Effects of Decursin and Angelica gigas Nakai Root Extract on Hair Growth in Mouse Dorsal Skin via Regulating Inflammatory Cytokines



Abstract
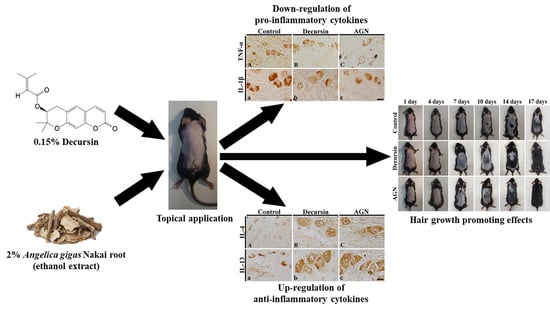
1. Introduction
2. Materials and Methods
2.1. Preparation of Decursin and AGN Root Extract
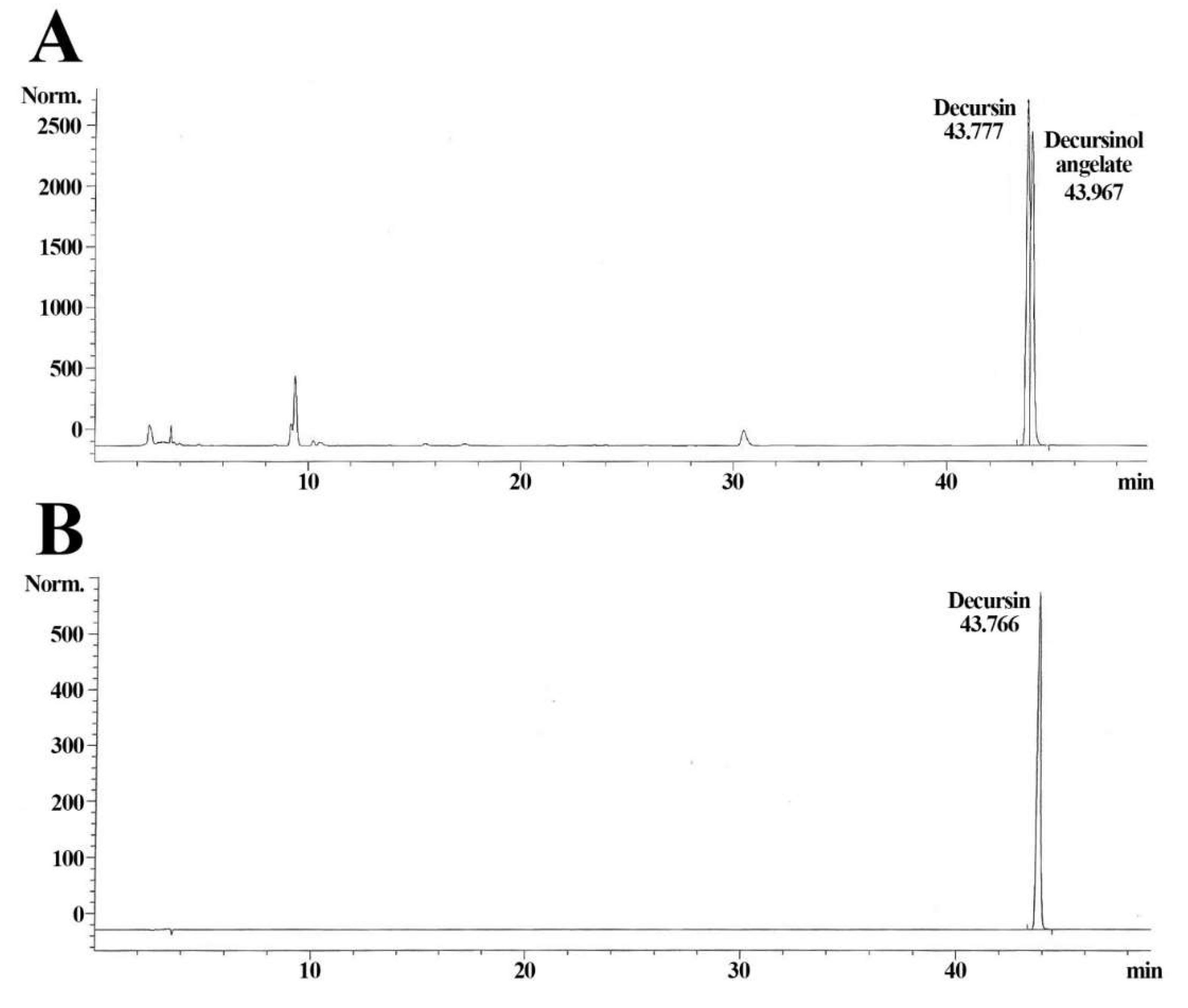
2.2. Qualitative Analysis of AGN Extract
2.3. Experimental Animals
2.4. Experimental Groups and Treatments of Decursin and AGN Extract
2.5. Western Blot Analysis
2.6. Preparation of Tissue Sections
2.7. Hematoxylin and Eosin (H&E) Staining
2.8. Immunohistochemistry
2.9. Data Analysis
2.10. Statistical Analysis
3. Results
3.1. Decursin of AGN Extract
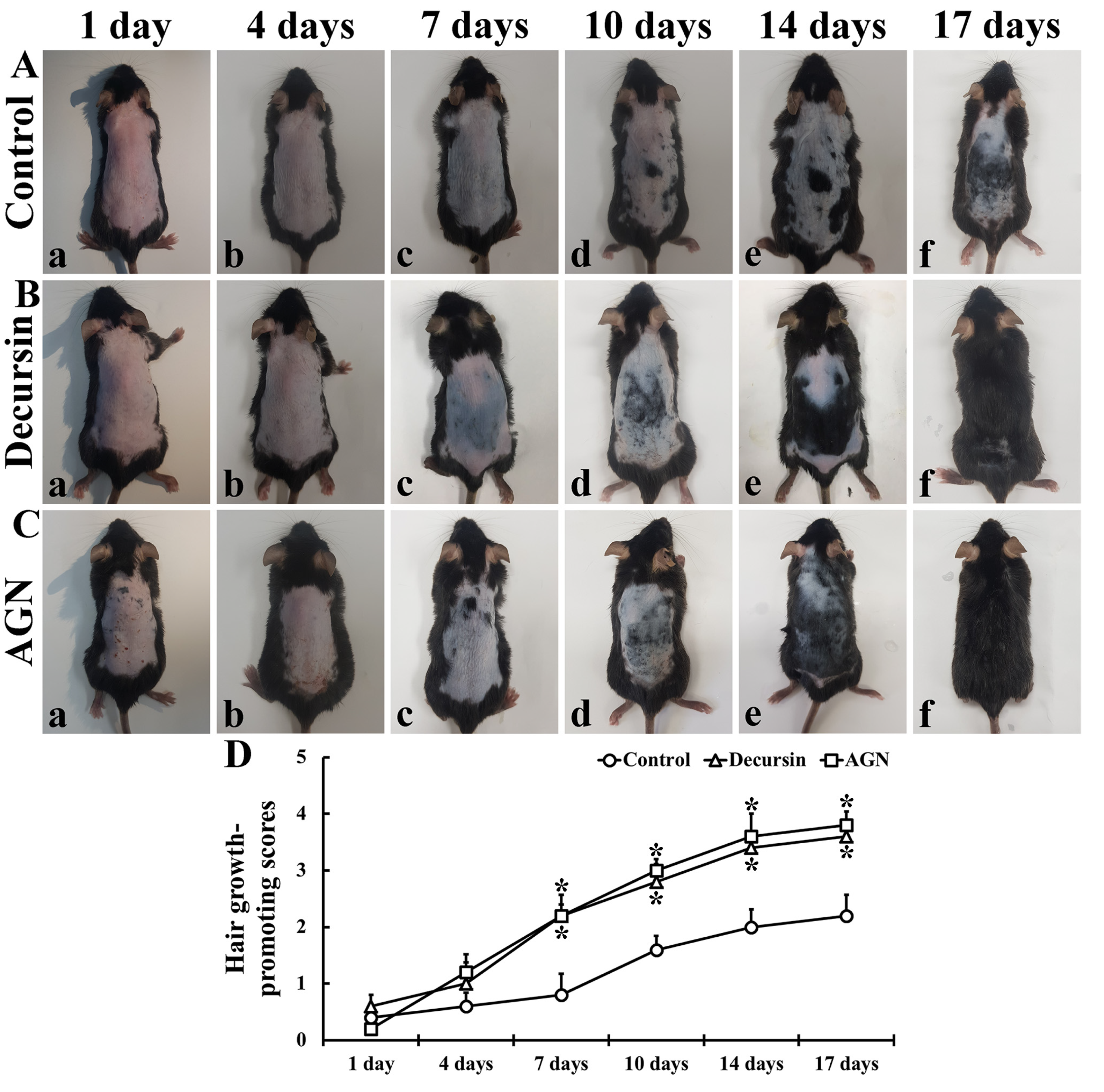
3.2. Hair Growth
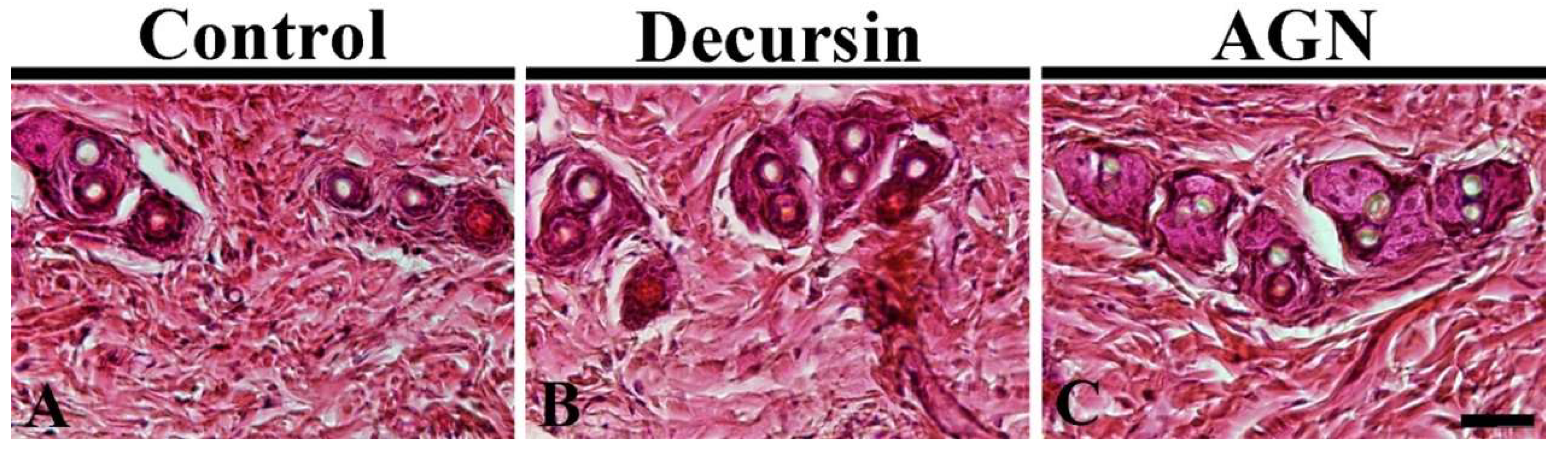
3.3. Hair Follicles
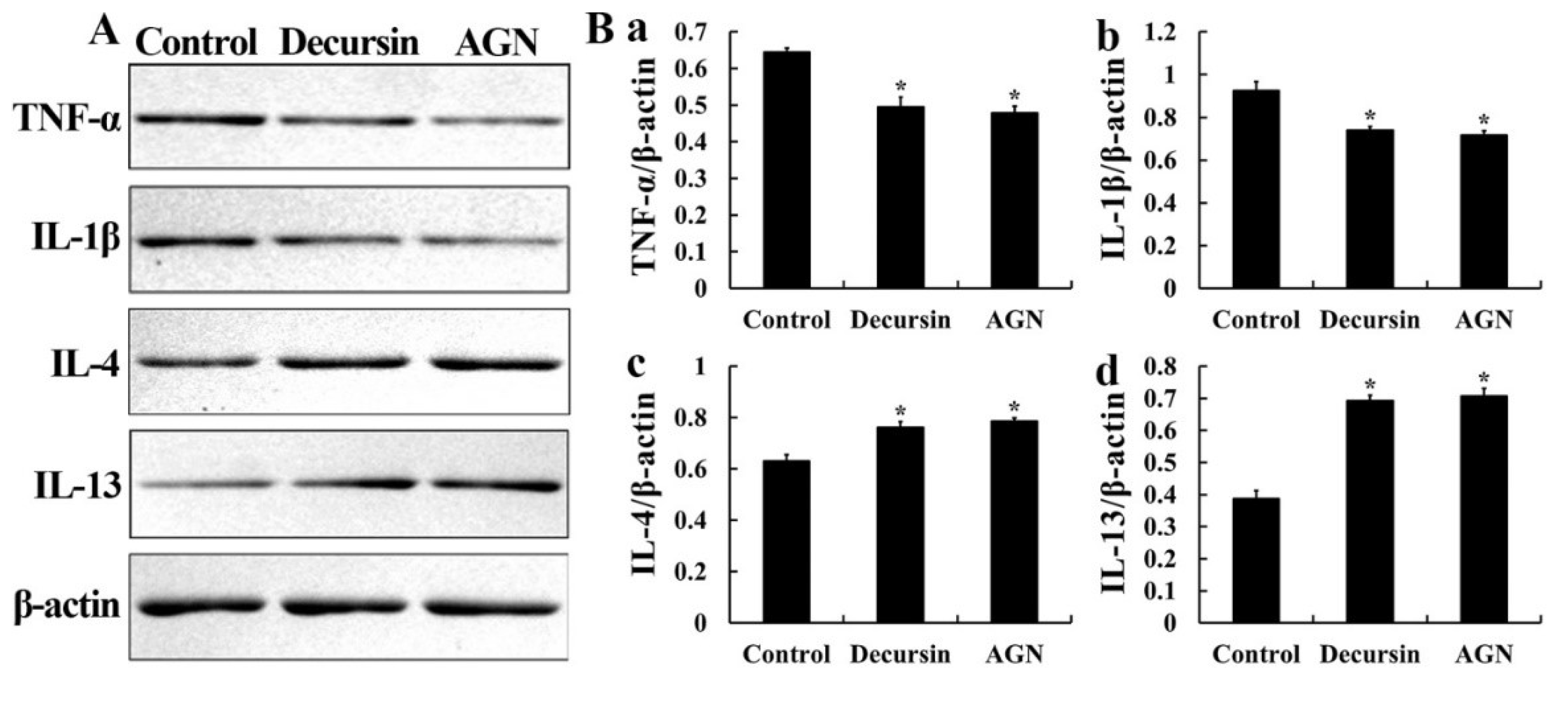
3.4. Levels of Pro- and Anti-Inflammatory Cytokines
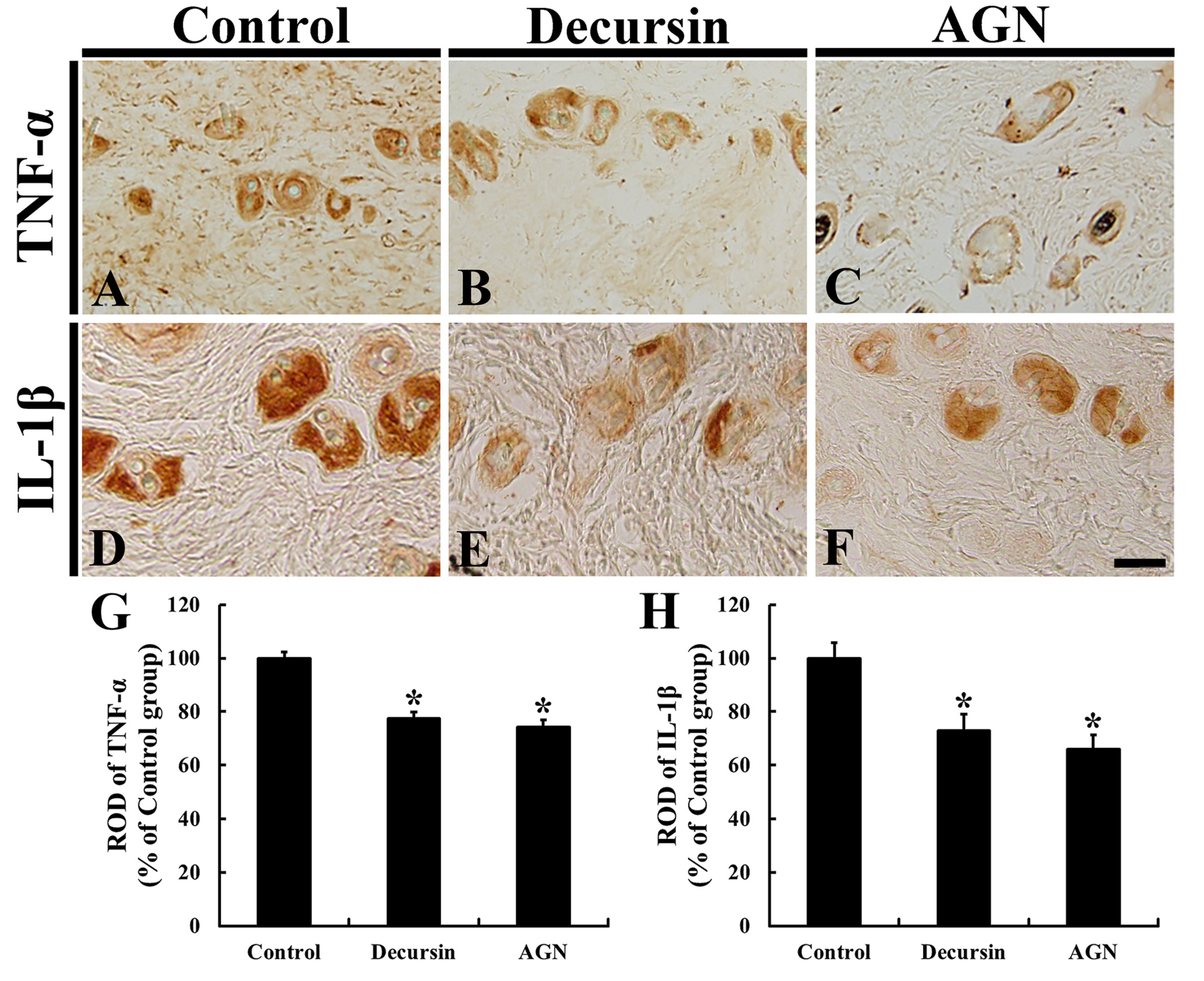
3.5. Pro-Inflammatory Cytokines
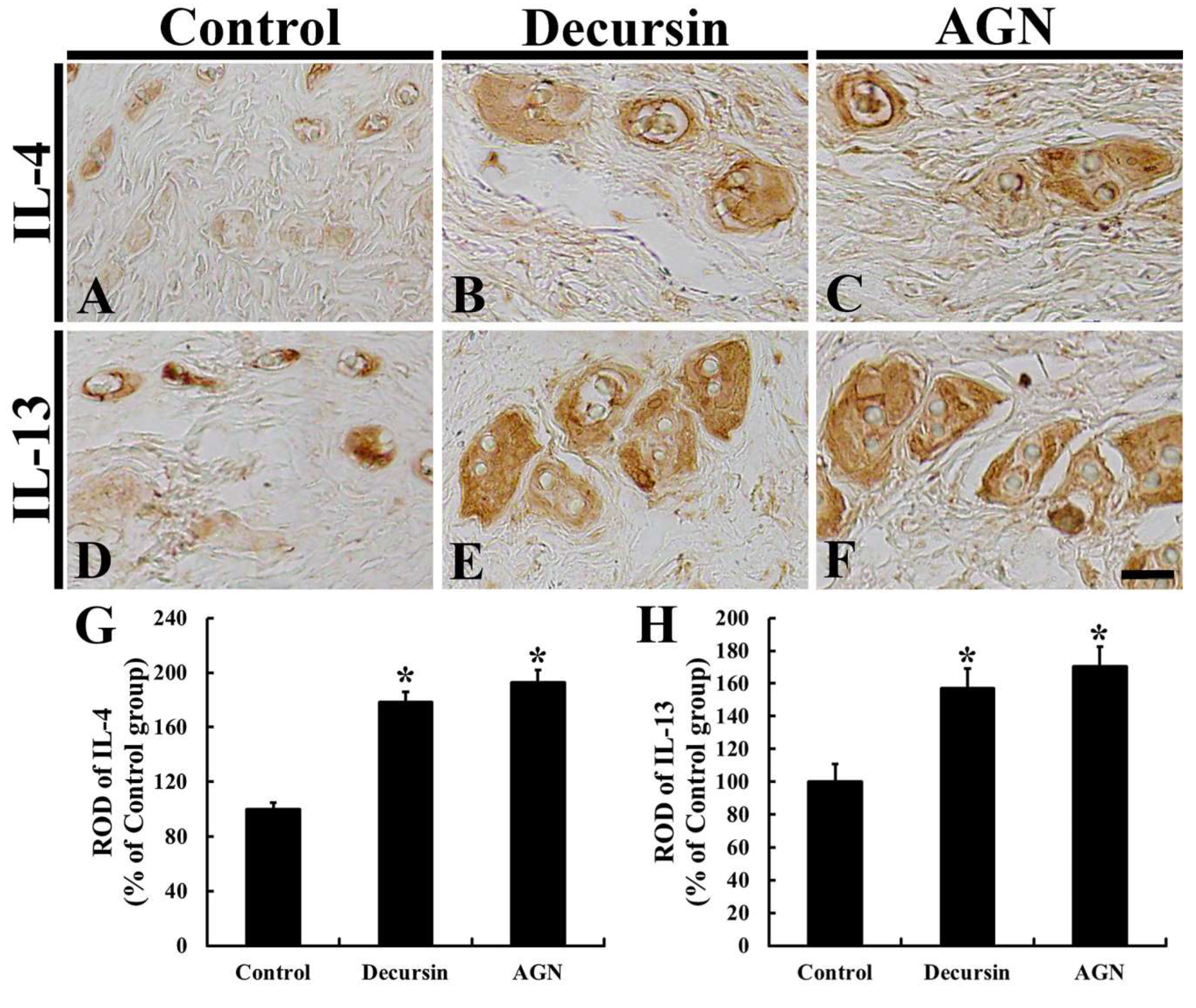
3.6. Anti-Inflammatory Cytokines
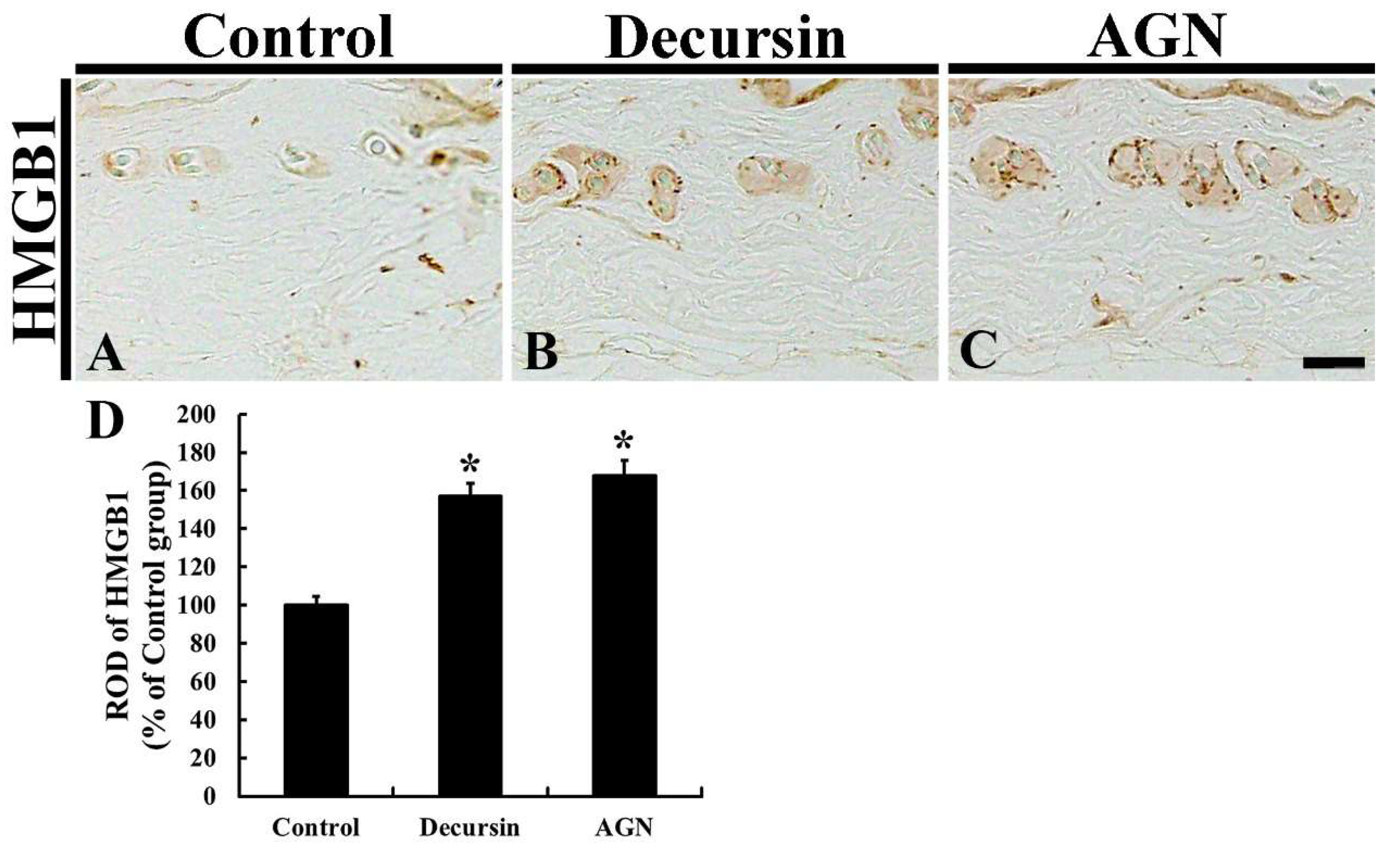
3.7. HMGB1 Immunoreactivity
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- O’Rahilly, R.; Müller, F. Basic Human Anatomy: A Regional Study of Human Structure; WB Saunders Company: Philadelphia, PA, USA, 1983. [Google Scholar]
- Rushton, D.H.; Norris, M.J.; Dover, R.; Busuttil, N. Causes of hair loss and the developments in hair rejuvenation. Int. J. Cosmet. Sci. 2002, 24, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Miao, Y.; Li, X.; Qu, Q.; Liu, Y.; Li, K.; Feng, C.; Hu, Z. The relationship between self-esteem and hair transplantation satisfaction in male androgenetic alopecia patients. J. Cosmet. Dermatol. 2019, 18, 1441–1447. [Google Scholar] [CrossRef] [PubMed]
- Phillips, T.G.; Slomiany, W.P.; Allison, R. Hair loss: Common causes and treatment. Am. Fam. Physician 2017, 96, 371–378. [Google Scholar] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, L.; Harris, M.C.; Kilpatrick, L.E. Cytokines and inflammatory response in the fetus and neonate. In Fetal and Neonatal Physiology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1241–1254.e1244. [Google Scholar]
- Opal, S.M.; DePalo, V.A. Anti-inflammatory cytokines. Chest 2000, 117, 1162–1172. [Google Scholar] [CrossRef]
- Khan, H.A.; Khan, I.; Lee, Y.-K. Role of immune factors in bioavailability and toxicity of carbon nanomaterials. In Fullerens, Graphenes and Nanotubes; Elsevier: Amsterdam, The Netherlands, 2018; pp. 601–630. [Google Scholar]
- Guo, H.; Cheng, Y.; Shapiro, J.; McElwee, K. The role of lymphocytes in the development and treatment of alopecia areata. Expert Rev. Clin. Immunol. 2015, 11, 1335–1351. [Google Scholar] [CrossRef] [PubMed]
- Carroll, J.M.; Crompton, T.; Seery, J.P.; Watt, F.M. Transgenic mice expressing ifn-gamma in the epidermis have eczema, hair hypopigmentation, and hair loss. J. Investig. Dermatol. 1997, 108, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Paus, R.; Bertolini, M. The role of hair follicle immune privilege collapse in alopecia areata: Status and perspectives. J. Investig. Dermatol. Symp. Proc. 2013, 16, S25–S27. [Google Scholar] [CrossRef] [PubMed]
- Ok, S.; Oh, S.R.; Jung, T.S.; Jeon, S.O.; Jung, J.W.; Ryu, D.S. Effects of angelica gigas nakai as an anti-inflammatory agent in in vitro and in vivo atopic dermatitis models. Evid. -Based Complement. Altern. Med. eCAM 2018, 2018, 2450712. [Google Scholar]
- Cho, J.H.; Kwon, J.E.; Cho, Y.; Kim, I.; Kang, S.C. Anti-inflammatory effect of angelica gigas via heme oxygenase (ho)-1 expression. Nutrients 2015, 7, 4862–4874. [Google Scholar] [CrossRef]
- Bae, U.-J.; Oh, M.-R.; Jung, T.-S.; Chae, S.-W.; Park, B.-H. Decursin and decursinol angelate-rich angelica gigas nakai extract suppresses de novo lipogenesis and alleviates nonalcoholic fatty liver disease and dyslipidemia in mice fed a high fat diet. J. Funct. Foods 2017, 31, 208–216. [Google Scholar] [CrossRef]
- Lee, J.K.; Jeong, J.W.; Jang, T.; Lee, G.W.; Han, H.; Kang, J.S.; Kim, I.H. Decursin attenuates kainic acid-induced seizures in mice. Neuroreport 2014, 25, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Lee, S.W.; Park, H.J.; Lee, S.H.; Im, W.K.; Kim, Y.D.; Kim, K.H.; Park, S.J.; Hong, S.; Jeon, S.H. Anti-cancer activity of angelica gigas by increasing immune response and stimulating natural killer and natural killer t cells. BMC Complement. Altern. Med. 2018, 18, 218. [Google Scholar] [CrossRef]
- Kang, S.Y.; Lee, K.Y.; Park, M.J.; Kim, Y.C.; Markelonis, G.J.; Oh, T.H. Decursin from angelica gigas mitigates amnesia induced by scopolamine in mice. Neurobiol. Learn. Mem. 2003, 79, 11–18. [Google Scholar] [CrossRef]
- Lee, J.; Kim, Y. The promoting effect of angelica gigas nakai and glycyrrhiza uralensis fischet oriental medicine complex extracts on hair growth. J. Cosmetol. Sci 2010, 6, 49–56. [Google Scholar]
- Lee, Y.-G.; Kim, J.-K. The study of the oriental medicine extract on the hair growth effect: I. The effect of the mixture extract of polygoni multiflori radix, angelicae gigantis radix and lycii fructus on the hair growth. Korea J. Herbol. 2004, 19, 83. [Google Scholar]
- Lee, J.Y.; Im, K.R.; Jung, T.K.; Lee, M.-H.; Yoon, K.-S. Medicinal herbal complex extract with potential for hair growth-promoting activity. J. Soc. Cosmet. Sci. Korea 2012, 38, 277–287. [Google Scholar]
- Huang, B.; Kang, B.-G.; Wang, Z.; Lim, S.S. Effect of ethanol extract of plant mixture on hair regeneration in human dermal papilla cells and c57bl/6j mice. J. Med. Plants Res. 2015, 9, 1103–1110. [Google Scholar]
- Ahmed, M.B.; Islam, S.U.; Lee, Y.S. Decursin negatively regulates lps-induced upregulation of the tlr4 and jnk signaling stimulated by the expression of prp4 in vitro. Anim. Cells Syst. 2020, 24, 44–52. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Lee, K.-Y.; Kim, T.-H.; Park, A.; Noh, H.-S.; Kim, S.-C.; Ahn, M.-S. Development and validation of analytical method for decursin in aerial parts of angelica gigas nakai extract. J. Food Hyg. Saf. 2019, 34, 52–57. [Google Scholar] [CrossRef]
- Patel, S.; Nag, M.K.; Sharma, V.; Chauhan, N.S.; Dixit, V. A comparative in vivo and in vitro evaluation of hair growth potential of extracts and an isolate from petroleum ether extract of cuscuta reflexa roxb. Beni-Suef Univ. J. Basic Appl. Sci. 2014, 3, 165–171. [Google Scholar] [CrossRef]
- Roy, R.K.; Thakur, M.; Dixit, V.K. Hair growth promoting activity of eclipta alba in male albino rats. Arch. Dermatol. Res. 2008, 300, 357–364. [Google Scholar] [CrossRef]
- Kim, Y.J.; Choi, M.J.; Bak, D.H.; Lee, B.C.; Ko, E.J.; Ahn, G.R.; Ahn, S.W.; Kim, M.J.; Na, J.; Kim, B.J. Topical administration of egf suppresses immune response and protects skin barrier in dncb-induced atopic dermatitis in nc/nga mice. Sci. Rep. 2018, 8, 11895. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.R.; Zhang, Y.L.; Yap, J.; Boisvert, W.A.; Lee, B.H. Hair growth potential of salvia plebeia extract and its associated mechanisms. Pharm. Biol. 2020, 58, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Boisvert, W.A.; Yu, M.; Choi, Y.; Jeong, G.H.; Zhang, Y.L.; Cho, S.; Choi, C.; Lee, S.; Lee, B.H. Hair growth-promoting effect of geranium sibiricum extract in human dermal papilla cells and c57bl/6 mice. BMC Complement. Altern. Med. 2017, 17, 109. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.-W.; Qu, F.-Z.; Zhang, Y.-M.; Su, G.-Y.; Zhao, Y.-Q. Hair growth promotion effect of cedrol cream and its dermatopharmacokinetics. RSC Adv. 2018, 8, 42170–42178. [Google Scholar] [CrossRef]
- Lu, S.; Xiao, X.; Cheng, M. Matrine inhibits il-1beta-induced expression of matrix metalloproteinases by suppressing the activation of mapk and nf-kappab in human chondrocytes in vitro. Int. J. Clin. Exp. Pathol. 2015, 8, 4764–4772. [Google Scholar]
- Ghasemian, M.; Owlia, S.; Owlia, M.B. Review of anti-inflammatory herbal medicines. Adv. Pharmacol. Sci. 2016, 2016, 9130979. [Google Scholar] [CrossRef]
- Moini Jazani, A.; Hamdi, K.; Tansaz, M.; Nazemiyeh, H.; Sadeghi Bazargani, H.; Fazljou, S.M.B.; Nasimi Doost Azgomi, R. Herbal medicine for oligomenorrhea and amenorrhea: A systematic review of ancient and conventional medicine. BioMed. Res. Int. 2018, 2018, 3052768. [Google Scholar] [CrossRef]
- Oh, S.R.; Ok, S.; Jung, T.S.; Jeon, S.O.; Park, J.M.; Jung, J.W.; Ryu, D.S. Protective effect of decursin and decursinol angelate-rich angelica gigas nakai extract on dextran sulfate sodium-induced murine ulcerative colitis. Asian Pac. J. Trop. Med. 2017, 10, 864–870. [Google Scholar] [CrossRef]
- Shin, S.; Jeon, J.H.; Park, D.; Jang, J.Y.; Joo, S.S.; Hwang, B.Y.; Choe, S.Y.; Kim, Y.B. Anti-inflammatory effects of an ethanol extract of angelica gigas in a carrageenan-air pouch inflammation model. Exp. Anim. 2009, 58, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.J.; Kwon, M.S.; Park, S.H.; Sim, Y.B.; Choi, S.M.; Huh, G.H.; Lee, J.K.; Suh, H.W. The analgesic effect of decursinol. Arch. Pharmcal Res. 2009, 32, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.N.; Khan, I.; Dey, D.K.; Cho, K.H.; Hwang, B.S.; Bae, K.B.; Kang, S.C.; Park, J.G. Decursinol angelate ameliorates 12-o-tetradecanoyl phorbol-13-acetate (tpa) -induced nf-kappab activation on mice ears by inhibiting exaggerated inflammatory cell infiltration, oxidative stress and pro-inflammatory cytokine production. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2019, 132, 110699. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Kim, S.H.; Jiang, C.; Lee, H.; Guo, J. Oriental herbs as a source of novel anti-androgen and prostate cancer chemopreventive agents. Acta Pharmacol. Sin. 2007, 28, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Sowndhararajan, K.; Kim, S. Neuroprotective and cognitive enhancement potentials of angelica gigas nakai root: A review. Sci. Pharm. 2017, 85, 21. [Google Scholar] [CrossRef]
- Islam, S.U.; Lee, J.H.; Shehzad, A.; Ahn, E.M.; Lee, Y.M.; Lee, Y.S. Decursinol angelate inhibits lps-induced macrophage polarization through modulation of the nfkappab and mapk signaling pathways. Molecules 2018, 23, 1880. [Google Scholar] [CrossRef]
- Choi, Y.E.; Ahn, H.; Ryu, J.H. Polyacetylenes from angelica gigas and their inhibitory activity on nitric oxide synthesis in activated macrophages. Biol. Pharm. Bull. 2000, 23, 884–886. [Google Scholar] [CrossRef]
- Kim, S.; Oh, H.-K.; Kim, J.-Y.; Hong, J.-W.; Cho, S.-I. A review of pharmacological effects of angelica gigas, angelica sinensis, angelica acutiloba and their bioactive compounds. J. Korean Med. 2011, 32, 1–24. [Google Scholar]
- Park, S.J.; Cha, H.S.; Lee, Y.H.; Kim, W.J.; Kim, D.H.; Kim, E.C.; Lee, K.H.; Kim, T.J. Effect of nodakenin on atopic dermatitis-like skin lesions. Biosci. Biotechnol. Biochem. 2014, 78, 1568–1571. [Google Scholar] [CrossRef]
- Hoffmann, R. The potential role of cytokines and t cells in alopecia areata. J. Investig. Dermatol. Symp. Proc. 1999, 4, 235–238. [Google Scholar] [CrossRef]
- Kim, S.N.; Akindehin, S.; Kwon, H.J.; Son, Y.H.; Saha, A.; Jung, Y.S.; Seong, J.K.; Lim, K.M.; Sung, J.H.; Maddipati, K.R.; et al. Anti-inflammatory role of 15-lipoxygenase contributes to the maintenance of skin integrity in mice. Sci. Rep. 2018, 8, 8856. [Google Scholar] [CrossRef] [PubMed]
- Weedon, D. Weedon’s Skin Pathology e-Book: Expert Consult-Online and Print; Elsevier Health Sciences: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Hwang, J.H.; Chu, H.; Ahn, Y.; Kim, J.; Kim, D.Y. Hmgb1 promotes hair growth via the modulation of prostaglandin metabolism. Sci. Rep. 2019, 9, 6660. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds AGN root extract is available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Components | |
|---|---|---|
| Coumarin Derivatives | Decursin | Major components |
| Decursinol | ||
| Decursinol angelate | ||
| Nodakenin, 4′′-Hydroxytigloyldecursinol, 4′′-hydroxydecursin, (2′′S,3′′S)-epoxyangeloyldecursinol, (2′′R,3′′R)-epoxyangeloyldecursinol, marmesinin, columbianetin-O-β-D-glucopyranoside, etc. | Other components | |
| Saccharides | Angelan (peptic polysaccharide) | Polysaccharide |
| Arabinose, Galactose, Galacturonic acid (sugar acid), etc. | Monosaccharides | |
| Polyacetylene | Octadeca-1,9-dien-4,6-diyn-3,8,18-triol, 18-acetoxy-octadeca-1,9-dien-4,6-diyn-3,8-diol, etc. | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, T.-K.; Kim, B.; Kim, D.W.; Ahn, J.H.; Sim, H.; Lee, J.-C.; Yang, G.E.; Her, Y.; Park, J.H.; Kim, H.S.; et al. Effects of Decursin and Angelica gigas Nakai Root Extract on Hair Growth in Mouse Dorsal Skin via Regulating Inflammatory Cytokines. Molecules 2020, 25, 3697. https://doi.org/10.3390/molecules25163697
Lee T-K, Kim B, Kim DW, Ahn JH, Sim H, Lee J-C, Yang GE, Her Y, Park JH, Kim HS, et al. Effects of Decursin and Angelica gigas Nakai Root Extract on Hair Growth in Mouse Dorsal Skin via Regulating Inflammatory Cytokines. Molecules. 2020; 25(16):3697. https://doi.org/10.3390/molecules25163697
Chicago/Turabian StyleLee, Tae-Kyeong, Bora Kim, Dae Won Kim, Ji Hyeon Ahn, Hyejin Sim, Jae-Chul Lee, Go Eun Yang, Young Her, Joon Ha Park, Hyun Sook Kim, and et al. 2020. "Effects of Decursin and Angelica gigas Nakai Root Extract on Hair Growth in Mouse Dorsal Skin via Regulating Inflammatory Cytokines" Molecules 25, no. 16: 3697. https://doi.org/10.3390/molecules25163697
APA StyleLee, T.-K., Kim, B., Kim, D. W., Ahn, J. H., Sim, H., Lee, J.-C., Yang, G. E., Her, Y., Park, J. H., Kim, H. S., Sim, T. H., Lee, H. S., & Won, M.-H. (2020). Effects of Decursin and Angelica gigas Nakai Root Extract on Hair Growth in Mouse Dorsal Skin via Regulating Inflammatory Cytokines. Molecules, 25(16), 3697. https://doi.org/10.3390/molecules25163697