Abstract
Intolerance of uncertainty (IU) and behavioral inhibition (BI) are personality traits associated with avoidance of the unfamiliar. Both are linked to anxiety and other disorders. However, most research on personality has relied on self-report, which may not correspond to actual behavior. An alternative is to observe behavior in computer-based tasks designed to assess personality. The current study sought to develop such a task, based on the conditioned place preference paradigm, which is sensitive to IU but not BI. Participants foraged for reward in a virtual environment consisting of multiple interconnected rooms. In the training phase, the rich room was paired with a higher number of wins than losses. The poor room was the opposite. In the test phase, participants could freely search any of the rooms, including a completely new room. Although most showed a strong initial preference for the new room, those with higher self-reported IU left this room faster, foraging there significantly less than those with lower IU. This preference also depended on information provided about the new room. There was a strong positive correlation between IU and BI; however, the latter was unrelated to behavior. Thus, the task captures a unique component of IU.
1. Introduction
Dealing with uncertainty poses a challenge that must be overcome by any organism. However, relatively few studies have examined individual differences in how people respond to uncertain situations (Carleton et al., 2016). The current study focuses on intolerance of uncertainty (IU)—a personality trait associated with aversion and avoidance of uncertainty (Carleton, 2016). Recently, there has been increased interest in IU as a possible common factor between several different types of disorders, including anxiety, depression, and substance use (Carleton et al., 2016). Thus, it is a personality trait with transdiagnostic relevance, that is also related to other traits linked to psychopathology, including behavioral inhibition (BI). BI can be defined as the tendency to avoid or withdraw from novelty in both social and non-social situations (Gladstone & Parker, 2005). It has been identified as a risk factor for generalized anxiety and social anxiety disorders (Clauss & Blackford, 2012; Muris et al., 2011) and is also related to the severity of self-reported post-traumatic stress disorder symptoms (Contractor et al., 2013; Myers et al., 2012).
As with most personality research, that on IU and BI has relied almost exclusively on self-report measures (Carleton et al., 2007; Freeston et al., 1994), which may not correspond to actual behavior. While behavioral tests can be more resource intensive, computer-based tasks that employ virtual environments may provide a cost-effective solution that can supplement self-report. Once developed and validated, these tasks can still be administered quickly to a large number of participants, either in the laboratory or online. For example, a computer-based task to assess BI has been developed (Myers et al., 2016), similar to an adventure game where participants guide an avatar through a series of choices that resemble a real-life situation (e.g., attending a party). A strong correlation (r > 0.78) was found between behavior in the task and scores on a self-report measure of BI, supporting that the task can either supplement traditional paper-and-pencil questionnaires, or replace them in populations (e.g., children) where they are less appropriate (Myers et al., 2016). In another example, the task resembled a spaceship shooter video game where participants had to learn to avoid incoming enemy fire. The avoidance response was faster to condition in participants with higher self-reported BI (Sheynin et al., 2014). Such tasks can help link the human and animal studies on conditioning and capture features of conditioning, such as resistance to extinction (Sheynin et al., 2016), important for clinical practice. They can also be used to study populations with clinical symptoms or diagnoses, such as post-traumatic stress disorder and substance use disorder, that involve persistent avoidance (Sheynin et al., 2016, 2017). In addition, since they resemble video games, these tasks can help mitigate the boredom and fatigue associated with standard conditioning procedures, and with questionnaires.
Prior research has sought to measure IU using a task based on the conditioned place preference (CPP) paradigm commonly used in non-human animals to assess the reward value of different types of stimuli, such as food, water, potential mates, or drugs of abuse (Radell et al., 2016; Tzschentke, 2007). In the human version of the task, participants guide an avatar through a virtual environment while foraging for reward, with some locations paired with a higher payoff than others. As in the animal version, participants are then allowed to freely explore the environment, and their preference for each location (i.e., virtual room) is quantified (Radell et al., 2016). However, this task, as well as the few other studies of CPP in humans (Linhardt et al., 2022) used two-room paradigms, where the choice is between two previously experienced, therefore familiar, locations. Instead, IU should be more important when the choice is between the familiar and the unknown.
The purpose of the current study was to test this possibility by introducing a completely new room that could be explored during the preference test, and by varying its similarity to a previously experienced room. It was hypothesized that participants with higher IU will be less likely to enter and forage in the new room compared to those with lower IU. If this was to avoid uncertainty, then providing information about this room (i.e., a cue signaling that it is similar to a previously experienced room) should reduce this difference. Thus, an interaction was predicted, where the difference in preference should be highest for uninformed participants with higher IU. A relatively high correlation between IU and BI has been reported (Radell & McGuire, 2021), suggesting considerable overlap between the two traits. Thus, a second goal of the study was to test whether the three-choice paradigm can distinguish between IU and BI. If so, this would help establish its discriminant validity.
2. Materials and Methods
2.1. Participants
An a priori power analysis based on a multiple regression with 4 predictors, assuming a small to medium effect size (f2 = 0.15), indicated that 85 participants would be needed for at least 80% power. The fourth predictor was included to account for the interaction term in the model. A total of 79 students were recruited from psychology courses at a private university in the northeastern United States through announcements made in class or posted on the course website. The only eligibility requirement was to be at least 18 years old. Those who chose to participate were compensated with extra credit in the course.
2.2. Procedure
2.2.1. Questionnaires
Participants completed a demographics questionnaire, the brief 12-item version of the Intolerance of Uncertainty Scale (IUS-12) (Carleton et al., 2007), and the Adult Measure of Behavioral Inhibition (AMBI) (Gladstone & Parker, 2005). In the current sample, internal consistency was α = 0.91 (95% CI [0.88, 0.94]) for the IUS-12 and α = 0.85 (95% CI [0.80, 0.90]) for the AMBI, indicating high reliability for both scales. The questionnaires were administered on a computer and always after the CPP task described below.
2.2.2. Conditioned Place Preference Task
Participants completed a computer-based CPP task modified from prior research (Radell et al., 2016, 2018). The task involved controlling a cartoon fox while searching for rewards (golden eggs) hidden in chests across different rooms. A script of the prompts seen by participants is included in Appendix A. Different rooms were paired with a different chance of reward or punishment (i.e., losing already obtained rewards), which they had to discover through trial-and-error. The task employed a posttest-only design, with a tutorial, training, and test phase (Figure 1A). The participants were first forced to enter each of the available rooms, one at a time, and allowed to forage inside. Unlike past research (Radell et al., 2016, 2018), which placed a limit on the amount of time participants could forage, the current version of the task limited the number of opportunities available to search for reward. Participants were given a set number of keys that they could use to open chests (35 per room during training, and 40 during the test phase), with the number of remaining keys displayed at the top of the screen. This was performed to make the number of opportunities available to acquire reinforcement explicit and encourage them to make more careful decisions about which locations they searched.
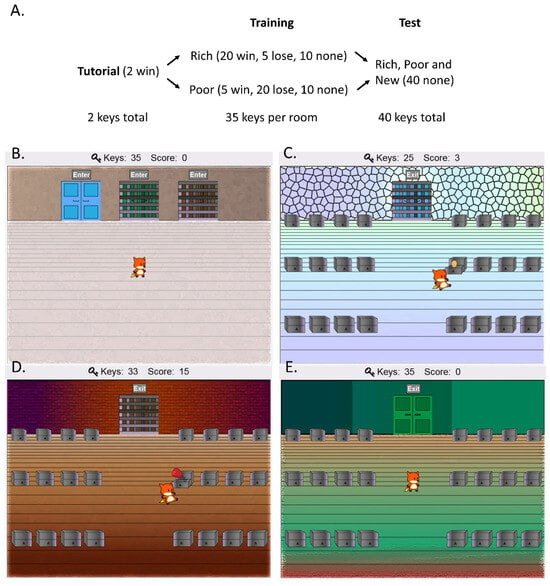
Figure 1.
Task design. (A) The task consisted of a tutorial, training, and test phases. The number of keys available to open chests in each phase are listed at the bottom, with the possible outcomes in each room listed in parenthesis. (B) Participants controlled a cartoon fox to forage for rewards (golden eggs) by opening chests in a virtual environment consisting of multiple rooms connected through a lobby area. Opportunities to forage were limited by the number of keys available to open chests. It was also possible to lose points when opening some of the chests. During the training phase, one room (the rich room) was associated with more wins but fewer losses while another room (the poor room) was the opposite. The rooms were visually distinct—the (C) rich room was always blue while the (D) poor room was always brown. The order in which participants experienced the rooms was counterbalanced. (E) During the test phase, participants could freely switch between rooms, and a new room became available. The door leading to the new room was either blue, matching that of the rich room, or a completely new color (green).
The training phase consisted of two conditioning sessions, one for each of the rooms located on the left and right. One of the rooms (the rich room) was paired with a higher number of wins (20 of 35 trials) but a lower number of losses (5 of 35 trials). The second room (the poor room) was the opposite. A win was paired with an animation where the fox discovered a golden egg inside the chest (Figure 1C) and the score counter at the top of the screen increased by one. In contrast, losses were paired with the fox becoming dazed by a boxing glove that sprung from the chest (Figure 1D) and a brief timeout period (2 sec) during which the participant lost control of the character. The score counter also decreased by one. Both rooms had an equal number of no outcome trials (10 of 35) on which neither reward nor punishment was given. On these trials, the chest appeared empty when opened. Unknown to the participants, the outcome of each trial was predetermined but in pseudorandom order.
At the start of the training phase, the fox was placed in a lobby area (Figure 1B) and could only enter one of the rooms (e.g., the rich room; Figure 1C)—the others remained locked. After using up the keys in that room, the fox was again placed in the lobby area, at which point the other room could be accessed (e.g., the poor room; Figure 1D). The order in which participants experienced each room was counterbalanced such that half of them were sent to the rich room first while the other half were sent to the poor room first. However, the rich room was always blue and located on the left, while the poor room was always brown and located on the right, for all participants. The room in the center was locked during the training phase. Since the overall number of wins and losses (across the two rooms) were equal (total of 25), all participants ended this phase with a score of zero.
At the start of the test phase, the fox was again placed in the lobby, but participants could now freely explore all of the rooms. At this time, a new room also became available (Figure 1E). The door to this room was always located in the center. To increase the familiarity of this room, for half of the participants assigned at random, the color of the door leading to this room matched that of the rich room (i.e., the door was blue)—for the remainder, the color was completely new (i.e., green; Figure 1B). Thus, preference for the new room could be modulated by the amount of generalization from the rich to the new room, or how much each room is coded in terms of color vs. location, since in one condition the rooms would be signaled by the same color. In this phase, none of the 40 trials were rewarded or punished to prevent the possibility of feedback resulting in additional conditioning and altering room preference. The percent of total keys used in each room was recorded as the main dependent measure.
2.3. Data Analysis
To examine whether IU was related to BI, a Pearson’s correlation was computed between scores on the IUS-12 and AMBI (as measures of IU and BI, respectively). To determine whether personality was related to foraging behavior in the task, a multiple regression was performed on the percent total keys used in the new room, as well as in the rich room during the posttest. For both analyses, the predictors entered in the model were new room door color (same as rich room vs. new), total IU and BI using a forced-entry method. To test the hypothesis that participants with higher IU would be more likely to forage in the new room when it was more familiar (i.e., when its door color matched that of the rich room), the regression model also included the interaction between IU and door color. The analyses were conducted in R (version 3.4.3; The R Software Foundation) and JASP (version 0.17.2.1; JASP Team). All tests were two-tailed. Data are available on the Open Science Framework.
3. Results
Two participants were excluded from the statistical analyses due to incomplete responses on the IUS-12. The remaining sample of 77 (85.71% female) had a mean age of 19.81 (SD = 2.26, range = 18–33). The sample was predominantly white (84.42%) with 5 participants self-reporting as black or African American, 2 as Asian, 4 who identified with more than one racial group, and 1 participant who chose not to answer this question. When asked about ethnicity, only 2 participants identified as Hispanic or Latino. The two most common majors reported were Psychology (49.35%, including double majors) and Nursing (22.08%).
Overall, ignoring the other factors, percent keys used in each room (i.e., foraging behavior) indicated a preference for the rich (M = 34.38%, SE = 2.37) and new rooms (M = 40.23%, SE = 2.31), over the poor room (M = 25.39%, SE = 1.86). This was confirmed by a repeated measures ANOVA (F(1.85, 140.80) = 7.76, p < 0.001, η2p = 0.093) with Greenhouse-Geisser correction applied, since Mauchly’s test indicated the assumption of sphericity was violated (χ2(2) = 6.22, p = 0.045). Post hoc comparisons with Holm adjustment confirmed the pattern described above: preference for the rich and new rooms was significantly higher than for the poor room (p = 0.038 and <0.001, respectively) with no difference between the rich and new rooms (p = 0.126). Most entered the new room first during the test (77.92% of the sample), with only 18.18% and 3.90% entering the rich and poor rooms first, respectively.
Total IU and BI were comparable between participants assigned to the same as rich vs. new information condition based on the new room’s door color. This was confirmed by Welch’s t-tests performed on total IU and BI (t(75) = 0.48, p = 0.633 and t(75) = 0.11, p = 0.91, respectively).
A multiple regression performed on percent keys used in the new room, including new room door color, IU, BI and the new room door color by IU interaction, returned a significant model (Table 1). The significant predictors were new room door color, IU, and their interaction. As expected, higher IU was associated with reduced foraging in the new room. However, this also depended on information provided about the room. Participants with higher IU foraged less in the new room only when its door color matched that of the rich room (Pearson’s r(35) = −0.45, p = 0.005, 95% CI −0.67 to −0.14; Figure 2A left panel). In contrast, there was no relationship between IU and foraging in the new room when the door color was new (Pearson’s r(38) = −0.11, p = 0.489, 95% CI −0.41 to 0.21; Figure 2A right panel). As expected, a significant positive correlation was found between IU and BI (Pearson’s r(75) = 0.57, p < 0.001, r2 = 0.32; Figure 2B). Despite this correlation, BI was not a significant predictor of foraging in the new room (Figure 2C) suggesting that this behavior taps into a relatively unique aspect of IU. Lastly, the significant effect of door color indicated that participants used fewer keys in the new room when its door color was new (M = 35.25%, SE = 2.32) than when it matched the rich room (M = 45.61%, SE = 3.96). To understand the micro-level behavior underlying the observed preference differences, changes in room occupancy over time during the test were also plotted (Figure 3), split by the main effects of door color and IU. Near the start of the test, occupancy of the new room was lower when the door color was familiar (Figure 3A) rather than new (Figure 3B). Though initial occupancy was similar between participants with low vs. high IU, those with low IU remained in the room longer (Figure 3C) compared to those with high IU (Figure 3D).

Table 1.
Summary of Multiple Regression Analysis for Variables Predicting Percent Keys Used in the New Room (n = 77).
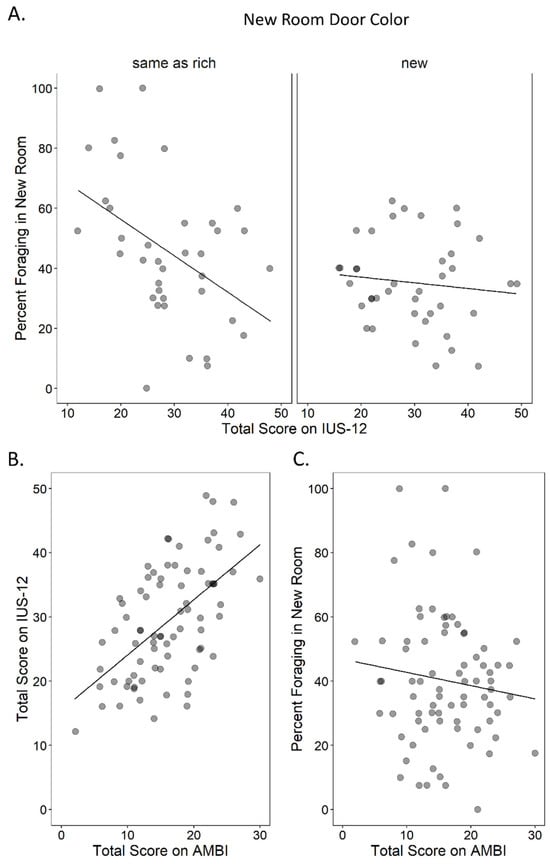
Figure 2.
Task results. (A) Foraging behavior expressed as mean percent keys used in the new room during the test phase as a function of total IU, split by whether the new room door color was the same as the rich room (left), or new (right). Higher IU was associated with less foraging, but only when the door color was the same as the rich room (r = −0.45, p = 0.005). (B) There was a significant positive correlation between IU and BI (r = 0.57, p < 0.001), but (C) BI was not related to foraging in the new room (r = −0.12, p = 0.300).
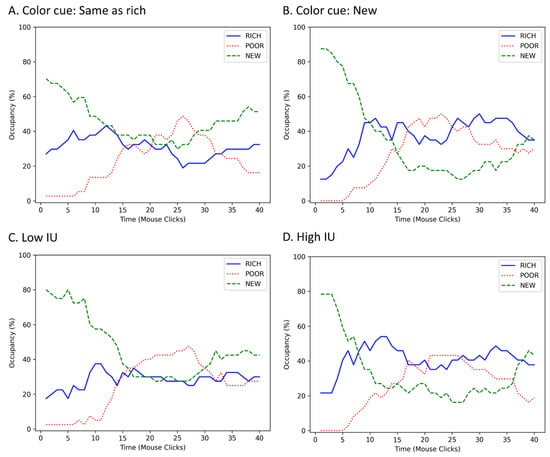
Figure 3.
Room occupancy over time during the test as a percent of total people within group. Based on visual inspection, (A) when the color of the door to the new room was familiar (i.e., same as that of the rich room), occupancy of the room was lower near the start of the test compared to (B) when the color was new. People with (C) low IU appear to remain in the new room longer than (D) people with high IU.
Multiple regression with the same predictors as above but performed on percent keys used in the rich room was not significant (R2 = 0.078, F(4, 72) = 1.524, p = 0.204). Thus, there was no evidence that IU or BI were related to foraging in the rich room.
4. Discussion
In partial support of the hypothesis, participants with higher IU showed a lower preference for a new room, and this depended on the information provided about the room. However, this interaction was in the opposite direction of what was predicted. It was expected that providing a cue suggesting that the new room is similar to the previously rewarded, familiar room, would reduce uncertainty, and therefore increase exploration. Instead, providing the cue reduced exploration. That is, higher IU was associated with less foraging in the new room, but only when the color of the door to that room was the same as that of the rich room (Figure 2A). There was no relationship between IU and foraging in the new room when the color of the door was completely new (Figure 2B).
How can this be explained? One possibility is that IU is more important not when there is a choice between familiar and unfamiliar, but rather when there is ambiguity between familiar and unfamiliar options. In this case, the choice was between two rooms, one familiar and one new, but the provided information (i.e., color cue) suggested that the new room was similar to one of the familiar rooms (the rich room). This may have increased avoidance, because the ambiguous information was interpreted more negatively, as in seen in anxiety disorders (Malivoire et al., 2019) that are related to IU (Carleton, 2012). However, this was not expressed in the first choice participants made at the start of the test—most still entered the new room first, regardless of IU. Instead, as suggested by changes in room occupancy, those with higher IU (Figure 3D) may have been more likely to leave the first room and go back to the rich room, after their initial foraging attempts did not result in reward. Thus, people with higher IU may be faster to change their behavior and exploit familiar alternatives when expectancies are violated, compared to those with lower IU, who tended to remain the new room longer (Figure 3C).
The strong preference for the new room at the start of the test suggests that most participants, regardless of IU, sought information about that room. Why did participants with higher IU not avoid this room instead? Avoidance might be more likely for contexts that elicit threat or worry, which is where this personality trait has been found to have a greater effect (Koerner et al., 2017). Although the participants could lose points in the task, this is a low intensity punisher that does not elicit the same level of response as the aversive stimuli (e.g., electric shock, lithium chloride) commonly used in conditioning experiments, as well as real-life situations that involve uncertainty and are perceived as threatening (e.g., taking an exam or interviewing for a job). Also, as with any behavior, punishment due to potential point loss may have been counteracted by concurrent reinforcement, such as that provided by exploring the new room. In a changing environment, which is also less predictable, seeking information to reduce uncertainty may be more important in the long-term (Anselme, 2023). Thus, exploration of new alternatives might be preferred over exploitation, at least at first.
The novelty preference observed here is consistent with animal CPP studies. For example, rats given a choice between a new and a familiar chamber show a preference for the new chamber (Parker, 1992). This preference can be abolished by dopamine receptor antagonists, which decrease dopamine activity, suggesting that exploring the new chamber is reinforcing (Bardo et al., 1989). Similarly, monkeys given a dopamine reuptake inhibitor, which increases dopamine activity, showed increased preference for novelty in a probabilistic decision-making task (Costa et al., 2014). Preference for novelty, however, likely depends on the type of reinforcer, and on species. For example, in a three-chamber apparatus, rats prefer a morphine or amphetamine-paired chamber over a novel chamber, but show the lowest preference for a saline-paired chamber (Parker, 1992). The value of the reinforcer used in the current study (i.e., points) is of course much lower than that of the highly addictive drugs typically used as reinforcers in animal studies. Other research has suggested that the addition of a novel choice can enhance the preference for an otherwise weak reinforcer. For example, zebrafish showed a conditioned place preference for caffeine only in an apparatus that had at least one novel chamber. In contrast to rats, they did not prefer the novel chamber over a familiar chamber (Faillace et al., 2018).
Motivation, and the perceived value of the stimuli will also likely affect the strength of the preference. A previous study in humans compared preference for two virtual rooms, one paired with chocolate, and found participants spent more time in the chocolate-paired room, but only if they were food deprived (Astur et al., 2014). Even so, preference has been previously shown for secondary reinforcers, including points (Astur et al., 2016) and money (Childs et al., 2017), in two-room paradigms. In the current study, despite the strong initial preference for the new room, overall preference was similar between the rich and new rooms, and was significantly lower for the poor room.
In contrast to the animal literature, CPP in people has been rarely studied, and a recent review found no place aversion studies (Linhardt et al., 2022). As such, it is unclear what task parameters can increase the reliability of place preference effects in humans, and their sensitivity to IU. For example, a prior study reported that college students with higher IU tended to return to the rich room, but only when first given a choice (Radell et al., 2016). That study used a two-room paradigm involving only reward, not punishment. The addition of a novel, third room in the current study may have precluded observing this relationship. The task in the current study involved both reward and punishment (e.g., the rich room was more rewarding but less punishing), and was not designed to establish whether any differences in preference are due to appetitive or aversive conditioning. For example, a higher preference for the rich room could be due to more appetitive conditioning, less aversive conditioning, or a combination of the two. This remains a direction for future research.
Another factor may be the type of environment—physical vs. virtual. Place preference effects appear to be more robust in a real, physical environment (Linhardt et al., 2022). It is also possible that a more realistic virtual environment (e.g., one observed through a virtual reality headset) would result in a stronger effect compared to three- and two-dimensional environments presented on a computer screen. However, in addition to being more resource intensive, virtual reality paradigms are not readily transferable online. The current task on other hand, can be reworked to run in a browser to reach larger, more representative samples. In the future, the task can also be supplemented with self-report to help understand the decisions participants made, as well as ratings to indicate how much they liked each room, to assess explicit knowledge of reward contingencies, as in some past studies of human place preference (Linhardt et al., 2022).
To increase the familiarity of the new room, information in the form of a color cue was provided but the participants were not screened for color blindness. This is a limitation of the study. However, this is unlikely to have influenced the results because most of the sample was female and Caucasian (about 86 and 84%, respectively). The prevalence of red-green color blindness—the most common form of color blindness—is less than 1% in Caucasian women (Birch, 2012). Though the prevalence is higher in men at about 8% (Birch, 2012), the pattern of results is the same even when the few men are excluded from the sample. Furthermore, the color of the door to the rich room (and the door to the new room, when it matched) was always blue, and should not be confused with that of the other rooms even by someone who is red-green colorblind. Future research should replicate this result with a different familiarity manipulation—for example, a verbal cue could be provided instead.
In the current study, the lower preference for the new room was associated only with higher IU, and not BI, despite the relatively high correlation between the two traits. This suggests that preference for novelty taps into a unique aspect of IU. To the author’s knowledge, this is the first study to examine the behavioral differences between IU and BI using a computer-based task. Other than BI, IU is related to several other constructs, including tolerance of ambiguity, indecisiveness, risk aversion, and curiosity. For example, in one study based on self-report, the weakest correlation found was with curiosity (r = −0.29) while the highest correlation, besides that with worry (r = 0.64), was with indecisiveness (r = 0.59) (Koerner et al., 2017). In a review of studies published between 1986 and 2012 (Rosen et al., 2014), only three measured IU and at least one related construct—tolerance of ambiguity, uncertainty orientation, and the need for cognitive closure. Thus, more research is needed to establish how IU differs from these, and other related constructs. Importantly, this includes their behavioral correlates.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Niagara University Institutional Review Board (2017-072, approval date 12 December 2017).
Informed Consent Statement
Informed consent was obtained from all participants prior to the start of the study procedures.
Data Availability Statement
The data are available on the Open Science Framework (https://osf.io/ynsfg/, accessed on 7 January 2025).
Conflicts of Interest
The author declares no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| CPP | Conditioned place preference |
| BI | Behavioral inhibition |
| IU | Intolerance of uncertainty |
Appendix A
Task Script
Help Kit the Fox collect golden eggs. First, you’ll practice how to collect eggs.
When you’re ready to begin, click start.
<after clicking start>
Click on the chests to search for golden eggs. Opening a chest will use up a key!
<after clicking on two chests to get two eggs (chests in the tutorial always contain an egg)>
Great! Now enter the next room by clicking on the door.
<after clicking on the door>
Next, you’ll start the real game.
Try to collect as many eggs as you can before your keys run out!
When you’re ready to begin, click start.
<after clicking start, in lobby with only one side door available, the other side door and center door are barred, 35 keys available>
<after using the keys, sent back to lobby, now with the other side door available, 35 keys again>
<after using keys, back to lobby, all three doors available, 40 keys>
You can now freely switch between rooms. A new room has also become available.
<the message above is displayed throughout the entire test phase>
References
- Anselme, P. (2023). Exploratory search: Information matters more than primary reward. Animal Behavior and Cognition, 10(4), 366–383. [Google Scholar] [CrossRef]
- Astur, R. S., Carew, A. W., & Deaton, B. E. (2014). Conditioned place preferences in humans using virtual reality. Behavioural Brain Research, 267, 173–177. [Google Scholar] [CrossRef]
- Astur, R. S., Palmisano, A. N., Carew, A. W., Deaton, B. E., Kuhney, F. S., Niezrecki, R. N., Hudd, E. C., Mendicino, K. L., & Ritter, C. J. (2016). Conditioned place preferences in humans using secondary reinforcers. Behavioural Brain Research, 297, 15–19. [Google Scholar] [CrossRef]
- Bardo, M. T., Neisewander, J. L., & Pierce, R. C. (1989). Novelty-induced place preference behavior in rats: Effects of opiate and dopaminergic drugs. Pharmacology Biochemistry and Behavior, 32(3), 683–689. [Google Scholar] [CrossRef]
- Birch, J. (2012). Worldwide prevalence of red-green color deficiency. Journal of the Optical Society of America A, 29(3), 313–320. [Google Scholar] [CrossRef] [PubMed]
- Carleton, R. N. (2012). The intolerance of uncertainty construct in the context of anxiety disorders: Theoretical and practical perspectives. Expert Review of Neurotherapeutics, 12(8), 937–947. [Google Scholar] [CrossRef]
- Carleton, R. N. (2016). Fear of the unknown: One fear to rule them all? Journal of Anxiety Disorders, 41, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Carleton, R. N., Duranceau, S., Shulman, E. P., Zerff, M., Gonzales, J., & Mishra, S. (2016). Self-reported intolerance of uncertainty and behavioural decisions. Journal of Behavior Therapy and Experimental Psychiatry, 51, 58–65. [Google Scholar] [CrossRef]
- Carleton, R. N., Norton, M. A. P. J., & Asmundson, G. J. G. (2007). Fearing the unknown: A short version of the intolerance of uncertainty scale. Journal of Anxiety Disorders, 21(1), 105–117. [Google Scholar] [CrossRef] [PubMed]
- Childs, E., Astur, R. S., & de Wit, H. (2017). Virtual reality conditioned place preference using monetary reward. Behavioural Brain Research, 322(Pt A), 110–114. [Google Scholar] [CrossRef]
- Clauss, J. A., & Blackford, J. U. (2012). Behavioral inhibition and risk for developing social anxiety disorder: A meta-analytic study. Journal of the American Academy of Child and Adolescent Psychiatry, 51(10), 1066–1075. [Google Scholar] [CrossRef]
- Contractor, A. A., Elhai, J. D., Ractliffe, K. C., & Forbes, D. (2013). PTSD’s underlying symptom dimensions and relations with behavioral inhibition and activation. Journal of Anxiety Disorders, 27(7), 645–651. [Google Scholar] [CrossRef]
- Costa, V. D., Tran, V. L., Turchi, J., & Averbeck, B. B. (2014). Dopamine modulates novelty seeking behavior during decision making. Behavioral Neuroscience, 128(5), 556–566. [Google Scholar] [CrossRef]
- Faillace, M. P., Pisera-Fuster, A., & Bernabeu, R. (2018). Evaluation of the rewarding properties of nicotine and caffeine by implementation of a five-choice conditioned place preference task in zebrafish. Progress in Neuro-Psychopharmacology and Biological Psychiatry, 84, 160–172. [Google Scholar] [CrossRef]
- Freeston, M. H., Rhéaume, J., Letarte, H., Dugas, M. J., & Ladouceur, R. (1994). Why do people worry? Personality and Individual Differences, 17(6), 791–802. [Google Scholar] [CrossRef]
- Gladstone, G., & Parker, G. (2005). Measuring a behaviorally inhibited temperament style: Development and initial validation of new self-report measures. Psychiatry Research, 135(2), 133–143. [Google Scholar] [CrossRef]
- Koerner, N., Mejia, T., & Kusec, A. (2017). What’s in a name? Intolerance of uncertainty, other uncertainty-relevant constructs, and their differential relations to worry and generalized anxiety disorder. Cognitive Behaviour Therapy, 46(2), 141–161. [Google Scholar] [CrossRef] [PubMed]
- Linhardt, M., Kiser, D. P., Pauli, P., & Hilger, K. (2022). Approach and avoidance beyond verbal measures: A quantitative meta-analysis of human conditioned place preference studies. Behavioural Brain Research, 426, 113834. [Google Scholar] [CrossRef] [PubMed]
- Malivoire, B. L., Marcos, M., Pawluk, E. J., Tallon, K., Kusec, A., & Koerner, N. (2019). Look before you leap: The role of negative urgency in appraisals of ambiguous and unambiguous scenarios in individuals high in generalized anxiety disorder symptoms. Cognitive Behaviour Therapy, 48(3), 217–240. [Google Scholar] [CrossRef] [PubMed]
- Muris, P., van Brakel, A. M., Arntz, A., & Schouten, E. (2011). Behavioral inhibition as a risk factor for the development of childhood anxiety disorders: A longitudinal study. Journal of Child and Family Studies, 20, 157–170. [Google Scholar] [CrossRef]
- Myers, C. E., Kostek, J. A., Ekeh, B., Sanchez, R., Ebanks-Williams, Y., Krusznis, A. L., Weinflash, N., & Servatius, R. J. (2016). Watch what I do, not what I say I do: Computer-based avatars to assess behavioral inhibition, a vulnerability factor for anxiety disorders. Computers in Human Behavior, 55, 804–816. [Google Scholar] [CrossRef] [PubMed]
- Myers, C. E., VanMeenen, K. M., & Servatius, R. J. (2012). Behavioral inhibition and PTSD symptoms in veterans. Psychiatry Research, 196(2–3), 271–276. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Parker, L. A. (1992). Place conditioning in a three- or four-choice apparatus: Role of stimulus novelty in drug-induced place conditioning. Behavioral Neuroscience, 106(2), 294–306. [Google Scholar] [CrossRef] [PubMed]
- Radell, M. L., Allen, M. T., Favaloro, B., Myers, C. E., Haber, P., Morley, K., & Moustafa, A. A. (2018). Intolerance of uncertainty and conditioned place preference in opioid addiction. PeerJ, 6, e4775. [Google Scholar] [CrossRef]
- Radell, M. L., & McGuire, B. M. (2021). Intolerance of uncertainty predicts fear of healthcare settings but not desire for testing during the novel coronavirus pandemic. Discover Psychology, 1(1), 10. [Google Scholar] [CrossRef]
- Radell, M. L., Myers, C. E., Beck, K. D., Moustafa, A. A., & Allen, M. T. (2016). The personality trait of intolerance to uncertainty affects behavior in a novel computer-based conditioned place preference task. Frontiers in Psychology, 7, 1175. [Google Scholar] [CrossRef]
- Rosen, N. O., Ivanova, E., & Knäuper, B. (2014). Differentiating intolerance of uncertainty from three related but distinct constructs. Anxiety, Stress & Coping, 27(1), 55–73. [Google Scholar] [CrossRef]
- Sheynin, J., Beck, K. D., Pang, K. C. H., Servatius, R. J., Shikari, S., Ostovich, J., & Myers, C. E. (2014). Behaviourally inhibited temperament and female sex, two vulnerability factors for anxiety disorders, facilitate conditioned avoidance (also) in humans. Behavioural Processes, 103, 228–235. [Google Scholar] [CrossRef]
- Sheynin, J., Moustafa, A. A., Beck, K. D., Servatius, R. J., Casbolt, P. A., Haber, P., Elsayed, M., Hogarth, L., & Myers, C. E. (2016). Exaggerated acquisition and resistance to extinction of avoidance behavior in treated heroin-dependent males. The Journal of Clinical Psychiatry, 77(3), 386–394. [Google Scholar] [CrossRef]
- Sheynin, J., Shind, C., Radell, M., Ebanks-Williams, Y., Gilbertson, M. W., Beck, K. D., & Myers, C. E. (2017). Greater avoidance behavior in individuals with posttraumatic stress disorder symptoms. Stress, 20(3), 285–293. [Google Scholar] [CrossRef]
- Tzschentke, T. M. (2007). Measuring reward with the conditioned place preference (CPP) paradigm: Update of the last decade. Addiction Biology, 12(3–4), 227–462. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).