Menaquinone-4 Amplified Glucose-Stimulated Insulin Secretion in Isolated Mouse Pancreatic Islets and INS-1 Rat Insulinoma Cells

 ,
,  ,
,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
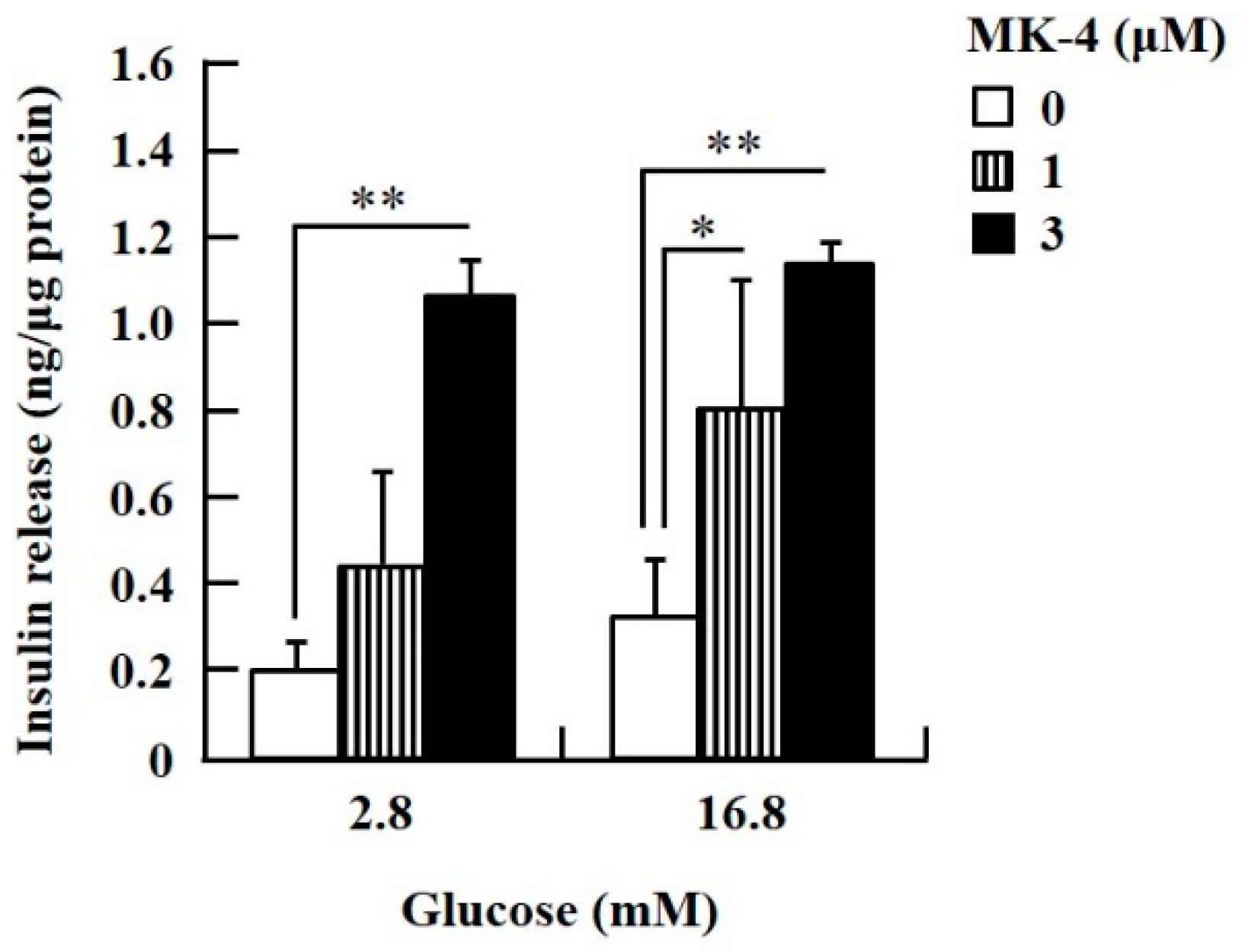
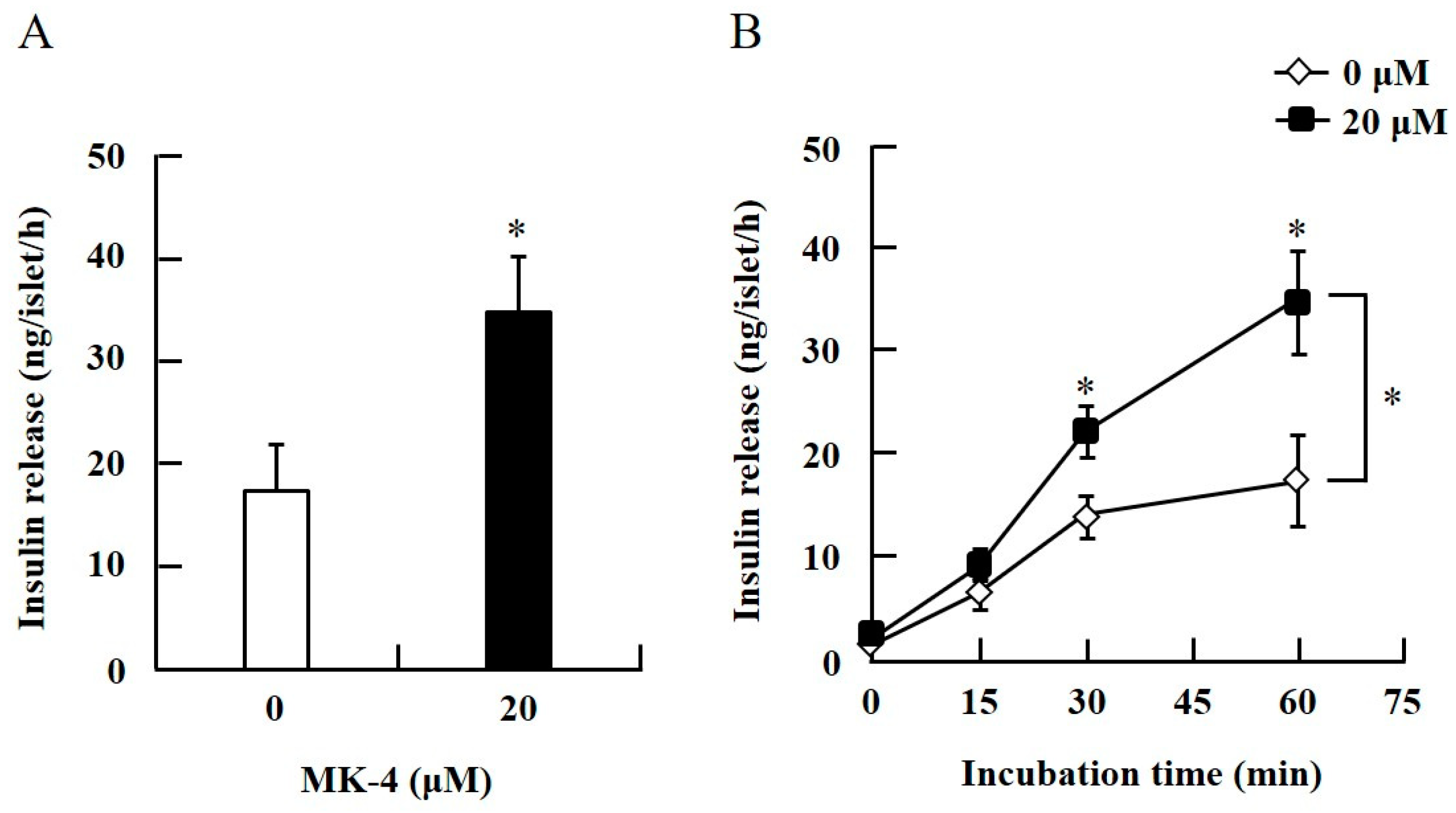
2.1. MK-4 Enhanced GSIS in Isolated Mouse Pancreatic Islets and Rat INS-1 Cells
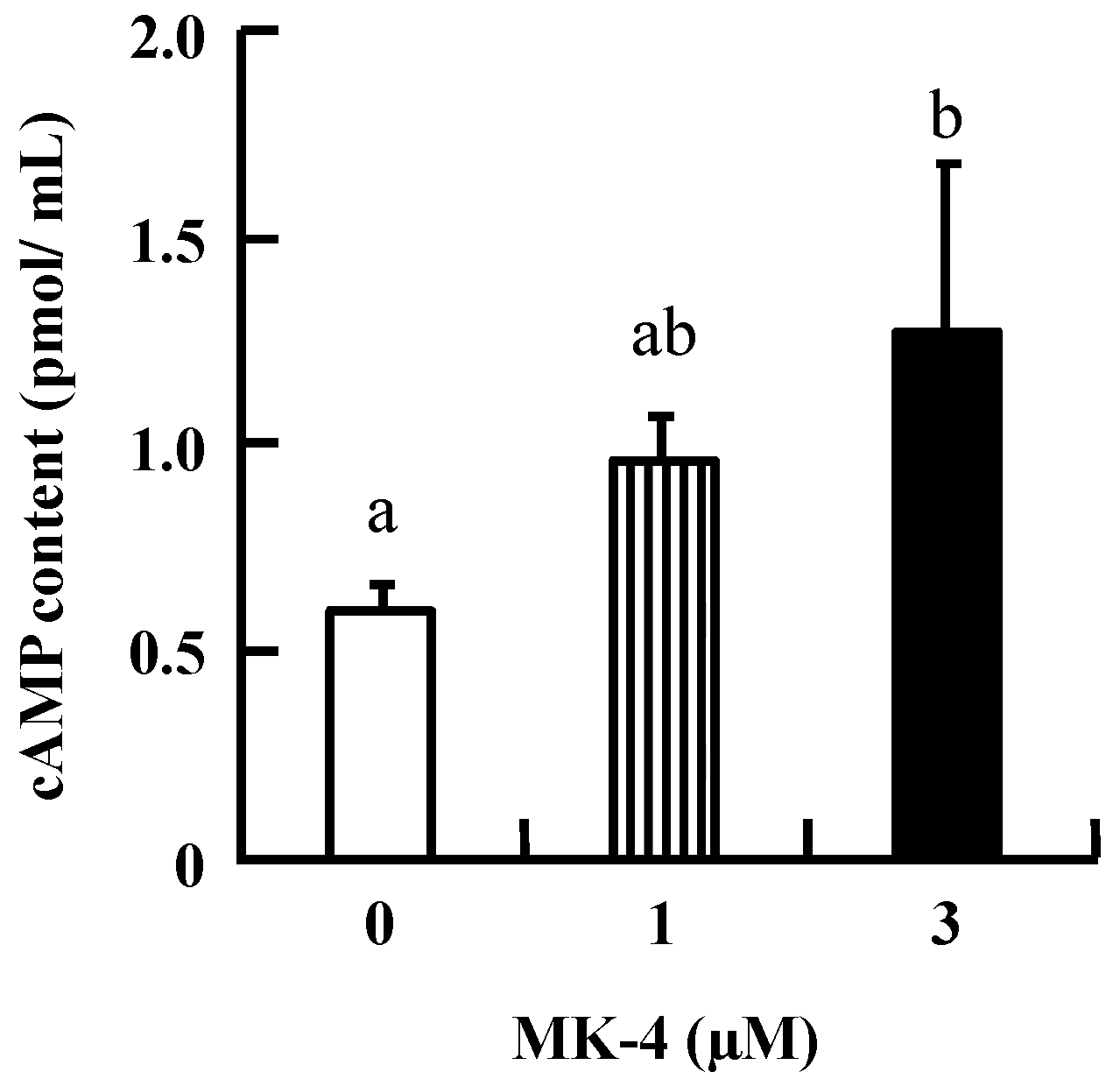
2.2. MK-4 Increased cAMP Levels in INS-1 Cells
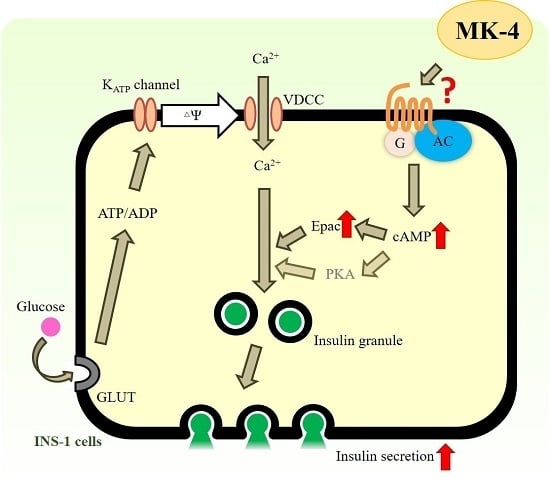
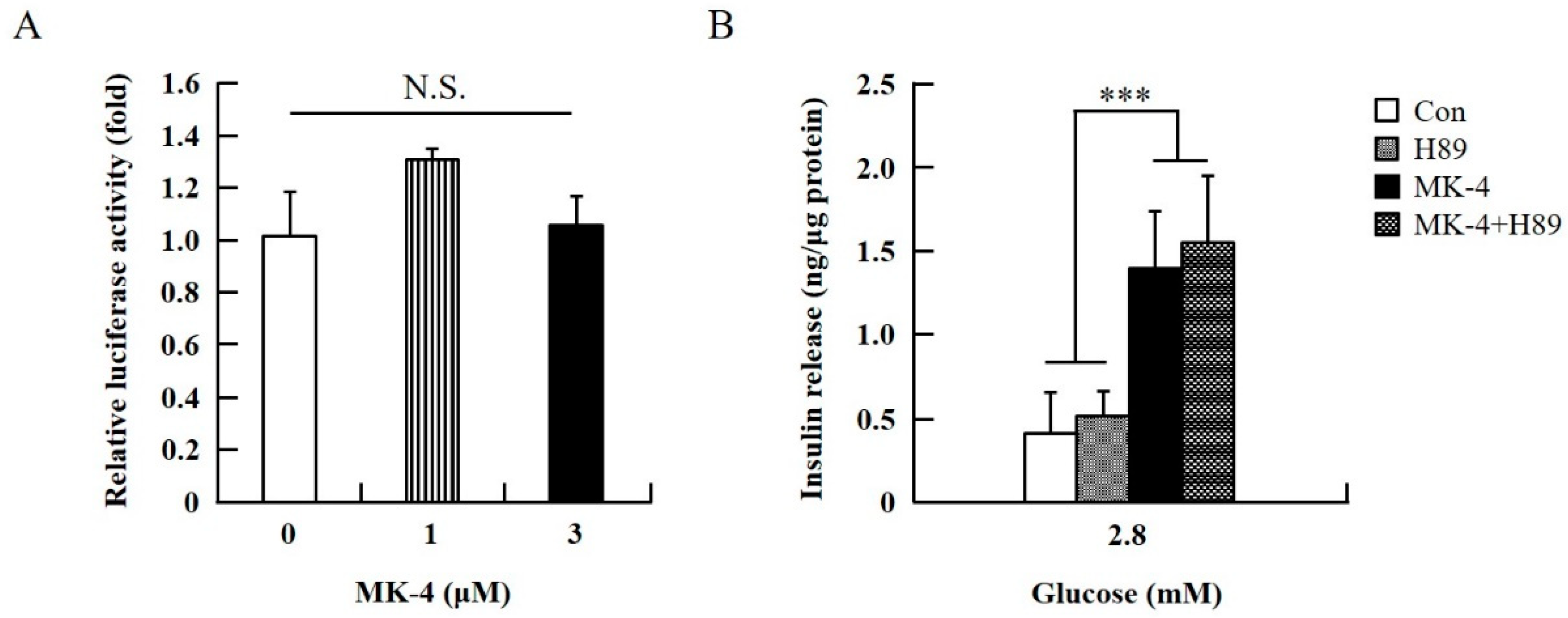
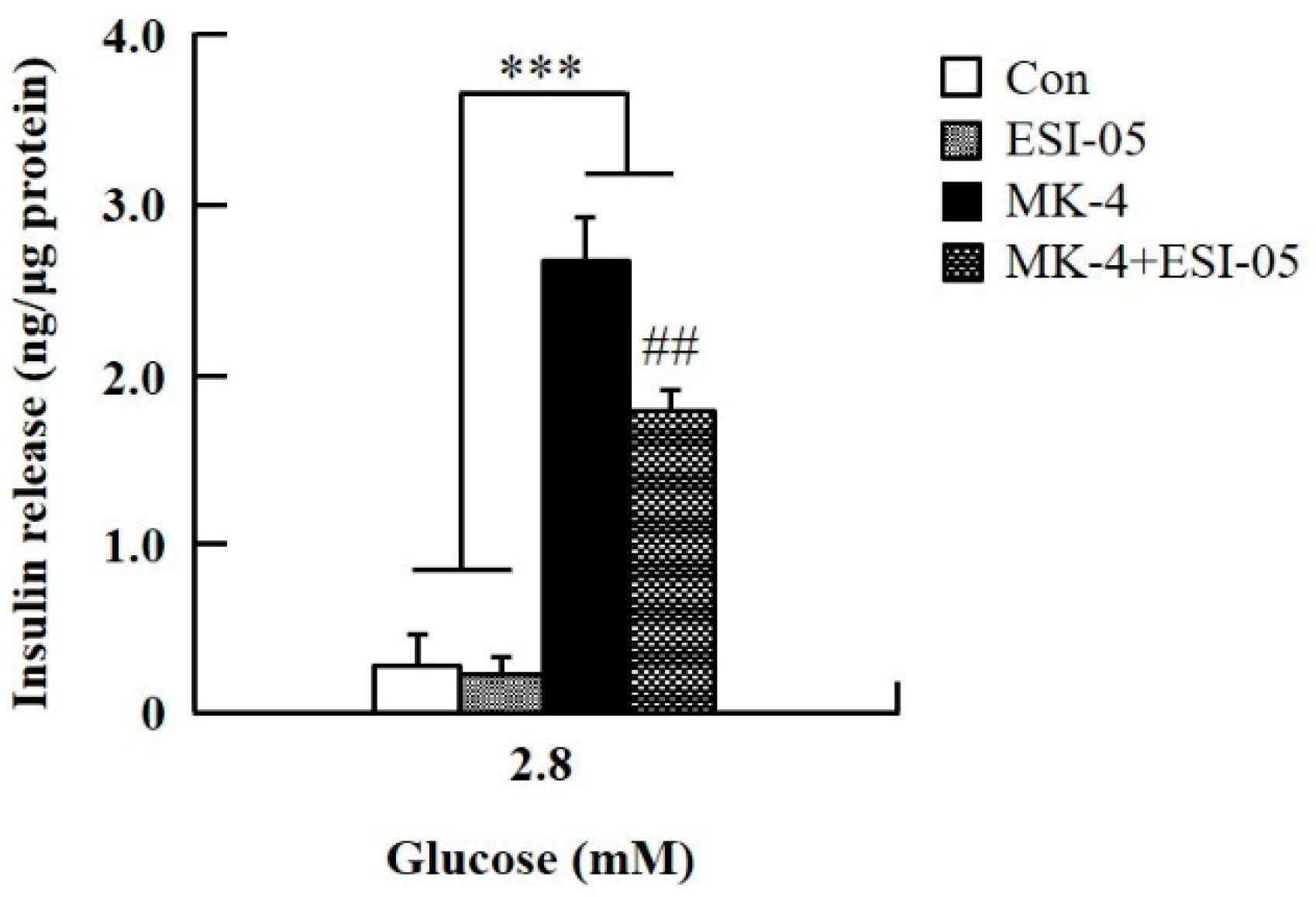
2.3. MK-4 Amplified GSIS by Regulating the cAMP/Epac-Dependent Pathway but not the cAMP/PKA-Dependent Pathway in INS-1 Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animals and Isolation of Pancreatic Islets
4.3. Glucose-Stimulated Insulin Secretion in Isolated Islets
4.4. Cell line and Culture Conditions
4.5. Cell Proliferation Assay
4.6. GSIS in INS-1 Cells
4.7. Measurement of cAMP Levels
4.8. CRE-Reporter Gene Assay
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MK-4 | menaquinone-4 |
| GSIS | glucose-stimulated insulin secretion |
| GIP | glucose-dependent insulinotropic peptide |
| GLP-1 | glucagon-like peptide-1 |
| T2DM | type 2 diabetes mellitus |
| CRE | cAMP response element |
| DDP-4 | dipeptidyl peptidase-4 |
| PKA | protein kinase A |
| Epac2 | exchange protein directly activated by cAMP 2 |
References
- Fu, X.; Harshman, S.G.; Shen, X.; Haytowitz, D.B.; Karl, J.P.; Wolfe, B.E.; Booth, S.L. Multiple Vitamin K Forms Exist in Dairy Foods. Curr. Dev. Nutr. 2017, 1, e000638. [Google Scholar] [CrossRef]
- Otsuka, M.; Kato, N.; Shao, R.X.; Hoshida, Y.; Ijichi, H.; Koike, Y.; Taniguchi, H.; Moriyama, M.; Shiratori, Y.; Kawabe, T.; Omata, M. Vitamin K2 inhibits the growth and invasiveness of hepatocellular carcinoma cells via protein kinase A activation. Hepatology 2004, 40, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, T.; Miyazawa, K.; Yoshida, T.; Ohyashiki, K. Combination of vitamin K2 plus imatinib mesylate enhances induction of apoptosis in small cell lung cancer cell lines. Int. J. Oncol. 2005, 26, 33–40. [Google Scholar] [CrossRef]
- Tokita, H.; Tsuchida, A.; Miyazawa, K.; Ohyashiki, K.; Katayanagi, S.; Sudo, H.; Enomoto, M.; Takagi, Y.; Aoki, T. Vitamin K2-induced antitumor effects via cell-cycle arrest and apoptosis in gastric cancer cell lines. Int. J. Mol. Med. 2006, 17, 235–243. [Google Scholar] [CrossRef]
- Enomoto, M.; Tsuchida, A.; Miyazawa, K.; Yokoyama, T.; Kawakita, H.; Tokita, H.; Naito, M.; Itoh, M.; Ohyashiki, K.; Aoki, T. Vitamin K2-induced cell growth inhibition via autophagy formation in cholangiocellular carcinoma cell lines. Int. J. Mol. Med. 2007, 20, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Tabb, M.M.; Sun, A.; Zhou, C.; Grün, F.; Errandi, J.; Romero, K.; Pham, H.; Inoue, S.; Mallick, S.; Lin, M.; Forman, B.M.; Blumberg, B. Vitamin K2 regulation of bone homeostasis is mediated by the steroid and xenobiotic receptor SXR. J. Biol. Chem. 2003, 278, 43919–43927. [Google Scholar] [CrossRef]
- Ichikawa, T.; Horie-Inoue, K.; Ikeda, K.; Blumberg, B.; Inoue, S. Steroid and xenobiotic receptor SXR mediates vitamin K2-activated transcription of extracellular matrix-related genes and collagen accumulation in osteoblastic cells. J. Biol. Chem. 2006, 281, 16927–16934. [Google Scholar] [CrossRef]
- Sultana, H.; Watanabe, K.; Rana, M.M.; Takashima, R.; Ohashi, A.; Komai, M.; Shirakawa, H. Effects of Vitamin K2 on the Expression of Genes Involved in Bile Acid Synthesis and Glucose Homeostasis in Mice with Humanized PXR. Nutrients 2018, 10, 982. [Google Scholar] [CrossRef] [PubMed]
- Ohsaki, Y.; Shirakawa, H.; Hiwatashi, K.; Furukawa, Y.; Mizutani, T.; Komai, M. Vitamin K suppresses lipopolysaccharide-induced inflammation in the rat. Biosci. Biotechnol. Biochem. 2006, 70, 926–932. [Google Scholar] [CrossRef]
- Ohsaki, Y.; Shirakawa, H.; Miura, A.; Giriwono, P.E.; Sato, S.; Ohashi, A.; Iribe, M.; Goto, T.; Komai, M. Vitamin K suppresses the lipopolysaccharide-induced expression of inflammatory cytokines in cultured macrophage-like cells via the inhibition of the activation of nuclear factor κB through the repression of IKKα/β phosphorylation. J. Nutr. Biochem. 2010, 21, 1120–1126. [Google Scholar] [CrossRef]
- Ito, A.; Shirakawa, H.; Takumi, N.; Minegishi, Y.; Ohashi, A.; Howlader, Z.H.; Ohsaki, Y.; Sato, T.; Goto, T.; Komai, M. Menaquinone-4 enhances testosterone production in rats and testis-derived tumor cells. Lipids Health Dis. 2011, 10, 158. [Google Scholar] [CrossRef]
- Orimo, H.; Shiraki, M.; Tomita, A.; Morii, H.; Fujita, T.; Ohata, M. Effects of menatetrenone on the bone and calcium metabolism in osteoporosis: A double-blind placebo-controlled study. J. Bone Miner. Metab. 1998, 16, 106–112. [Google Scholar] [CrossRef]
- Shiraki, M.; Shiraki, Y.; Aoki, C.; Miura, M. Vitamin K2 (menatetrenone) effectively prevents fractures and sustains lumbar bone mineral density in osteoporosis. J. Bone Miner. Res. 2000, 15, 515–521. [Google Scholar] [CrossRef]
- Knapen, M.H.; Schurgers, L.J.; Vermeer, C. Vitamin K2 supplementation improves hip bone geometry and bone strength indices in postmenopausal women. Osteoporos. Int. 2007, 18, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Fujita, T.; Kishimoto, H.; Makino, T.; Nakamura, T.; Nakamura, T.; Sato, T.; Yamazaki, K. Randomized controlled study on the prevention of osteoporotic fractures (OF study): A phase IV clinical study of 15-mg menatetrenone capsules. J. Bone Miner. Metab. 2009, 27, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, H.; Katsurai, T.; Komai, M. Conversion of menaquinone-4 in animal organs and it functions. Jpn. Oil Chem. Soc. 2014, 14, 547–553. [Google Scholar]
- Henquin, J.C. Triggering and amplifying pathways of regulation of insulin secretion by glucose. Diabetes 2000, 49, 1751–1760. [Google Scholar] [CrossRef] [PubMed]
- Lipshutz, A.K.; Gropper, M.A. Perioperative glycemic control: An evidence-based review. Anesthesiology 2009, 110, 408–421. [Google Scholar] [CrossRef]
- Meier, J.J.; Nauck, M.A.; Schmidt, W.E.; Gallwitz, B. Gastric inhibitory polypeptide: The neglected incretin revisited. Regul. Pept. 2002, 107, 1–13. [Google Scholar] [CrossRef]
- Vilsbøll, T.; Krarup, T.; Madsbad, S.; Holst, J.J. Both GLP-1 and GIP are insulinotropic at basal and postprandial glucose levels and contribute nearly equally to the incretin effect of a meal in healthy subjects. Regul. Pept. 2003, 114, 115–121. [Google Scholar] [CrossRef]
- Drucker, D.J. The biology of incretin hormones. Cell Metab. 2006, 3, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, R.; Matsuyama, T.; Namba, M.; Watanabe, N.; Itoh, H.; Kono, N.; Tarui, S. Glucagonostatic and insulinotropic action of glucagonlike peptide I-(7–36)-amide. Diabetes 1989, 38, 902–905. [Google Scholar] [CrossRef]
- Prigeon, R.L.; Quddusi, S.; Paty, B.; D’Alessio, D.A. Suppression of glucose production by GLP-1 independent of islet hormones: A novel extrapancreatic effect. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E701–E707. [Google Scholar] [CrossRef]
- Odegaard, J.I.; Chawla, A. Pleiotropic actions of insulin resistance and inflammation in metabolic homeostasis. Science 2013, 339, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Holst, J.J.; Knop, F.K.; Vilsbøll, T.; Krarup, T.; Madsbad, S. Loss of incretin effect is a specific; important, and early characteristic of type 2 diabetes. Diabetes Care 2011, 34, S251–S257. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M.A.; Baller, B.; Meier, J.J. Gastric inhibitory polypeptide and glucagon-like peptide-1 in the pathogenesis of type 2 diabetes. Diabetes 2004, 53, S190–S196. [Google Scholar] [CrossRef] [PubMed]
- Cryer, P.E. Glycemic goals in diabetes: Trade-off between glycemic control and iatrogenic hypoglycemia. Diabetes 2014, 63, 2188–2195. [Google Scholar] [CrossRef] [PubMed]
- Beulens, J.W.; van der A, D.L.; Grobbee, D.E.; Sluijs, I.; Spijkerman, A.M.; van der Schouw, Y.T. Dietary phylloquinone and menaquinones intakes and risk of type 2 diabetes. Diabetes Care 2010, 33, 1699–1705. [Google Scholar] [CrossRef]
- Kumar, R.; Binkley, N.; Vella, A. Effect of phylloquinone supplementation on glucose homeostasis in humans. Am. J. Clin. Nutr. 2010, 92, 1528–1532. [Google Scholar] [CrossRef] [PubMed]
- Ibarrola-Jurado, N.; Salas-Salvadó, J.; Martínez-González, M.A.; Bulló, M. Dietary phylloquinone intake and risk of type 2 diabetes in elderly subjects at high risk of cardiovascular disease. Am. J. Clin. Nutr. 2012, 96, 1113–1118. [Google Scholar] [CrossRef] [PubMed]
- Rasekhi, H.; Karandish, M.; Jalali, M.T.; Mohammad-Shahi, M.; Zarei, M.; Saki, A.; Shahbazian, H. The effect of vitamin K1 supplementation on sensitivity and insulin resistance via osteocalcin in prediabetic women: A double-blind randomized controlled clinical trial. Eur. J. Clin. Nutr. 2015, 69, 891–895. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Jacques, P.F.; Meigs, J.B.; Saltzman, E.; Shea, M.K.; Gundberg, C.; Dawson-Hughes, B.; Dallal, G.; Booth, S.L. Effect of vitamin K supplementation on insulin resistance in older men and women. Diabetes Care 2008, 31, 2092–2096. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Booth, S.L.; Meigs, J.B.; Saltzman, E.; Jacques, P.F. Phylloquinone intake; insulin sensitivity; and glycemic status in men and women. Am. J. Clin. Nutr. 2008, 88, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, N.; Wakabayashi, I.; Sakamoto, K. Low vitamin K intake effects on glucose tolerance in rats. Int. J. Vitam. Nutr. Res. 1999, 69, 27–31. [Google Scholar] [CrossRef]
- Sakamoto, N.; Nishiike, T.; Iguchi, H.; Sakamoto, K. Relationship between acute insulin response and vitamin K intake in healthy young male volunteers. Diabetes Nutr. Metab. 1999, 12, 37–41. [Google Scholar] [PubMed]
- Sone, H.; Ito, M.; Sugiyama, K.; Ohneda, M.; Maebashi, M.; Furukawa, Y. Biotin enhances glucose-stimulated insulin secretion in the isolated perfused pancreas of the rat. J. Nutr. Biochem. 1999, 10, 237–243. [Google Scholar] [CrossRef]
- Knop, F.K.; Vilsbøll, T.; Højberg, P.V.; Larsen, S.; Madsbad, S.; Vølund, A.; Holst, J.J.; Krarup, T. Reduced incretin effect in type 2 diabetes: Cause or consequence of the diabetic state? Diabetes 2007, 56, 1951–1959. [Google Scholar] [CrossRef]
- Nauck, M.; Stöckmann, F.; Ebert, R.; Creutzfeldt, W. Reduced incretin effect in type 2 (non-insulin-dependent) diabetes. Diabetologia 1986, 29, 46–52. [Google Scholar] [CrossRef]
- Ahrén, B. Incretin dysfunction in type 2 diabetes: Clinical impact and future perspectives. Diabetes Metab. 2013, 39, 195–201. [Google Scholar] [CrossRef]
- Herzberg-Schafer, S.; Heni, M.; Stefan, N.; Haring, H.U.; Fritsche, A. Impairment of GLP1-induced insulin secretion: Role of genetic background, insulin resistance and hyperglycaemia. Diabetes Obes. Metab. 2012, 14, 85–90. [Google Scholar] [CrossRef]
- Tasyurek, H.M.; Altunbas, H.A.; Balci, M.K.; Sanlioglu, S. Incretins: Their physiology and application in the treatment of diabetes mellitus. Diabetes Metab. Res. Rev. 2014, 30, 354–371. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J.; Nauck, M.A. The incretin system: Glucagon-like peptide-1 receptor agonists and dipeptidyl peptidase-4 inhibitors in type 2 diabetes. Lancet 2006, 368, 1696–1705. [Google Scholar] [CrossRef]
- Aroda, V.R.; DeYoung, M.B. Clinical implications of exenatide as a twice-daily or once-weekly therapy for type 2 diabetes. Postgrad Med. 2011, 123, 228–238. [Google Scholar] [CrossRef]
- Franks, A.S.; Lee, P.H.; George, C.M. Pancreatitis: A potential complication of liraglutide? Ann. Pharmacother. 2012, 46, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J.; Sherman, S.I.; Bergenstal, R.M.; Buse, J.B. The safety of incretin-based therapies-review of the scientific evidence. J. Clin. Endocrinol. Metab. 2011, 96, 2027–2031. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.H.; Stockton, M.D.; Franks, A.S. Acute pancreatitis associated with liraglutide. Ann. Pharmacother. 2011, 45, e22. [Google Scholar] [CrossRef]
- Anderson, S.L.; Trujillo, J.M. Association of pancreatitis with glucagon-like peptide-1 agonist use. Ann. Pharmacother. 2010, 44, 904–909. [Google Scholar] [CrossRef]
- Nyborg, N.C.; Molck, A.M.; Madsen, L.W.; Knudsen, L.B. The human GLP-1 analog liraglutide and the pancreas: Evidence for the absence of structural pancreatic changes in three species. Diabetes 2012, 61, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Barnett, A. DPP-4 inhibitors and their potential role in the management of type 2 diabetes. Int. J. Clin. Pract. 2006, 60, 1454–1470. [Google Scholar] [CrossRef]
- Engel, S.S.; Williams-Herman, D.E.; Golm, G.T.; Clay, R.J.; Machotka, S.V.; Kaufman, K.D.; Goldstein, B.J. Sitagliptin: Review of preclinical and clinical data regarding incidence of pancreatitis. Int. J. Clin. Pract. 2010, 64, 984–990. [Google Scholar] [CrossRef]
- Pratley, R.E.; Jauffret-Kamel, S.; Galbreath, E.; Holmes, D. Twelve-week monotherapy with the DPP-4 inhibitor vildagliptin improves glycemic control in subjects with type 2 diabetes. Horm. Metab. Res. 2006, 38, 423–428. [Google Scholar] [CrossRef] [PubMed]
- VanDeKoppel, S.; Choe, H.M.; Sweet, B.V. Managed care perspective on three new agents for type 2 diabetes. J. Manag. Care Pharm. 2008, 14, 363–380. [Google Scholar] [CrossRef] [PubMed]
- Ristic, S.; Byiers, S.; Foley, J.; Holmes, D. Improved glycaemic control with dipeptidyl peptidase-4 inhibition in patients with type 2 diabetes: Vildagliptin (LAF237) dose response. Diabetes Obes. Metab. 2005, 7, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Shimano, H.; Yamamoto, T.; Ishikawa, M.; Kumadaki, S.; Matsuzaka, T.; Nakagawa, Y.; Yahagi, N.; Nakakuki, M.; Hasty, A.H.; et al. Palmitate impairs and eicosapentaenoate restores insulin secretion through regulation of SREBP-1c in pancreatic islets. Diabetes 2008, 57, 2382–2392. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, N.; Shibasaki, T.; Kashima, Y.; Miki, T.; Takahashi, K.; Ueno, H.; Sunaga, Y.; Yano, H.; Matsuura, Y.; Iwanaga, T.; Takai, Y.; Seino, S. cAMP-GEFII is a direct target of cAMP in regulated exocytosis. Nat. Cell. Biol. 2000, 2, 805–811. [Google Scholar] [CrossRef]
- Kashima, Y.; Miki, T.; Shibasaki, T.; Ozaki, N.; Miyazaki, M.; Yano, H.; Seino, S. Critical role of cAMP-GEFII--Rim2 complex in incretin-potentiated insulin secretion. J. Biol. Chem. 2001, 276, 46046–46053. [Google Scholar] [CrossRef]
- Holz, G.G. Epac: A new cAMP-binding protein in support of glucagon-like peptide-1 receptor-mediated signal transduction in the pancreatic beta-cell. Diabetes 2004, 53, 5–13. [Google Scholar] [CrossRef]
- Shibasaki, T.; Takahashi, H.; Miki, T.; Sunaga, Y.; Matsumura, K.; Yamanaka, M.; Zhang, C.; Tamamoto, A.; Satoh, T.; Miyazaki, J.; Seino, S. Essential role of Epac2/Rap1 signaling in regulation of insulin granule dynamics by cAMP. Proc. Natl. Acad. Sci. USA 2007, 104, 19333–19338. [Google Scholar] [CrossRef]
- Zhang, C.L.; Katoh, M.; Shibasaki, T.; Minami, K.; Sunaga, Y.; Takahashi, H.; Yokoi, N.; Iwasaki, M.; Miki, T.; Seino, S. The cAMP sensor Epac2 is a direct target of antidiabetic sulfonylurea drugs. Science 2009, 325, 607–610. [Google Scholar] [CrossRef]
- Ho, H.J.; Shirakawa, H.; Komai, M. Menquinone-4 enhances steroidogenesis in testis derived tumor cells via the elevation of cAMP level. In Vitamin K2—Vital for Health and Wellbeing; InTech: Rijeka, Croatia, 2017; pp. 189–198. [Google Scholar]
- Asfari, M.; Janjic, D.; Meda, P.; Li, G.; Halban, P.A.; Wollheim, C.B. Establishment of 2-mercaptoethanol-dependent differentiated insulin-secreting cell lines. Endocrinology 1992, 130, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Mataunaga, T.; Li, S.; Adachi, T.; Joo, E.; Gu, N.; Yamazaki, N.; Yasuda, K.; Kondoh, T.; Tsuda, K. Hyperoxia reverses glucotoxicity-induced inhibition of insulin secretion in rat INS-1 β cells. Biosci. Biotech. Biochem. 2014, 78, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Horiuchi, H.; Harada, N.; Adachi, T.; Nakano, Y.; Inui, H.; Yamaji, R. S-Equol enantioselectively activates cAMP-protein kinase A signaling and reduces alloxan-induced cell death in INS-1 pancreatic β-cells. J. Nutr. Sci. Vitaminol. 2014, 60, 291–296. [Google Scholar] [CrossRef]
- Ho, H.J.; Shirakawa, H.; Yoshida, R.; Ito, A.; Maeda, M.; Goto, T.; Komai, M. Geranylgeraniol enhances testosterone production via the cAMP/protein kinase A pathway in testis-derived I-10 tumor cells. Biosci. Biotechnol. Biochem. 2016, 80, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Suemori, H.; Kadodawa, Y.; Goto, K.; Araki, I.; Kondoh, H.; Nakatsuni, N. A mouse embryonic stem cell line showing pluripotency of differentiation in early embryos and ubiquitous beta-galactosidase expression. Cell Differ. Dev. 1990, 29, 181–186. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, H.-J.; Shirakawa, H.; Hirahara, K.; Sone, H.; Kamiyama, S.; Komai, M. Menaquinone-4 Amplified Glucose-Stimulated Insulin Secretion in Isolated Mouse Pancreatic Islets and INS-1 Rat Insulinoma Cells. Int. J. Mol. Sci. 2019, 20, 1995. https://doi.org/10.3390/ijms20081995
Ho H-J, Shirakawa H, Hirahara K, Sone H, Kamiyama S, Komai M. Menaquinone-4 Amplified Glucose-Stimulated Insulin Secretion in Isolated Mouse Pancreatic Islets and INS-1 Rat Insulinoma Cells. International Journal of Molecular Sciences. 2019; 20(8):1995. https://doi.org/10.3390/ijms20081995
Chicago/Turabian StyleHo, Hsin-Jung, Hitoshi Shirakawa, Keisukei Hirahara, Hideyuki Sone, Shin Kamiyama, and Michio Komai. 2019. "Menaquinone-4 Amplified Glucose-Stimulated Insulin Secretion in Isolated Mouse Pancreatic Islets and INS-1 Rat Insulinoma Cells" International Journal of Molecular Sciences 20, no. 8: 1995. https://doi.org/10.3390/ijms20081995
APA StyleHo, H.-J., Shirakawa, H., Hirahara, K., Sone, H., Kamiyama, S., & Komai, M. (2019). Menaquinone-4 Amplified Glucose-Stimulated Insulin Secretion in Isolated Mouse Pancreatic Islets and INS-1 Rat Insulinoma Cells. International Journal of Molecular Sciences, 20(8), 1995. https://doi.org/10.3390/ijms20081995