The Effects of the Combination of Oral Lactoferrin and Iron Injection on Iron Homestasis, Antioxidative Abilities and Cytokines Activities of Suckling Piglets


Simple Summary
Abstract
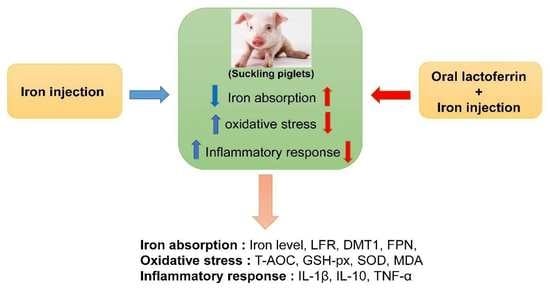
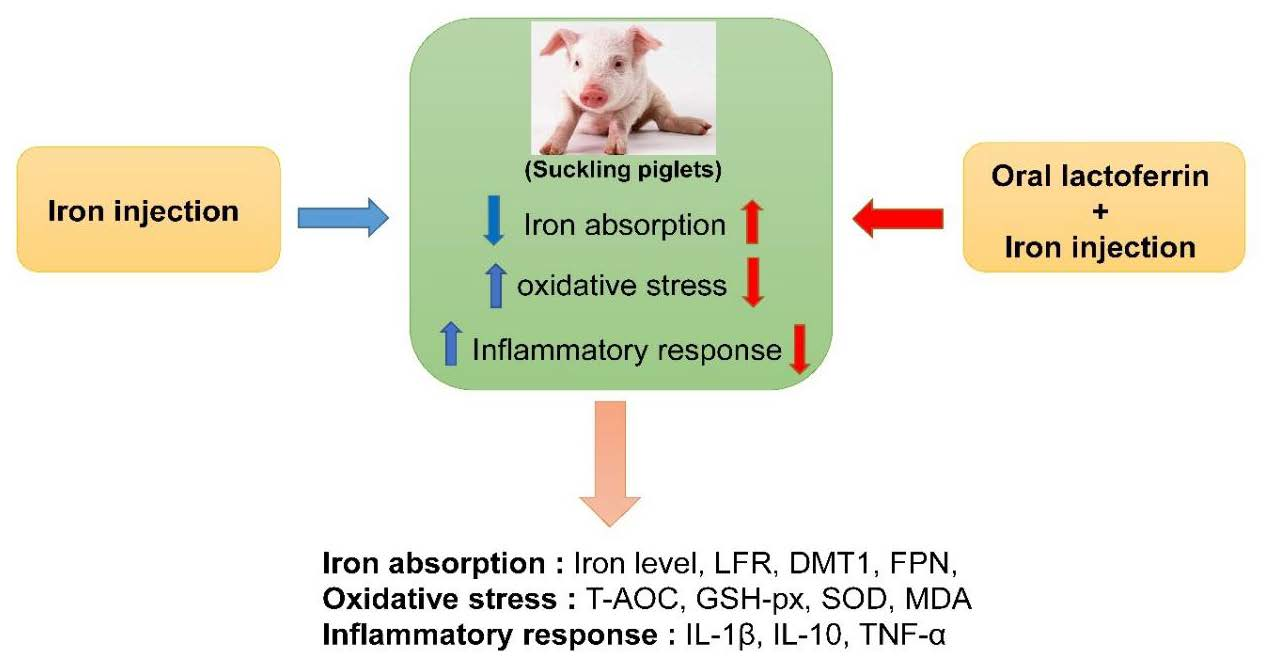
1. Introduction
2. Materials and Methods
2.1. Animals and Samples
2.2. Iron Parameters Analysis
2.3. Gene Expression Analysis
2.4. Protein Extraction and Western Blot Analysis
2.5. Antioxidant Status Analysis
2.6. Serum Cytokines Analyses
2.7. Statistical Analysis
3. Results
3.1. Effects of Lactoferrin on Serum Iron Parameters of Suckling Piglets
3.2. Effects of Lactoferrin on Iron Levels in the Tissues and Digesta of Suckling Piglets
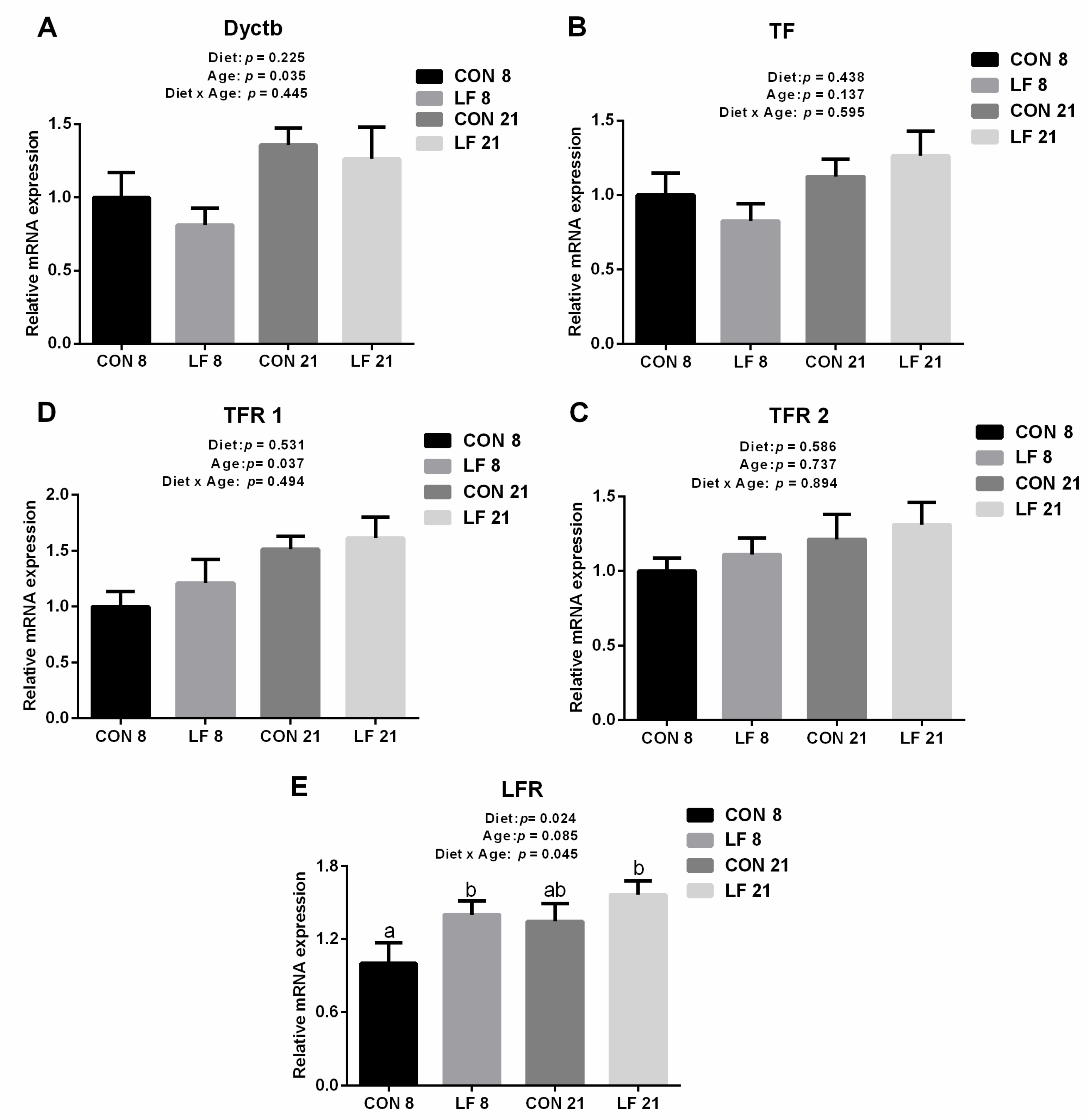
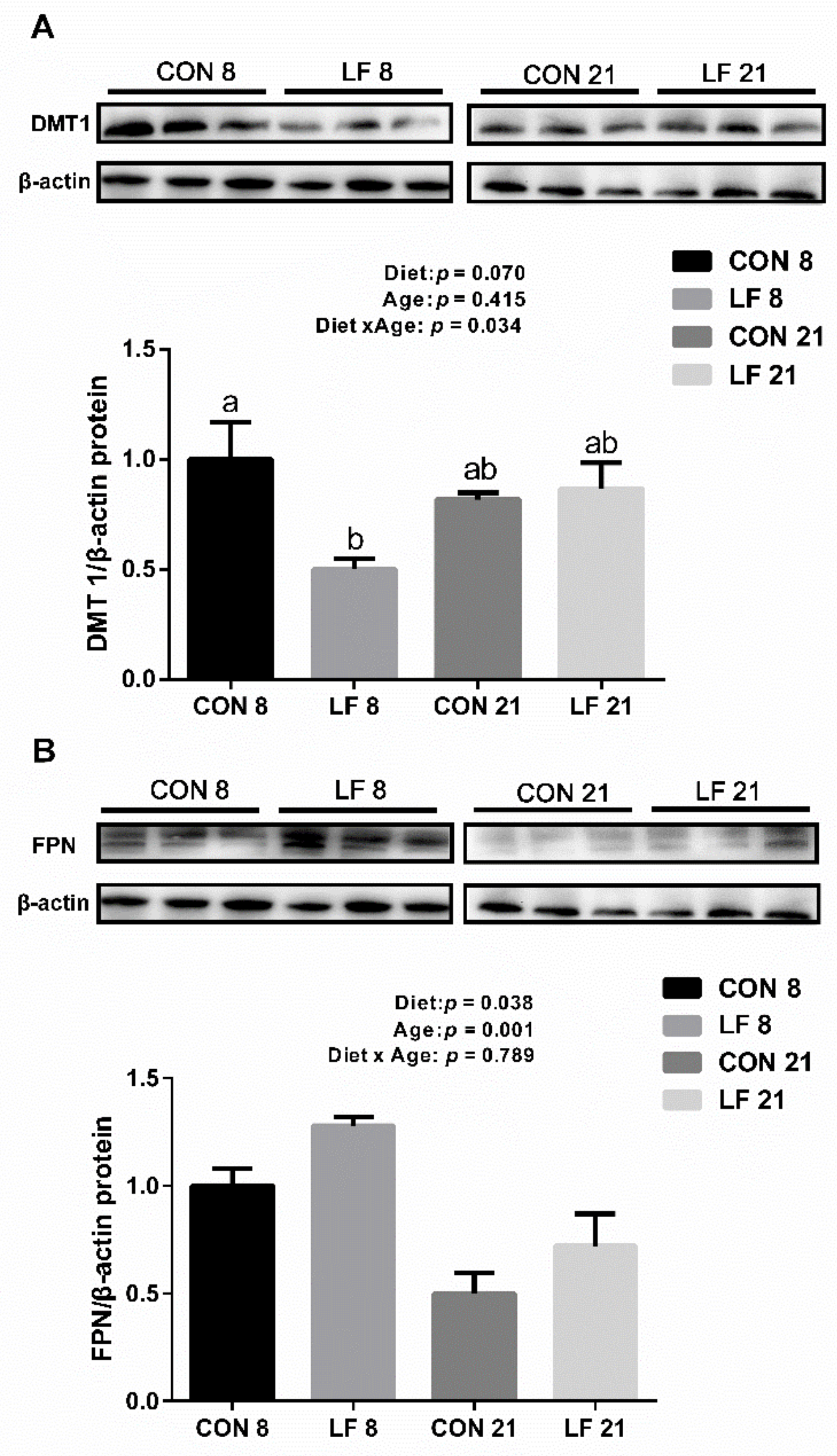
3.3. Effects of Lactoferrin on the Expressions of Gene and Protein Related to Iron Metabolism in the Duodenum of Suckling Piglets
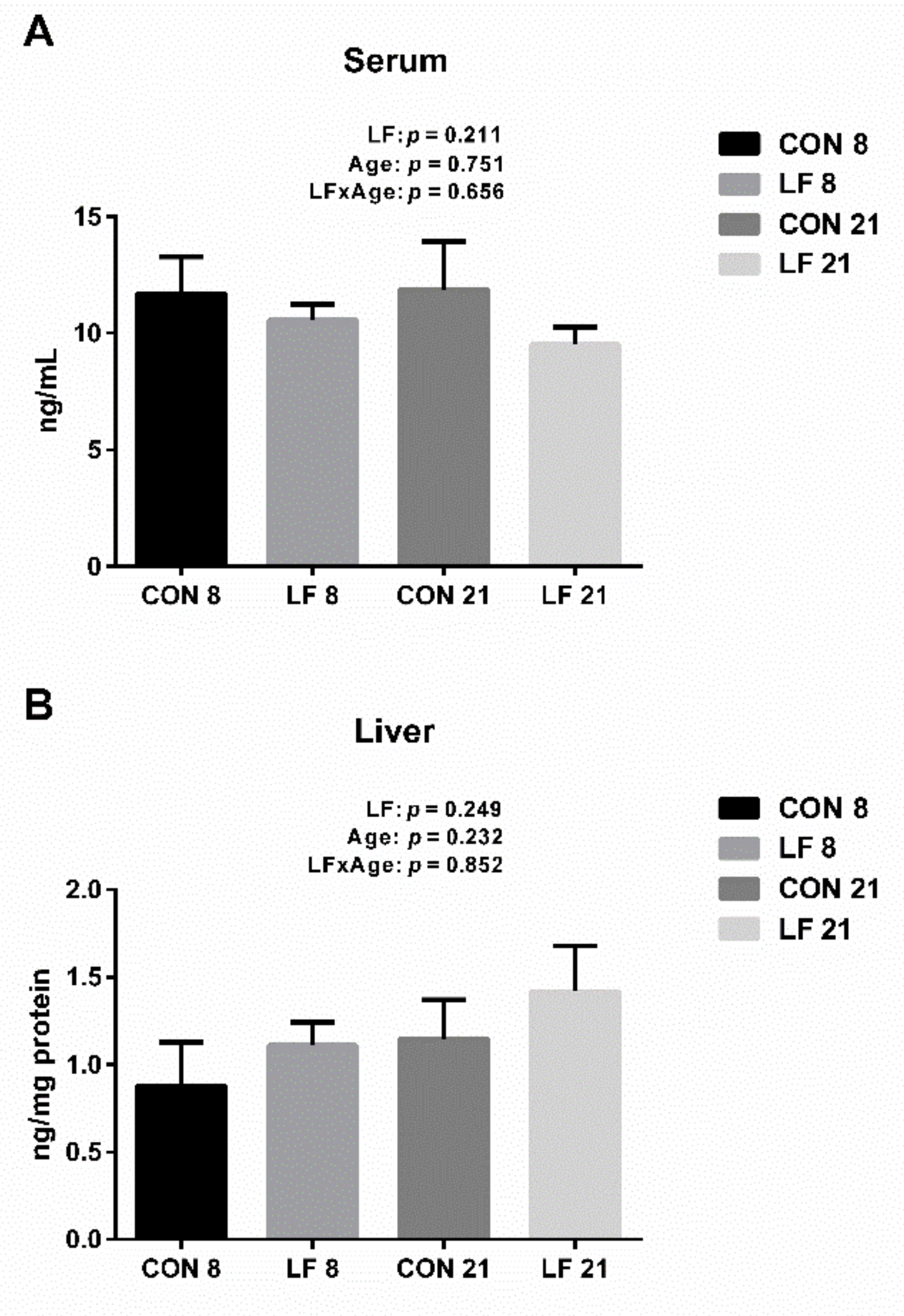
3.4. Effects of Lactoferrin on Hepcidin Levels in the Serum and Liver of Suckling Piglets
3.5. Effects of Lactoferrin on Antioxidant Ability in the Serum and Duodenum of Suckling Piglets
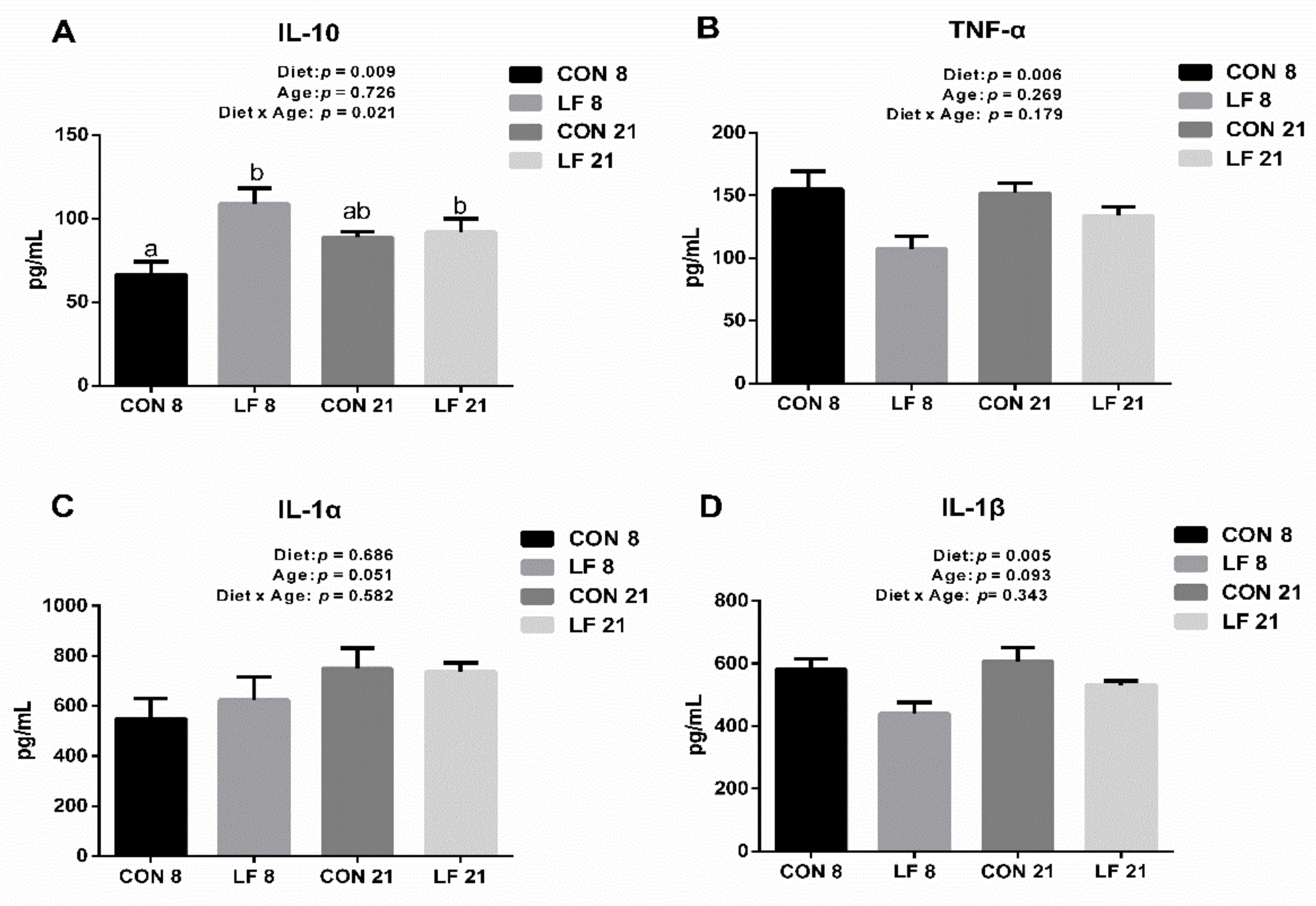
3.6. Effects of Lactoferrin on Cytokines Levels in the Serum of Suckling Piglets
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fajt, Z.; Baňoch, T.; Vašek, J.; Vaňhara, J.; Blahová, J.; Kazatelová, Z.; Fiorino, E.; Svoboda, M. Influence of iron deficiency on thyroidal hormone production in piglets before weaning. Acta Vet. Brno 2016, 85, 17–22. [Google Scholar] [CrossRef]
- Touati, D. Iron and oxidative stress in bacteria. Arch. Biochem. Biophys. 2000, 373, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lia, M.; Yin, Q.; Dang, X.; Chang, J.; Zuo, R.; Zheng, Q. Effect of different iron loads on serum and tissue biochemical parameters and liver hepcidin mRNA abundance of neonatal piglets. Arch. Anim. Nutr. 2011, 65, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Perri, A.M.; Friendship, R.M.; Harding, J.C.; O’Sullivan, T.L. An investigation of iron deficiency and anemia in piglets and the effect of iron status at weaning on post-weaning performance. J. Swine Health Prod. 2016, 24, 10–20. [Google Scholar]
- Lipiński, P.; Starzyński, R.R.; Canonne-Hergaux, F.; Tudek, B.; Oliński, R.; Kowalczyk, P.; Dziaman, T.; Thibaudeau, O.; Gralak, M.A.; Smuda, E. Benefits and Risks of Iron Supplementation in Anemic Neonatal Pigs. Am. J. Pathol. 2010, 177, 1233–1243. [Google Scholar] [CrossRef]
- Pu, Y.; Li, S.; Xiong, H.; Zhang, X.; Wang, Y.; Du, H. Iron promotes intestinal development in neonatal piglets. Nutrients 2018, 10, 726. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.E.; Wessling-Resnick, M. Iron metabolism and the innate immune response to infection. Microbes Infect. 2012, 14, 207–216. [Google Scholar] [CrossRef]
- Egeli, A.K.; Framstad, T. Evaluation of the efficacy of perorally administered glutamic acid-chelated iron and iron-dextran injected subcutaneously in Duroc and Norwegian Landrace piglets. J. Vet. Med. Ser. A 2010, 45, 53–61. [Google Scholar] [CrossRef]
- Staroń, R.; Van Swelm, R.P.; Lipiński, P.; Gajowiak, A.; Lenartowicz, M.; Bednarz, A.; Gajewska, M.; Pieszka, M.; Laarakkers, C.M.; Swinkels, D.W. Urinary Hepcidin Levels in Iron-Deficient and Iron-Supplemented Piglets Correlate with Hepcidin Hepatic mRNA and Serum Levels and with Body Iron Status. PLoS ONE 2015, 10, e0136695. [Google Scholar] [CrossRef]
- Mayeur, S.; Spahis, S.; Pouliot, Y.; Levy, E. Lactoferrin, a pleiotropic protein in health and disease. Antioxid. Redox Sign. 2016, 24, 813–836. [Google Scholar] [CrossRef]
- Rosalba, P.; Francesco, T.; Francesca, B.; Enrica, P.; Valeria, E.; Massimo, M.; Piera, V. Oral administration of lactoferrin increases hemoglobin and total serum iron in pregnant women. Biochem. Cell Biol. 2006, 84, 377–380. [Google Scholar]
- Jungmi, K.; Hyun-Dong, P.; Yoh-Chang, Y.; Eunju, P. Whey protein inhibits iron overload-induced oxidative stress in rats. J. Nutr. Sci. Vitaminol. 2013, 59, 198–205. [Google Scholar]
- Pulina, M.; Sokolov, A.; Zakharova, E.; Kostevich, V.; Vasilyev, V. Effect of lactoferrin on consequences of acute experimental hemorrhagic anemia in rats. Bull. Exp. Biol. Med. 2010, 149, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Paesano, R.; Berlutti, F.; Pietropaoli, M.; Goolsbee, W.; Pacifici, E.; Valenti, P. Lactoferrin efficacy versus ferrous sulfate in curing iron disorders in pregnant and non-pregnant women. Int. J. Immunopathol. Pharmacol. 2010, 23, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B. Nutritional roles of lactoferrin. Curr. Opin. Clin. Nutr. 2009, 12, 293–297. [Google Scholar] [CrossRef]
- Burrow, H.; Kanwar, R.K.; Mahidhara, G.; Kanwar, J.R. Effect of selenium-saturated bovine lactoferrin (Se-bLF) on antioxidant enzyme activities in human gut epithelial cells under oxidative stress. Anticancer Agents Med. Chem. 2011, 11, 762–771. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Yang, H.; Lv, B.; Zhao, D.; Wang, J.; Zhu, W. Dynamic changes of fatty acids and minerals in sow milk during lactation. J. Anim. Physiol. Anim. Nutr. 2019, 103, 603–611. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wang, J.; Tian, S.; Yu, H.; Wang, J.; Zhu, W. Response of Colonic Mucosa-Associated Microbiota Composition, Mucosal Immune Homeostasis, and Barrier Function to Early Life Galactooligosaccharides Intervention in Suckling Piglets. J. Agric. Food Chem. 2018, 67, 578–588. [Google Scholar] [CrossRef]
- Almond, G.; Byers, E.; Seate, J.; Boyer, P. Supplemental iron dextran injections: Influence on hemoglobin concentrations and piglet growth. J. Swine Health Prod. 2017, 25, 308–312. [Google Scholar]
- Pluske, J.; Williams, I.; Zak, L.; Clowes, E.; Cegielski, A.; Aherne, F. Feeding lactating primiparous sows to establish three divergent metabolic states: III. Milk production and pig growth. J. Anim. Sci. 1998, 76, 1165–1171. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Wu, S.; Wang, S. Serum and milk lactoferrin concentration and the correlation with some blood components in lactating sows. Res. Vet. Sci. 2000, 69, 95–97. [Google Scholar] [CrossRef]
- Fernández-Menéndez, S.; Fernández-Sánchez, M.L.; González-Iglesias, H.; Fernández-Colomer, B.; López-Sastre, J.; Sanz-Medel, A. Iron bioavailability from supplemented formula milk: Effect of lactoferrin addition. Eur. J. Nutr. 2016, 56, 2611–2620. [Google Scholar] [CrossRef]
- Martini, L.A.; Laurie, T.; Wood, R.J. Iron treatment downregulates DMT1 and IREG1 mRNA expression in Caco-2 cells. J. Nutr. 2002, 132, 693. [Google Scholar] [CrossRef]
- Igor, T.; Elmar, A.; Milan, T.; Manfred, N.; Markus, S.; Andrea, S.; Thomas, S.; Lukas, E.; Witcher, D.R.; Murphy, A.T. Regulation of iron homeostasis in anemia of chronic disease and iron deficiency anemia: Diagnostic and therapeutic implications. Blood 2009, 113, 5277–5286. [Google Scholar]
- Rolfs, A.; Bonkovsky, H.L.; Kohlroser, J.G.; Mcneal, K.; Sharma, A.; Berger, U.V.; Hediger, M.A. Intestinal expression of genes involved in iron absorption in humans. Am. J. Physiol. Gastrointest Liver Physiol. 2002, 282, G598–G607. [Google Scholar] [CrossRef]
- Ward, D.M.; Kaplan, J. Ferroportin-mediated iron transport: Expression and regulation. Biochim. Biophys. Acta 2012, 1823, 1426–1433. [Google Scholar] [CrossRef] [PubMed]
- Ivana, D.D.; Diane Mcvey, W.; Charles, L.; Vaughn, M.B.; Elizabeta, N.; Sundquist, W.I.; Tomas, G.; Giovanni, M.; Jerry, K. The molecular mechanism of hepcidin-mediated ferroportin down-regulation. Mol. Biol. Cell. 2007, 18, 2569–2578. [Google Scholar]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Lopez, V.; Shafizadeh, T.B.; Halsted, C.H.; Bo, L. Cloning of a pig homologue of the human lactoferrin receptor: Expression and localization during intestinal maturation in piglets. Comp. Biochem. Phys. A Mol. Integr. Physiol. 2007, 148, 584–590. [Google Scholar] [CrossRef]
- Brock, J.H. Lactoferrin–50 years on. Biochem. Cell Biol. 2012, 90, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Jiang, F.; Zhang, J.; Xu, Q.; Chen, D.; Yu, B.; Mao, X.; Yu, J.; Luo, Y.; He, J. Amniotic fluid metabolomics and biochemistry analysis provides novel insights into the diet-regulated foetal growth in a pig model. Sci. Rep. 2017, 7, 44782. [Google Scholar] [CrossRef] [PubMed]
- Hao, R.; Li, Q.; Zhao, J.; Li, H.; Wang, W.; Gao, J. Effects of grape seed procyanidins on growth performance, immune function and antioxidant capacity in weaned piglets. Livest. Sci. 2015, 178, 237–242. [Google Scholar] [CrossRef]
- Xu, J.; Xu, C.; Chen, X.; Cai, X.; Yang, S.; Sheng, Y.; Wang, T. Regulation of an antioxidant blend on intestinal redox status and major microbiota in early weaned piglets. Nutrition 2014, 30, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, C.; An, Z.; Liu, J.; Feng, J. Effect of dietary bovine lactoferrin on performance and antioxidant status of piglets. Anim. Feed Sci. Tech. 2008, 140, 326–336. [Google Scholar] [CrossRef]
- Brambilla, D.; Mancuso, C.; Scuderi, M.R.; Bosco, P.; Cantarella, G.; Lempereur, L.; Di Benedetto, G.; Pezzino, S.; Bernardini, R. The role of antioxidant supplement in immune system, neoplastic, and neurodegenerative disorders: A point of view for an assessment of the risk/benefit profile. Nutr. J. 2008, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, M. Effects of antioxidants on immune system ageing. Eur. J. Clin. Nutr. 2002, 56, S5–S8. [Google Scholar] [CrossRef]
- Yuan, W.; Jin, H.; Ren, Z.; Deng, J.; Zuo, Z.; Wang, Y.; Deng, H.; Deng, Y. Effects of antibacterial peptide on humoral immunity in weaned piglets. Food Agric. Immunol. 2015, 26, 682–689. [Google Scholar] [CrossRef]
- Donovan, S.M. The role of lactoferrin in gastrointestinal and immune development and function: A preclinical perspective. J. Pediatr. 2016, 173, S16–S28. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) | Accession Number |
|---|---|---|
| Dcytb | Forward: TCCACGCAGGGTTGAATAC | NM_001128452·1 |
| Reverse: GCCCAAGGAAGCAGAAAGAC | ||
| TF | Forward: GTATCCGCAGAAAACACCG | NM_001244653 |
| Reverse: AGGACAGGCACCAGACCAC | ||
| TFR 1 | Forward: CAGTTGAACAGAATGGCACG | NM_214001 |
| Reverse: CAGACTCAGACCCATCTCCCT | ||
| TFR 2 | Forward: CAGACTCAGACCCATCTCCCT | XM_003124374 |
| Reverse: CCTCCCAGTCTCCCCTTAT | ||
| LFR | Forward: GTGGGYGATCGCTGGTCCA | NM_017625 |
| Reverse: CCTCCTCCACCRATGCAGTG | ||
| β-actin | Forward: ATGCTTCTAGACGGACTGCG | XM_003357928.4 |
| Reverse: GTTTCAGGAGGCTGGCATGA |
| Items | Day 8 | Day 21 | SEM 2 | p-Values | ||||
|---|---|---|---|---|---|---|---|---|
| CON | LF | CON | LF | Diet | Age | Diet × Age | ||
| Hemoglobin, g/L | 91.01 a | 105.12 b | 71.65 c | 77.22 c | 6.89 | 0.041 | <0.001 | 0.045 |
| Serum Iron, μmol/L | 31.49 | 42.35 | 28.44 | 33.54 | 2.68 | 0.075 | 0.169 | 0.124 |
| TIBC 3, μmol/L | 134.98 | 138.96 | 123.76 | 139.27 | 12.35 | 0.551 | 0.451 | 0.651 |
| Items | Day 8 | Day 21 | SEM 2 | p-Values | ||||
|---|---|---|---|---|---|---|---|---|
| CON | LF | CON | LF | Diet | Age | Diet × Age | ||
| Liver, mg/kg | 253.94 | 269.68 | 173.99 | 214.45 | 36.51 | 0.039 | 0.298 | 0.215 |
| Spleen, mg/kg | 318.72 | 299.68 | 149.47 | 131.42 | 52.03 | 0.732 | 0.005 | 0.732 |
| Kindey, mg/kg | 40.75 | 65.22 | 35.87 | 33.66 | 2.68 | 0.215 | <0.001 | 0.175 |
| Muscle, mg/kg | 13.59 | 14.21 | 8.56 | 9.30 | 1.68 | 0.418 | <0.001 | 0.433 |
| Dudenum, mg/kg | 2.84 | 4.47 | 1.38 | 1.41 | 0.61 | 0.188 | 0.001 | 0.201 |
| Jejunum, mg/kg | 2.25 | 1.64 | 1.15 | 0.90 | 0.32 | 0.197 | 0.010 | 0.577 |
| Ileum, mg/kg | 2.51 | 2.04 | 0.88 | 1.81 | 0.45 | 0.137 | 0.052 | 0.615 |
| Digesta 3, mg/kg | 1.06 | 0.65 | 1.83 | 1.16 | 0.11 | <0.001 | <0.001 | 0.217 |
| Items 2 | Day 8 | Day 21 | SEM 3 | p-Values | ||||
|---|---|---|---|---|---|---|---|---|
| CON | LF | CON | LF | Diet | Age | Diet × Age | ||
| SOD, U/mL | 75.85 | 79.82 | 72.26 | 80.25 | 10.55 | 0.711 | 0.919 | 0.903 |
| T-AOC, U/mL | 11.07 a | 9.03 a | 3.01 b | 13.05 a | 1.75 | 0.032 | 0.253 | 0.003 |
| MDA, U/mL | 1.95 | 2.64 | 4.13 | 2.84 | 0.84 | 0.737 | 0.167 | 0.260 |
| GSH-px, U/mL | 467.69 | 753.84 | 823.07 | 1012.82 | 103.09 | 0.048 | 0.037 | 0.709 |
| Items 2 | Day 8 | Day 21 | SEM 3 | p-Values | ||||
|---|---|---|---|---|---|---|---|---|
| CON | LF | CON | LF | Diet | Age | Diet × Age | ||
| SOD, U/mg protein | 39.53 a,b | 36.73 a,b | 33.59 a | 47.53 b | 5.09 | 0.136 | 0.508 | 0.030 |
| T-AOC, U/mg protein | 0.71 a | 0.54 a,b | 0.30 b | 0.51 a,b | 0.11 | 0.784 | 0.020 | 0.039 |
| MDA, U/mg protein | 0.98 | 0.84 | 0.52 | 0.37 | 0.09 | 0.140 | <0.001 | 0.950 |
| GSH-px, U/mg protein | 76.13 | 79.09 | 76.07 | 127.98 | 12.96 | 0.038 | 0.088 | 0.087 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, P.; Zhao, D.; Zhao, F.; Wang, J.; Zhu, W. The Effects of the Combination of Oral Lactoferrin and Iron Injection on Iron Homestasis, Antioxidative Abilities and Cytokines Activities of Suckling Piglets. Animals 2019, 9, 438. https://doi.org/10.3390/ani9070438
Hu P, Zhao D, Zhao F, Wang J, Zhu W. The Effects of the Combination of Oral Lactoferrin and Iron Injection on Iron Homestasis, Antioxidative Abilities and Cytokines Activities of Suckling Piglets. Animals. 2019; 9(7):438. https://doi.org/10.3390/ani9070438
Chicago/Turabian StyleHu, Ping, Daoyuan Zhao, Fangzhou Zhao, Jing Wang, and Weiyun Zhu. 2019. "The Effects of the Combination of Oral Lactoferrin and Iron Injection on Iron Homestasis, Antioxidative Abilities and Cytokines Activities of Suckling Piglets" Animals 9, no. 7: 438. https://doi.org/10.3390/ani9070438
APA StyleHu, P., Zhao, D., Zhao, F., Wang, J., & Zhu, W. (2019). The Effects of the Combination of Oral Lactoferrin and Iron Injection on Iron Homestasis, Antioxidative Abilities and Cytokines Activities of Suckling Piglets. Animals, 9(7), 438. https://doi.org/10.3390/ani9070438