282–1294. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Wang, Z.; Li, Y.; Hu, X.Q.; Wang, X.Y. Effects of lipopolysaccharide core sugar deficiency on colanic acid biosynthesis in Escherichia coli. J. Bacteriol. 2016, 198, 1576–1584. [Google Scholar] [CrossRef] [PubMed]
- Konovalova, A.; Perlman, D.H.; Cowles, C.E.; Silhavy, T.J. Transmembrane domain of surface-exposed outer membrane lipoprotein RcsF is threaded through the lumen of β-barrel proteins. Proc. Natl. Acad. Sci. USA 2014, 111, E4350–E4358. [Google Scholar] [CrossRef] [PubMed]
- Konovalova, A.; Mitchell, A.M.; Silhavy, T.J. A lipoprotein/β-barrel complex monitors lipopolysaccharide integrity transducing information across the outer membrane. eLife 2016, 5, e15276. [Google Scholar] [CrossRef] [PubMed]
- Girgis, H.S.; Liu, Y.; Ryu, W.S.; Tavazoie, S. A comprehensive genetic characterization of bacterial motility. PLoS Genet. 2007, 3, 1644–1660. [Google Scholar] [CrossRef]
- Yethon, J.A.; Heinrichs, D.E.; Monteiro, M.A.; Perry, M.B.; Whitfield, C. Involvement of waaY, waaQ, and waaP in the modification of Escherichia coli lipopolysaccharide and their role in the formation of a stable outer membrane. J. Biol. Chem. 1998, 273, 26310–26316. [Google Scholar] [CrossRef]
- Yethon, J.A.; Whitfield, C. Purification and characterization of WaaP from Escherichia coli, a lipopolysaccharide kinase essential for outer membrane stability. J. Biol. Chem. 2001, 276, 5498–5504. [Google Scholar] [CrossRef]
- Hancock, R.E.; Diamond, G. The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol. 2000, 8, 402–410. [Google Scholar] [CrossRef]
- Papo, N.; Shai, Y. A molecular mechanism for lipopolysaccharide protection of Gram-negative bacteria from antimicrobial peptides. J. Biol. Chem. 2005, 280, 10378–10387. [Google Scholar] [CrossRef]
- Mireles, J.R.; Toguchi, A.; Harshey, R.M. Salmonella enterica serovar typhimurium swarming mutants with altered biofilm-forming abilities: Surfactin inhibits biofilm formation. J. Bacteriol. 2001, 183, 5848–5854. [Google Scholar] [CrossRef] [PubMed]
- Nakao, R.; Ramstedt, M.; Wai, S.N.; Uhlin, B.E. Enhanced biofilm formation by Escherichia coli LPS mutants defective in Hep biosynthesis. PLoS ONE 2012, 7, e51241. [Google Scholar] [CrossRef] [PubMed]
- Al-Hendy, A.; Toivanen, P.; Skurnik, M. Lipopolysaccharide O side chain of Yersinia enterocolitica O:3 is an essential virulence factor in an orally infected murine model. Infect. Immun. 1992, 60, 870–875. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Bai, J.Q.; Xu, J.H.; Huang, C.; Chen, J.Y. Differential regulation of physiological activities by RcsB and OmpR in Yersinia enterocolitica. FEMS Microbiol. Lett. 2019, 336, fnz210. [Google Scholar] [CrossRef] [PubMed]
- Fang, N.; Yang, H.Y.; Fang, H.H.; Liu, L.; Zhang, Y.Q.; Wang, L.; Han, Y.P.; Zhou, D.S.; Yang, R.F. RcsAB is a major repressor of Yersinia biofilm development through directly acting on hmsCDE, hmsT, and hmsHFRS. Sci. Rep. 2015, 5, 9566. [Google Scholar] [CrossRef]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef]
- Yan, F.L.; Dang, Q.F.; Liu, C.S.; Yan, J.Q.; Wang, T.; Fan, B.; Cha, D.S.; Li, X.L.; Liang, S.G.; Zhang, Z.Z. 3,6-O-[N-(2-Aminoethyl)-acetamide-yl]-chitosan exerts antibacterial activity by a membrane damage mechanism. Carbohydr. Polym. 2016, 149, 102–111. [Google Scholar] [CrossRef]
- Matatkova, O.; Pospisilova, D.; Michailidu, J.; Jaros, P.; Masak, J. Effect of subinhibitory concentration of antibiotics on Rhodococcus erythropolis and Pseudomonas fluorescens biofilm formation. Chem. Pap. 2019, 73, 1113–1119. [Google Scholar] [CrossRef]
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef]
- Nagy, G.; Dobrindt, U.; Schneider, G.; Khan, A.S.; Hacker, J.; Emody, L. Loss of regulatory protein RfaH attenuates virulence of uropathogenic Escherichia coli. Infect. Immun. 2002, 70, 4406–4413. [Google Scholar] [CrossRef]
- Bengoechea, J.A.; Skurnik, M. Temperature-regulated efflux pump/potassium antiporter system mediates resistance to cationic antimicrobial peptides in Yersinia. Mol. Microbiol. 2000, 37, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Sledjeski, D.D.; Gottesman, S. Osmotic shock induction of capsule synthesis in Escherichia coli K-12. J. Bacteriol. 1996, 178, 1204–1206. [Google Scholar] [CrossRef] [PubMed]
- Callewaert, L.; Vanoirbeek, K.G.A.; Lurquin, I.; Michiels, C.W.; Aertsen, A. The Rcs system two-component system regulates expression of lysozyme inhibitors and is induced by exposure to lysozyme. J. Bacteriol. 2009, 191, 1979–1981. [Google Scholar] [CrossRef] [PubMed]
- Farizano, J.V.; Torres, M.A.; Pescaretti, M.D.L.M.; Delgado, M.A. The RcsCDB regulatory system plays a crucial role in the protection of Salmonella enterica serovar Typhimurium against oxidative stress. Microbiology 2014, 160, 2190–2199. [Google Scholar] [CrossRef]
- Hirakawa, H.; Nishino, K.; Yamada, J.; Hirata, T.; Yamaguchi, A. Betalactam resistance modulated by the overexpression of response regulators of two-component signal transduction systems in Escherichia coli. J. Antimicrob. Chemother. 2003, 52, 576–582. [Google Scholar] [CrossRef]
- Farris, C.; Sanowar, S.; Bader, M.W.; Pfuetzner, R.; Miller, S.I. Antimicrobial peptides activate the Rcs regulon through the outer membrane lipoprotein RcsF. J. Bacteriol. 2010, 192, 4894–4903. [Google Scholar] [CrossRef]
- Bengoechea, J.A.; Najdenski, H.; Skurnik, M. Lipopolysaccharide O antigen status of Yersinia enterocolitica O:8 is essential for virulence and absence of O antigen affects the expression of other Yersinia virulence factors. Mol. Microbiol. 2004, 52, 451–469. [Google Scholar] [CrossRef]
- Bengoechea, J.A.; Zhang, L.; Toivanen, P.; Skurnik, M. Regulatory network of lipopolysaccharide O-antigen biosynthesis in Yersinia enterocolitica includes cell envelope-dependent signals. Mol. Microbiol. 2002, 44, 1045–1062. [Google Scholar] [CrossRef]
- Faille, C.; Lemy, C.; Allion-Maurer, A.; Zoueshtiagh, F. Evaluation of the hydrophobic properties of latex microspheres and Bacillus spores. Influence of the particle size on the data obtained by the MATH method (microbial adhesion to hydrocarbons). Colloid Surf. B Biointerfaces 2019, 182, 110398. [Google Scholar] [CrossRef]
- Meng, J.; Xu, J.G.; Chen, J.Y. The role of osmoregulated periplasmic glucans in the biofilm antibiotic resistance of Yersinia enterocolitica. Microb. Pathog. 2020, 147, 104284. [Google Scholar] [CrossRef]
- Meng, J.; Wang, B.Y.; Liu, D.Y.; Tao, C.; Wang, Z.W.; Zhao, X.M. High-yield anaerobic succinate production by strategically regulating multiple metabolic pathways based on stoichiometric maximum in Escherichia coli. Microb. Cell Fact. 2016, 15, 141. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Relevant Characteristics | Sources |
|---|---|---|
| Y. enterocolitica | ||
| YE | WT, Serotype O:8, Biotype 1B, pYV- | [34] |
| YE-W | ΔwaaF | This study |
| YE-W+ | ΔwaaF, PBADwaaF; Ampr | This study |
| YE-F | ΔrcsF | [10] |
| YE-C | ΔrcsC | [10] |
| YE-B | ΔrcsB | [34] |
| YE-WF | ΔwaaF, ΔrcsF | This study |
| YE-WC | ΔwaaF, ΔrcsC | This study |
| YE-WB | ΔwaaF, ΔrcsB | This study |
| E. coli | ||
| S17-1 λpir | recA1, thi, pro, hsdR-M+, RP4:2-Tc::Mu-Kan::Tn7, λpir | [34] |
| DH5a | F-, φ80lacZΔM15, Δ(lacZYA-argF)U169, deoR, recA1, endA1, hsdR17(rk−, mk+), phoA, supE44, λ-, thi-1, gyrA96, relA1 | [34] |
| Plasmids | ||
| pDS132 | conditional replication vector; R6K origin, mobRK4 transfer origin, sucrose-inducible-sacB; Cmr | [34] |
| pDS132-ΔwaaF | upstream and downstream fragments of waaF gene were cloned into pDS132; Cmr | This study |
| pDS132-ΔrcsF | upstream and downstream fragments of rcsF gene were cloned into pDS132; Cmr | [10] |
| pDS132-ΔrcsC | upstream and downstream fragments of rcsC gene were cloned into pDS132; Cmr | [10] |
| pDS132-ΔrcsB | upstream and downstream fragments of rcsB gene were cloned into pDS132; Cmr | [34] |
| pBAD24 | AraC, promoter PBAD; Ampr | [34] |
| pBAD24-waaF | AraC, PBAD-waaF; Ampr | This study |
| Primer | Sequences (5′→3′) |
|---|---|
| waaF-up-F | ATCGCATGCTGCCACAAGCTGATTCACAGA |
| waaF-up-R | ACCGTTTATCAATTCCTTGCAGCAAGTTATT |
| waaF-down-F | AATAACTTGCTGCAAGGAATTGATAAACGGTTGCATGTATTGATCGTTAAAA |
| waaF-down-R | ATTCAGAGCTCCTGCGCAATAGCATAATCGCC |
| p-waaF-F | ATGCGTCGACATGAAAATACTGGTAATCG |
| p-waaF-R | CCCAAGCTTTTAATCGCCCTCTTTCACA |
| rcsF-up-F | ACTGCATGCAAATCATTGGAAGAACTGCAAC |
| rcsF-down-R | ACTGCGAGCTCCTTTGCGGTAGGCTGGGCGTG |
| rcsC-up-F | ACTGCATGCCTCAATGGCGACGATCGGGTTA |
| rcsC-down-R | ACTGCGAGCTCCAGATTTAGCCATAATAGTAC |
| rcsB-up-F | ACTGCATGCAGAAGTGCGTTCTATAATCACA |
| rcsB-down-R | ACTGCGAGCTCATCTGGATGAGAATGCAGATC |
| Primer | Sequences (5′→3′) | Amplicon Size (bp) |
|---|---|---|
| q-flhDC-F | CCTCAGCGATGTTTCGTCTC | 176 |
| q-flhDC-R | CTGCAAGTCATCCACACGAG | |
| q-hmsHFRS-F | GATGATGTACCGCCTCCAGA | 96 |
| q-hmsHFRS-R | GTGAATAGTTTCCCGCGCAT | |
| q-hmsT-F | TATAATCGCCGTGGGTTGGA | 144 |
| q-hmsT-R | CACTAAGGCTTGGTCTCCCA | |
| 16S rRNA-F | GCACGTAATGGTGGGAACTC | 183 |
| 16S rRNA-R | CTCCAATCCGGACTACGACA |
Sample Availability: Samples of the compounds are available from the authors. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, J.; Xu, J.; Huang, C.; Chen, J. Rcs Phosphorelay Responses to Truncated Lipopolysaccharide-Induced Cell Envelope Stress in Yersinia enterocolitica. Molecules 2020, 25, 5718. https://doi.org/10.3390/molecules25235718
Meng J, Xu J, Huang C, Chen J. Rcs Phosphorelay Responses to Truncated Lipopolysaccharide-Induced Cell Envelope Stress in Yersinia enterocolitica. Molecules. 2020; 25(23):5718. https://doi.org/10.3390/molecules25235718
Chicago/Turabian StyleMeng, Jiao, Junhong Xu, Can Huang, and Jingyu Chen. 2020. "Rcs Phosphorelay Responses to Truncated Lipopolysaccharide-Induced Cell Envelope Stress in Yersinia enterocolitica" Molecules 25, no. 23: 5718. https://doi.org/10.3390/molecules25235718
APA StyleMeng, J., Xu, J., Huang, C., & Chen, J. (2020). Rcs Phosphorelay Responses to Truncated Lipopolysaccharide-Induced Cell Envelope Stress in Yersinia enterocolitica. Molecules, 25(23), 5718. https://doi.org/10.3390/molecules25235718