Lactoferrin-Derived Peptides as a Control Strategy against Skinborne Staphylococcal Biofilms

 ,
,  ,
,  ,
, 

Abstract
1. Introduction
2. Experimental Section
2.1. Bacteria and Culture Conditions
2.2. Static Biofilm Formation
2.3. Antimicrobial Activity of HLF
2.4. LC/MS/MS Analysis of HLF
2.5. Purification of Antimicrobial Peptides (AMPs) by Gel Filtration Chromatography (GFC) and Their Antimicrobial Activity
2.6. Evaluation of Antibiofilm Activity and MBIC Determination
2.7. Biofilm Eradication by HLF
2.8. Biofilm Eradication by Dipping in HLF Solutions
2.9. Eradication by Sprayed HLF Solution
2.10. Statistical Analyses
3. Results
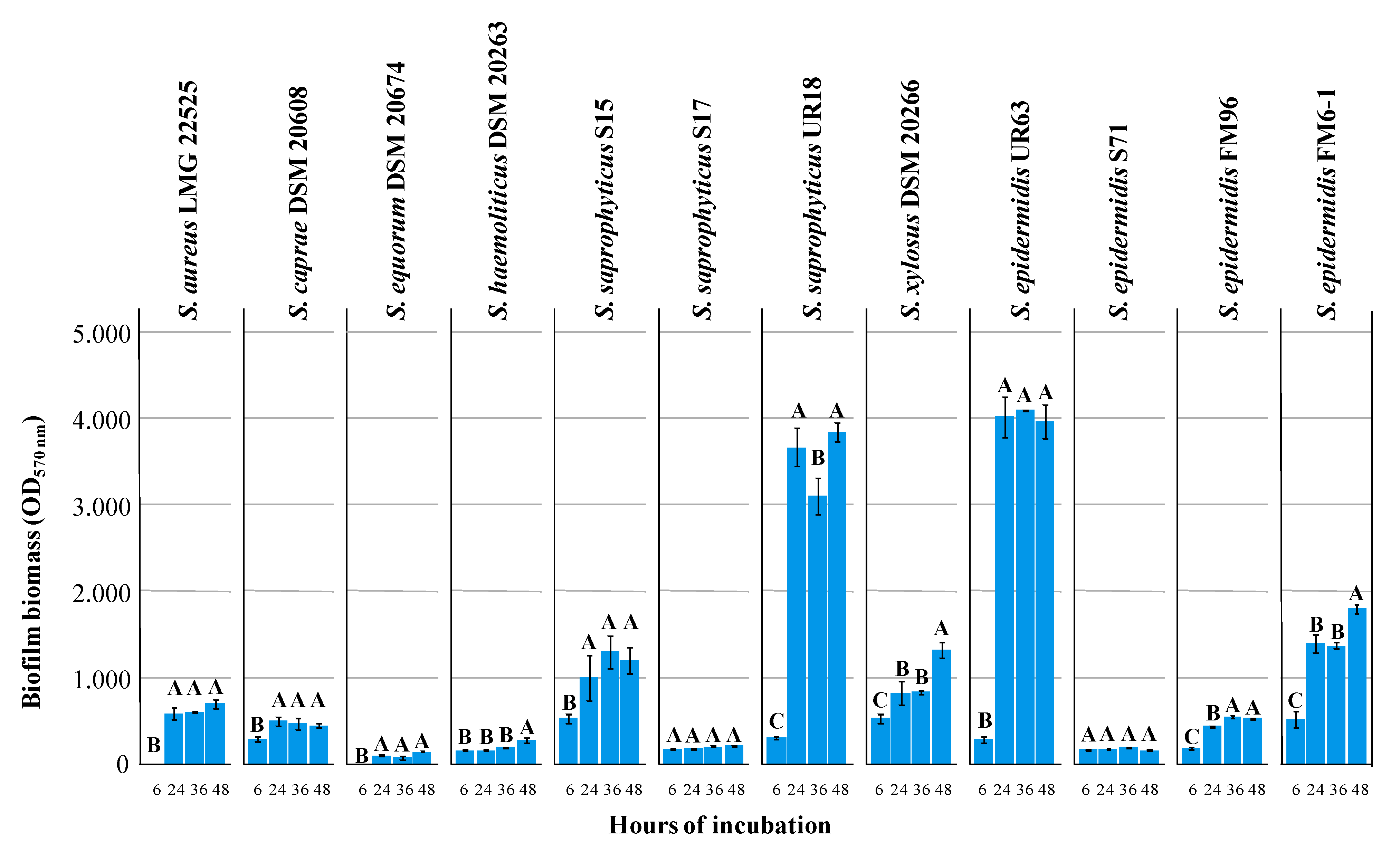
3.1. Biofilm Formation and Minimal Inhibitory Concentration (MIC) of Lactoferrin Hydrolysate (HLF) against Selected Strains
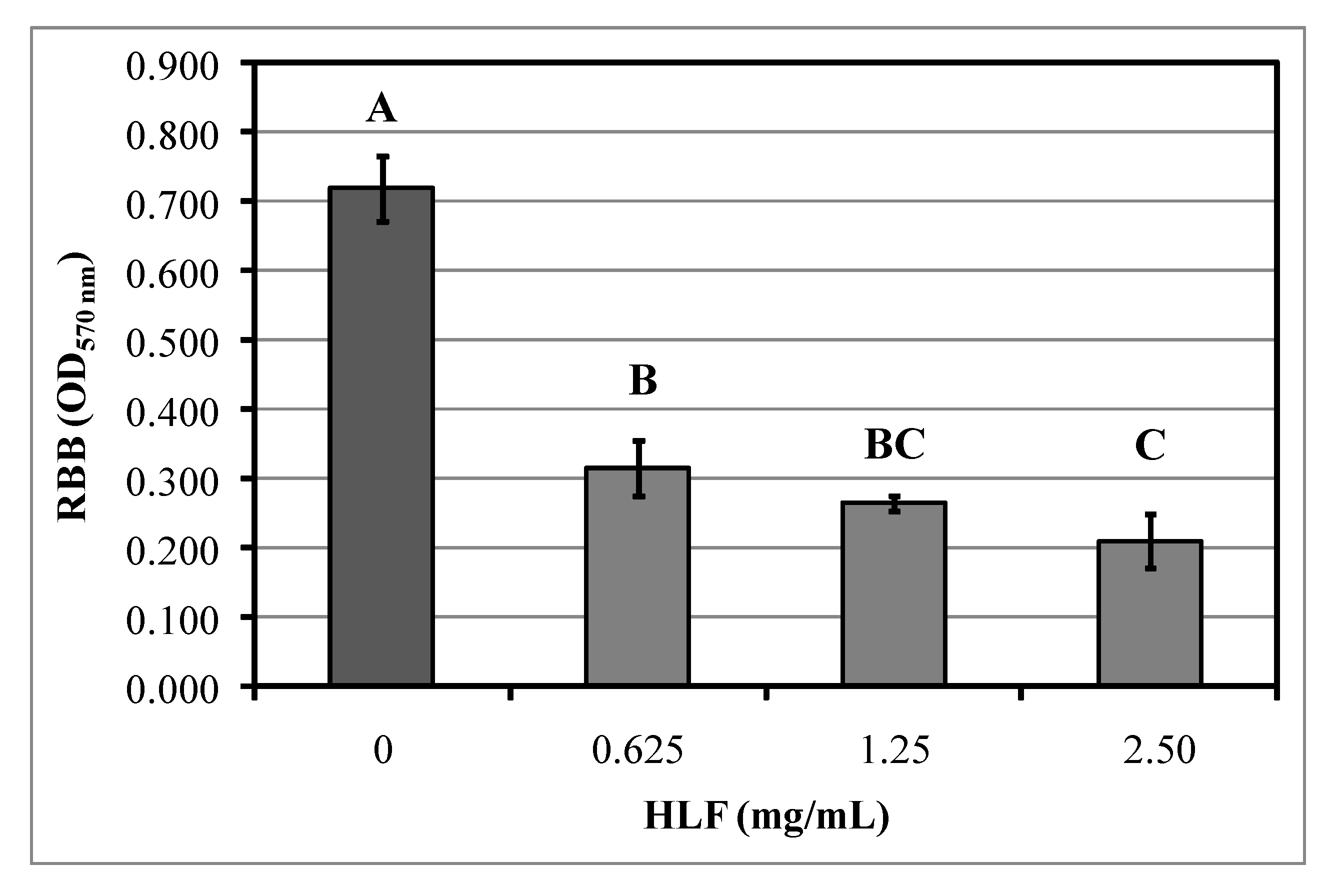
3.2. Identification of Peptides Endowed with Antimicrobial Activity
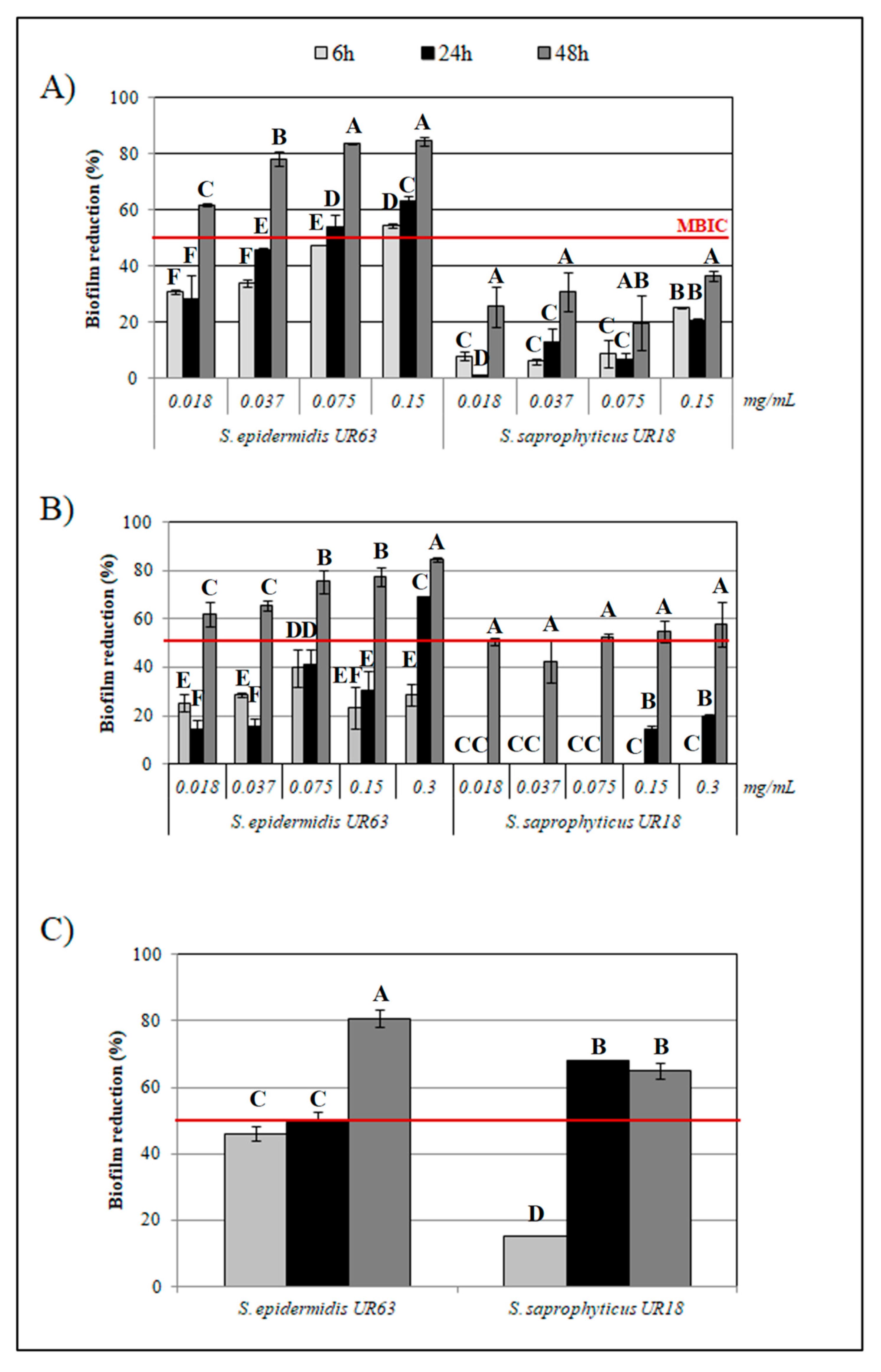
3.3. Antibiofilm Activity by HLF and Synthetic Peptides
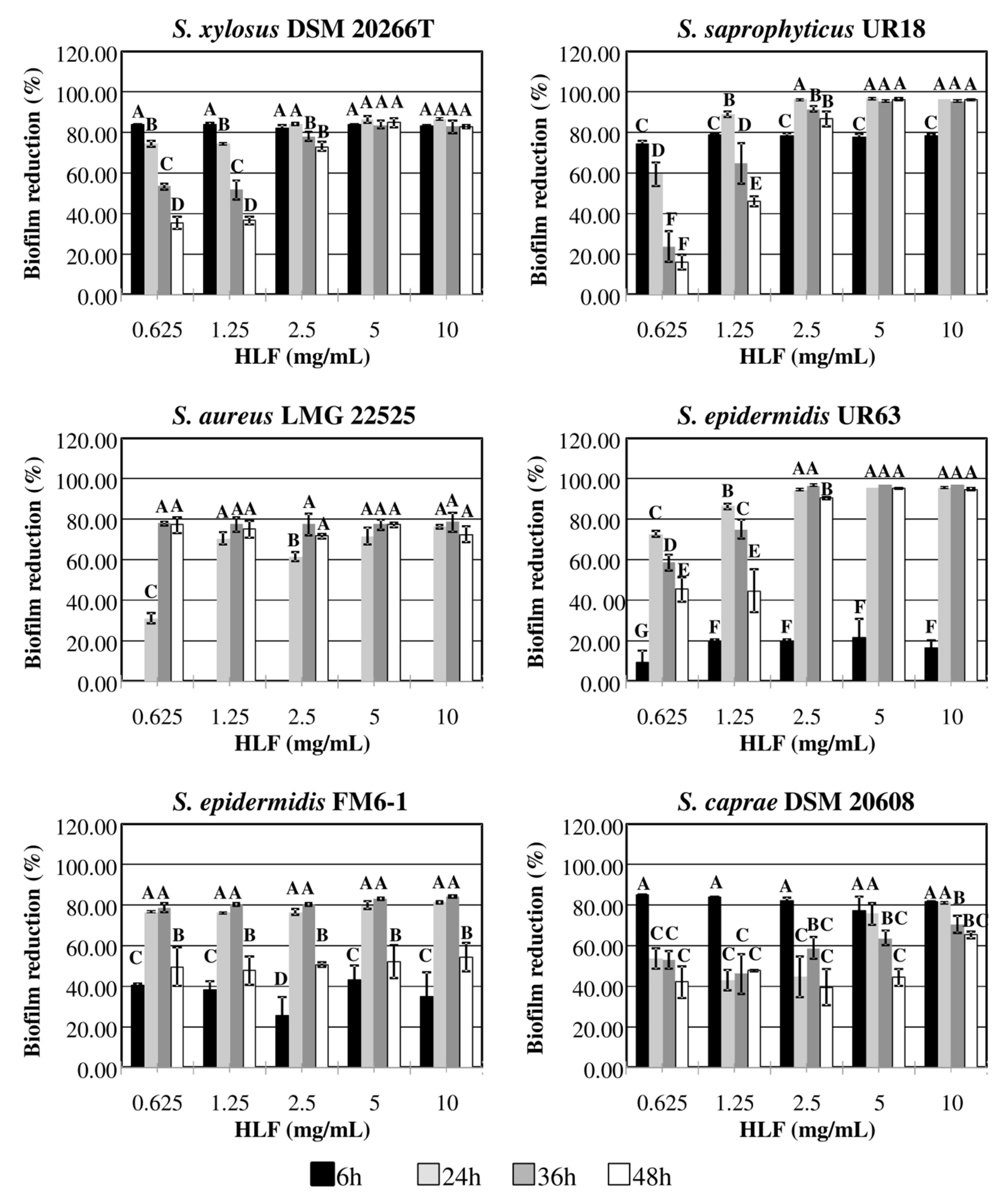
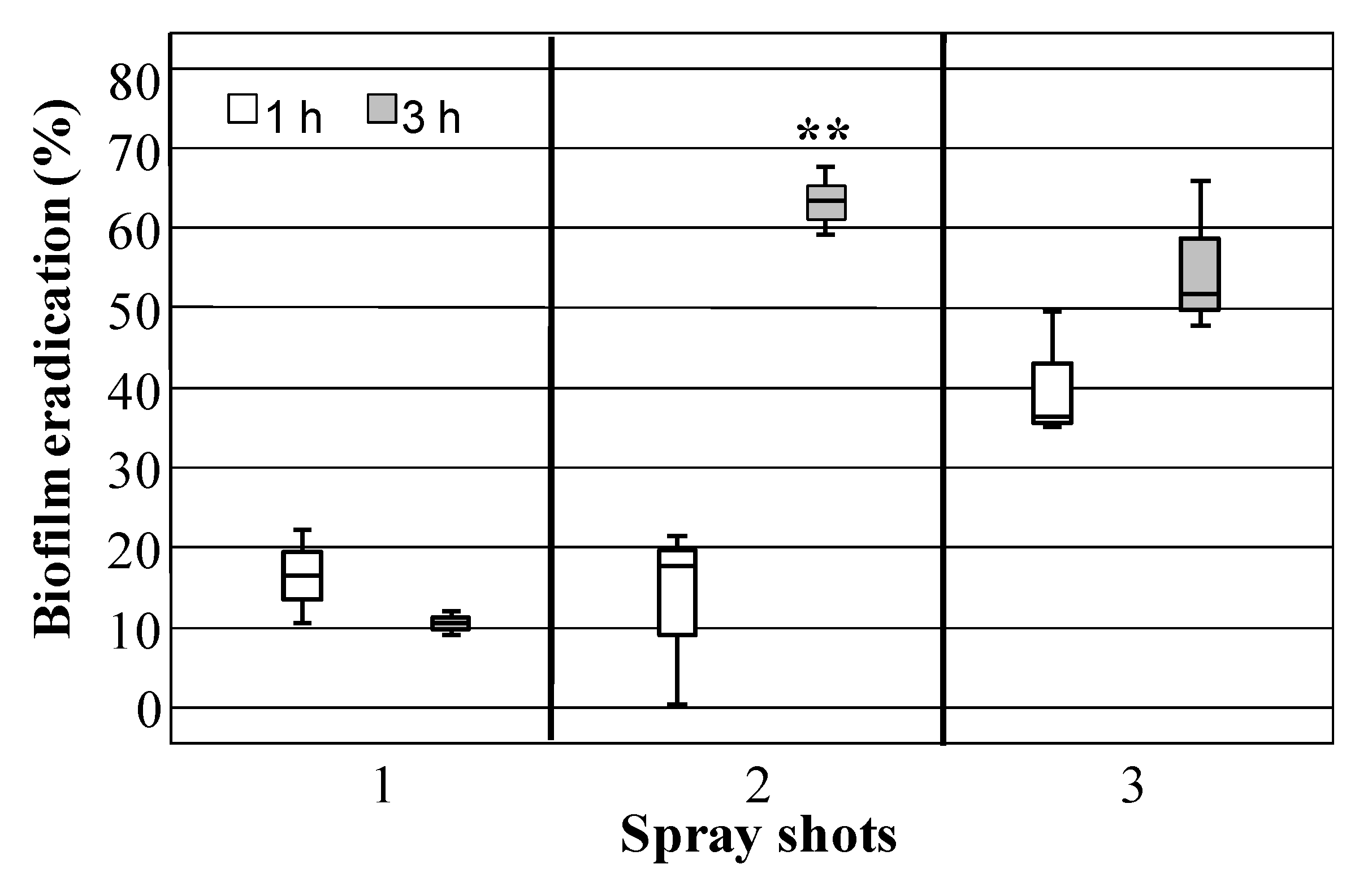
3.4. Staphylococcal Biofilm Eradication by HLF
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CoPS | coagulase positive pathogens strains |
| CoNS | coagulase negative strains |
| AR | antibiotic resistance |
| AMPs | Antimicrobial peptides |
| BLF | Bovine Lactoferrin |
| HLF | Bovine Lactoferrin hydrolysate |
| LFcinB | lactoferricin B |
| LFmpin | lactoferrampin |
| CV | crystal violet |
| OD | optical density |
| MIC | Minimal inhibitory concentration |
| MBC | minimum bactericidal concentration |
| MBIC | Minimum Biofilm Inhibitory Concentration |
| MBEC | Minimal biofilm-eradication concentration |
| LSD | Least significant difference |
References
- Foster, T. Staphylococcus. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch: Galveston, TX, USA, 1996; Chapter 12. Available online: https://www.ncbi.nlm.nih.gov/books/NBK8448/ (accessed on 26 June 2020).
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, Y.S.; Yang, J.Y.; Lee, S.H.; Park, Y.Y.; Kweon, M.N. The resident pathobiont Staphylococcus xylosus in Nfkbiz-deficient skin accelerates spontaneous skin inflammation. Sci. Rep. 2017, 7, 6348. [Google Scholar] [CrossRef]
- Percival, S.L.; Emanuel, C.; Cutting, K.F.; Williams, D.W. Microbiology of the skin and the role of biofilms in infection. Int. Wound J. 2012, 9, 14–32. [Google Scholar] [CrossRef]
- Park, Y.J.; Kim, C.W.; Lee, H.K. Interactions between Host Immunity and Skin-Colonizing Staphylococci: No Two Siblings Are Alike. Int. J. Mol. Sci. 2019, 20, 718. [Google Scholar] [CrossRef]
- Chen, Y.E.; Fischbach, M.A.; Belkaid, Y. Skin microbiota–host interactions. Nature 2018, 553, 427–436. [Google Scholar] [CrossRef]
- Musthaq, S.; Mazuy, A.; Jakus, J. The microbiome in dermatology. Clin. Dermatol. 2018, 36, 390–398. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcus epidermidis—the accidental pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef]
- Minhas, G.S.; Bawdon, D.; Herman, R.; Rudden, M.; Stone, A.P.; James, A.G.; Thomas, G.H.; Newstead, S. Structural basis of malodour precursor transport in the human axilla. eLife 2018, 7, e34995. [Google Scholar] [CrossRef]
- Lopedota, A.; Cutrignelli, A.; Laquintana, V.; Franco, M.; Donelli, D.; Ragni, L.; Tongiani, S.; Denora, N. β-cyclodextrin in personal care formulations: Role on the complexation of malodours causing molecules. Int. J. Cosm. Sci. 2015, 37, 438–445. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcal Biofilms. In Gram-Positive Pathogens, 3nd ed.; Fischetti, V.A., Novick, R.P., Ferretti, J.J., Portnoy, D.A., Braunstein, M., Rood, J.I., Eds.; ASM Press: Washington, DC, USA, 2019; Chapter 43; pp. 699–711. [Google Scholar]
- Yong, Y.Y.; Dykes, G.A.; Choo, W.S. Biofilm formation by staphylococci in health-related environments and recent reports on their control using natural compounds. Crit. Rev. Microbiol. 2019, 45, 201–222. [Google Scholar] [CrossRef] [PubMed]
- Brandwein, M.; Steinberg, D.; Meshner, S. Microbial biofilms and the human skin microbiome. NPJ Biofilms Microbiomes 2016, 2, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ashrafi, M.; Novak-Frazer, L.; Morris, J.; Baguneid, M.; Rautemaa-Richardson, R.; Bayat, A. Electrical stimulation disrupts biofilms in a human wound model and reveals the potential for monitoring treatment response with volatile biomarkers. Wound Repair Regen. 2019, 27, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Ashrafi, M.; Novak-Frazer, L.; Bates, M.; Baguneid, M.; Alonso-Rasgado, T.; Xia, G.; Rautemaa-Richardson, R.; Bayat, A. Validation of biofilm formation on human skin wound models and demonstration of clinically translatable bacteria-specific volatile signatures. Sci. Rep. 2018, 8, 9431–9446. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.; Russo, J.; Fiegel, J.; Brogden, N. Antibiotic delivery strategies to treat skin infections when innate antimicrobial defense Fails. Antibiotics 2020, 9, 56. [Google Scholar] [CrossRef]
- Voegele, P.; Badiola, J.; Schmidt-Malan, S.M.; Karau, M.J.; Greenwood-Quaintance, K.E.; Mandrekar, J.N.; Patel, R. Antibiofilm activity of electrical current in a catheter model. Antimicrob.Agents Chemother. 2016, 60, 1476–1480. [Google Scholar] [CrossRef]
- Caputo, L.; Quintieri, L.; Cavalluzzi, M.M.; Lentini, G.; Habtemariam, S. Antimicrobial and antibiofilm activities of citrus water-extracts obtained by microwave-assisted and conventional methods. Biomedicines 2018, 6, 70. [Google Scholar] [CrossRef]
- Morán, A.; Gutiérrez, S.; Martínez-Blanco, H.; Ferrero, M.A.; Monteagudo-Mera, A.; Rodríguez-Aparicio, L.B. Non-toxic plant metabolites regulate Staphylococcus viability and biofilm formation: A natural therapeutic strategy useful in the treatment and prevention of skin infections. Biofouling 2014, 30, 1175–1182. [Google Scholar] [CrossRef]
- Ricciardelli, A.; Casillo, A.; Papa, R.; Monti, D.M.; Imbimbo, P.; Vrenna, G.; Artini, M.; Selan, L.; Corsaro, M.M.; Tutino, M.L.; et al. Pentadecanal inspired molecules as new anti-biofilm agents against Staphylococcus epidermidis. Biofouling 2018, 34, 1110–1120. [Google Scholar] [CrossRef]
- Banerjee, S.; Vishakha, K.; Das, S.; Dutta, M.; Mukherjee, D.; Mondal, J.; Ganguli, A. Antibacterial, anti-biofilm activity and mechanism of action of Pancreatin doped Zinc oxide nanoparticles against methicillin resistant Staphylococcus aureus. Colloids Surf. B 2020, 110921. [Google Scholar] [CrossRef]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial peptides and their therapeutic potential for bacterial skin infections and wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef] [PubMed]
- Galdiero, E.; Lombardi, L.; Falanga, A.; Libralato, G.; Guida, M.; Carotenuto, R. Biofilms: Novel strategies based on antimicrobial peptides. Pharmaceutics 2019, 11, 322. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.D.; Sothiselvam, S.; Lu, T.K.; de la Fuente-Nunez, C. Peptide design principles for antimicrobial applications. J. Mol. Biol. 2019, 431, 3547–3567. [Google Scholar] [CrossRef] [PubMed]
- Maccari, G.; Di Luca, M.; Nifosì, R. In silico design of antimicrobial peptides. In Computational Peptidology, 1st ed.; Zhou, P., Huang, J., Eds.; Humana Press: New York, NY, USA, 2015; pp. 195–219. [Google Scholar]
- Caputo, L.; Quintieri, L.; Morea, M.; Baruzzi, F. Antimicrobial activity of a meat-borne Bacillus subtilis strain against food pathogens. Eur. Food Res. Technol. 2011, 232, 183–189. [Google Scholar] [CrossRef]
- Waghu, F.H.; Idicula-Thomas, S. Collection of antimicrobial peptides database and its derivatives: Applications and beyond. Protein Sci. 2020, 29, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.; García-Vaquero, M. Bioactive compounds from fermented food products. In Novel Food Fermentation Technologies, 1st ed.; Ojha, K.S., Tiwari, B.K., Eds.; Springer International Publishing AG: Cham, Switzerland, 2016; pp. 293–310. [Google Scholar]
- Quintieri, L.; Zühlke, D.; Fanelli, F.; Caputo, L.; Liuzzi, V.C.; Logrieco, A.F.; Hirschfeld, C.; Becher, D.; Riedel, K. Proteomic analysis of the food spoiler Pseudomonas fluorescens ITEM 17298 reveals the antibiofilm activity of the pepsin-digested bovine lactoferrin. Food Microbiol. 2019, 82, 177–193. [Google Scholar] [CrossRef]
- Quintieri, L.; Fanelli, F.; Zühlke, D.; Caputo, L.; Logrieco, A.F.; Albrecht, D.; Riedel, K. Biofilm and pathogenesis-related proteins in the foodborne P. fluorescens ITEM 17298 with distinctive phenotypes during cold storage. Front. Microbiol. 2020, 11, 991. [Google Scholar] [CrossRef]
- Cosseau, C.; Romano-Bertrand, S.; Duplan, H.; Lucas, O.; Ingrassia, I.; Pigasse, C.; Roques, C.; Jumas-Bilak, E. Proteobacteria from the human skin microbiota: Species-level diversity and hypotheses. One Health 2016, 2, 33–41. [Google Scholar] [CrossRef]
- Quintieri, L.; Caputo, L.; Monaci, L.; Deserio, D.; Morea, M.; Baruzzi, F. Antimicrobial efficacy of pepsin-digested bovine lactoferrin on spoilage bacteria contaminating traditional mozzarella cheese. Food Microbiol. 2012, 31, 64–71. [Google Scholar] [CrossRef]
- Ammons, M.C.; Copié, V. Mini-review: Lactoferrin: A bioinspired, anti-biofilm therapeutic. Biofouling 2013, 29, 443–455. [Google Scholar] [CrossRef]
- Takayama, Y.; Aoki, R. Roles of lactoferrin on skin wound healing. Biochem. Cell Biol. 2011, 90, 497–503. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter dish biofilm formation assay. JoVE 2011, 47, e2437. [Google Scholar] [CrossRef]
- European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Diseases (ESCMID). Determination of minimum inhibitory concentrations (MICs) of antibacterial agents by broth dilution. Clin. Microbiol. Infect. 2003, 9, ix–xv. [Google Scholar] [CrossRef]
- Bakkiyaraj, D.; Sritharadol, R.; Padmavathi, A.R.; Nakpheng, T.; Srichana, T. Anti-biofilm properties of a mupirocin spray formulation against Escherichia coli wound infections. Biofouling 2017, 33, 591–600. [Google Scholar] [CrossRef]
- Packiavathy, I.A.; Priya, S.; Pandian, S.K.; Ravi, A.V. Inhibition of biofilm development of uropathogens by curcumin—an anti-quorum sensing agent from Curcuma longa. Food Chem. 2014, 148, 453–460. [Google Scholar] [CrossRef]
- Kosikowska, P.; Lesner, A. Antimicrobial peptides (AMPs) as drug candidates: A patent review (2003–2015). Expert Opin. Ther. Pat. 2016, 26, 689–702. [Google Scholar] [CrossRef]
- Fleischer, D.M.; Venter, C.; Vandenplas, Y. Hydrolyzed formula for every infant? In Protein in Neonatal and Infant Nutrition: Recent Updates; Bathia, J., Shamir, R., Vandeplas, Y., Eds.; Karger: Basel, Switzerland, 2016; Volume 86, pp. 51–65. [Google Scholar]
- Lima, T.N.; PedrialiMoraes, C.A. Bioactive peptides: Applications and relevance for cosmeceuticals. Cosmetics 2018, 5, 21. [Google Scholar] [CrossRef]
- Suwal, S.; Rozoy, E.; Manenda, M.; Doyen, A.; Bazinet, L. Comparative study of in situ and ex situ enzymatic hydrolysis of milk protein and separation of bioactive peptides in an electromembrane reactor. ACS Sustain. Chem. Eng. 2017, 5, 5330–5340. [Google Scholar] [CrossRef]
- Batoni, G.; Maisetta, G.; Esin, S. Antimicrobial peptides and their interaction with biofilms of medically relevant bacteria. BBA-Biomembranes 2016, 1858, 1044–1060. [Google Scholar] [CrossRef]
- Méric, G.; Mageiros, L.; Pensar, J.; Laabei, M.; Yahara, K.; Pascoe, B.; Kittiwan, N.; Tadee, P.; Post, V.; Lamble, S.; et al. Disease-associated genotypes of the commensal skin bacterium Staphylococcus epidermidis. Nat. Commun. 2018, 9, 5034. [Google Scholar] [CrossRef] [PubMed]
- Deutch, C.E. Inhibition of urease activity in the urinary tract pathogens Staphylococcus saprophyticus and Proteus mirabilis by dimethylsulfoxide (DMSO). J. Appl. Microbiol. 2019, 128, 1514–1523. [Google Scholar] [CrossRef] [PubMed]
- Gowda, A.; Pensiero, A.L.; Packer, C.D. Staphylococcus caprae: A Skin Commensal with Pathogenic Potential. Cureus 2018, 10, e3485. [Google Scholar] [CrossRef] [PubMed]
- Quintieri, L.; Caputo, L.; Morea, M.; Baruzzi, F. Control of Mozzarella spoilage bacteria by using bovine lactoferrin pepsin-digested hydrolysate. In Worldwide Research Efforts in the Fighting against Microbial Pathogens: From Basic Research to Technological Developments; Mendèz-Vilas, A., Ed.; Brown Walker Press: Boca Raton, FL, USA, 2013; pp. 118–122. [Google Scholar]
- Quintieri, L.; Carito, A.; Pinto, L.; Calabrese, N.; Baruzzi, F.; Caputo, L. Application of lactoferricin B to control microbial spoilage in cold stored fresh foods. In Multidisciplinary Approach for Studying and Combating Microbial Pathogens; Mendèz-Vilas, A., Ed.; Microbiology series volume 3; Brown Walker Press: Boca Raton, FL, USA, 2015; pp. 58–62. [Google Scholar]
- Chen, P.W.; Jheng, T.T.; Shyu, C.L.; Mao, F.C. Synergistic antibacterial efficacies of the combination of bovine lactoferrin or its hydrolysate with probiotic secretion in curbing the growth of meticillin-resistant Staphylococcus aureus. J. Med. Microbiol. 2013, 62, 1845–1851. [Google Scholar] [CrossRef] [PubMed]
- van der Kraan, M.I.; Nazmi, K.; Teeken, A.; Groenink, J.; van ‘t Hof, W.; Veerman, E.C.; Bolscher, J.G.; NieuwAmerongen, A.V. Lactoferrampin, an antimicrobial peptide of bovine lactoferrin, exerts its candidacidal activity by a cluster of positively charged residues at the C-terminus in combination with a helix-facilitating N-terminal part. Biol. Chem. 2005, 386, 137–142. [Google Scholar] [CrossRef]
- van der Kraan, M.I.; Nazmi, K.; van ‘t Hof, W.; Nieuw Amerongen, A.V.; Veerman, E.C.; Bolscher, J.G. Distinct bactericidal activities of bovine lactoferrin peptides LFampin 268-284 and LFampin 265-284: Asp-Leu-Ile makes a difference. Biochem. Cell Biol. 2006, 84, 358–362. [Google Scholar] [CrossRef]
- Bolscher, J.G.; van der Kraan, M.I.; Nazmi, K.; Kalay, H.; Grün, C.H.; van’t Hof, W. A one-enzyme strategy to release an antimicrobial peptide from the LFampin-domain of bovine lactoferrin. Peptides 2006, 27, 1–9. [Google Scholar] [CrossRef]
- Cruz, J.; Ortiz, C.; Guzmán, F.; Cárdenas, C.; Fernandez-Lafuente, R.; Torres, R. Design and activity of novel lactoferrampin analogues against O157: H7 enterohemorrhagic Escherichia coli. Biopolymers 2014, 101, 319–328. [Google Scholar] [CrossRef]
- Ruangcharoen, S.; Suwannarong, W.; Lachica, M.R.C.T.; Bolscher, J.G.; Nazmi, K.; Khunkitti, W.; Taweechaisupapong, S. Killing activity of LFchimera on periodontopathic bacteria and multispecies oral biofilm formation in vitro. World J. Microb. Biot. 2017, 33, 167. [Google Scholar] [CrossRef]
- Morici, P.; Fais, R.; Rizzato, C.; Tavanti, A.; Lupetti, A. Inhibition of Candida albicans biofilm formation by the synthetic lactoferricin derived peptide hLF1-11. PLoS ONE 2016, 11, e0167470. [Google Scholar] [CrossRef]
- Sánchez-Gómez, S.; Ferrer-Espada, R.; Stewart, P.S.; Pitts, B.; Lohner, K.; de Tejada, G.M. Antimicrobial activity of synthetic cationic peptides and lipopeptides derived from human lactoferricin against Pseudomonas aeruginosa planktonic cultures and biofilms. BMC Microbiol. 2015, 15, 137. [Google Scholar] [CrossRef]
- Svendsen, J.S.M.; Grant, T.M.; Rennison, D.; Brimble, M.A.; Svenson, J. Very short and stable lactoferricin-derived antimicrobial peptides: Design principles and potential uses. Accounts Chem. Res. 2019, 52, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Raksha, L.; Gangashettappa, N.; Shantala, G.B.; Nandan, B.R.; Sinha, D. Study of biofilm formation in bacterial isolates from contact lens wearers. Indian J. Ophthalmol. 2020, 68, 23. [Google Scholar] [PubMed]
- Xiao, A.; Dhand, C.; Leung, C.M.; Beuerman, R.W.; Ramakrishna, S.; Lakshminarayanan, R. Strategies to design antimicrobial contact lenses and contact lens cases. J. Mater. Chem. B 2018, 6, 2171–2186. [Google Scholar] [CrossRef] [PubMed]
- Sharadha, M.; Gowda, D.V.; Gupta, V.; Akhila, A.R. An overview on topical drug delivery system–Updated review. Int. J. Res. Pharm. Sci. 2020, 11, 368–385. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HLF (mg/mL) | ||
|---|---|---|
| Strains | MIC | MBC |
| S. epidermidis FM6–1 | 20 | 20 |
| S. epidermidis UR63 | 10 | 10 |
| S. saprophyticus UR18 | 20 | >20 |
| S. xylosus DSM 20266T | 20 | >20 |
| S. aureus LMG 22525 | >20 | >20 |
| S. caprae DSM 20608 | 20 | 20 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quintieri, L.; Caputo, L.; Monaci, L.; Cavalluzzi, M.M.; Denora, N. Lactoferrin-Derived Peptides as a Control Strategy against Skinborne Staphylococcal Biofilms. Biomedicines 2020, 8, 323. https://doi.org/10.3390/biomedicines8090323
Quintieri L, Caputo L, Monaci L, Cavalluzzi MM, Denora N. Lactoferrin-Derived Peptides as a Control Strategy against Skinborne Staphylococcal Biofilms. Biomedicines. 2020; 8(9):323. https://doi.org/10.3390/biomedicines8090323
Chicago/Turabian StyleQuintieri, Laura, Leonardo Caputo, Linda Monaci, Maria Maddalena Cavalluzzi, and Nunzio Denora. 2020. "Lactoferrin-Derived Peptides as a Control Strategy against Skinborne Staphylococcal Biofilms" Biomedicines 8, no. 9: 323. https://doi.org/10.3390/biomedicines8090323
APA StyleQuintieri, L., Caputo, L., Monaci, L., Cavalluzzi, M. M., & Denora, N. (2020). Lactoferrin-Derived Peptides as a Control Strategy against Skinborne Staphylococcal Biofilms. Biomedicines, 8(9), 323. https://doi.org/10.3390/biomedicines8090323