Tamarind Trypsin Inhibitor in Chitosan–Whey Protein Nanoparticles Reduces Fasting Blood Glucose Levels without Compromising Insulinemia: A Preclinical Study

 , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Tamarind Trypsin Inhibitor (TTI) Extraction and Quantification
2.2. Synthesis and Characterization of Nanoparticles
2.3. The Half Maximal Inhibitory Concentration (IC50) and Incorporation Efficiency of TTI in Nanoparticles (%)
2.4. In Vitro Stability in Simulated Physiological Conditions of the Gastrointestinal Tract
2.4.1. Oral
2.4.2. Gastric
2.4.3. Intestinal
2.5. Effect of ECW on Biochemical Parameters (Lipid Profile, Glycemia, Insulin, HOMA—IR, and HOMA-β) in Wistar Rats Overfed with HGLI Diet
2.5.1. Preclinical Studies
2.5.2. Biochemical Parameters
2.5.3. Microscopic Analysis of the Pancreas
2.6. Statistical Analysis
3. Results
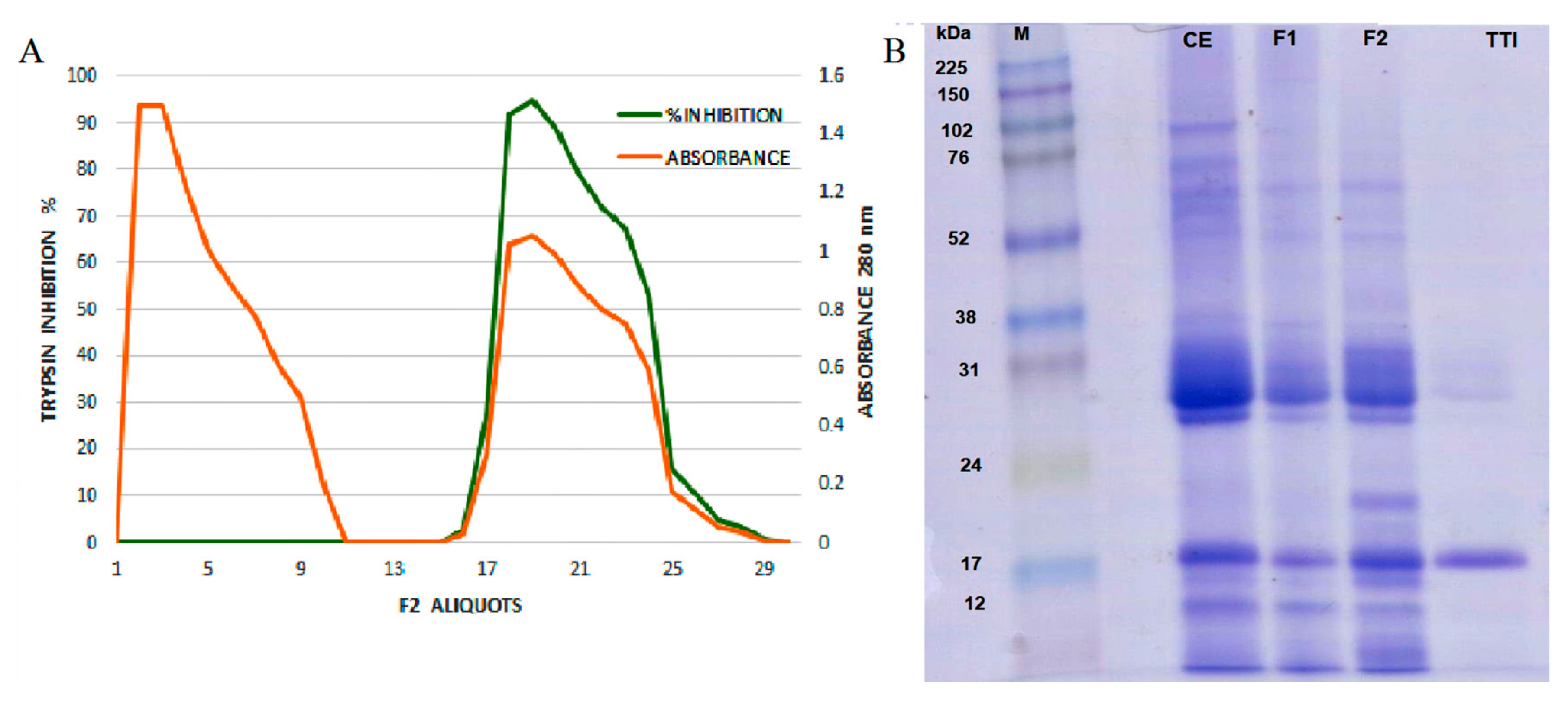
3.1. Tamarind Trypsin Inhibitor (TTI) Extraction and Quantification
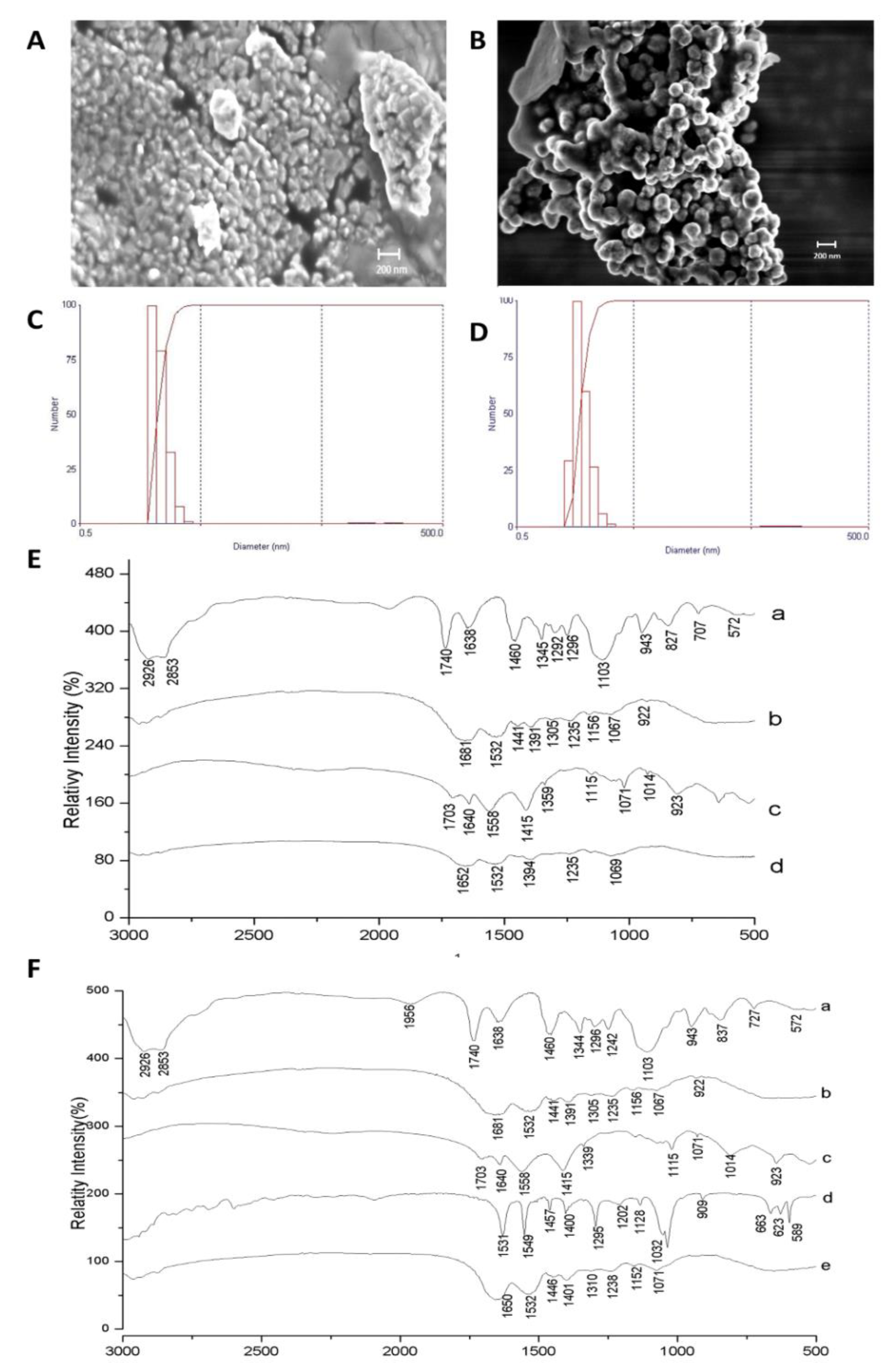
3.2. Characterization of the Nanoformulations Obtained (CW and ECW)
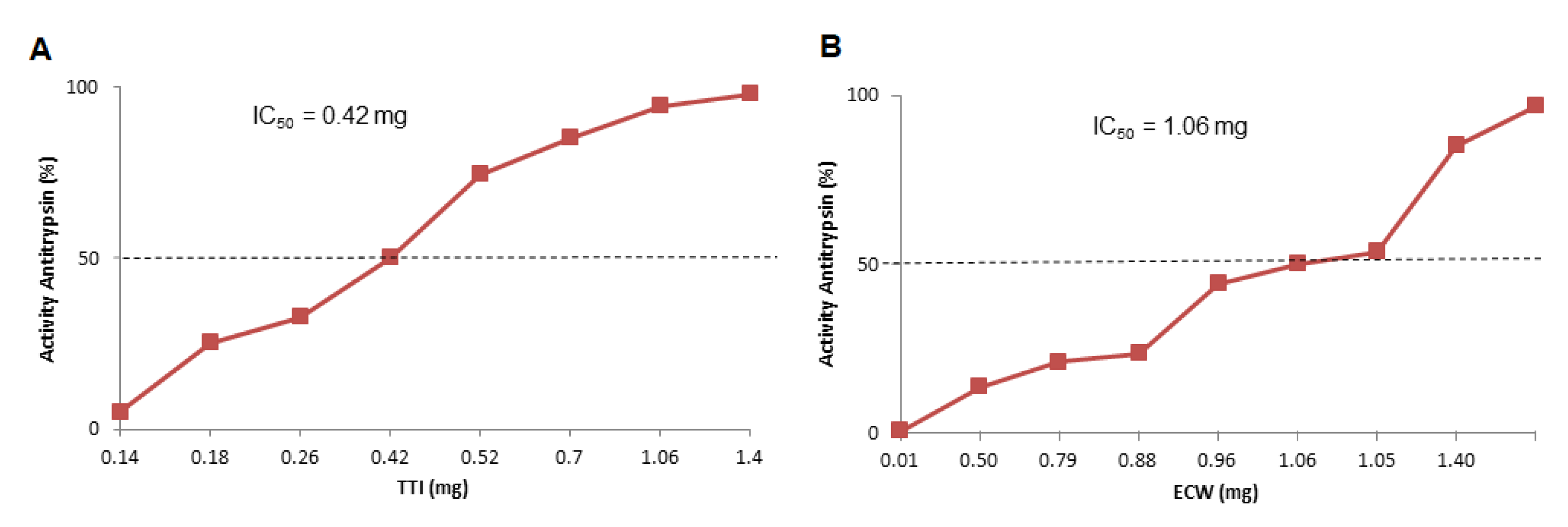
3.3. Encapsulation Incorporation Efficiency
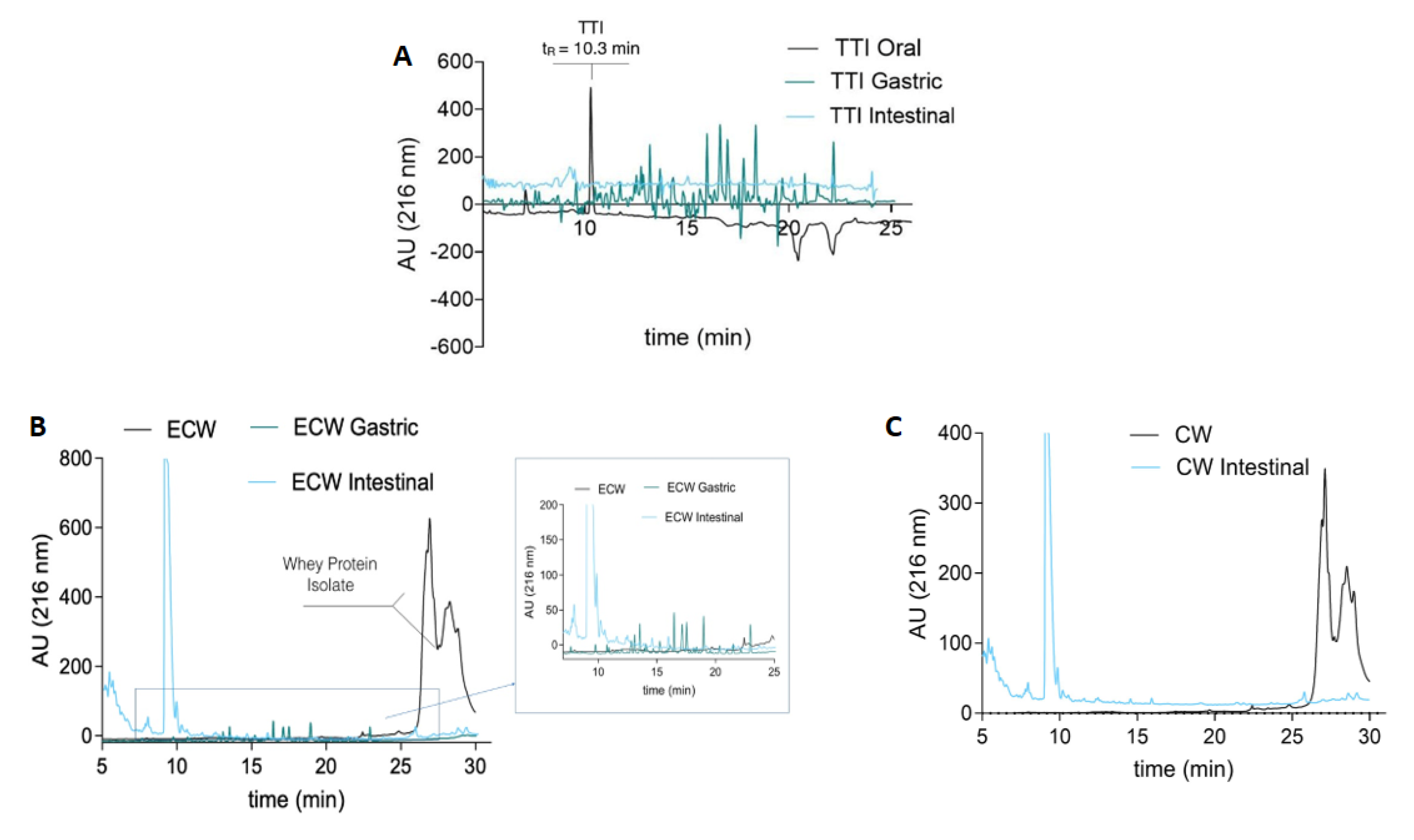
3.4. In Vitro Stability in Simulated Physiological Conditions of the Gastrointestinal Tract
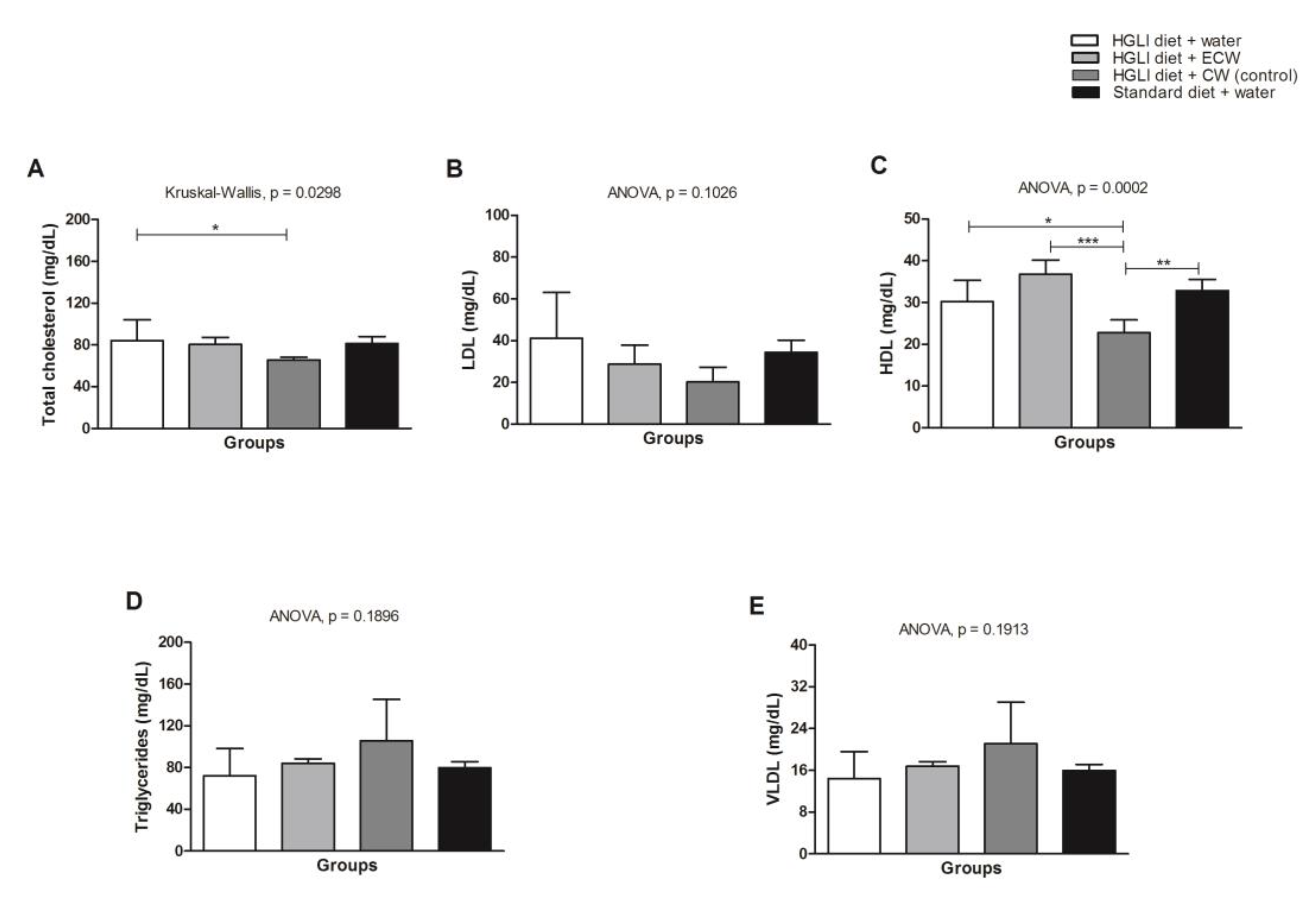
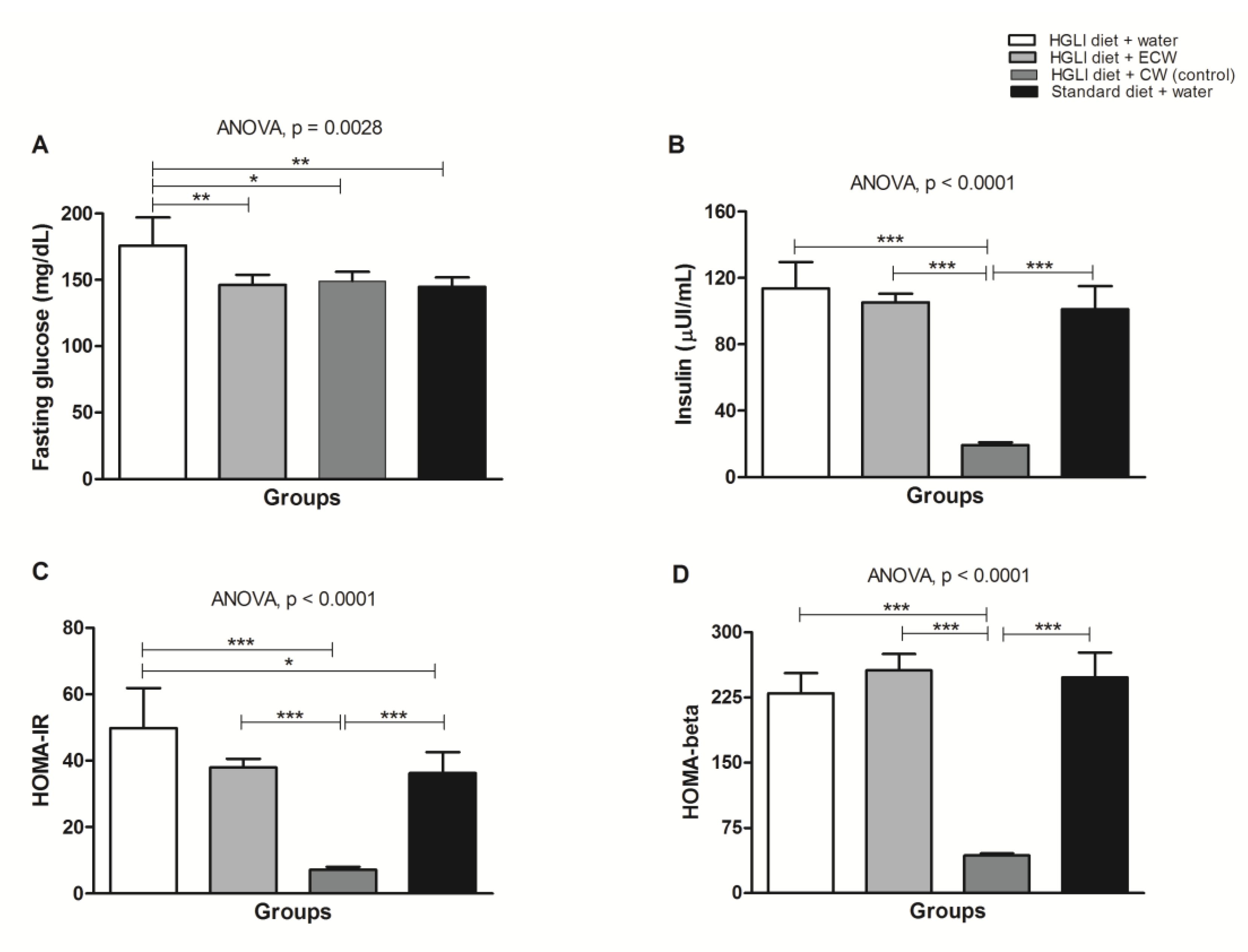
3.5. Evaluation of In Vivo Biochemical Parameters
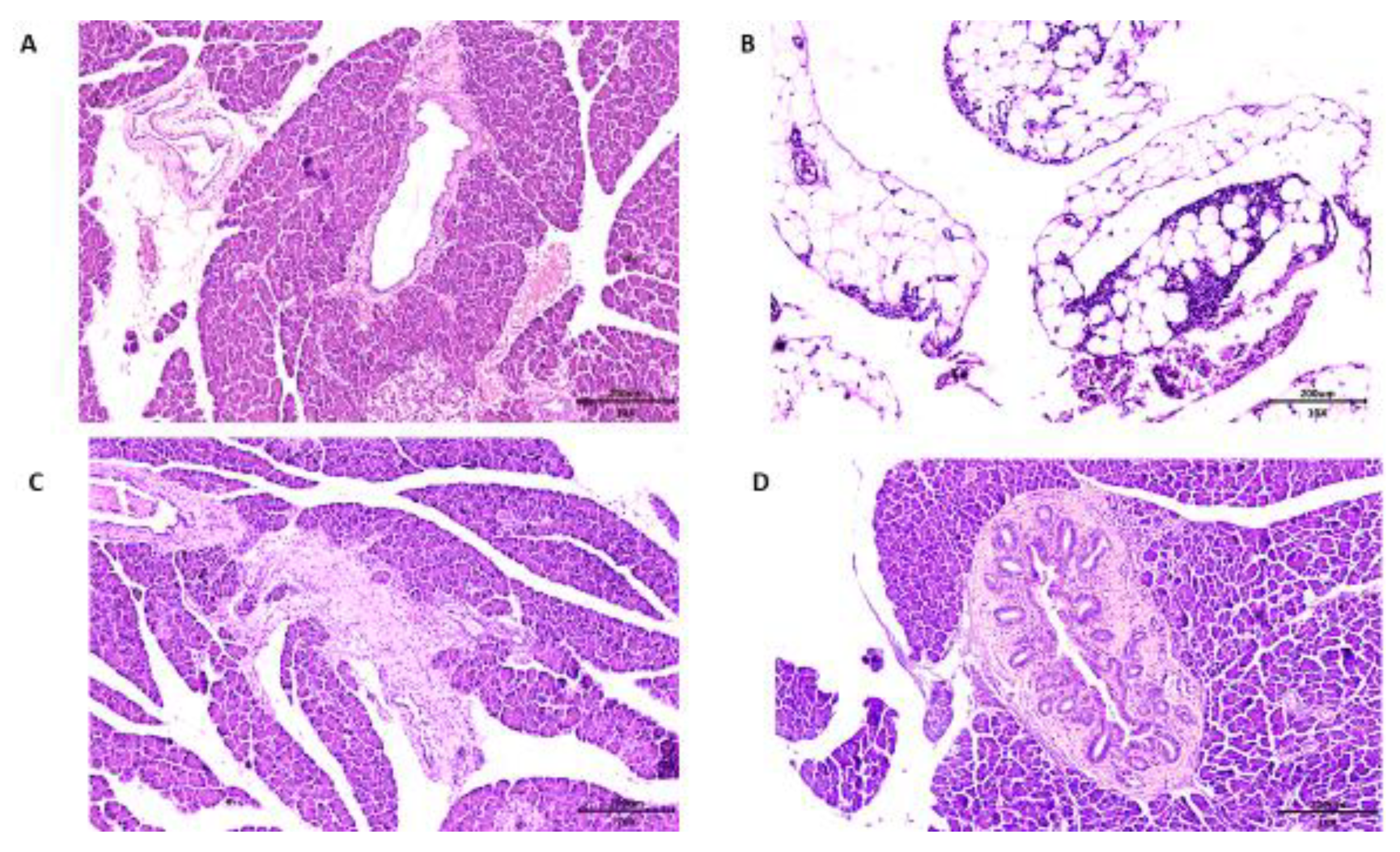
3.6. Pancreas Histopathology of Wistar Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sempértegui, F.; Estrella, B.; Tucker, K.L.; Hamer, D.H.; Narvaez, X.; Sempértegui, M.; Griffiths, J.K.; Noel, S.E.; Dallal, G.E.; Selhub, J.; et al. Metabolic syndrome in elderly living in marginal peri-urban communities in Quito, Ecuador. Public Health Nutr. 2010, 14, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Housseinpour-niazi, S.; Mirmiran, P.; Sohrab, G.; Hosseini-Esfahani, F.; Azizi, F. Inverse association between fruit, legume, and cereal fiber and the risk of metabolic syndrome: Tehrn Lipid and Glucose study. Diabetes Res. Clin. Pract. 2011, 94, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Lee, J.E.; Song, W.O.; Paik, H.Y.; Song, Y. Carbohydrate intake and refined-grain consumption in the Korean adult population. J. Acad. Nutr. Diet. 2014, 114, 54–62. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association Diabetes Care. Pharmacologic Approaches to Glycemic Treatment. Stand. Med. Care Diabetes 2019, 42, 90–102. [Google Scholar]
- Choudhury, H.; Pandey, M.; Hua, C.K.; Mun, C.S.; Jing, J.K.; Kong, L.; Ern, L.Y.; Ashraf, N.A.; Kit, S.W.; Yee, T.S.; et al. An update on natural compounds in the remedy of diabetes mellitus: A systematic review. J. Tradit. Complement. Med. 2018, 8, 361–376. [Google Scholar] [CrossRef] [PubMed]
- De Castro, R.J.S.; Domingues, M.A.F.; Ohara, A.; Okuro, P.K.; dos Santos, J.G.; Brexo, R.P.; Sato, H.H. Whey protein as a key component in food systems: Physicochemical properties, production technologies and applications. Food Struct. 2017, 14, 17–29. [Google Scholar] [CrossRef]
- Ozorio, L.; Silva, L.P.; Prates, M.V.; Bloch, C., Jr.; Takeiti, C.Y.; Gomes, D.M.; da Silva-Santos, J.E.; Deliza, R.; Brígida, A.I.S.; Furtado, A.; et al. Whey hydrolysate-based ingredient with dual functionality: From production to consumer’s evaluation. Food Res. Int. 2019, 122, 123–128. [Google Scholar] [CrossRef]
- Kohda, N.; Inoue, S.; Noda, T.; Saito, T. Effects of a chitosan intake on the fecal excretion of dioxins and fat in rats. Biosci. Biotechnol. Biochem. 2012, 76, 1544–1548. [Google Scholar] [CrossRef][Green Version]
- Jae-Young, J.; Pyo-Jam, P.; Bokyung, K.; Se-Kwon, K. Antihypertensive activity of chitin derivatives. Biopolymers 2006, 83, 250–254. [Google Scholar]
- Yemişci, M.; Gürsoy-Özdemir, Y.; Caban, S.; Bodur, E.; Çapan, Y.; Dalkara, T. Transport of a Caspase Inhibitor Across the Blood–Brain Barrier by Chitosan Nanoparticles. Nanomedicine—Cancer, Diabetes, and Cardiovascular, Central Nervous System, Pulmonary and Inflammatory Diseases. Methods Enzymol. 2012, 508, 253–269. [Google Scholar]
- Serquiz, A.C.; Machado, R.J.; Serquiz, R.P.; Lima, V.C.; de Carvalho, F.M.C.; Carneiro, M.A.; Maciel, B.L.; Uchôa, A.F.; Santos, E.A.; Morais, A.H. Supplementation with a new trypsin inhibitor from peanut is associated with reduced fasting glucose, weight control, and increased plasma CCK secretion in an animal model. J. Enzyme Inhib. Med. Chem. 2016, 29, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.; Serquiz, A.C.; Silva, P.; Barbosa, P.; Sampaio, T.B.M.; Junior, A.; Oliveira, A.S.D.; Machado, R.J.A.; Maciel, B.L.L.; Uchôa, A.F.; et al. Trypsin inhibitor from Tamarindusindica L. seeds reduces weight gain and food consumption and increases plasmatic cholecystokinin levels. Clinics 2015, 70, 136–143. [Google Scholar] [CrossRef]
- Carvalho, F.; Lima, V.; Costa, I.; Medeiros, A.; Serquiz, A.; Lima, M.; Serquiz, R.; Maciel, B.; Uchôa, A.; Santos, E.; et al. A Trypsin Inhibitor from Tamarind Reduces Food Intake and Improves Inflammatory Status in Rats with Metabolic Syndrome Regardless of Weight Loss. Nutrients 2016, 8, 544. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.; Lima, V.C.; Costa, I.S.; Luz, A.; Ladd, F.V.; Serquiz, A.C.; Bortolin, R.H.; Silbiger, V.N.; Maciel, B.L.; Santos, E.A.; et al. Anti TNF-α Agent Tamarind Kunitz Trypsin Inhibitor Improves Lipid Profile of Wistar Rats Presentins Dyslipidemia and Diet-induced Obesity Regardlessof PPAT-Induction. Nutrients 2019, 11, 512. [Google Scholar] [CrossRef]
- Muhammad, D.R.A.; Doost, A.S.; Gupta, V.; Sintangad, M.D.; de Walle, D.V.; de Meeren, P.V.; Dewettincka, K. Stability and functionality of xanthan gum-shellac nanoparticles for the encapsulation of cinnamon bark extract. Food Hydrocoll. 2020, 100, 105377. [Google Scholar] [CrossRef]
- De Queiroz, J.L.C.; Costa, R.O.D.A.; Matias, L.L.R.; De Medeiros, A.F.; Gomes, A.F.T.; Pais, T.D.S.; Passos, T.S.; Maciel, B.L.L.; Dos Santos, E.A.; Morais, A.H.D.A. Chitosan-whey protein nanoparticles improve encapsulation efficiency and stability of a trypsin inhibitor isolated from Tamarindusindica L. Food Hydrocoll. 2018, 84, 247–256. [Google Scholar] [CrossRef]
- Costa, R.O.A. Avaliação da Citotoxicidade e Toxicidade Subaguda do Inibidor de Tripsina de Sementes de Tamarindo (Tamarindus, L. indica L.) Nanoencapsulado. Master’s Thesis, Federal University of Rio Grande, Natal, Brazil, 2019. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kakade, M.L.; Simons, N.; Liener, I. An evaluation of natural vs synthetic substrates for measuring antitryptic activity of soy bean sample. Cereal Chem. 1969, 46, 518–526. [Google Scholar]
- Kumari, R.; Gupta, S.; Singh, A.R.; Ferosekhan, S.; Kothari, D.C.; Pal, A.K.; Jadhao, S.B. Chitosan Nanoencapsulated Exogenous Trypsin Biomimics Zymogen-Like Enzyme in Fish Gastrointestinal Tract. PLoS ONE 2016, 8, e74743. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carriere, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Committiee for de Update of the Guide of the Care and Use of Laboratory Animals. Guide of Care and Use of Laboratory Animals; Division of Earth and Life Studies, Institute of Laboratory Animals Research: Washington, DC, USA, 2005; Volume 202, pp. 334–3313. [Google Scholar]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and B-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, M.A.; Mazaro, R. Princípios Éticos e Práticos do uso de Animais de Experimentação; UNIFESP: São Paulo, Brazil, 2004. [Google Scholar]
- Martins, L.B.; de Oliveira, M.C.; Menezes-Garcia, Z.; Rodrigues, D.F.; Lana, J.P.; Vieira, L.Q.; Teixeira, M.M.; Ferreira, A.V.M. Paradoxical role of tumor of necrosis factor on metabolic dysfunction and adipose tissue expansion in mice. Nutrition 2018, 50, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Costa, I.S.; Medeiros, A.F.; Carvalho, F.M.; Lima, V.C.; Serquiz, R.P.; Serquiz, A.C.; Silbiger, V.N.; Bortolin, R.H.; Maciel, B.L.; Santos, E.A.; et al. Satietogenic Protein from Tamarind Seeds Decreases Food Intake, Leptin Plasma and Gene Expression in Obese Wistar Rats. Obes. Facts 2018, 11, 440–453. [Google Scholar] [CrossRef] [PubMed]
- Pagels, R.F.; Prud’homme, R.K. Polymeric nanoparticles and microparticles for the delivery of peptides, biologics, and soluble therapeutics. J. Control. Release 2015, 219, 519–535. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Guidance on risk assessment of the aplication of nanosciense and nanotechnologies in the food and feed chain: Part 1, human and animal health. EFSA J. 2018, 16, 5327. [Google Scholar]
- Chanphai, P.; Tajmir-Riahi, H.A. Trypsin and trypsin inhibitor bind PAMAM nanoparticles: Effect of hydrophobicity on protein-polymer conjugation. J. Colloid Interface Sci. 2016, 461, 419–424. [Google Scholar] [CrossRef]
- Chanphai, P.; Tajmir-Riahi, H.A. Chitosan nanoparticles conjugate with trypsin and trypsin inhibitor. Carbohydr. Polym. 2016, 144, 346–352. [Google Scholar] [CrossRef]
- Kannel, W.B. Coronary heart disease risk factors in the elderly. Am. J. Geriatr. Cardiol. 2002, 11, 101–107. [Google Scholar] [CrossRef]
- Singh, J.; Cumming, E.; Manoharan, G.; Kalasz, H.; Adeghate, E. Medicinal chemistry of the anti-diabetic effects of Momordica charantia: Active constituents and modes of actions. Open. Med. Chem. J. 2011, 5, 70–77. [Google Scholar] [CrossRef]
- Lo, H.Y.; Ho, T.Y.; Li, C.C.; Chen, J.C.; Liu, J.J.; Hsiang, C.Y. A novel insulin receptor-binding protein from Momordica charantia enhances glucose uptake and glucose clearance in vitro and in vivo through triggering insulin receptor signaling pathway. J. Agric. Food Chem. 2014, 62, 8952–8966. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.Y.; Ho, T.Y.; Lin, C.; Li, C.C.; Hsiang, C.Y. Momordica charantia and its novel polypeptide regulate glucose homeostasis in mice via binding to insulin receptor. J. Agric. Food Chem. 2013, 61, 2461–2468. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.Y.; Li, C.C.; Ho, T.Y.; Hsiang, C.Y. Identification of the bioactive and consensus peptide motif from Momordica charantia insulin receptor-binding protein. Food Chem. 2016, 204, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Mahboobi, S.; Rahimi, F.; Jafarnejad, S. Effects of Prebiotic and Synbiotic Supplementation on Glycaemia and Lipid Profile in Type 2 Diabetes: A Meta-Analysis of Randomized Controlled Trials. Adv. Pharm. Bull. 2018, 8, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.H.P.; Hae-Soo, K. Antidiabetic Activity of Nanopowdered Chitosan in db/db Mice MiHwa. Food Sci. Biotechnol. 2010, 19, 1245–1250. [Google Scholar] [CrossRef]
- Liu, S.H.; Chang, Y.H.; Chiang, M.T. Chitosan Reduces Gluconeogenesis and Increases Glucose Uptake in Skeletal Muscle in Streptozotocin-Induced Diabetic Rats. J. Agric. Food Chem. 2010, 58, 5795–5800. [Google Scholar] [CrossRef]
- Zhao, L.; Sun, T.; Wang, L. Chitosan oligosaccharide improves the therapeutic efficacy of sitagliptin for the therapy of Chinese elderly patients with type 2 diabetes mellitus. Clin. Risk Manag. 2017, 13, 739–750. [Google Scholar] [CrossRef]
- Liu, X.; Yang, F.; Song, T.; Zeng, A.; Whang, Q.; Sun, S.; Shen, J. Therapeutic Effect of Carboxymethylated and Quanternized Chitosan on Insulin Resistance in High-Fat-Diet-Induced Rats and 3T3-L1 Adipocytes. J. Biomater. Sci. Polym. 2012, 23, 1271–1284. [Google Scholar] [CrossRef]
- Petersen, B.L.; Ward, L.S.; Bastian, E.D.; Jenkins, A.L.; Campbell, J.; Vuksan, V. A whey protein supplement decreases post-prandial glycemia. Nutr. J. 2009, 8, 47. [Google Scholar] [CrossRef]
- Bravata, D.M.; Wells, C.K.; Concato, J.; Kernan, W.N.; Brass, L.M.; Gulanski, B.I. Two measures of insulin sensitivity provided similar information in US population. J. Clin. Epidemiol. 2004, 57, 1214–1217. [Google Scholar] [CrossRef]
- Oliveira, E.P.; Lima, M.D.; Souza, A.; Mirabeau, L.A. Síndrome Metabólica, seus fenótipos e resistência à insulina pelo HOMA-RI. Arq. Bras. Endocrinol. Metabol. 2007, 5, 1506–1515. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lima, V.C.O.; Piuvezam, G.; Maciel, B.L.L.; Morais, A.H.A. Trypsin inhibitors: Promising candidate satietogenic proteins as complementary treatment for obesity and metabolic disorders? J. Enzym. Inhib. Med. Chem. 2019, 34, 405–419. [Google Scholar]
- Van der Gronde, T.; Hartog, A.; Van Hees, C.; Pellikaan, H.; Pieters, T. Systematic review of the mechanisms and evidence behind the hypocholesterolaemic effects of HPMC, pectin and chitosan in animal trials. Food Chem. 2016, 15, 746–759. [Google Scholar] [CrossRef] [PubMed]
- Hernández-González, S.O.; González-Ortiz, M.; Martínez-Abundis, E.; Robles-Cervantes, J.A. Chitosan improves insulin sensitivity as determined by the euglycemic-hyperinsulinemic clamp technique in obese subjects. Nutr. Res. 2010, 30, 392–395. [Google Scholar] [CrossRef]
- Karadeniz, F.; Kim, S.K. Antidiabetic activities of chitosan and its derivatives: A mini review. Adv. Food Nutr. Res. 2014, 73, 33–44. [Google Scholar]
- Zhou, Q.; Melton, D.A. Pancreas regeneration. Nature 2018, 557, 351–358. [Google Scholar] [CrossRef]
- Segerstolpe, Å.; Palasantza, A.; Eliasson, P.; Andersson, E.M.; Andréasson, A.C.; Sun, X.; Picelli, S.; Sabirsh, A.; Clausen, M.; Bjursell, M.K.; et al. Single-Cell Transcriptome Profiling of Human Pancreatic Islets in Health and Type 2 Diabetes. Cell Metab. 2016, 24, 593–607. [Google Scholar] [CrossRef]
- Luz, A.B.S.; dos Santos Figueredo, J.B.; Salviano, B.D.P.D.; Aguiar, A.J.F.C.; Pinheiro, L.G.S.D.; Krause, M.F.D.; da Silva Camillo, C.; Ladd, F.V.L.; Bortolin, R.H.; Silbiger, V.N.; et al. Adipocytes and intestinal epithelium dysfunctions linking obesity to inflammation induced by high glycemic index pellet-diet in Wistar rats. Biosci. Rep. 2018, 38, BSR20180304. [Google Scholar] [CrossRef]
- Ragavan, B.; Krishnakumari, S. Efect of Tarjuna stem bark extract on histopathology of liver, kidney and pancreas of alloxaninduced diabetic rats. Afr. J. Biomed. Res. 2006, 9, 189–197. [Google Scholar]
- Simon, J.P.; Baskaran, U.L.; Shallauddin, K.B.; Ramalingam, G.; Prince, S.E. Evidence of antidiabetic activity of Spirulinafusiformis against streptozotocin-induced diabetic Wistar albino rats. Biotech 2018, 8, 129. [Google Scholar]
- Castro, C.A.D.; da Silva, K.A.; Buffo, M.M.; Pinto, K.N.Z.; Duarte, F.D.O.; Nonaka, K.O.; Aníbal, F.D.F.; Duarte, A. Experimental type 2 diabetes induction reduces serum vaspin, but not serum omentin, in Wistar rats. Int. J. Exp. Pathol. 2017, 98, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Eliakim-Ikechukwu, C.F.; Obri, I. Histological changes in the pancreas following administration of ethanolic extract of alchorneacordifolia leaf in alloxan-induced diabetic Wistar rats. Niger. J. Physiol. Sci. 2009, 24, 153–155. [Google Scholar] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical | Stock Solution | SSF (pH 7) | SGF (pH 3) | SIF (pH 7) | ||||
|---|---|---|---|---|---|---|---|---|
| g/L | mol/L | mL | mmol/L | mL | mmol/L | mL | mmol/L | |
| KCl | 37.3 | 0.5 | 15.1 | 15.1 | 6.9 | 6.9 | 6.8 | 6.8 |
| KH2PO4 | 68.0 | 0.5 | 3.7 | 3.7 | 0.9 | 0.9 | 0.8 | 0.8 |
| NaHCO3 | 84.0 | 1.0 | 6.8 | 13.6 | 12.5 | 25.0 | 42.5 | 85.0 |
| NaCl | 117 | 2.0 | - | - | 11.8 | 47.2 | 9.6 | 38.4 |
| MgCl2(H2O)6 | 30.5 | 0.15 | 0.5 | 0.15 | 0.4 | 0.12 | 1.1 | 0.33 |
| (NH4)2CO3 | 48.0 | 0.5 | 0.06 | 0.06 | 0.5 | 0.5 | - | - |
| CaCl2(H2O)2 | 44.1 | 0.3 | 1.5 | 0.15 | 0.6 | |||
| HCl | - | 6.0 | 0.9 | 1.1 | 13.0 | 15.6 | 0.7 | 8.4 |
| Groups | N | Treatment |
|---|---|---|
| 1 | 5 | HGLI Diet + 1 mL of water per gavage |
| 2 | 5 | HGLI Diet + 1 mL of ECW (12.5 mg/kg per gavage) |
| 3 | 5 | HGLI Diet + 1mL of CW (10.0 mg/kg per gavage) |
| 4 | 5 | Standard Diet + 1 mL of water per gavage |
| Parameters | Mean (SD) |
|---|---|
| Fasting blood glucose (mg/dL) | 88.80 (17.87) |
| Insulin (µU/mL) | 12.17 (0.73) |
| HOMA-IR | 2.67 (0.63) |
| HOMA—BETA | 47.44 (9.07) |
| Total cholesterol (mg/dL) | 112.00 (54.00) |
| HDL-c (mg/dL) | 23.40 (4.04) |
| LDL-c (mg/dL) | 22.76 (4.05) |
| VLDL-c (mg/dL) | 20.65 (5.59) |
| Triglycerides (mg/dL) | 100.18 (29.80) |
| Antitrypsin Activity (%) | |||||
|---|---|---|---|---|---|
| Aliquots | TTI Mean % | ECW Mean% | Water Mean % | TTI * Mean % | ECW * Mean% |
| Control | - | - | - | 100 | 100 |
| Oral | 100 | 100 | 0.0 | 100 | 100 |
| Gastric | 0.0 | 95.5 | 0.0 | 85.0 | 100 |
| Intestinal | - | - | 0.0 | 50.0 | 90.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matias, L.L.R.; Costa, R.O.A.; Passos, T.S.; Queiroz, J.L.C.; Serquiz, A.C.; Maciel, B.L.L.; Santos, P.P.A.; Camillo, C.S.; Gonçalves, C.; Amado, I.R.; et al. Tamarind Trypsin Inhibitor in Chitosan–Whey Protein Nanoparticles Reduces Fasting Blood Glucose Levels without Compromising Insulinemia: A Preclinical Study. Nutrients 2019, 11, 2770. https://doi.org/10.3390/nu11112770
Matias LLR, Costa ROA, Passos TS, Queiroz JLC, Serquiz AC, Maciel BLL, Santos PPA, Camillo CS, Gonçalves C, Amado IR, et al. Tamarind Trypsin Inhibitor in Chitosan–Whey Protein Nanoparticles Reduces Fasting Blood Glucose Levels without Compromising Insulinemia: A Preclinical Study. Nutrients. 2019; 11(11):2770. https://doi.org/10.3390/nu11112770
Chicago/Turabian StyleMatias, Lídia L. R., Rafael O. A. Costa, Thaís S. Passos, Jaluza L. C. Queiroz, Alexandre C. Serquiz, Bruna L. L. Maciel, Pedro P. A. Santos, Christina S. Camillo, Catarina Gonçalves, Isabel R. Amado, and et al. 2019. "Tamarind Trypsin Inhibitor in Chitosan–Whey Protein Nanoparticles Reduces Fasting Blood Glucose Levels without Compromising Insulinemia: A Preclinical Study" Nutrients 11, no. 11: 2770. https://doi.org/10.3390/nu11112770
APA StyleMatias, L. L. R., Costa, R. O. A., Passos, T. S., Queiroz, J. L. C., Serquiz, A. C., Maciel, B. L. L., Santos, P. P. A., Camillo, C. S., Gonçalves, C., Amado, I. R., Pastrana, L., & Morais, A. H. A. (2019). Tamarind Trypsin Inhibitor in Chitosan–Whey Protein Nanoparticles Reduces Fasting Blood Glucose Levels without Compromising Insulinemia: A Preclinical Study. Nutrients, 11(11), 2770. https://doi.org/10.3390/nu11112770