Circulating and Adipose Tissue Fatty Acid Composition in Black South African Women with Obesity: A Cross-Sectional Study

 , , ,
, , ,
Abstract
:1. Introduction
2. Methods
2.1. Study Participants and Ethical Considerations
2.2. Body Composition and Body Fat Distribution
2.3. Fasting Blood Samples and Frequently Sampled Intravenous Glucose Tolerance Test
2.4. Adipose Tissue Sample Collection
2.5. Determination of RBC-TPL and SAT Fatty Acid Composition
3. Statistical Analyses
4. Results
4.1. Participant Characteristics and Dietary Intake
4.2. Tissue-Specific Fatty Acid Composition
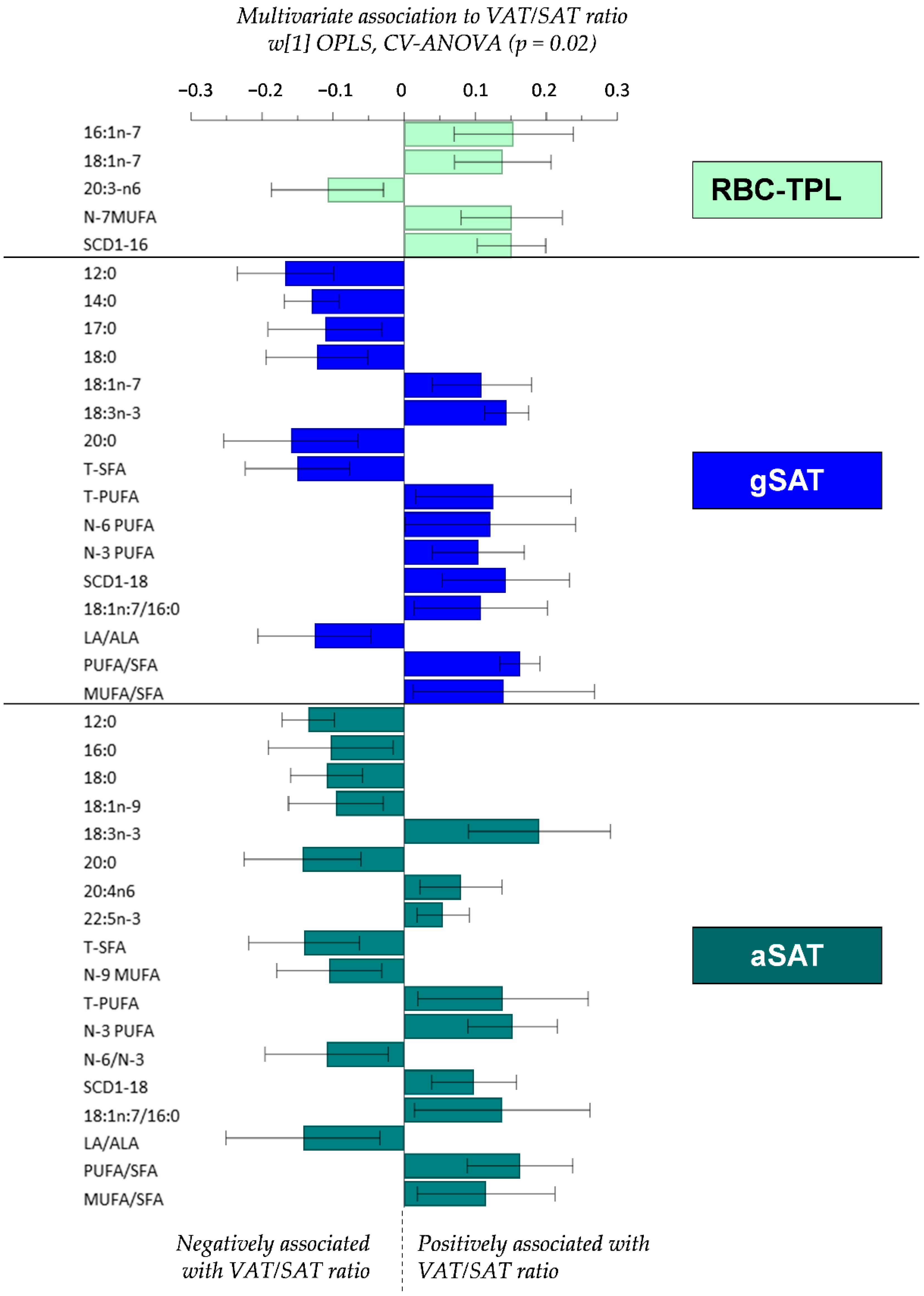
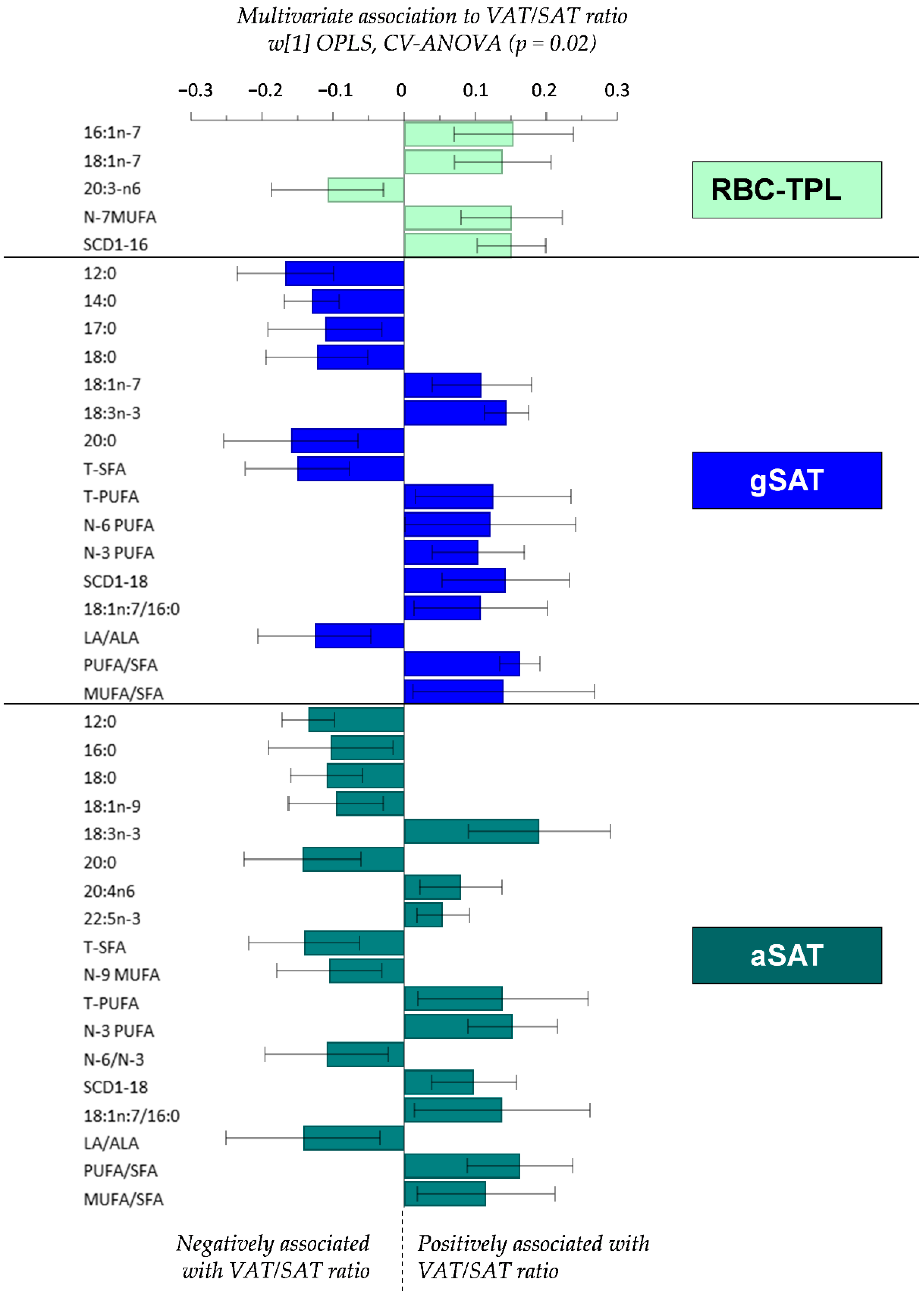
4.3. Relationship between Fatty Acid Profiles and the VAT/SAT Ratio
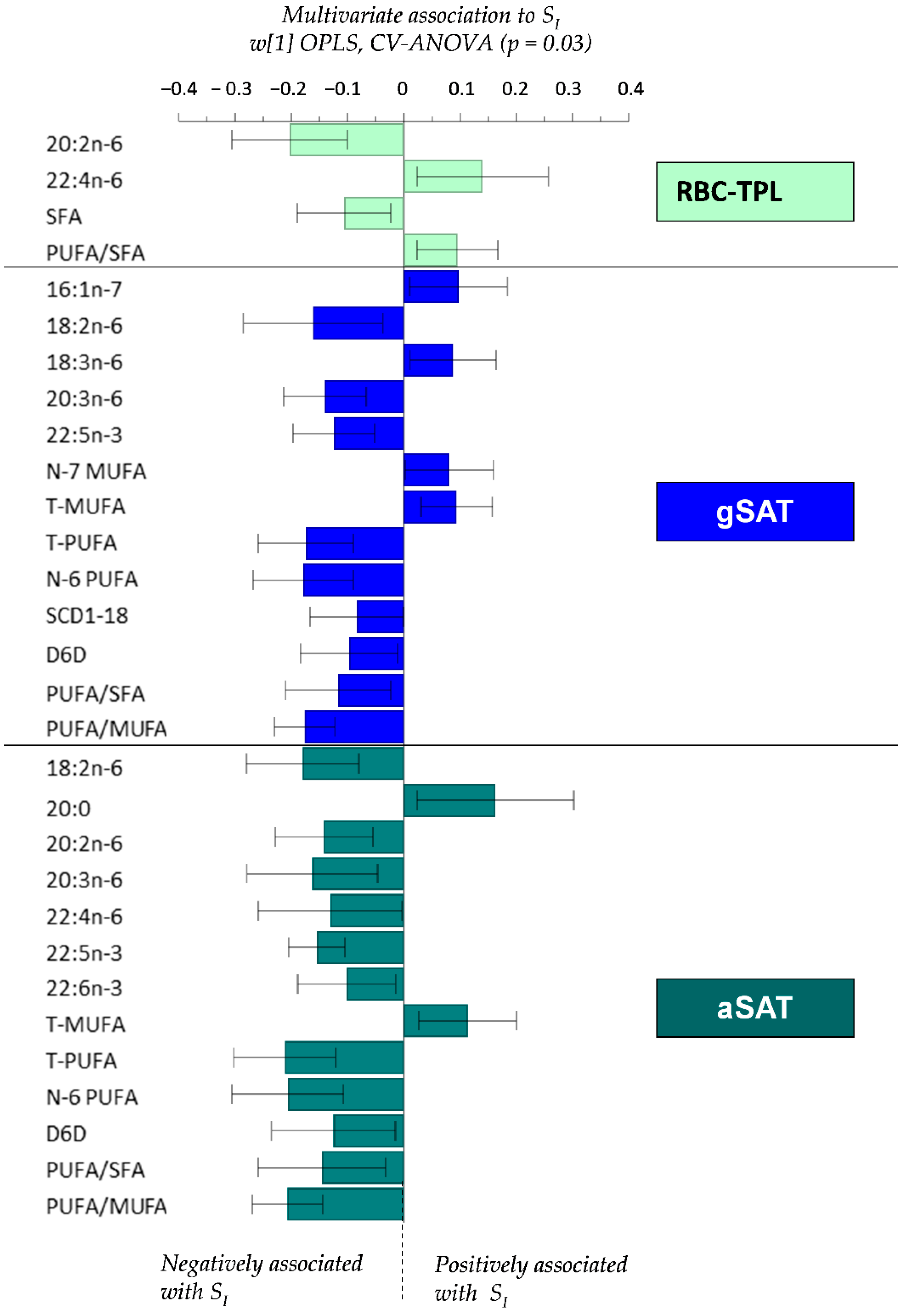
4.4. Relationship between Fatty Acid Profiles and SI
5. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Shay, C.M.; Carnethon, M.R.; Church, T.R.; Hankinson, A.L.; Chan, C.; Jacobs, D.R., Jr.; Lewis, C.E.; Schreiner, P.J.; Sternfeld, B.; Sidney, S. Lower extremity fat mass is associated with insulin resistance in overweight and obese individuals: The CARDIA study. Obesity 2011, 19, 2248–2253. [Google Scholar] [CrossRef] [PubMed]
- Preis, S.R.; Massaro, J.M.; Robins, S.J.; Hoffmann, U.; Vasan, R.S.; Irlbeck, T.; Meigs, J.B.; Sutherland, P.; D’Agostino, R.B., Sr.; O’donnell, C.J.; et al. Abdominal subcutaneous and visceral adipose tissue and insulin resistance in the Framingham heart study. Obesity 2010, 18, 2191–2198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goedecke, J.H.; Levitt, N.S.; Evans, J.; Ellman, N.; Hume, D.J.; Kotze, L.; Tootla, M.; Victor, H.; Keswell, D. The role of adipose tissue in insulin resistance in women of African ancestry. J. Obes. 2013, 2013, 952916. [Google Scholar] [CrossRef] [PubMed]
- Goedecke, J.H.; Levitt, N.S.; Utzschneider, K.M.; Faulenbach, M.V.; Dave, J.A.; West, S.; Victor, H.; Evans, J.; Olsson, T.; Walker, B.R.; et al. Differential effects of abdominal adipose tissue distribution on insulin sensitivity in black and white South African women. Obesity 2009, 17, 1506–1512. [Google Scholar] [CrossRef]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef] [Green Version]
- Keswell, D.; Tootla, M.; Goedecke, J.H. Associations between body fat distribution, insulin resistance and dyslipidaemia in black and white South African women. Cardiovasc. J. Afr. 2016, 27, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Joubert, J.; Norman, R.; Bradshaw, D.; Goedecke, J.H.; Steyn, N.P.; Puoane, T. Estimating the burden of disease attributable to excess body weight in South Africa in 2000. S. Afr. Med. J. 2007, 97, 683–690. [Google Scholar]
- Joffe, Y.T.; van der Merwe, L.; Evans, J.; Collins, M.; Lambert, E.V.; September, A.V.; Goedecke, J.H. Interleukin-6 Gene Polymorphisms, Dietary Fat Intake, Obesity and Serum Lipid Concentrations in Black and White South African Women. Nutrients 2014, 6, 2436–2465. [Google Scholar] [CrossRef] [Green Version]
- Goff, L.M.; Griffin, B.A.; Lovegrove, J.A.; Sanders, T.A.; Jebb, S.A.; Bluck, L.J.; Frost, G.S. Ethnic differences in beta-cell function, dietary intake and expression of the metabolic syndrome among UK adults of South Asian, black African-Caribbean and white-European origin at high risk of metabolic syndrome. Diabetes Vasc. Dis. Res. 2013, 10, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Mathias, R.A.; Sergeant, S.; Ruczinski, I.; Torgerson, D.G.; Hugenschmidt, C.E. The impact of FADS genetic variants on ω6 polyunsaturated fatty acid metabolism in African Americans. BMC Genet. 2011, 12, 50. [Google Scholar] [CrossRef] [Green Version]
- Sergeant, S.; Hugenschmidt, C.E.; Rudock, M.E.; Ziegler, J.T.; Ivester, P.; Ainsworth, H.C.; Vaidya, D.; Case, D.; Langefeld, C.D.; Freedman, B.I.; et al. Differences in arachidonic acid levels and fatty acid desaturase (FADS) gene variants in African Americans and European Americans with diabetes or the metabolic syndrome. Br. J. Nutr. 2011, 107, 547–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodson, L.; Skeaff, C.M.; Fielding, B.A. Fatty acid composition of adipose tissue and blood in humans and its use as a biomarker of dietary intake. Prog. Lipid Res. 2008, 47, 348–380. [Google Scholar] [CrossRef] [PubMed]
- Markussen, M.S.; Veierod, M.B.; Ursin, G.; Andersen, L.F. The effect of under-reporting of energy intake on dietary patterns and on the associations between dietary patterns and self-reported chronic disease in women aged 50–69 years. Br. J. Nutr. 2016, 116, 547–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunes, O.; Tascilar, E.; Sertoglu, E.; Tas, A.; Serdar, M.A.; Kaya, G.; Kayadibi, H.; Ozcan, O. Associations between erythrocyte membrane fatty acid compositions and insulin resistance in obese adolescents. Chem. Phys. Lipids 2014, 184, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Rise, P.; Eligini, S.; Ghezzi, S.; Colli, S.; Galli, C. Fatty acid composition of plasma, blood cells and whole blood: Relevance for the assessment of the fatty acid status in humans. Prostaglandins Leukot Essent Fatty Acids 2007, 76, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Perona, J.S. Membrane lipid alterations in the metabolic syndrome and the role of dietary oils. Biochim. Biophys. Acta 2017, 1859, 1690–1703. [Google Scholar] [CrossRef]
- Warensjo, E.; Rosell, M.; Hellenius, M.L.; Vessby, B.; de Faire, U.; Riserus, U. Associations between estimated fatty acid desaturase activities in serum lipids and adipose tissue in humans: Links to obesity and insulin resistance. Lipids Health Dis. 2009, 8, 37. [Google Scholar] [CrossRef] [Green Version]
- Mahendran, Y.; Agren, J.; Uusitupa, M.; Cederberg, H.; Vangipurapu, J.; Stancakova, A.; Schwab, U.; Kuusisto, J.; Laakso, M. Association of erythrocyte membrane fatty acids with changes in glycemia and risk of type 2 diabetes. Am. J. Clin. Nutr. 2014, 99, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Sjogren, P.; Sierra-Johnson, J.; Gertow, K.; Rosell, M.; Vessby, B.; de Faire, U.; Hamsten, A.; Hellenius, M.L.; Fisher, R.M. Fatty acid desaturases in human adipose tissue: Relationships between gene expression, desaturation indexes and insulin resistance. Diabetologia 2008, 51, 328–335. [Google Scholar] [CrossRef] [Green Version]
- Roberts, R.; Hodson, L.; Dennis, A.L.; Neville, M.J.; Humphreys, S.M.; Harnden, K.E.; Micklem, K.J.; Frayn, K.N. Markers of de novo lipogenesis in adipose tissue: Associations with small adipocytes and insulin sensitivity in humans. Diabetologia 2009, 52, 882–890. [Google Scholar] [CrossRef] [Green Version]
- Blüher, M. Adipose Tissue Dysfunction Contributes to Obesity Related Metabolic Diseases. Best Pract. Res. Clin. Endocrinol. Metab. 2013, 27, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Roden, M.; Price, T.B.; Perseghin, G.; Petersen, K.F.; Rothman, D.L.; Cline, G.W.; Shulman, G.I. Mechanism of free fatty acid induced insulin resistance in humans. J. Clin. Investig. 1996, 97, 2859–2865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherer, P.E. Adipose tissue: From lipid storage compartment to endocrine organ. Diabetes 2006, 55, 1537–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karastergiou, K.; Fried, S.K.; Xie, H.; Lee, M.J.; Divoux, A.; Rosencrantz, M.A.; Chang, R.J.; Smith, S.R. Distinct developmental signatures of human abdominal and gluteal subcutaneous adipose tissue depots. J. Clin. Endocrinol. Metab. 2013, 98, 362–371. [Google Scholar] [CrossRef]
- Karpe, F.; Pinnick, K.E. Biology of upper-body and lower-body adipose tissue--link to whole-body phenotypes. Nat. Rev. Endocrinol. 2015, 11, 90–100. [Google Scholar] [CrossRef]
- Karastergiou, K.; Smith, S.R.; Greenberg, A.S.; Fried, S.K. Sex differences in human adipose tissues—The biology of pear shape. Biol. Sex Differ. 2012, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Nikolaidis, M.G.; Mougios, V. Effects of Exercise on the Fatty-Acid Composition of Blood and Tissue Lipids. Sports Med. Open 2004, 34, 1051–1076. [Google Scholar] [CrossRef]
- Goedecke, J.H.; Mendham, A.E.; Clamp, L.; Nono Nankam, P.A.; Fortuin-de Smidt, M.C.; Phiri, L.; Micklesfield, L.K.; Keswell, D.; Woudberg, N.J.; Lecour, S.; et al. An Exercise Intervention to Unravel the Mechanisms Underlying Insulin Resistance in a Cohort of Black South African Women: Protocol for a Randomized Controlled Trial and Baseline Characteristics of Participants. JMIR Res. Protoc. 2018, 7, e75. [Google Scholar] [CrossRef] [Green Version]
- Clamp, L.D.; Mendham, A.E.; Kroff, J.; Goedecke, J.H. Higher baseline fat oxidation promotes gynoid fat mobilization in response to a 12 week exercise intervention in sedentary, obese black South African women. Appl. Physiol. Nutr. Metab. 2019. [Google Scholar] [CrossRef]
- Goedecke, J.H.; Micklesfield, L.K.; Levitt, N.S.; Lambert, E.V.; West, S.; Maartens, G.; Dave, J.A. Effect of different antiretroviral drug regimens on body fat distribution of HIV-infected South African women. AIDS Res. Hum. Retrovir. 2013, 29, 557–563. [Google Scholar] [CrossRef]
- Micklesfield, L.K.; Goedecke, J.H.; Punyanitya, M.; Wilson, K.E.; Kelly, T.L. Dual-energy X-ray performs as well as clinical computed tomography for the measurement of visceral fat. Obesity 2012, 20, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Fortuin-de Smidt, M.C.; Mendham, A.E.; Hauksson, J.; Hakim, O.; Stefanovski, D.; Clamp, L.; Phiri, L.; Swart, J.; Goff, L.M.; Micklesfield, L.K.; et al. Effect of exercise training on insulin sensitivity, hyperinsulinemia and ectopic fat in black South African women: A randomized controlled trial. Eur. J. Endocrinol. 2020. [Google Scholar] [CrossRef]
- Bergman, R.; Ider, Y.; Bowden, C.; Cobelli, C. Quantitative estimation of insulin sensitivity. Am. J. Physiol. 1979, 236, E667–E677. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Chimhashu, T.; Malan, L.; Baumgartner, J.; van Jaarsveld, P.J.; Galetti, V.; Moretti, D.; Smuts, C.M.; Zimmermann, M.B. Sensitivity of fatty acid desaturation and elongation to plasma zinc concentration: A randomised controlled trial in Beninese children. Br. J. Nutr. 2018, 119, 610–619. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.Y.; Chapkin, R.S. Importance of dietary γ-linolenic acid in human health and nutrition. J. Nutr. 1998, 128, 1411–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jump, D.B. Fatty acid regulation of hepatic lipid metabolism. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Kroger, J.; Zietemann, V.; Enzenbach, C.; Weikert, C.; Jansen, E.H.J.M.; Döring, F.; Joost, H.; Boeing, H.; Schulze, M.B. Erythrocyte membrane phospholipid fatty acids, desaturase activity, and dietary fatty acids in relation to risk of type 2 diabetes in the European prospective investigation into cancer and nutrition (EPIC)-potsdam study. Am. J. Clin. Nutr. 2011, 93, 127–142. [Google Scholar] [CrossRef] [Green Version]
- Martinelli, N.G.D.; Malerba, G.; Guarini, P.; Illig, T.; Trabetti, E.; Sandri, M.; Friso, S.; Pizzolo, F.; Schaeffer, L.; Heinrich, J.; et al. FADS genotypes and desaturase activity estimated by the ratio of arachidonic acid to linoleic acid are associated with inflammation and coronary artery disease. Am. J. Clin. Nutr. 2008, 88, 941–949. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, L.; Trygg, J.; Wold, S. CV-ANOVA for significance testing of PLS and OPLS® models. J. Chemom. 2008, 22, 594–600. [Google Scholar] [CrossRef]
- Efron, B.; Gong, G. A Leisurely Look at the Bootstrap, the Jackknife, and Cross-Validation. Am. Stat. 1983, 37, 36–48. [Google Scholar]
- Abraham, T.M.; Pedley, A.; Massaro, J.M.; Hoffmann, U.; Fox, C.S. Association between visceral and subcutaneous adipose depots and incident cardiovascular disease risk factors. Circulation 2015, 132, 1639–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacIntyre, U.; Kruger, H.; Venter, C.; Vorster, H. Dietary intakes of an African population in different stages of transition in the North West Province, South Africa: The THUSA study. Nutr. Res. 2002, 22, 239–256. [Google Scholar] [CrossRef]
- Garaulet, M.; Pérez-Llamas, F.; Pérez-Ayala, M.; Martínez, P.; Sánchez de Medina, F.; Tebar, F.J.; Zamora, S. Site-specific differences in the fatty acid composition of abdominal adipose tissue in an obese population from a Mediterranean area: Relation with dietary fatty acids, plasma lipid profile, serum insulin, and central obesity. Am. J. Clin. Nutr. 2001, 74, 585–591. [Google Scholar] [CrossRef] [Green Version]
- Gower, B.A.; Alvarez, J.A.; Bush, N.C.; Hunter, G.R. Insulin sensitivity affects propensity to obesity in an ethnic-specific manner: Results from two controlled weight loss intervention studies. Nutr. Metab. 2013, 10, 3. [Google Scholar] [CrossRef] [Green Version]
- Gianmattia, G.; Ferreri, C.; Marfella, R.; Pournaras, D.; Roux, C.; del Genio, F.; Paolo, L.; Tolone, S.; Docimo, L.; Puca, A.A. Morbid Obesity is Associated to Altered Fatty Acid Profile of Erythrocyte Membranes. J. Diabetes Metab. 2015, 6, 1–4. [Google Scholar]
- Cordero, A.; González, J.E.; Perona, J.S.; Padilla, L.; Ferre, A.; Hita, O.; Baeza, R.; Barrilao, G.; Rivas, G. Obesity and its relation with markers of inflammation and erythrocyte fatty acids in a group of overweight adolescents. Nutr. Hosp. 2012, 27, 161–164. [Google Scholar]
- Mika, A.; Sledzinski, T. Alterations of specific lipid groups in serum of obese humans: A review. Obes. Rev. 2017, 18, 247–272. [Google Scholar] [CrossRef]
- Mika, A.; Kaska, L.; Korczynska, J.; Mirowska, A.; Stepnowski, P.; Proczko, M.; Ratnicki-Sklucki, K.; Goyke, E.; Sledzinski, T. Visceral and subcutaneous adipose tissue stearoyl-CoA desaturase-1 mRNA levels and fatty acid desaturation index positively correlate with BMI in morbidly obese women. Eur. J. Lipid Sci. Technol. 2015, 117, 926–932. [Google Scholar] [CrossRef]
- Ojwang, A.A.; Kruger, H.S.; Zec, M.; Ricci, C.; Pieters, M.; Kruger, I.M.; Wentzel-Viljoen, E.; Smuts, C.M. Plasma phospholipid fatty acid patterns are associated with adiposity and the metabolic syndrome in black South Africans: A cross-sectional study. Cardiovasc. J. Afr. 2019, 30, 228–238. [Google Scholar] [CrossRef]
- Malcom, G.T.; Bhattacharyya, A.K.; Velez-Duran, M.; Guzman, M.A.; Oalmann, M.C.; Strong, J.P. Fatty acid composition of adipose tissue in humans: Differences between subcutaneous sites. Am. J. Clin. Nutr. 1989, 50, 288–291. [Google Scholar] [CrossRef] [PubMed]
- Phinney, S.D.; Stern, J.S.; Burke, K.E.; Tang, A.B.; Miller, G.; Holman, R.T. Human subcutaneous adipose tissue shows site-specific differences in fatty acid composition. Am. J. Clin. Nutr. 1994, 60, 725–729. [Google Scholar] [CrossRef]
- Calder, P.C.; Harvey, D.J.; Pond, C.M.; Newsholme, E.A. Site-specific differences in the fatty acid composition of human adipose tissue. Lipids 1992, 27, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Manson, J.E.; Rautiainen, S.; Gaziano, J.M.; Buring, J.E.; Tsai, M.Y.; Sesso, H.D. A prospective study of erythrocyte polyunsaturated fatty acid, weight gain, and risk of becoming overweight or obese in middle-aged and older women. Eur. J. Nutr. 2016, 55, 687–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amri, E.Z.; Ailhaud, G.; Grimaldi, P.A. Fatty acids as signal transducing molecules: Involvement in the differentiation of preadipose to adipose cells. J. Lipid Res. 1994, 35, 930–937. [Google Scholar] [PubMed]
- Aoun, M.; Michel, F.; Fouret, G.; Schlernitzauer, A.; Ollendorff, V.; Wrutniak-Cabello, C.; Cristol, J.P.; Carbonneau, M.A.; Coudray, C.; Feillet-Coudray, C. A grape polyphenol extract modulates muscle membrane fatty acid composition and lipid metabolism in high-fat--high-sucrose diet-fed rats. Br. J. Nutr. 2011, 106, 491–501. [Google Scholar] [CrossRef] [Green Version]
- Micallef, M.; Munro, I.; Phang, M.; Garg, M. Plasma n-3 Polyunsaturated Fatty Acids are negatively associated with obesity. Br. J. Nutr. 2009, 102, 1370–1374. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Bhulaidok, S.; Cai, Z.; Xu, T.; Xu, F.; Wahlqvist, M.L.; Li, D. Plasma phospholipids n-3 polyunsaturated fatty acid is associated with metabolic syndrome. Mol. Nutr. Food Res. 2010, 54, 1628–1635. [Google Scholar] [CrossRef]
- Fickova, M.; Hubert, P.; Crémel, G.; Leray, C. Dietary (n-3) and (n-6) polyunsaturated fatty acids rapidly modify fatty acid composition and insulin effects in rat adipocytes. J. Nutr. 1998, 128, 512–519. [Google Scholar] [CrossRef]
- Aglago, E.K.; Biessy, C.; Torres-Mejia, G.; Angeles-Llerenas, A.; Gunter, M.J.; Romieu, I.; Chajes, V. Association between serum phospholipid fatty acid levels and adiposity in Mexican women. J. Lipid Res. 2017, 58, 1462–1470. [Google Scholar] [CrossRef] [Green Version]
- Mayneris-Perxachs, J.; Guerendiain, M.; Castellote, A.I.; Estruch, R.; Covas, M.I.; Fito, M.; Salas-Salvado, J.; Martinez-Gonzalez, M.A.; Aros, F.; Lamuela-Raventos, R.M. Plasma fatty acid composition, estimated desaturase activities, and their relation with the metabolic syndrome in a population at high risk of cardiovascular disease. Clin. Nutr. 2014, 33, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Kabagambe, E.K.; Tsai, M.Y.; Hopkins, P.N.; Ordovas, J.M.; Peacock, J.M.; Borecki, I.B.; Arnett, D.K. Erythrocyte fatty acid composition and the metabolic syndrome: A National Heart, Lung, and Blood Institute GOLDN study. Clin. Chem. 2008, 54, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Yary, T.; Voutilainen, S.; Tuomainen, T.P.; Ruusunen, A.; Nurmi, T.; Virtanen, J.K. Omega-6 polyunsaturated fatty acids, serum zinc, delta-5- and delta-6-desaturase activities and incident metabolic syndrome. J. Hum. Nutr. Diet. 2017, 30, 506–514. [Google Scholar] [CrossRef]
- Rosqvist, F.; Bjermo, H.; Kullberg, J.; Johansson, L.; Michaelsson, K.; Ahlstrom, H.; Lind, L.; Riserus, U. Fatty acid composition in serum cholesterol esters and phospholipids is linked to visceral and subcutaneous adipose tissue content in elderly individuals: A cross-sectional study. Lipids Health Dis. 2017, 16, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gertow, K.; Rosell, M.; Sjogren, P.; Eriksson, P.; Vessby, B.; de Faire, U.; Hamsten, A.; Hellenius, M.L.; Fisher, R.M. Fatty acid handling protein expression in adipose tissue, fatty acid composition of adipose tissue and serum, and markers of insulin resistance. Eur. J. Clin. Nutr. 2006, 60, 1406–1413. [Google Scholar] [CrossRef]
- Iggman, D.; Arnlov, J.; Vessby, B.; Cederholm, T.; Sjogren, P.; Riserus, U. Adipose tissue fatty acids and insulin sensitivity in elderly men. Diabetologia 2010, 53, 850–857. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
| Variables | Values (n = 41) |
|---|---|
| Age (year) | 23 (21–27) |
| Body composition | |
| BMI (kg/m2) | 33.9 ± 2.8 |
| Waist circumference (cm) | 103.8 ± 8.0 |
| Hip circumference (cm) | 116 (114–121) |
| WHR | 0.9 (0.8–0.9) |
| Body FM (%) | 50.4 ± 12.0 |
| Android FM (%) | 8.1 ± 1.1 |
| Gynoid FM (%) | 18.6 ± 2.0 |
| VAT (cm3) | 900 ± 345 |
| SAT (cm3) | 5454 ± 1547 |
| VAT/SAT | 0.2 ± 0.2 |
| Insulin sensitivity | |
| Fasting glucose (mmol/L) | 2777.6 (2027.6–4287.5) |
| Fasting Insulin (µIU/mL) | 13.1 (8. –16.7) |
| SI (mU/L)−1min−1 | 2.5 ± 1.4 |
| Variables | Median (25th–75th Percentile) |
|---|---|
| Energy (kJ) | 12507 (9330–15374) |
| Total protein (g) | 96.1 (67.5–119.9) |
| Protein (%E) | 13.2 (12.2–14.1) |
| Total CHO (g) | 372.7 (295.1–464.7) |
| CHO (%E) | 53.1 (49.0–54.8) |
| Total Fat (g) | 113.0 (71.5–131.4) |
| Fat (% E) | 33.4 (30.1–36.5) |
| SFA (g) | 31.4 (18.7–42.3) |
| SFA (%E) | 9.5 (7.9–11.6) |
| MUFA (g) | 36.0 (24.1–44.6) |
| MUFA (%E) | 11.4 (10.1–12.5) |
| PUFA (g) | 25.8 (18.0–32.5) |
| PUFA (%E) | 7.9 (7.2–9.8) |
| RBC-TPL | gSAT | aSAT | p Values | |
|---|---|---|---|---|
| Saturated fatty acids (SFAs) | ||||
| 14:0 (Myristic acid) | 0.24 ± 0.06 a,b | 2.77 ± 0.62 | 2.89 ± 0.59 | <0.001 |
| 16:0 (Palmitic acid) | 21.88 ± 2.06 | 21.70 ± 1.12 | 22.52 ± 1.11 c | 0.024 |
| 18:0 (Stearic acid) | 16.31 ± 0.78 a,b | 5.25 ± 1.53 | 5.75 ± 1.49 c | <0.001 |
| 20:0 (Arachidic acid) | 0.35 ± 0.04 a,b | 0.16 ± 0.06 | 0.16 ± 0.05 | <0.001 |
| 22:0 (Behenic acid) | 1.39 ± 0.20 a,b | 0.06 ±0.03 | 0.05 ± 0.01 | <0.001 |
| 24:0 (Lignoceric acid) | 3.99 ± 0.64 a,b | 0.05 ± 0.02 | 0.04 ± 0.01 | <0.001 |
| Total SFAs | 44.16 ± 1.68 a,b | 30.76 ± 2.88 | 32.23 ± 2.46 c | <0.001 |
| Mono-unsaturated fatty acids (MUFAs) | ||||
| 16:1n-7 (Palmitoleic acid) | 0.24 ± 0.07 a,b | 6.61 ± 2.09 c | 5.93 ± 1.68 | <0.001 |
| 18:1n-7 (cis-Vaccenic acid) | 1.11 ± 0.22 a,b | 3.00 ± 0.49 c | 2.79 ± 0.43 | <0.001 |
| 18:1n-9 (Oleic acid) | 10.70 ± 0.87 a,b | 32.56 ± 0.92 | 32.49 ± 1.01 | <0.001 |
| 20:1n-9 (Eicosenoic acid) | 0.18 ± 0.02 a,b | 0.71 ± 0.10 | 0.69 ± 0.11 | <0.001 |
| Total n-7 MUFAs | 1.34 ± 0.27 a,b | 9.61 ± 2.50 c | 8.71 ± 2.02 | <0.001 |
| Total n-9 MUFAs | 14.23 ± 0.89 a,b | 33.31 ± 0.94 | 33.22 ± 1.04 | <0.001 |
| Total MUFAs | 15.57 ± 0.97 a,b | 42.92 ± 2.64 c | 41.93 ± 2.25 | <0.001 |
| Poly-unsaturated fatty acids (PUFAs) | ||||
| 20:5n-3 (EPA) | 0.47 ± 0.23 a,b | 0.09 ± 0.04 | 0.08 ± 0.04 | <0.001 |
| 22:5n-3 (DPA n-3) | 1.96 ± 0.31 a,b | 0.27 ± 0.09 | 0.24 ± 0.08< | <0.001 |
| 22:6n-3 (DHA) | 4.45 ± 0.81 a,b | 0.24 ± 0.08 | 0.21 ± 0.08< | <0.001 |
| 18:2n-6 (LA) | 11.71 ± 1.31 a,b | 22.33 ± 1.64 | 22.15 ± 1.55 | <0.001 |
| 20:2n-6 (Eicosadienoic acid) | 0.35 ± 0.04 a,b | 0.50 ± 0.10 | 0.48 ± 0.12 | <0.001 |
| 20:3n-6 (DGLA) | 1.53 ± 0.22 a,b | 0.43 ±0.12 c | 0.37 ± 0.12 | <0.001 |
| 20:4n-6 (AA) | 15.74 ± 1.19 a,b | 0.73 ±0.18 | 0.67 ± 0.17 | <0.001 |
| 22:4n-6 (Adrenic acid) | 3.37 ± 0.59 a,b | 0.30 ± 0.09 | 0.27± 0.09 | <0.001 |
| 22:5n-6 (DPA-n-6) | 0.68 ± 0.16 a,b | 0.08 ± 0.04 | 0.08 ± 0.03 | <0.001 |
| Total n-3 | 6.90 ± 1.87 a,b | 1.76 ± 0.33 | 1.66 ± 0.32 | <0.001 |
| Total n-6 | 33.38 ± 1.87 a,b | 24.56 ± 1.73 | 24.18 ± 1.64 | <0.001 |
| Total PUFAs | 40.27 ± 1.77 a,b | 26.32 ± 1.95 | 25.84 ± 1.80 | <0.001 |
| Estimated enzyme activities/indices | ||||
| D5D | 10.57 ± 1.95 a,b | 1.79 ± 0.43 | 1.90 ± 0.37 | <0.001 |
| D6D | 0.13 ± 0.02 a,b | 0.019 ± 0.01 | 0.016 ± 0.01 | <0.001 |
| SCD1-16 | 0.01 ± 0.00 a,b | 0.31 ± 0.11 c | 0.27 ± 0.08 | <0.001 |
| SCD1-18 | 0.68 ± 0.01 a,b | 0.65 ± 0.30 c | 0.53 ± 0.21 | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nono Nankam, P.A.; van Jaarsveld, P.J.; Chorell, E.; Fortuin-de Smidt, M.C.; Adams, K.; Blüher, M.; Olsson, T.; Mendham, A.E.; Goedecke, J.H. Circulating and Adipose Tissue Fatty Acid Composition in Black South African Women with Obesity: A Cross-Sectional Study. Nutrients 2020, 12, 1619. https://doi.org/10.3390/nu12061619
Nono Nankam PA, van Jaarsveld PJ, Chorell E, Fortuin-de Smidt MC, Adams K, Blüher M, Olsson T, Mendham AE, Goedecke JH. Circulating and Adipose Tissue Fatty Acid Composition in Black South African Women with Obesity: A Cross-Sectional Study. Nutrients. 2020; 12(6):1619. https://doi.org/10.3390/nu12061619
Chicago/Turabian StyleNono Nankam, Pamela A., Paul J. van Jaarsveld, Elin Chorell, Melony C. Fortuin-de Smidt, Kevin Adams, Matthias Blüher, Tommy Olsson, Amy E. Mendham, and Julia H. Goedecke. 2020. "Circulating and Adipose Tissue Fatty Acid Composition in Black South African Women with Obesity: A Cross-Sectional Study" Nutrients 12, no. 6: 1619. https://doi.org/10.3390/nu12061619