Abstract
The naturally colorful fur of the Rex rabbit is becoming increasingly popular in the modern textile market. Our previous study found that POU class 2 homeobox 1 gene (POU2F1) potentially affects the expression of genes involved in fur color formation in the Rex rabbit, but the function and regulation of POU2F1 has not been reported. In this study, the expression patterns of POU2F1 in Rex rabbits of various colors, as well as in different organs, were analyzed by RT-qPCR. Interference and overexpression of POU2F1 were used to identify the potential effects of POU2F1 on other genes related to fur color formation. The results show that the levels of POU2F1 expression were significantly higher in the dorsal skin of the brown and protein yellow Rex rabbits, compared with that of the black one. POU2F1 mRNAs were widespread in the tissues examined in this study and showed the highest level in the lungs. By transfecting rabbit melanocytes with an POU2F1-overexpression plasmid, we found that the POU2F1 protein was located at the nucleus, and the protein showed the classic characteristics of a transcription factor. In addition, abnormal expression of POU2F1 significantly affected the expression of pigmentation-related genes, including SLC7A11, MITF, SLC24A5, MC1R, and ASIP, revealing the regulatory roles of POU2F1 on pigmentation. The results provide the basis for further exploration of the role of POU2F1 in fur color formation of the Rex rabbit.
1. Introduction
Rex rabbit fur is becoming a fashionable raw material for modern textiles and has outstanding features, including aesthetic appeal, smoothness, softness, lightness, flexibility, and heat-retaining properties. The multiple natural colors of the Rex rabbit have made it increasingly popular. In particular, some special colors, such as chinchilla, brown, protein yellow, and protein chinchilla, have the advantages of being not only attractive but also nontoxic and ecological, as no artificial dyes are required [1]. The huge demand in the market for fashionable Rex rabbit fur has promoted research into the characteristics of its fur color.
The formation of fur color in mammals is a complex process influenced by diverse factors, including the environment, management, and the genetic background [2,3]. Briefly, the relative quantity and distribution of melanocyte products, such as eumelanin and pheomelanin, determine the fur color [4,5]. It has been reported that gene expression affects the pigmentation of rabbit fur through various mechanisms; for example, some genes, such as SLC7A11 [6,7,8], melanocortin receptor 1 (MC1R) [9,10,11], microphthalmia-associated transcription factor (MITF) [12], and ASIP [13], are likely to influence the production and deposition of melanin.
Recently, SLC7A11 was found to be modulated by POU class 2 homeobox 1 gene (POU2F1) in rabbit pigmentation [14]. POU2F1, also known as OCT1, OTF1, or oct-1B [15], is a potent regulator of stress responses, metabolism, and tumorigenicity and is itself regulated by phosphorylation, ubiquitination, O-GlcNAcylation, and other mechanisms [16]. Recent studies into POU2F1 have focused on its impacts on cancers and tumors [17,18,19,20], especially hepatocellular carcinoma [21,22]. Furthermore, POU2F1 was identified as a transcription factor that binds to the promoter region of SLC7A11 via two binding sites in the Rex rabbit [14]. However, little is known about the role of POU2F1 in fur color formation in mammals, and the effects of the gene on the fur color of the Rex rabbit remain unclear.
Therefore, this study is the next stage of our research to analyze the potential impacts of POU2F1 on the significant genes involved in the formation of Rex rabbit fur color. The expression pattern of POU2F1 in the dorsal skin of the Rex rabbit with different fur colors, and in different organs, was analyzed by RT-qPCR. Additionally, a pcDNA3.1(+)-Myc-POU2F1 vector and siRNAs were constructed to analyze the potential regulatory roles of POU2F1 on SLC7A11, as well as some other pigmentation-related genes, such as SLC24A5, MITF, MC1R, CREB1, and ASIP, to reveal the regulatory role of POU2F1 in fur color formation of the Rex rabbit.
2. Materials and Methods
2.1. Animals and Samples
Eighteen 6-month-old Rex rabbits with 6 different fur colors (n = 3 for each color), including black (BL), chinchilla (CH), white (WH), brown (BR), protein yellow (PY), and protein chinchilla (PC), were provided by a rabbit breeding farm in Yuyao, Zhejiang, China. A 1 cm2 sample of dorsal skin tissue was collected from Rex rabbits of each color (n = 3) after anesthesia by injection of sodium pentobarbital solution (0.7%) into the ear vein. Tissue samples of different organs (heart, liver, spleen, lung, kidney, jejunum, colon, ileum, cecum, rectum, dorsal skin, sacculus rotundus, and gizzard) were collected from other white and black Rex rabbits (n = 3, respectively). White is the most common color of Rex rabbit and is widely used for fur production around the world because it is easily dyed and has good plasticity, while black rabbits were chosen for their striking contrast. These rabbits were euthanized by ear vein injection of 25 mL air after deep anesthesia, and organ samples were collected about 5 min after confirmation of the absence of heartbeat and death. All tissue samples were placed in liquid nitrogen immediately after being cut into small pieces and stored at −80 °C until use. The experimental procedures were approved by the Animal Care and Use Committee of Yangzhou University (Yangzhou, China, 24 October 2017, No. 201710001).
2.2. Melanocyte Culture and RNA Extraction
Melanocytes were separated by two-step enzymatic digestion from a 1.5 × 1.5 cm2 section of dorsal skin from white Rex rabbits according to our previous report [23]. Total RNA of the dorsal skin and organs was extracted using RNAsimple Total RNA kit (Tiangen, Beijing, China), and total RNA from melanocytes was extracted using Trizol reagent (Invitrogen, Carlsbad, CA, USA), according to the manufacturer’s instructions. Electrophoresis with 1% agarose gel was used to monitor RNA degradation and contamination. RNA purity and concentration were measured using a NanoPhotometer spectrophotometer (Thermo Fisher Scientific, Wilmington, NC, USA).
2.3. Real-Time qPCR
The dorsal skin and organs were submitted for quantitative real-time PCR to detect the expression levels of POU2F1. Briefly, cDNA was synthesized from approximately 1 μg RNA using HiScript II Q Select RT SuperMix for qPCR (Vazyme, Nanjing, China). Then, the cDNA was used for RT-qPCR with the AceQ qPCR SYBRR Green Master Mix (Vazyme, Nanjing, China), based on the manufacturer’s instructions. To improve the data reliability, cDNA of each sample was tested three times for technical replication, and three biological replicates were tested for each group. In order to prevent deviation due to possible unstable expression of reference genes, glyceraldehyde 3-phosphate dehydrogenase (GAPDH), β-actin, and hypoxanthine phosphoribosyltransferase 1 (HPRT1) were used as reference genes [24]. The primer sequences for RT-qPCR are listed in Table 1. The output data were analyzed using QuantStudioR 5 (Applied Biosystems), and the expression levels were analyzed according to the 2−∆∆Ct method [25].

Table 1.
Primer sequences for RT-qPCR.
2.4. Construction of pcDNA3.1(+)-Myc-POU2F1 Vector
The PrimeScript™ 1st Strand cDNA Synthesis Kit (Takara, Beijing, China) was used to synthesize Rex rabbit skin cDNA of high quality for the construction of the POU2F1 overexpression vector. The CDS sequence of the POU2F1 gene was amplified by PCR using Phanta Max Super-Fidelity DNA Polymerase (Vazyme, Nanjing, China), according to the mRNA sequence of rabbit POU2F1 (NC_013681.1). The amplified CDS sequence of POU2F1 was subcloned into an NheI- and EcoRI-digested pcDNA3.1(+)-Myc vector (Invitrogen) (Forward primer: gggagacccaagctggctagcATGGCGGACGGAGGAGCA; Reverse primer: gtagtcggatcctttgaattcCTGTGCCTTGGAGGCGGT), and the recombinant plasmid was named pcDNA3.1(+)-Myc-POU2F1 (Figure 1) for the subsequent steps. The pcDNA3.1(+)-Myc-POU2F1 was transferred into melanocytes to overexpress the POU2F1 gene, and the expression levels of the fur-color-related genes (SLC24A5, SLC7A11, MITF, MC1R, CREB1, and ASIP) were detected by RT-qPCR, according to the above method.
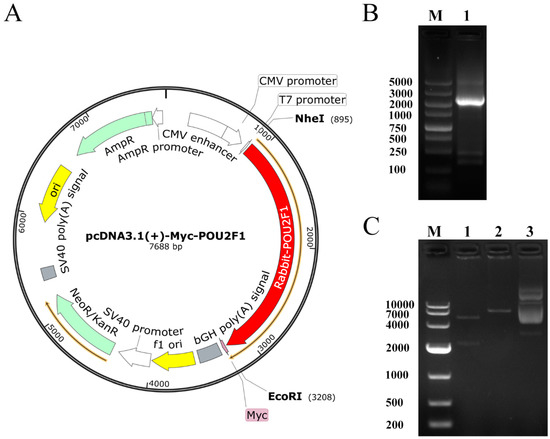
Figure 1.
Identification and construction of pcDNA3.1(+)-Myc-POU2F1. (A) Plasmid map of pcDNA3.1(+)-Myc-POU2F1. The gene sequence encoding POU2F1 was inserted into the pcDNA3.1(+)-Myc vector between the corresponding restriction sites NheI and EcoRI. (B) PCR products of POU2F1 gene. M, DL5000 DNA Marker, Lane 1, POU2F1 mRNA sequence. (C) Identification of pcDNA3.1(+)-Myc-POU2F1 digested by NheI and EcoRI. M, DL10000 DNA Marker. Lane 1, production of pcDNA3.1(+)-Myc-POU2F1 after NheI and EcoRI digestion. Lane 2, production of pcDNA3.1(+)-Myc-POU2F1 after EcoRI digestion. Lane 3, pcDNA3.1(+)-Myc-POU2F1 before digestion.
2.5. Subcellular Localization of POU2F1 Protein
PSORT (www.psort.org) was used to predict the localization of the POU2F1 protein. The pcDNA3.1(+)-Myc-POU2F1 was transferred into melanocytes using ExFect 2000 (Vazyme, Nanjing, China), according to the manufacturer’s instructions with 1 μg pcDNA3.1(+)-Myc-POU2F1 in 1 μL ExFect 2000 for each well, and cultured in a 24-well plate with CO2 at 37 °C for 24 h. Then, 4% paraformaldehyde was used to fix the cells at room temperature (RT) for 20 min, followed by penetration in 0.3% Triton X-100 (Solarbio, Beijing, China) at RT for 10 min and blocking in goat serum at 37 °C for 30 min. The cells were incubated with the primary antibody (Myc) at 4 °C overnight and with the second antibody (Cy3-conjugate) at RT for 2 h. After dying in DAPI for 10 min, the cells were observed and photographed under a fluorescent inverted microscope. The cells were washed with PBS (three times for 5 min/time) between each of the above steps.
2.6. Inhibition of POU2F1 by siRNA
Three siRNAs (with 5′ FAM modification) and negative control siRNAs were ordered from Suzhou GenePharma Co., Ltd. (Suzhou, China) (Table 2). The siRNAs were diluted into 20 μM with DEPC water, and then the complexed 2 μL diluted siRNAs and 1 μL Lipofectamine 2000 (InvitrogenTM) were transferred to the melanocytes when the cell confluence reached approximately 70%. After 24 h, fluorescence microscopy was used to detect the transfection efficiency. The total RNA extracted from the cells after transfection was used for RT-qPCR to examine the expression levels of POU2F1 and other genes, according to the above methods.
Table 2.
Primer sequences of siRNAs.
2.7. Statistical Analysis
All values in the text and figures are presented as the means ± standard deviation (SD) calculated by GraphPad Prism software (GraphPad Software, La Jolla, CA, USA). A paired T-test from SPSS 25 was performed between 2 groups (control and each test group). Statistical differences are presented at probability levels of p < 0.05 (*), p < 0.01 (**), and p < 0.001 (***).
3. Results
3.1. POU2F1 Gene Expression in Dorsal Skin of Rex Rabbits with Different Fur Colors
The expression levels of POU2F1 in the dorsal skin of Rex rabbits with different fur colors were detected by RT-qPCR. As shown in Figure 2A, POU2F1 was generally expressed in the dorsal skin of Rex rabbits of each color but showed significantly different expression levels (p < 0.05). POU2F1 had the lowest expression level in the BL-Rex rabbit, which was used as a control to calculate the fold change of POU2F1 gene expression in Rex rabbits with other fur colors. The highest expression levels of POU2F1 were seen in the BR-Rex rabbit (15.63 times), followed by the PY-Rex rabbit (12.37 times), which were significantly higher than that in Rex rabbits with the other four fur colors (p < 0.05). There was no significant difference in POU2F1 expression between CH, PC, and WH-Rex rabbits, in which POU2F1 expression was generally low at around 1–3 times the levels in the BL-Rex rabbit.
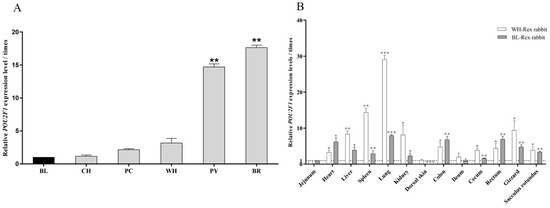
Figure 2.
Expression patterns of POU2F1 gene. (A) The expression level of POU2F1 in the dorsal skin of Rex rabbits with different fur colors. BL: black, CH: chinchilla, WH: white, BR: brown, PY: protein yellow, and PC: protein chinchilla. POU2F1 expression levels in the dorsal skin of the black Rex rabbit were used as the control to calculate the relative expression levels in Rex rabbits of other colors in figure A. (B) The expression level of POU2F1 in different organ tissues of the white and black Rex rabbit. POU2F1 expression levels in jejunum of the white and black Rex rabbit were used as controls for calculating the relative expression level in corresponding Rex rabbits in figure B. * p < 0.05, ** p < 0.01, and *** p < 0.001.
3.2. The Tissue Distribution of POU2F1 Gene Expression in WH- and BL-Rex Rabbits
POU2F1 expression in different organs of the WH- and BL-Rex rabbits is shown in Figure 2B. The relative expression levels of POU2F1 in different organs of BL-Rex rabbits normalized by three reference genes (GAPDH, β-actin and HPRT1) were generally consistent (Supplementary Figure S1). POU2F1 expression in the jejunum, which had a relatively low and stable expression level in BL- and WH-Rex rabbits, was used as a control (defined as 1) to calculate the fold change in POU2F1 expression in other organs. POU2F1 was widely expressed in different organs of the WH- and BL-Rex rabbits, and almost all other organs (except dorsal skin and ileum) had significantly higher expression levels than jejunum (p < 0.05). In addition, POU2F1 showed the highest expression levels in the lung in both WH- and BL-Rex rabbits (29.15 and 7.99 times higher, respectively), while the dorsal skin of BL-Rex rabbit had significantly lower POU2F1 expression (0.06 times).
3.3. Subcellular Localization of POU2F1 Protein
PSORT predicted that 95.7% of the POU2F1 protein would be located at the nucleus. Subcellular localization analysis was used to determine the localization of the POU2F1 protein. As shown in Figure 3A, the empty pcDNA3.1(+)-Myc vector without the POU2F1 gene was randomly distributed throughout the whole cell, including the nucleus and cytoplasm. The pcDNA3.1(+)-Myc-POU2F1 was only found in the nucleus (Figure 3B), indicating that the localization of the POU2F1 protein in the dorsal skin cells of Rex rabbits was nuclear, which is consistent with the prediction.
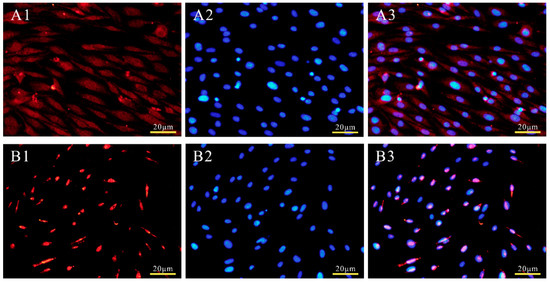
Figure 3.
Subcellular localization of POU2F1 protein in rabbit melanocytes. The pcDNA3.1(+)-Myc (A1–A3) and recombinant plasmids pcDNA3.1(+)-Myc-POU2F1 (B1–B3) were transiently transfected into rabbit melanocytes. Cells in groups A and B are melanocytes magnified 40 times by fluorescence inverted microscope. A1 (B1) is the Myc-tag dyed by Cy3, and A2 (B2) is the DAPI-dyed nucleus, which are merged in A3 (B3).
3.4. The Effects of POU2F1 Gene on Other Pigmentation-Related Genes
POU2F1 siRNA interference and overexpression were performed in melanocytes to analyze the potential functional mechanisms of POU2F1 during melanin deposition in the dorsal skin of Rex rabbits, followed by detection of the expression levels of pigment-related genes (SLC7A11, SLC24A5, MITF, MC1R, CREB1 and ASIP) by RT-qPCR. siRNA1, siRNA2, and siRNA3 were successfully transferred into melanocytes (Figure 4A); siRNA3 had the best interference efficiency (85.09%) (Figure 4B,C) and was used for POU2F1 interference in the next stage of the experiment.
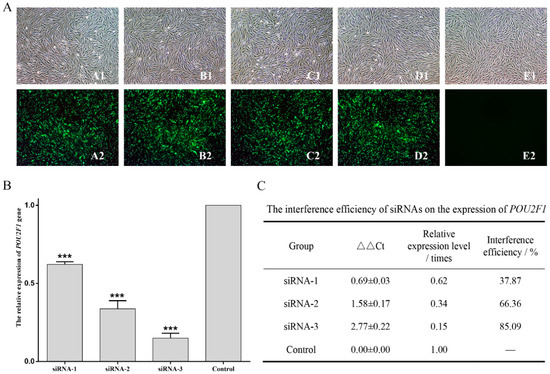
Figure 4.
Interference efficiency of siRNAs on POU2F1 expression. (A) Melanocyte morphology 24 h after FAM-siRNAs transfection. (A–E) represent siRNA-1, siRNA-2, siRNA-3, NC, and blank, respectively. (B,C) show interference efficiency of siRNAs on POU2F1 expression. *** p < 0.001.
The POU2F1 gene was overexpressed by 1326.3 times the baseline by pcDNA3.1(+)-Myc-POU2F1 (Figure 5A). Simultaneously, the SLC7A11 gene was significantly downregulated 0.55 times (Figure 5B), indicating the negative regulation of the SLC7A11 gene by POU2F1. In addition, MITF and SLC24A5 were downregulated (0.56 and 0.73 times, respectively), but ASIP was significantly upregulated 3.76 times. However, when POU2F1 expression was inhibited by siRNA3 (Figure 5C), the SLC7A11 gene showed significant overexpression at 1.58 times the baseline (Figure 5D), emphasizing the inhibitory role of POU2F1 on SLC7A11. In addition, ASIP was significantly downregulated 0.63 times, and MC1R was significantly upregulated 2.62 times.
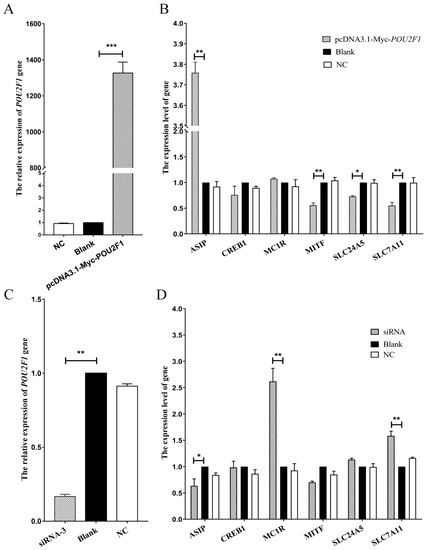
Figure 5.
Expression levels of SLC7A11 and other pigmentation-related genes after the overexpression and interference of POU2F1. (A,C) are the relative gene expression levels of POU2F1 in rabbit melanocytes transfected with pcDNA3.1(+)-Myc-POU2F1 vector and siRNA-3, respectively; (B,D) are the relative expression levels of other pigmentation-related genes in rabbit melanocytes transfected with pcDNA3.1(+)-Myc-POU2F1 vector and siRNA-3, respectively. The relative expression levels of the above genes were measured by RT-qPCR and calculated by the 2−∆∆Ct method. * p < 0.05, ** p < 0.01, and *** p < 0.001.
4. Discussion
In this study, we compared the expression levels of POU2F1 in Rex rabbits of various colors to investigate the possible impacts of the gene on fur color. POU2F1 was generally but differently expressed in the dorsal skin of Rex rabbits, revealing its potential roles in fur color formation. POU2F1 showed the lowest expression levels in BL-Rex rabbits and the highest expression levels in the BR-Rex rabbit. In addition, the opposite pattern of expression of the SLC7A11 gene was identified in Rex rabbits with the same color [14]. This is consistent with the negative regulation of the SLC7A11 gene by POU2F1, which was reported previously. Additionally, SLC7A11 was reported to positively regulate melanin levels, and higher SLC7A11 gene expression levels led to increased melanin deposition [14]. This may explain why POU2F1 and SLC7A11 showed the highest and lowest expression levels, respectively, in BL-Rex rabbits.
Additionally, the tissue distribution of POU2F1 expression was analyzed to explore potential differential expression. Because the use of GAPDH as a reference gene to correct gene expression levels in the Rex rabbit was relatively novel, another two reference genes (β-actin and HPRT1) were employed for the detection of POU2F1 expression levels. The expression levels of POU2F1 normalized by GAPDH, HPRT1 and β-actin were roughly consistent, which revealed the usability of GAPDH as a reference gene in the Rex rabbit and improved the reliability of the data in this study. POU2F1 was widely expressed in different organs of white and black Rex rabbits, with the highest expression levels in the lungs. POU2F1 has been reported to be involved in carcinogenesis and other disease processes by promoting cell proliferation and metastasis [26], e.g., in type 2 diabetes [15,27], head and neck cancer [28], cervical cancer [19], and liver cancer [29]. The wide involvement of POU2F1 in various kinds of cancer and in the immune response to cancers suggests its general expression in different organs, which was also found in this study. Interestingly, most recent studies on the POU2F1 gene have focused on its regulatory impacts on liver cancer [21,22,29]. POU2F1 was found to be highly expressed in the liver of Rex rabbits, suggesting a potential immune-related role for the POU2F1 gene, a phenomenon which deserves further research. In addition, POU2F1 showed the highest expression levels in the lungs, which may be related to the expression and important roles of SLC7A11 in response to lung cancer [30,31].
As a transcription factor, POU2F1 was predicted to be expressed and to have some biological functions in the nucleus of melanocytes of the Rex rabbit, as was reported in human HEK-293T cells [32]. A subcellular localization study confirmed the prediction that the POU2F1 protein is located at the nucleus and does not have a transmembrane structure, which are classical characteristics of transcription factors. POU2F1 played biological roles in the nucleus of melanocytes, suggesting it has a potential role as a transcription factor regulating the expression of some other genes. Overexpression and interference of POU2F1 revealed its negative regulatory effect on the SLC7A11 gene, which was consistent with the findings of our previous study. Abnormal expression of POU2F1 not only regulated the expression of the SLC7A11 gene but also some other fur-color-related genes. The positive influence of POU2F1 on the expression of ASIP was also identified in this study, which fits with the negative relationship between ASIP and SLC7A11 [14]. In addition, expression of the MC1R gene was upregulated when POU2F1 expression was affected. Both ASIP and MC1R have been reported to be involved in the melanogenesis pathway and the regulation of fur color formation [9,10,13]. Furthermore, the overexpression of POU2F1 negatively regulated the expression of SLC24A5 and MITF, which are related to pigmentation. The findings of various studies indicate the important role SLC24A5 has on human skin pigmentation: it regulates human epidermal melanogenesis and related diseases, such as nonsyndromic oculocutaneous albinism [33,34,35]. MITF plays significant roles in signal transduction and transcription in melanocytes during the color formation of skin and fur [36,37]. In addition, ASIP, MC1R, SLC24A5, and MITF were found to not only affect the formation of human skin color [38,39] but also affect fur color in different animals, including mice [37], sheep [40,41], horses [42,43,44], pigs [45], and rhesus macaques [46]. The significant associations between POU2F1 and other fur-color-related genes further revealed the potential impacts of POU2F1 on Rex rabbit fur color.
5. Conclusions
The POU2F1 gene was significantly more highly expressed in the dorsal skin of brown and protein yellow Rex rabbits compared with that of Rex rabbits with black, chinchilla, white, and protein chinchilla fur. POU2F1 affects the expression of genes, including SLC24A5, SLC7A11, MITF, MC1R, and ASIP, that are related to the formation of fur color in Rex rabbits. This study highlighted the potential regulatory role of POU2F1 in the fur color of Rex rabbits.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4425/11/5/575/s1, Figure S1: Expression level of POU2F1 in different organ tissues of Black Rex rabbit.
Author Contributions
Conceptualization, N.Y.; Formal analysis, B.Z.; Funding acquisition, X.W.; Investigation, N.Y., Z.B. and M.L.; Resources, S.H.; Supervision, X.W.; Writing—original draft, N.Y.; Writing—review & editing, Y.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Modern Agricultural Industrial System Special Funding, grant number CARS-43-A-1.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tang, J.; Zhang, Y. Color Rabbit Hair Property and Market Foreground. Adv. Mater. Res. 2011, 332, 35–40. [Google Scholar] [CrossRef]
- Kulikov, A.V.; Bazhenova, E.Y.; Kulikova, E.A.; Fursenko, D.V.; Trapezova, L.I.; Terenina, E.E.; Mormede, P.; Popova, N.K.; Trapezov, O.V. Interplay between aggression, brain monoamines and fur color mutation in the American mink. Genes Brain Behav. 2016, 15, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Cieslak, M.; Reissmann, M.; Hofreiter, M.; Ludwig, A. Colours of domestication. Biol. Rev. 2011, 86, 885–899. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.-S. An Updated Review of Tyrosinase Inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef] [PubMed]
- Sturm, R.A. Molecular genetics of human pigmentation diversity. Hum. Mol. Genet. 2009, 18, R9–R17. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kanai, Y.; Chairoungdua, A.; Cha, S.H.; Matsuo, H.; Kim, D.K.; Inatomi, J.; Sawa, H.; Ida, Y.; Endou, H. Human cystine/glutamate transporter: cDNA cloning and upregulation by oxidative stress in glioma cells. Biochim. Biophys. Acta (BBA) Biomembr. 2001, 1512, 335–344. [Google Scholar] [CrossRef]
- Chintala, S.; Li, W.; Lamoreux, M.L.; Ito, S.; Wakamatsu, K.; Sviderskaya, E.V.; Bennett, D.C.; Park, Y.-M.; Gahl, W.A.; Huizing, M.; et al. Slc7a11 gene controls production of pheomelanin pigment and proliferation of cultured cells. Proc. Natl. Acad. Sci. USA 2005, 102, 10964–10969. [Google Scholar] [CrossRef]
- Li, H.-T.; He, X.; Zhou, Z.; Zhao, S.-H.; Zhang, W.-X.; Liu, G.; Zhao, Z.-S.; Jia, B. Expression levels of Slc7a11 in the skin of Kazakh sheep with different coat colors. Hereditas (Beijing) 2012, 34, 1314–1319. [Google Scholar] [CrossRef]
- Luo, H.; Hou, J.N.; Li, Y.-M.; Yan, S.Q. Cloning and Sequence of MC1R Gene 5′-Flanking Sequence in Rabbit. China Anim. Husb. Vet. Med. 2011, 38, 89–92. [Google Scholar]
- Yang, G.-L.; Fu, D.-L.; Lang, X.; Wang, Y.-T.; Cheng, S.-R.; Fang, S.-L.; Luo, Y.-Z. Mutations in MC1R Gene Determine Black Coat Color Phenotype in Chinese Sheep. Sci. World J. 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- Thomas, A.C.; Heux, P.; Santos, C.; Arulvasan, W.; Solanky, N.; Carey, M.E.; Gerrelli, D.; Kinsler, V.A.; Etchevers, H.C. Widespread dynamic and pleiotropic expression of the melanocortin-1-receptor (MC1R) system is conserved across chick, mouse and human embryonic development. Birth Defects Res. 2018, 110, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Flesher, J.L.; Paterson-Coleman, E.K.; Vasudeva, P.; Ruiz-Vega, R.; Marshall, M.; Pearlman, E.; MacGregor, G.R.; Neumann, J.; Ganesan, A.K. Delineating the role of MITF isoforms in pigmentation and tissue homeostasis. Pigment Cell Melanoma Res. 2019, 33, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Fontanesi, L.; Forestier, L.; Allain, D.; Scotti, E.; Beretti, F.; Deretz-Picoulet, S.; Pecchioli, E.; Vernesi, C.; Robinson, T.; Malaney, J.L.; et al. Characterization of the rabbit agouti signaling protein (ASIP) gene: Transcripts and phylogenetic analyses and identification of the causative mutation of the nonagouti black coat colour. Genomics 2010, 95, 166–175. [Google Scholar] [CrossRef]
- Chen, Y.; Hu, S.; Mu, L.; Zhao, B.; Wang, M.; Yang, N.; Bao, G.; Zhu, C.; Wu, X. Slc7a11 Modulated by POU2F1 is Involved in Pigmentation in Rabbit. Int. J. Mol. Sci. 2019, 20, 2493. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.C.Y.; Lam, V.K.L.; Tam, C.H.T.; Chan, A.W.; So, W.-Y.; Ma, R.C.; Zee, B.; Waye, M.M.Y.; Mak, W.W.; Hu, C.; et al. Association of the POU class 2 homeobox 1 gene (POU2F1) with susceptibility to Type 2 diabetes in Chinese populations. Diabet. Med. 2010, 27, 1443–1449. [Google Scholar] [CrossRef]
- Kang, J.; Shen, Z.; Lim, J.-M.; Handa, H.; Wells, L.; Tantin, D. Regulation of Oct1/Pou2f1 transcription activity by O-GlcNAcylation. FASEB J. 2013, 27, 2807–2817. [Google Scholar] [CrossRef]
- Xiao, Y.; Xiang, L.; Dai, W.; Tang, W.; Wang, J.; Zhang, Y.; Wu, X.; Liu, G.; Chen, Y.; Zhu, H.; et al. The POU2F1/miR-4490/USP22 Axis Regulates Cell Proliferation and Metastasis in Gastric Cancer. Available online: https://ssrn.com/abstract=3384868 (accessed on 5 May 2019). [CrossRef]
- Dey, A.; Sen, S.; Maulik, U. Studying POU2F1 to Unveil the Structural Facet for Pan-Cancer Analysis Considering the Functional Annotations and Sequence-Structure Space Paradigm. bioRxiv 2019, 806489. [Google Scholar]
- Zhang, R.; Lu, H.; Lyu, Y.-Y.; Yang, X.-M.; Zhu, L.-Y.; Yang, G.-D.; Jiang, P.-C.; Re, Y.; Song, W.-W.; Wang, J.-H. E6/E7-P53-POU2F1-CTHRC1 axis promotes cervical cancer metastasis and activates Wnt/PCP pathway. Sci. Rep. 2017, 7, 44744. [Google Scholar] [CrossRef]
- Vázquez-Arreguín, K.; Bensard, C.; Schell, J.C.; Swanson, E.; Chen, X.; Rutter, J.; Tantin, D. Oct1/Pou2f1 is selectively required for colon regeneration and regulates colon malignancy. PLoS Genet. 2019, 15, e1007687. [Google Scholar] [CrossRef]
- Zhong, Y.; Huang, H.; Chen, M.; Huang, J.; Wu, Q.; Yan, G.-R.; Chen, D. POU2F1 over-expression correlates with poor prognoses and promotes cell growth and epithelial-to-mesenchymal transition in hepatocellular carcinoma. Oncotarget 2017, 8, 44082–44095. [Google Scholar] [CrossRef]
- Hong, Y.Z.; Guan, Y.C.; Shi, P.W.; Yu, C.; Jian, P.H. POU2F1 promotes growth and metastasis of hepatocellular carcinoma through the FAT1 signaling pathway. Am. J. Cancer Res. 2017, 7, 1665–1679. [Google Scholar]
- Chen, Y.; Hu, S.; Wang, M.; Zhao, B.; Yang, N.; Li, J.; Chen, Q.; Liu, M.; Zhou, J.; Bao, G.; et al. Characterization and Establishment of an Immortalized Rabbit Melanocyte Cell Line Using the SV40 Large T Antigen. Int. J. Mol. Sci. 2019, 20, 4874. [Google Scholar] [CrossRef] [PubMed]
- Svingen, T.; Letting, H.; Hadrup, N.; Hass, U.; Vinggaard, A.M. Selection of reference genes for quantitative RT-PCR (RT-qPCR) analysis of rat tissues under physiological and toxicological conditions. PeerJ 2015, 3, 855. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.-H.; Cao, Y.-M.; Huang, Y.; Shi, Q.-W.; Guo, J.-H.; Fan, Z.-W.; Li, J.-G.; Chen, B.-W.; Wu, B.-Y. Long non-coding RNA TUG1 contributes to tumorigenesis of human osteosarcoma by sponging miR-9-5p and regulating POU2F1 expression. Tumor Biol. 2016, 37, 15031–15041. [Google Scholar] [CrossRef] [PubMed]
- Batool, A.; Jahan, N.; Sun, Y.; Hanif, A.; Xue, H. Genetic association of IDE, POU2F1, PON1, IL1 alpha and IL1 beta with type 2 diabetes in Pakistani population. Mol. Biol. Rep. 2014, 41, 3063–3069. [Google Scholar] [CrossRef]
- Sharpe, D.J.; Orr, K.S.; Moran, M.; White, S.J.; McQuaid, S.; Lappin, T.R.; Thompson, A.; James, J. POU2F1 activity regulates HOXD10 and HOXD11 promoting a proliferative and invasive phenotype in Head and Neck cancer. Oncotarget 2014, 5, 8803–8815. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Sun, X.; Mei, C.; Zha, X. MiR-449a promotes liver cancer cell apoptosis by down-regulation of Calpain6 and POU2F1. Oncotarget 2015, 7, 13491–13501. [Google Scholar] [CrossRef]
- Moreno, J.A.; Lambros, M.P.; Ouchi, K.; Kuwahara, Y.; Iehara, T.; Konishi, E.; Hosoi, H. Abstract 3209: The expression of SLC7A11 transporter in lung and pancreatic cancer tissues at different stages of development. Tumor Biol. 2015, 75, 3209. [Google Scholar] [CrossRef]
- Ji, X.; Qian, J.; Rahman, S.M.J.; Siska, P.J.; Zou, Y.; Harris, B.K.; Hoeksema, M.D.; Trenary, I.A.; Heidi, C.; Eisenberg, R.; et al. xCT (SLC7A11)-mediated metabolic reprogramming promotes non-small cell lung cancer progression. Oncogene 2018, 37, 5007–5019. [Google Scholar] [CrossRef]
- Gao, J.; Chen, Y.; Yang, Y.; Liang, J.; Xie, J.; Liu, J.; Li, S.; Zheng, G.; Xie, L.; Zhang, R. The transcription factor Pf-POU3F4 regulates expression of the matrix protein genes Aspein and Prismalin-14 in pearl oyster (Pinctada fucata). FEBS J. 2016, 283, 1962–1978. [Google Scholar] [CrossRef] [PubMed]
- Wei, A.; Zang, D.-J.; Zhang, Z.; Liu, X.-Z.; He, X.; Yang, L.; Wang, Y.; Zhou, Z.; Zhang, M.-R.; Dai, L.-L.; et al. Exome Sequencing Identifies SLC24A5 as a Candidate Gene for Nonsyndromic Oculocutaneous Albinism. J. Investig. Dermatol. 2013, 133, 1834–1840. [Google Scholar] [CrossRef] [PubMed]
- Ginger, R.S.; Askew, S.E.; Ogborne, R.M.; Wilson, S.; Ferdinando, D.; Dadd, T.; Smith, A.M.; Kazi, S.; Szerencsei, R.T.; Winkfein, R.J.; et al. SLC24A5 Encodes atrans-Golgi Network Protein with Potassium-dependent Sodium-Calcium Exchange Activity That Regulates Human Epidermal Melanogenesis. J. Biol. Chem. 2007, 283, 5486–5495. [Google Scholar] [CrossRef] [PubMed]
- Lamason, R.; Mohideen, M.-A.P.; Mest, J.R.; Wong, A.C.; Norton, H.L.; Aros, M.C.; Jurynec, M.; Mao, X.; Humphreville, V.R.; Humbert, J.E.; et al. SLC24A5, a Putative Cation Exchanger, Affects Pigmentation in Zebrafish and Humans. Science 2005, 310, 1782–1786. [Google Scholar] [CrossRef] [PubMed]
- Goding, C.R. Mitf from neural crest to melanoma: Signal transduction and transcription in the melanocyte lineage. Genes Dev. 2000, 14, 1712–1728. [Google Scholar] [PubMed]
- Bauer, G.L.; Praetorius, C.; Bergsteinsdóttir, K.; Hallsson, J.H.; Gísladóttir, B.K.; Schepsky, A.; Swing, D.A.; O’Sullivan, T.N.; Arnheiter, H.; Bismuth, K.; et al. The Role of MITF Phosphorylation Sites During Coat Color and Eye Development in Mice Analyzed by Bacterial Artificial Chromosome Transgene Rescue. Genetics 2009, 183, 581–594. [Google Scholar] [CrossRef]
- Jacobs, L.C.; Hamer, M.A.; Gunn, D.A.; Deelen, J.; Lall, J.S.; Van Heemst, D.; Uh, H.-W.; Hofman, A.; Uitterlinden, A.G.; Griffiths, C.E.M.; et al. A Genome-Wide Association Study Identifies the Skin Color Genes IRF4, MC1R, ASIP, and BNC2 Influencing Facial Pigmented Spots. J. Investig. Dermatol. 2015, 135, 1735–1742. [Google Scholar] [CrossRef]
- Landi, M.T.; Kanetsky, P.A.; Tsang, S.; Gold, B.; Munroe, D.; Rebbeck, T.; Swoyer, J.; Ter-Minassian, M.; Hedayati, M.; Grossman, L.; et al. MC1R, ASIP, and DNA Repair in Sporadic and Familial Melanoma in a Mediterranean Population. J. Natl. Cancer Inst. 2005, 97, 998–1007. [Google Scholar] [CrossRef]
- Fontanesi, L.; Dall’Olio, S.; Beretti, F.; Portolano, B.; Russo, V. Coat colours in the Massese sheep breed are associated with mutations in the agouti signalling protein (ASIP) and melanocortin 1 receptor (MC1R) genes. Animal 2010, 5, 8–17. [Google Scholar] [CrossRef]
- Han, J.; Yang, M.; Yue, Y.; Guo, T.; Liu, J.; Niu, C.; Yang, B. Analysis of agouti signaling protein (ASIP) gene polymorphisms and association with coat color in Tibetan sheep (Ovis aries). Genet. Mol. Res. 2015, 14, 1200–1209. [Google Scholar] [CrossRef]
- Rieder, S.; Taourit, S.; Mariat, D.; Langlois, B.; Guérin, G. Mutations in the agouti (ASIP), the extension (MC1R), and the brown (TYRP1) loci and their association to coat color phenotypes in horses (Equus caballus). Mamm. Genome 2001, 12, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Kreuzer, S. A Mutation in the Equine SLC24A5 Gene Is Associated With A Dilution Of Black Horses. In Proceedings of the World Congress on Genetics Applied to Livestock Production, Leipzig, Germany, 1–6 August 2010. [Google Scholar]
- Haase, B.; Signer-Hasler, H.; Binns, M.M.; Obexer-Ruff, G.; Hauswirth, R.; Bellone, R.R.; Burger, D.; Rieder, S.; Wade, C.M.; Leeb, T. Accumulating Mutations in Series of Haplotypes at the KIT and MITF Loci Are Major Determinants of White Markings in Franches-Montagnes Horses. PLoS ONE 2013, 8, e75071. [Google Scholar] [CrossRef] [PubMed]
- Mao, H.; Ren, J.; Ding, N.; Xiao, S.; Huang, L. Genetic variation within coat color genes of MC1R and ASIP in Chinese brownish red Tibetan pigs. Anim. Sci. J. 2010, 81, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Bradley, B.J.; Gerald, M.S.; Widdig, A.; Mundy, N.I. Coat Color Variation and Pigmentation Gene Expression in Rhesus Macaques (Macaca mulatta). J. Mamm. Evol. 2012, 20, 263–270. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).