
On the Incompleteness of the Coelacanth Fossil Record


Abstract
1. Introduction
2. Materials and Methods
3. Result
3.1. Stages of Coelacanth Research History
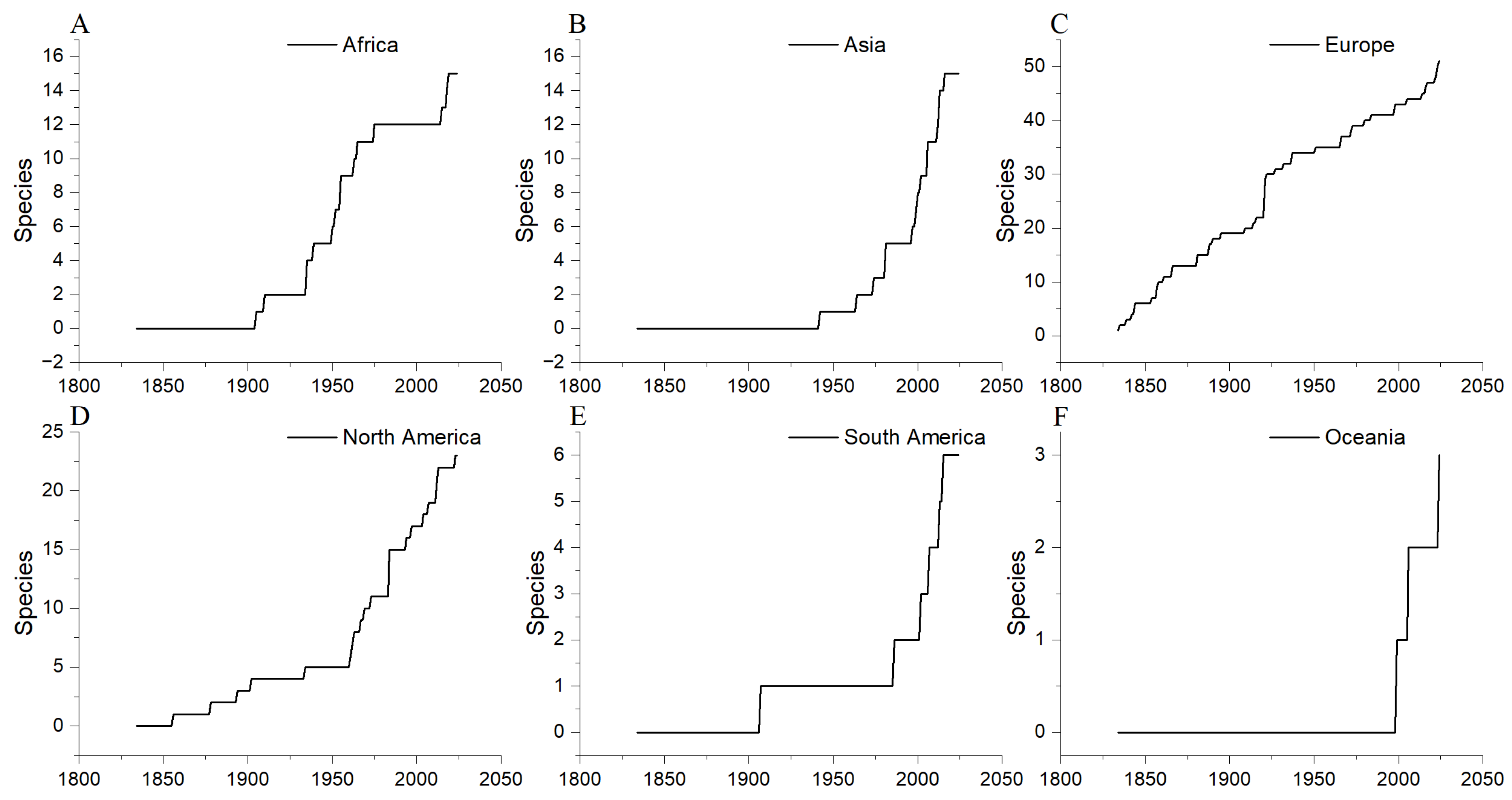
3.2. Spatial Heterogeneity in Coelacanth Fossil Record
3.3. Geographic Distribution of Coelacanths: From the Paleozoic to the Present
3.4. Evaluating Coelacanth Research Efforts by Continent Using Fossil Site Density
3.5. Temporal Heterogeneity in Coelacanth Fossil Record
3.6. Sampling Curves Across Different Time Intervals
3.7. Assessing Coelacanth Species Counts at Documented Sites via Fossil Richness
3.8. Assessing Coelacanth Fossil Record Based on Phylogenetic Diversity
4. Discussion
4.1. Spatial and Temporal Assessment of Coelacanth Fossil Record
4.2. Validity of Diversity Peaks
4.3. Factors Contributing to Incomplete Coelacanth Fossil Record
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Darwin, C. On the Origin of the Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life; John Murray: London, UK, 1859. [Google Scholar]
- Behrensmeyer, A.K.; Kidwell, S.M. Taphonomy’s Contributions to Paleobiology. Paleobiology 1985, 11, 105–119. [Google Scholar]
- Allison, P.A.; Bottjer, D.J. Taphonomy: Bias and Process Through Time. In Taphonomy; Allison, P.A., Bottjer, D.J., Eds.; Topics in Geobiology; Springer: Dordrecht, The Netherlands, 2010; Volume 32, pp. 1–17. [Google Scholar] [CrossRef]
- Smith, A.B.; McGowan, A.J. The Shape of the Phanerozoic Marine Palaeodiversity Curve: How Much Can Be Predicted from the Sedimentary Rock Record of Western Europe? Palaeontology 2007, 50, 765–774. [Google Scholar] [CrossRef]
- Raup, D.M. Taxonomic Diversity during the Phanerozoic: The Increase in the Number of Marine Species since the Paleozoic May Be More Apparent than Real. Science 1972, 177, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Kidwell, S.M.; Holland, S.M. The Quality of the Fossil Record: Implications for Evolutionary Analyses. Annu. Rev. Ecol. Syst. 2002, 33, 561–588. [Google Scholar] [CrossRef]
- Smith, A. Large-Scale Heterogeneity of the Fossil Record: Implications for Phanerozoic Biodiversity Studies. Philos. Trans. R. Soc. B Biol. Sci. 2001, 356, 351–367. [Google Scholar] [CrossRef]
- Benton, M.J. Diversification and Extinction in the History of Life. Science 1995, 268, 52–58. [Google Scholar] [CrossRef]
- Kidwell, S.M.; Flessa, K.W. The Quality of the Fossil Record: Populations, Species, and Communities. Annu. Rev. Ecol. Syst. 1995, 26, 269–299. [Google Scholar] [CrossRef]
- Friedman, M.; Sallan, L.C. Five Hundred Million Years of Extinction and Recovery: A Phanerozoic Survey of Large-Scale Diversity Patterns in Fishes. Palaeontology 2012, 55, 707–742. [Google Scholar] [CrossRef]
- Guinot, G.; Adnet, S.; Cappetta, H. An Analytical Approach for Estimating Fossil Record and Diversification Events in Sharks, Skates, and Rays. PLoS ONE 2012, 7, e44632. [Google Scholar] [CrossRef]
- Cleary, T.J.; Moon, B.C.; Dunhill, A.M.; Benton, M.J. The Fossil Record of Ichthyosaurs, Completeness Metrics and Sampling Biases. Palaeontology 2015, 58, 521–536. [Google Scholar] [CrossRef]
- Mannion, P.D.; Upchurch, P. Completeness Metrics and the Quality of the Sauropodomorph Fossil Record through Geological and Historical Time. Paleobiology 2010, 36, 283–302. [Google Scholar] [CrossRef]
- Benton, M.J.; Ruta, M.; Dunhill, A.M.; Sakamoto, M. The First Half of Tetrapod Evolution, Sampling Proxies, and Fossil Record Quality. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 372, 18–41. [Google Scholar] [CrossRef]
- Brocklehurst, N.; Upchurch, P.; Mannion, P.D.; O’Connor, J. The Completeness of the Fossil Record of Mesozoic Birds: Implications for Early Avian Evolution. PLoS ONE 2012, 7, e39056. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, G.T.; Friedman, M. A Survey of Palaeontological Sampling Biases in Fishes Based on the Phanerozoic Record of Great Britain. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 372, 5–17. [Google Scholar] [CrossRef]
- Henderson, S.; Dunne, E.M.; Fasey, S.A.; Giles, S. The Early Diversification of Ray-finned Fishes (Actinopterygii): Hypotheses, Challenges, and Future Prospects. Biol. Rev. Camb. Philos. Soc. 2023, 98, 284–315. [Google Scholar] [CrossRef]
- Huxley, T.H. Illustrations of the Structure of the Crossopterygian Ganoids; Longmans, Green, Reader, & Dyer: London, UK, 1866. [Google Scholar]
- Simpson, G.G. Tempo and Mode in Evolution; Columbia University Press: New York, NY, USA, 1944. [Google Scholar] [CrossRef]
- Agassiz, L. Recherches sur les Poissons Fossiles; Petitpierre et Prince: Neuchâtel, Switzerland, 1839; Volume 2. [Google Scholar]
- Smith, J.L. A Living Fish of Mesozoic Type. Nature 1939, 143, 455–456. [Google Scholar] [CrossRef]
- Schaeffer, B. A Revision of Coelacanthus Newarki and Notes on the Evolution of the Girdles and Basal Plates of the Median Fins in the Coelacanthini; The American Museum of Natural History: New York, NY, USA, 1941; No. 1110. [Google Scholar]
- Cloutier, R. Patterns, Trends, and Rates of Evolution within the Actinistia. In The Biology of Latimeria chalumnae and Evolution of Coelacanths; Springer: Berlin/Heidelberg, Germany, 1991; pp. 23–58. [Google Scholar]
- Forey, P. The Coelacanth as a Living Fossil. In Living Fossils; Springer: Berlin/Heidelberg, Germany, 1984; pp. 166–169. [Google Scholar]
- Forey, P. Golden Jubilee for the Coelacanth Latimeria chalumnae. Nature 1988, 336, 727–732. [Google Scholar] [CrossRef]
- Clement, A.M.; Cloutier, R.; Lee, M.S.Y.; King, B.; Vanhaesebroucke, O.; Bradshaw, C.J.A.; Dutel, H.; Trinajstic, K.; Long, J.A. A Late Devonian Coelacanth Reconfigures Actinistian Phylogeny, Disparity, and Evolutionary Dynamics. Nat. Commun. 2024, 15, 7529. [Google Scholar] [CrossRef]
- Cavin, L.; Tong, H.; Buffetaut, E.; Wongko, K.; Suteethorn, V.; Deesri, U. The First Fossil Coelacanth from Thailand. Diversity 2023, 15, 286. [Google Scholar] [CrossRef]
- Manuelli, L.; Mondéjar Fernández, J.; Dollman, K.; Jakata, K.; Cavin, L. The Most Detailed Anatomical Reconstruction of a Mesozoic Coelacanth. PLoS ONE 2024, 19, e0312026. [Google Scholar] [CrossRef]
- Ferrante, C.; Cavin, L. Early Mesozoic Burst of Morphological Disparity in the Slow-Evolving Coelacanth Fish Lineage. Sci. Rep. 2023, 13, 11356. [Google Scholar] [CrossRef] [PubMed]
- Toriño, P.; Gausden, S.F.; Etches, S.; Rankin, K.; Marshall, J.E.; Gostling, N.J. An Enigmatic Large Mawsoniid Coelacanth (Sarcopterygii, Actinistia) from the Upper Jurassic Kimmeridge Clay Formation of England. J. Vertebr. Paleontol. 2022, 42, e2125813. [Google Scholar] [CrossRef]
- Toriño, P.; Soto, M.; Perea, D. A Comprehensive Phylogenetic Analysis of Coelacanth Fishes (Sarcopterygii, Actinistia) with Comments on the Composition of the Mawsoniidae and Latimeriidae: Evaluating Old and New Methodological Challenges and Constraints. Hist. Biol. 2021, 33, 3423–3443. [Google Scholar] [CrossRef]
- Ferrante, C.; Menkveld-Gfeller, U.; Cavin, L. The First Jurassic Coelacanth from Switzerland. Swiss J. Palaeontol. 2022, 141, 15. [Google Scholar] [CrossRef]
- Forey, P.L.; Cloutier, R. Literature Relating to Fossil Coelacanths. In The Biology of Latimeria chalumnae and Evolution of Coelacanths; Springer: Berlin/Heidelberg, Germany, 1991; pp. 391–402. [Google Scholar]
- Kadarusman; Sugeha, H.Y.; Pouyaud, L.; Hocdé, R.; Hismayasari, I.B.; Gunaisah, E.; Paradis, E. A Thirteen-Million-Year Divergence Between Two Lineages of Indonesian Coelacanths. Sci. Rep. 2020, 10, 192. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 4. [Google Scholar]
- Bapst, D.W. paleotree: An R Package for Paleontological and Phylogenetic Analyses of Evolution. Methods Ecol. Evol. 2012, 3, 803–807. [Google Scholar] [CrossRef]
- Bell, M.A.; Lloyd, G.T. strap: An R Package for Plotting Phylogenies Against Stratigraphy and Assessing Their Stratigraphic Congruence. Palaeontology 2014, 58, 379–389. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 15 June 2024).
- Agassiz, L. Abgerissene Bemerkungen Über Fossile Fische. Neues Jahrb. Mineral. Geogn. Geol. Petrefaktenkunde 1834, 1834, 379–390. [Google Scholar]
- Reis, O.M. Die Coelacanthinen mit besonderer Berücksichtigung der im Weißen Jura Bayerns vorkommenden Gattungen. Palaeontographica 1888, 35, 1–96. [Google Scholar]
- Stensiö, E.A. Triassic Fishes from Spitzbergen; A. Holzhausen: Vienna, Austria, 1921. [Google Scholar] [CrossRef]
- Renesto, S.; Kustatscher, E. A Coelacanth Fish from the Anisian (Middle Triassic) of the Dolomites. Riv. Ital. Paleontol. Stratigr. 2019, 125. [Google Scholar] [CrossRef]
- Newberry, J.S. Description of Several New Genera and Species of Fossil Fishes from the Carboniferous Strata of Ohio. Proc. Acad. Nat. Sci. USA 1856, 8, 96–100. [Google Scholar]
- Stensiö, E.A. Triassic Fishes from East Greenland. Meddelelser Grønland Cph. 1932, 83, 1–305. [Google Scholar]
- Lund, R.; Lund, W. New Genera and Species of Coelacanths from the Bear Gulch Limestone (Lower Carboniferous) of Montana (USA). Geobios 1984, 17, 237–244. [Google Scholar] [CrossRef]
- Broom, R. On a Species of Coelacanthus from the Upper Beaufort Beds of Aliwal North. Rec. Albany Mus. Grahamst. 1905, 1, 338–339. [Google Scholar]
- Moy-Thomas, J.A. The Coelacanth Fishes from Madagascar. Geol. Mag. 1935, 72, 213–227. [Google Scholar] [CrossRef]
- Clément, G. The Actinistian (Sarcopterygii) Piveteauia madagascariensis Lehman from the Lower Triassic of Northwestern Madagascar: A Redescription on the Basis of New Material. J. Vertebr. Paleontol. 1999, 19, 234–242. [Google Scholar] [CrossRef]
- Yabumoto, Y.; Brito, P.M.; Iwata, M.; Abe, Y. A New Triassic Coelacanth, Whiteia uyenoteruyai (Sarcopterygii, Actinistia) from Madagascar and Paleobiogeography of the Family Whiteiidae. Bull. Kitakyushu Mus. Nat. Hist. Hum. Hist. Ser. A (Nat. Hist.) 2019, 17, 15–27. [Google Scholar]
- Woodward, A.S. Some New and Little-Known Upper Cretaceous Fishes from Mount Lebanon. Ann. Mag. Nat. Hist. 1942, 9, 537–568. [Google Scholar] [CrossRef]
- Yabumoto, Y. A New Coelacanth from the Early Cretaceous of Brazil (Sarcopterygii, Actinistia). Paleontol. Res. 2002, 6, 343–350. [Google Scholar] [CrossRef]
- Maisey, J.G. Coelacanths from the Lower Cretaceous of Brazil; American Museum of Natural History: New York, NY, USA, 1986; p. 32. [Google Scholar]
- Arratia, G.; Schultze, H.P. A New Fossil Actinistian from the Early Jurassic of Chile and Its Bearing on the Phylogeny of Actinistia. J. Vertebr. Paleontol. 2015, 35, e983524. [Google Scholar] [CrossRef]
- Liu, H.T. A New Coelacanth from the Marine Lower Triassic of NW Kwangsi, China. Vertebr. Palasiat. 1964, 8, 211–214. (In Chinese) [Google Scholar]
- Wang, N.; Liu, H.T. Coelacanth Fishes from the Marine Permian of Zhejiang, South China. Vertebr. Palasiat. 1981, 19, 305. [Google Scholar]
- Wen, W.; Zhang, Q.-Y.; Hu, S.-X.; Benton, M.J.; Zhou, C.-Y.; Tao, X.; Huang, J.-Y.; Chen, Z.-Q. Coelacanths from the Middle Triassic Luoping Biota, Yunnan, South China, with the Earliest Evidence of Ovoviviparity. Acta Palaeontol. Pol. 2013, 58, 175–193. [Google Scholar] [CrossRef]
- Long, J.A. A New Genus of Fossil Coelacanth (Osteichthyes: Coelacanthiformes) from the Middle Devonian of Southeastern Australia. Rec. West. Aust. Mus. Suppl. 1999, 57, 37–53. [Google Scholar]
- Johanson, Z.; Long, J.A.; Talent, J.A.; Janvier, P.; Warren, J.W. Oldest coelacanth, from the Early Devonian of Australia. Biol. Lett. 2006, 2, 443–446. [Google Scholar] [CrossRef]
- Pouyaud, L.; Wirjoatmodjo, S.; Rachmatika, I.; Tjakrawidjaja, A.; Hadiaty, R.; Hadie, W. A New Species of Coelacanth. C. R. L’acad. Sci.—Ser. III—Sci. Vie 1999, 322, 261–267. [Google Scholar] [CrossRef]
- Forey, P. History of the Coelacanth Fishes; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- Zhu, M.; Yu, X.B. A Primitive Fish Close to the Common Ancestor of Tetrapods and Lungfish. Nature 2002, 418, 767–770. [Google Scholar] [CrossRef]
- Cavin, L.; Valentin, X.; Garcia, G. A New Mawsoniid Coelacanth (Actinistia) from the Upper Cretaceous of Southern France. Cretac. Res. 2016, 62, 65–73. [Google Scholar] [CrossRef]
- Mawson, J.; Woodward, A.S. On the Cretaceous Formation of Bahia (Brazil), and on Vertebrate Fossils Collected Therein. Q. J. Geol. Soc. 1907, 63, 128. [Google Scholar] [CrossRef]
- Aureliano, T.; Ghilardi, A.M.; Duque, R.R.C.; Barreto, A.M.F. On the Occurrence of Pterosauria in Exu, Pernambuco (Lower Cretaceous Romualdo Formation, Araripe Basin), Northeastern Brazil. Estud. Geológicos 2014, 24, 15. [Google Scholar] [CrossRef]
- Schluter, D. The Ecology of Adaptive Radiation; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Schultze, H.-P. Crossopterygier mit Heterozerker Schwanzflosse aus dem Oberdevon Kanadas, Nebst einer Beschreibung von Onychodontida-Resten aus Dem Mitteldevon Spaniens und aus dem Karbon der USA. Palaeontogr. Abt. A 1973, 188, 188–208. [Google Scholar]
- Forey, P.L.; Ahlberg, P.E.; Lukševičs, E.; Zupinš, I. A New Coelacanth from the Middle Devonian of Latvia. J. Vertebr. Paleontol. 2000, 20, 243–252. [Google Scholar] [CrossRef]
- Friedman, M.; Coates, M. A Newly Recognized Fossil Coelacanth Highlights the Early Morphological Diversification of the Clade. Proc. R. Soc. B Biol. Sci. 2006, 273, 245–250. [Google Scholar] [CrossRef]
- Wendruff, A.J.; Wilson, M.V. A Fork-Tailed Coelacanth, Rebellatrix divaricerca, gen. et sp. nov. (Actinistia, Rebellatricidae, fam. nov.), from the Lower Triassic of Western Canada. J. Vertebr. Paleontol. 2012, 32, 499–511. [Google Scholar] [CrossRef]
- Cavin, L.; Mennecart, B.; Obrist, C.; Costeur, L.; Furrer, H. Heterochronic Evolution explains novel body shape in a Triassic Coelacanth from Switzerland. Sci. Rep. 2017, 7, 13695. [Google Scholar] [CrossRef]
- Dai, X.; Davies, J.H.F.L.; Yuan, Z.; Brayard, A.; Ovtcharova, M.; Xu, G.; Liu, X.; Smith, C.P.A.; Schweitzer, C.E.; Li, M.; et al. A Mesozoic Fossil Lagerstätte from 250.8 Million Years Ago Shows a Modern-Type Marine Ecosystem. Science 2023, 379, 567–572. [Google Scholar] [CrossRef]
- Clément, G. A New Coelacanth (Actinistia, Sarcopterygii) from the Jurassic of France, and the Question of the Closest Relative Fossil to Latimeria. J. Vertebr. Paleontol. 2005, 25, 481–491. [Google Scholar] [CrossRef]
- Eastman, C.R. Catalogue of the Fossil Fishes in the Carnegie Museum. Part III. Catalogue of the Fossil Fishes from the Lithographic Stone of Cerin, France. Mem. Carnegie Mus. 1914, 6, 349–388. [Google Scholar]
- Lambers, P. On the Ichthyofauna of the Solnhofen Lithographic Limestone (Upper Jurassic, Germany); Rijksuniversiteit: Groningen, The Netherlands, 1992. [Google Scholar]
- Villalobos-Segura, E.; Stumpf, S.; Türtscher, J.; Jambura, P.L.; Begat, A.; López-Romero, F.A.; Fischer, J.; Kriwet, J. A Synoptic Review of the Cartilaginous Fishes (Chondrichthyes: Holocephali, Elasmobranchii) from the Upper Jurassic Konservat-Lagerstätten of Southern Germany: Taxonomy, Diversity, and Faunal Relationships. Diversity 2023, 15, 386. [Google Scholar] [CrossRef]
- Jain, S.L. Indocoelacanthus robustus n. gen., n. sp. (Coelacanthidae, Lower Jurassic), the First Fossil Coelacanth from India. J. Paleontol. 1974, 48, 49–62. [Google Scholar]
- Woodward, A.S. Catalogue of the Fossil Fishes in the British Museum (Natural History); Trustees of the British Museum: London, UK, 1895. [Google Scholar]
- Benton, M.J.; Emerson, B.C. How Did Life Become So Diverse? The Dynamics of Diversification According to the Fossil Record and Molecular Phylogenetics. Palaeontology 2007, 50, 23–40. [Google Scholar] [CrossRef]
- Ye, S.; Peters, S.E. Bedrock Geological Map Predictions for Phanerozoic Fossil Occurrences. Paleobiology 2023, 49, 394–413. [Google Scholar] [CrossRef]
- Meyers, S.R.; Peters, S.E. A 56 Million Year Rhythm in North American Sedimentation during the Phanerozoic. Earth Planet. Sci. Lett. 2011, 303, 174–180. [Google Scholar] [CrossRef]
- Peters, S.E.; Husson, J.M. Sediment Cycling on Continental and Oceanic Crust. Geology 2017, 45, 323–326. [Google Scholar] [CrossRef]
- Brito, P.M. Description of Aspidorhynchus arawaki from the Late Jurassic of Cuba (Actinopterygii: Halecostomi). Mesoz. Fishes 1999, 2, 239–248. [Google Scholar]
- Holland, S.M. The Non-Uniformity of Fossil Preservation. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150130. [Google Scholar] [CrossRef]
- Li, Q.; Liu, Y.; Li, G.; Wang, Z.; Zheng, Z.; Sun, Y.; Lei, N.; Li, Q.; Zhang, W. Review of the Impact of Whale Fall on Biodiversity in Deep-Sea Ecosystems. Front. Ecol. Evol. 2022, 10, 885572. [Google Scholar] [CrossRef]
- Curray, J.R.; Moore, D.G. Sedimentary and Tectonic Processes in the Bengal Deep-Sea Fan and Geosyncline. In The Geology of Continental Margins; Burk, C.A., Drake, C.L., Eds.; Springer: Berlin/Heidelberg, Germany, 1974; pp. 617–627. [Google Scholar] [CrossRef]
- Sibert, E.C.; Norris, R.D. New Age of Fishes Initiated by the Cretaceous-Paleogene Mass Extinction. Proc. Natl. Acad. Sci. USA 2015, 112, 8537–8542. [Google Scholar] [CrossRef]
- Allison, P.A.; Briggs, D.E.G. Exceptional Fossil Record: Distribution of Soft-Tissue Preservation through the Phanerozoic. Geology 1993, 21, 527–530. [Google Scholar] [CrossRef]
- Münster, G. von Beitrag Zur Kenntniss Einiger Neuen Seltenen Versteinerungen Aus Den Lithographischen Schiefern in Baiern. Neues Jahrb. Mineral. Geogn. Geol. Petrefakten-Kunde 1842, 1842, 35–46. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EU | NA | AF | AS | SA | OC | |
|---|---|---|---|---|---|---|
| ERSD | 1834 | 1856 | 1905 | 1942 | 1907 | 1999 |
| Num_Gen | 51 | 23 | 15 | 15 | 6 | 3 |
| Num_Sp | 93 | 67 | 30 | 17 | 15 | 3 |
| Num_Spe/Mkm2 | 9.14 | 2.71 | 0.98 | 0.38 | 0.84 | 0.35 |
| Paleo_Dist | Conc | Conc | Scat | Scat | Scat | Scat |
| Meso_Dist | Conc | Conc | Mod | Mod | Mod | Scat |
| D1 | D2 | D3 | C1 | C2 | P1 | P2 | P3 | T1 | T2 | T3 | J1 | J2 | J3 | K1 | K2 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Num_Gen_RT | 3 | 5 | 7 | 7 | 2 | 2 | 1 | 4 | 14 | 13 | 7 | 7 | 3 | 9 | 4 | 4 |
| Num_Sp | 3 | 5 | 7 | 8 | 4 | 1 | 0 | 4 | 21 | 13 | 8 | 7 | 2 | 17 | 10 | 8 |
| Chao1_Div | 3.5 | 6.4 | 7.2 | 8.5 | 4.0 | 1.0 | 0.0 | 4.9 | 22.5 | 15.5 | 9.5 | 8.4 | 2.0 | 26.2 | 16.0 | 8.0 |
| iChao1_Div | 3.5 | 9.3 | 7.3 | 8.5 | 4.0 | 1.0 | 0.0 | 5.5 | 23.8 | 17.6 | 13.0 | 11.3 | 2.1 | 32.8 | 16.0 | 8.0 |
| ACE_Div | 4.2 | 11.0 | 7.8 | 8.7 | 4.0 | 1.0 | 0.0 | 5.7 | 22.7 | 16.9 | 11.4 | 10.1 | 3.1 | 27.9 | 15.8 | 8.0 |
| PD_Inc_Gen | 1 | 4 | 4 | 7 | 1 | 2 | 1 | 2 | 9 | 8 | 4 | 6 | 3 | 8 | 4 | 3 |
| PD_Gen | 7 | 9 | 7 | 10 | 4 | 4 | 3 | 4 | 14 | 13 | 11 | 15 | 12 | 12 | 6 | 4 |
| PD_Gen_Diff | 6 | 5 | 3 | 3 | 3 | 2 | 2 | 2 | 5 | 5 | 7 | 9 | 9 | 4 | 2 | 1 |
| PD_Inc_Gen/Ma | 0.04 | 0.38 | 0.17 | 0.20 | 0.04 | 0.08 | 0.07 | 0.26 | 1.91 | 0.78 | 0.11 | 0.22 | 0.23 | 0.48 | 0.09 | 0.09 |
| PD_Gen/Ma | 0.27 | 0.85 | 0.29 | 0.28 | 0.16 | 0.15 | 0.22 | 0.53 | 2.98 | 1.27 | 0.31 | 0.56 | 0.91 | 0.73 | 0.13 | 0.12 |
| PD_Gen/Ma_Diff | 0.23 | 0.47 | 0.13 | 0.08 | 0.12 | 0.08 | 0.15 | 0.26 | 1.06 | 0.49 | 0.20 | 0.34 | 0.68 | 0.24 | 0.04 | 0.03 |
| Samp_Size | 12 | 19 | 48 | 221 | 197 | 21 | 0 | 8 | 118 | 78 | 44 | 23 | 4 | 67 | 127 | 112 |
| Rare_Curve | RS | RS | AP | AP | PL | / | / | RS | AP | AP | AP | RS | RS | RS | RS | PL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Z.; Cavin, L.; Song, H. On the Incompleteness of the Coelacanth Fossil Record. Foss. Stud. 2025, 3, 10. https://doi.org/10.3390/fossils3030010
Yuan Z, Cavin L, Song H. On the Incompleteness of the Coelacanth Fossil Record. Fossil Studies. 2025; 3(3):10. https://doi.org/10.3390/fossils3030010
Chicago/Turabian StyleYuan, Zhiwei, Lionel Cavin, and Haijun Song. 2025. "On the Incompleteness of the Coelacanth Fossil Record" Fossil Studies 3, no. 3: 10. https://doi.org/10.3390/fossils3030010
APA StyleYuan, Z., Cavin, L., & Song, H. (2025). On the Incompleteness of the Coelacanth Fossil Record. Fossil Studies, 3(3), 10. https://doi.org/10.3390/fossils3030010