
Effects of Grazing and Shrub Management on Species Composition and Soil Properties in Patagonian Grasslands


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Sites
2.3. Estimation of Grazing Intensity
2.4. Experimental Design
2.5. Vegetation Monitoring
2.6. Soil Sampling and Analysis
2.7. Statistical Analysis
3. Results
3.1. Vegetation Response to Interventions and Grazing
3.2. Changes in Soil Properties Due to Interventions and Grazing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| gl | Sum of Squares | Mean Squares | F. model | R2 | Pr(>F) | ||
|---|---|---|---|---|---|---|---|
| treatment | 5 | 0.68 | 0.14 | 2.17 | 0.13 | 0.001 | *** |
| YEAR | 1 | 0.58 | 0.58 | 9.35 | 0.11 | 0.000 | *** |
| treatment:YEAR | 5 | 0.14 | 0.03 | 0.45 | 0.03 | 0.998 | |
| Residuals | 60 | 3.75 | 0.06 | 0.73 | |||
| Total | 71 | 5.16 | 1 |
References
- Godde, C.M.; Garnett, T.; Thornton, P.K.; Ash, A.J.; Herrero, M. Grazing Systems Expansion and Intensification: Drivers, Dynamics, and Trade-Offs. Glob. Food Secur. 2018, 16, 93–105. [Google Scholar] [CrossRef]
- D’Odorico, P.; Okin, G.S.; Bestelmeyer, B.T. A Synthetic Review of Feedbacks and Drivers of Shrub Encroachment in Arid Grasslands. Ecohydrology 2012, 5, 520–530. [Google Scholar] [CrossRef]
- León, R.J.; Aguiar, M.R. El Deterioro Por Uso Pasturil En Estepas Herbáceas Patagónicas [Sheep Grazing Causes Important Disturbances in Western Patagonian Semi-Arid Grasslands]. Phytocoenologia 1985, 13, 181–196. [Google Scholar] [CrossRef]
- López, D.R.; Brizuela, M.A.; Willems, P.; Aguiar, M.R.; Siffredi, G.; Bran, D. Linking Ecosystem Resistance, Resilience, and Stability in Steppes of North Patagonia. Ecol. Indic. 2013, 24, 1–11. [Google Scholar] [CrossRef]
- Nota, G.; Svensk, M.; Barberis, D.; Frund, D.; Pagani, R.; Pittarello, M.; Probo, M.; Ravetto Enri, S.; Lonati, M.; Lombardi, G. Foraging Behavior of Highland Cattle in Silvopastoral Systems in the Alps. Agrofor. Syst. 2024, 98, 491–505. [Google Scholar] [CrossRef]
- Villagra, E.S.; Pelliza, A.; Willems, P.; Siffredi, G.; Bonvissuto, G.; Villagra, E.S.; Pelliza, A.; Willems, P.; Siffredi, G.; Bonvissuto, G. What Do Domestic Livestock Eat in Northern Patagonian Rangelands? Anim. Prod. Sci. 2013, 53, 360–367. [Google Scholar] [CrossRef]
- Ambrosino, M.L.; Torres, Y.A.; Lucero, C.T.; Lorda, G.S.; Ithurrart, L.S.; Martínez, J.M.; Armando, L.V.; Garayalde, A.; Busso, C.A. Impacts of Shrubs on Soil Quality in the Native Monte Rangelands of Southwestern Buenos Aires, Argentina. Land Degrad. Dev. 2023, 34, 3406–3417. [Google Scholar] [CrossRef]
- Bisigato, A.J.; Bertiller, M.B. Grazing Effects on Patchy Dryland Vegetation in Northern Patagonia. J. Arid Environ. 1997, 36, 639–653. [Google Scholar] [CrossRef]
- Viglizzo, E.F.; Ricard, M.F.; Taboada, M.A.; Vázquez-Amábile, G. Reassessing the Role of Grazing Lands in Carbon-Balance Estimations: Meta-Analysis and Review. Sci. Total Environ. 2019, 661, 531–542. [Google Scholar] [CrossRef]
- Kröpfl, A.I.; Cecchi, G.A.; Villasuso, N.M.; Distel, R.A. Degradation and Recovery Processes in Semi-Arid Patchy Rangelands of Northern Patagonia, Argentina. Land Degrad. Dev. 2011, 24, 393–399. [Google Scholar] [CrossRef]
- Khalil, M.I.; Francaviglia, R.; Henry, B.; Klumpp, K.; Koncz, P.; Llorente, M.; Madari, B.E.; Muñoz-Rojas, M.; Nerger, R. Strategic Management of Grazing Grassland Systems to Maintain and Increase Organic Carbon in Soils. In CO2 Sequestration; IntechOpen: London, UK, 2019; pp. 45–64. ISBN 978-1-83962-993-8. [Google Scholar] [CrossRef]
- Wonkka, C.L.; West, J.B.; Twidwell, D.; Rogers, W.E. Grass Mortality and Turnover Following Core Rangeland Restoration Practices. Rangel. Ecol. Manag. 2017, 70, 290–300. [Google Scholar] [CrossRef]
- López, D.R.; Cavallero, L.; Willems, P.; Bestelmeyer, B.; Brizuela, M. Degradation Influences Equilibrium and non-Equilibrium Dynamics in Rangelands: Implications in Resilience and Stability. Appl. Veg. Sci. 2022, 25, e12670. [Google Scholar] [CrossRef]
- Lin, Y.; Hong, M.; Han, G.; Zhao, M.; Bai, Y.; Chang, S.X. Grazing Intensity Affected Spatial Patterns of Vegetation and Soil Fertility in a Desert Steppe. Agric. Ecosyst. Environ. 2010, 138, 282–292. [Google Scholar] [CrossRef]
- Biancari, L.; Oñatibia, G.R.; Gaitán, J.J.; Aguiar, M.R. Coexistence of Grasses and Shrubs in Patagonian Steppes. Norm or Exception? J. Veg. Sci. 2023, 34, e13177. [Google Scholar] [CrossRef]
- López, D.R.; Cavallero, L.; Brizuela, M.A.; Aguiar, M.R. Ecosystemic Structural-Functional Approach of the State and Transition Model. Appl. Veg. Sci. 2011, 14, 6–16. [Google Scholar] [CrossRef]
- Howard, K.S.C.; Eldridge, D.J.; Soliveres, S. Positive Effects of Shrubs on Plant Species Diversity Do Not Change along a Gradient in Grazing Pressure in an Arid Shrubland. Basic Appl. Ecol. 2012, 13, 159–168. [Google Scholar] [CrossRef]
- Noble, J.C.; Walker, P. Integrated Shrub Management in Semi-Arid Woodlands of Eastern Australia: A Systems-Based Decision Support Model. Agric. Syst. 2006, 88, 332–359. [Google Scholar] [CrossRef]
- Adema, E.O.; Buschiazzo, D.E.; Babinec, F.J.; Rucci, T.E.; Gomez Hermida, V.F. Mechanical Control of Shrubs in a Semiarid Region of Argentina and Its Effect on Soil Water Content and Grassland Productivity. Agric. Water Manag. 2004, 68, 185–194. [Google Scholar] [CrossRef]
- Rago, M.M.; Urretavizcaya, M. Acumulación de Biomasa En Juveniles de Arbustos Nativos Expuestos a Distintos Niveles de Luz Asociados a Esquemas de Manejo Forestal de Coníferas Exóticas. Ecol. Austral 2023, 33, 198–210. [Google Scholar] [CrossRef]
- Velázquez-Martí, B. Situación de Los Sistemas de Aprovechamiento de Los Residuos Forestales Para Su Utilización Energética. Ecosistemas 2006, 15, 77–86. [Google Scholar]
- Kimiti, D.W.; Riginos, C.; Belnap, J. Low-Cost Grass Restoration Using Erosion Barriers in a Degraded African Rangeland. Restor. Ecol. 2017, 25, 376–384. [Google Scholar] [CrossRef]
- Kowaljow, E.; Rostagno, C.M. Enramado y Riego Como Alternativas de Rehabilitación de Regiones Semiáridas Afectadas Por El Tendido de Ductos. Ecol. Austral 2013, 23, 62–69. [Google Scholar] [CrossRef]
- del Valle, H.F.; Labraga, J.C.; Goergen, J. Biozonas de La Región Patagónica. In Evaluación del Estado Actual de la Desertificación en Áreas Representativas de la Patagonia; Proyecto INTA-GTZ (LUDEPA-SME): Río Gallegos, Argentina, 1995; pp. 37–55. [Google Scholar]
- Veblen, T.T.; Lorenz, D.C. Recent Vegetation Changes along the Forest/Steppe Ecotone of Northern Patagonia. Ann. Assoc. Am. Geogr. 1988, 78, 93–111. [Google Scholar] [CrossRef]
- Paruelo, J.; Beltrán, A.B.; Jobbágy, E.; Sala, O.E.; Golluscio, R. The Climate of Patagonia: General Patterns and Controls on Biotic Processes. Ecol. Austral 1998, 8, 85–101. [Google Scholar]
- Bertiller, M.B.; Defose, G.E. Grazing and Plant Growth Interactions in a Semiarid Festuca Pallescens Grassland (Patagonia). Rangel. Ecol. Manag./J. Range Manag. Arch. 1990, 43, 300–303. [Google Scholar] [CrossRef]
- Anchorena, J.; Cingolani, A.M. Identifying Habitat Types in a Disturbed Area of the Forest-Steppe Ecotone of Patagonia. Plant Ecol. 2002, 158, 97–112. [Google Scholar] [CrossRef]
- Roig, F.A. La Vegetación de La Patagonia. In Flora Patagónica; Correa, M.N., Ed.; INTA: Buenos Aires, Argentina, 1998; Volume I, pp. 48–166. [Google Scholar]
- Leon, R.J.C.; Bran, D.; Collantes, M.; Paruelo, J.M.; Soriano, A. Grandes Unidades de Vegetación de La Patagonia Extra Andina. Ecología Austral 1998, 8, 125–144. [Google Scholar]
- Irisarri, J. La Propuesta de Reclasificación de Los Andepts de Argentina, de Acuerdo al Orden Andisoles. In Proceedings of the Workshop Soil Taxonomy; Instituto Nacional de Tecnología Agropecuaria, AICET, Asociación Argentina de la Ciencia del Suelo: Rafaela, Argentina, 2000; pp. 18–27. [Google Scholar]
- Borrelli, P.; Oliva, G.E. Efectos de Los Animales Sobre Los Pastizales. In Ganadería Ovina Sustentable en la Patagonia Austral; INTA Ediciones: Buenos Aires, Argentina, 2001; pp. 99–128. [Google Scholar]
- Paruelo, J.M.; Pütz, S.; Weber, G.; Bertiller, M.B.; Golluscio, R.A.; Aguiar, M.R.; Wiegand, T. Long-Term Dynamics of a Semiarid Grass Steppe under Stochastic Climate and Different Grazing Regimes: A Simulation Analysis. J. Arid Environ. 2008, 72, 2211–2231. [Google Scholar] [CrossRef]
- Willis, B. El Norte de La Patagonia: Naturaleza y Riquezas; Scribner Press: New York, NY, USA, 1914; Volume 1. [Google Scholar]
- INTA Censo Ganadero Chubut 1937–2002. Available online: https://sipas.inta.gob.ar/modulos/info-estrategica/Estad%C3%ADsticas%20y%20censos/CENSOS%20GANADEROS/CENSO%20GANADERA%20CHUBUT%201937-2002.pdf (accessed on 31 October 2019).
- Rusch, V.E.; Cavallero, L.; López, D.R. El Modelo de Estados y Transiciones Como Herramienta Para La Aplicación de La Ley de Bosques. Patagon. For. 2016, XXI, 20–27. [Google Scholar]
- Bottaro, H.; Ejarque, M.; Perez Centeno, M.; Preda, G. Cambios En La Producción Agropecuaria de Chubut En El Período Intercensal 2002–2018; INTA: Buenos Aires, Argentina, 2021; pp. 322–339. ISBN 978-987-47691-2-1. [Google Scholar]
- Irisarri, J.; Mendía, J.; Roca, C.; Buduba, C.G.; Valenzuela, F.; Epele, F.; Fraseto, F.; Ostertag, G.; Bobadilla, S.; Andenmatten, E.; et al. Zonificación de Las Tierras Para La Aptitud Forestal de La Provincia Del Chubut; Dirección General de Bosques y Parques de la Provincia del Chubut: Rawson, Argentina, 1995. [Google Scholar]
- Scarnecchia, D.; Gaskins, C. Modeling Animal-Unit-Equivalents for Beef Cattle. Agric. Syst. 1987, 23, 19–26. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Golluscio, R.A.; Guerschman, J.P.; Cesa, A.; Jouve, V.V.; Garbulsky, M.F. Regional Scale Relationships between Ecosystem Structure and Functioning: The Case of the Patagonian Steppes. Glob. Ecol. Biogeogr. 2004, 13, 385–395. [Google Scholar] [CrossRef]
- Milchunas, D.G.; Lauenroth, W.K. Quantitative Effects of Grazing on Vegetation and Soils over a Global Range of Environments. Ecol. Monogr. 1993, 63, 327–366. [Google Scholar] [CrossRef]
- Passera, C.B.; Borsetto, O.; Candia, R.J.; Stasi, C.R. Shrub Control and Seeding Influences on Grazing Capacity in Argentina. J. Range Manag. 1992, 45, 480–482. [Google Scholar] [CrossRef]
- Loguercio, G.A.; Jovanovski, A.; Molina, J.; Pantaenius, P.; Ríos, F.; Masucheli, M.; Oses, C.; MohrBell, D. Residuos de Biomasa de Forestaciones y Aserraderos de la Región Andina de las Provincias de Neuquén y Chubut: Evaluación Preliminar de Oferta. Available online: http://www.ciefap.org.ar/digitalizado/Publ_tec_34_Informe_final_residuos_biomasa_provincias_Neuquen_Chubut.pdf (accessed on 11 April 2024).
- Laclau, P. Biomass and Carbon Sequestration of Ponderosa Pine Plantations and Native Cypress Forests in Northwest Patagonia. For. Ecol. Manag. 2003, 180, 317–333. [Google Scholar] [CrossRef]
- Goodall, D.W. Some Considerations in the Use of Point Quadrats for the Analysis of Vegetation. Aust. J. Biol. Sci. 1952, 5, 1–41. [Google Scholar] [CrossRef]
- Funk, F.A.; Peter, G.; Loydi, A.; Kröpfl, A.I.; Distel, R.A. Recuperación Estructural y Funcional de Los Espacios Entre Arbustos al Cabo de 10 Años de Exclusión Del Pastoreo En Una Estepa Semiárida Del Noreste de La Patagonia. Ecol. Austral 2012, 22, 195–202. [Google Scholar] [CrossRef]
- IRAM-SAGyP NORMA 29571-1; Calidad Ambiental—Calidad Del Suelo. Determinación de Materia Orgánica En Suelos. Parte 1-Método de Pérdida de Masa Por Calcinación. IRAM: Buenos Aires, Argentina, 2008.
- Chepil, W.S. Field Structure of Cultivated Soils with Special Reference to Erodibility by Wind. Soil Sci. Soc. Am. J. 1953, 17, 185–190. [Google Scholar] [CrossRef]
- Arriaga, F.J.; Lowery, B.; Mays, M.D. A Fast Method for Determining Soil Particle Size Distribution Using a Laser Instrument. Soil Sci. 2006, 171, 663–674. [Google Scholar] [CrossRef]
- Van den Brink, P.J.; ter Braak, C.J.F. Principal Response Curves: Analysis of Time-Dependent Multivariate Responses of Biological Community to Stress. Environ. Toxicol. Chem. 1999, 18, 138–148. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Šmilauer, P. Topics in Constrained and Unconstrained Ordination. Plant Ecol. 2015, 216, 683–696. [Google Scholar] [CrossRef]
- Buckley, H.L.; Day, N.J.; Case, B.S.; Lear, G. Measuring Change in Biological Communities: Multivariate Analysis Approaches for Temporal Datasets with Low Sample Size. PeerJ 2021, 9, e11096. [Google Scholar] [CrossRef] [PubMed]
- Auber, A.; Travers-Trolet, M.; Villanueva, M.C.; Ernande, B. A New Application of Principal Response Curves for Summarizing Abrupt and Cyclic Shifts of Communities over Space. Ecosphere 2017, 8, e02023. [Google Scholar] [CrossRef]
- De Cáceres, M.; Coll, L.; Legendre, P.; Allen, R.B.; Wiser, S.K.; Fortin, M.; Condit, R.; Hubbell, S. Trajectory Analysis in Community Ecology. Ecol. Monogr. 2019, 89, e01350. [Google Scholar] [CrossRef]
- Minchin, P.R. An Evaluation of the Relative Robustness of Techniques for Ecological Ordination. In Theory and Models in Vegetation Science; Prentice, I.C., Van Der Maarel, E., Eds.; Springer: Dordrecht, The Netherlands, 1987; pp. 89–107. ISBN 978-94-010-8303-4. [Google Scholar]
- Campanella, M.V.; Rostagno, C.M.; Videla, L.S.; Bisigato, A.J. Land Degradation Affects Shrub Growth Responses to Precipitation in a Semiarid Rangeland of North-Eastern Patagonia (Argentina). Austral Ecol. 2018, 43, 280–287. [Google Scholar] [CrossRef]
- Cesa, A.; Paruelo, J.M. Changes in Vegetation Structure Induced by Domestic Grazing in Patagonia (Southern Argentina). J. Arid Environ. 2011, 75, 1129–1135. [Google Scholar] [CrossRef]
- Bertiller, M.B.; Defossé, G.E. Estepas Graminosas de Festuca Pallescens En El SW Del Chubut. In Secuencias de Deterioro en Distintos Ambientes Patagónicos. Su Caracterización Mediante el Modelo de Estados y Transiciones; Paruelo, J.M., Bertiller, M.B., Schlichter, T.M., Coronato, F., Eds.; Convenio Argentino-Alemán de Cooperación Técnica, INTA-GTZ, LUDEPA-SME: San Carlos de Bariloche, Argentina, 1993; pp. 14–22. [Google Scholar]
- Vogel, B.; Rostagno, C.M.; Molina, L.; Antilef, M.; La Manna, L. Cushion Shrubs Encroach Subhumid Rangelands and Form Fertility Islands along a Grazing Gradient in Patagonia. Plant Soil 2022, 475, 623–643. [Google Scholar] [CrossRef]
- Fagundes, M.; Weisser, W.; Ganade, G. The Role of Nurse Successional Stages on Species-Specific Facilitation in Drylands: Nurse Traits and Facilitation Skills. Ecol. Evol. 2018, 8, 5173–5184. [Google Scholar] [CrossRef]
- Maestre, F.T.; Escudero, A. Is the Patch Size Distribution of Vegetation a Suitable Indicator of Deserti Cation Processes? Ecology 2009, 90, 1729–1735. [Google Scholar] [CrossRef]
- Gaitán, J.J.; Bran, D.; Oliva, G.E.; Maestre, F.T.; Aguiar, M.R.; Jobbágy, E.; Buono, G.; Ferrante, D.; Nakamatsu, V.; Ciari, G.; et al. Plant Species Richness and Shrub Cover Attenuate Drought Effects on Ecosystem Functioning across Patagonian Rangelands. Biol. Lett. 2014, 10, 20140673. [Google Scholar] [CrossRef]
- Verón, S.R.; Paruelo, J.M. Desertification Alters the Response of Vegetation to Changes in Precipitation. J. Appl. Ecol. 2010, 47, 1233–1241. [Google Scholar] [CrossRef]
- Nosetto, M.D.; Jobbágy, E.G.; Paruelo, J.M. Carbon Sequestration in Semi-Arid Rangelands: Comparison of Pinus Ponderosa Plantations and Grazing Exclusion in NW Patagonia. J. Arid Environ. 2006, 67, 142–156. [Google Scholar] [CrossRef]
- Carrera, A.L.; Bertiller, M.B. Relationships among Plant Litter, Fine Roots, and Soil Organic C and N across an Aridity Gradient in Northern Patagonia, Argentina. Ecoscience 2010, 17, 276–286. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.; Fu, B.; Zhao, W.; Wang, S. Vegetation Responses and Trade-Offs with Soil-Related Ecosystem Services after Shrub Removal: A Meta-Analysis. Land Degrad. Dev. 2019, 30, 1219–1228. [Google Scholar] [CrossRef]
- Gaujour, E.; Amiaud, B.; Mignolet, C.; Plantureux, S. Factors and Processes Affecting Plant Biodiversity in Permanent Grasslands. A Review. Agron. Sustain. Dev. 2012, 32, 133–160. [Google Scholar] [CrossRef]
- Pittarello, M.; Probo, M.; Lonati, M.; Lombardi, G. Restoration of Sub-Alpine Shrub-Encroached Grasslands through Pastoral Practices: Effects on Vegetation Structure and Botanical Composition. Appl. Veg. Sci. 2016, 19, 381–390. [Google Scholar] [CrossRef]
- Romero Ovalle, P.E.; Bisigato, A.J.; Campanella, M.V. Soil Erosion Facilitates Shrub Encroachment in Patagonian Herbaceous Steppes. Land Degrad. Dev. 2021, 32, 3377–3385. [Google Scholar] [CrossRef]
- Colazo, J.C.; Carfagno, P.; Gvozdenovich, J.; Buschiazzo, D. Soil Erosion. In The Soils of Argentina; Rubio, G., Lavado, R.S., Pereyra, F.X., Eds.; World Soils Book Series; Springer International Publishing: Cham, Switzerland, 2019; pp. 239–250. ISBN 978-3-319-76853-3. [Google Scholar]
- Buschiazzo, D.E.; Panebianco, J.E.; Guevara, G.; Rojas, J.; Zurita, J.J.; Bran, D.E.; López, D.R.; Gaitán, J.J.; Hurtado, P. Incidencia Potencial de La Erosión Eólica Sobre La Degradación Del Suelo y La Calidad Del Aire En Distintas Regiones de La Argentina. Cienc Suelo Argent. 2009, 27, 255–260. [Google Scholar]
- Kiem, R.; Kögel-Knabner, I. Refractory Organic Carbon in Particle-Size Fractions of Arable Soils II: Organic Carbon in Relation to Mineral Surface Area and Iron Oxides in Fractions <6 Μm. Org. Geochem. 2002, 33, 1699–1713. [Google Scholar] [CrossRef]
- Cambardella, C.A.; Elliott, E.T. Methods for Physical Separation and Characterization of Soil Organic Matter Fractions. In Soil Structure/Soil Biota Interrelationships; Brussaard, L., Kooistra, M.J., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 449–457. ISBN 978-0-444-81490-6. [Google Scholar]
- La Manna, L.; Buduba, C.G.; Rostagno, C.M. Soil Erodibility and Quality of Volcanic Soils as Affected by Pine Plantations in Degraded Rangelands of NW Patagonia. Eur. J. For. Res. 2016, 135, 643–655. [Google Scholar] [CrossRef]
- McBride, S.G.; Levi, E.M.; Nelson, J.A.; Archer, S.R.; Barnes, P.W.; Throop, H.L.; Predick, K.; McCulley, R.L. Soil-Litter Mixing Mediates Drivers of Dryland Decomposition along a Continuum of Biotic and Abiotic Factors. Ecosystems 2023, 26, 1349–1366. [Google Scholar] [CrossRef]
- Naeth, M.A.; Bailey, A.W.; Pluth, D.J.; Chanasyk, D.S.; Hardin, R.T. Grazing Impacts on Litter and Soil Organic Matter in Mixed Prairie and Fescue Grassland Ecosystems of Alberta. J. Range Manag. 1991, 44, 712. [Google Scholar] [CrossRef]
- Oñatibia, G.R.; Aguiar, M.R. Continuous Moderate Grazing Management Promotes Biomass Production in Patagonian Arid Rangelands. J. Arid. Environ. 2016, 125, 73–79. [Google Scholar] [CrossRef]


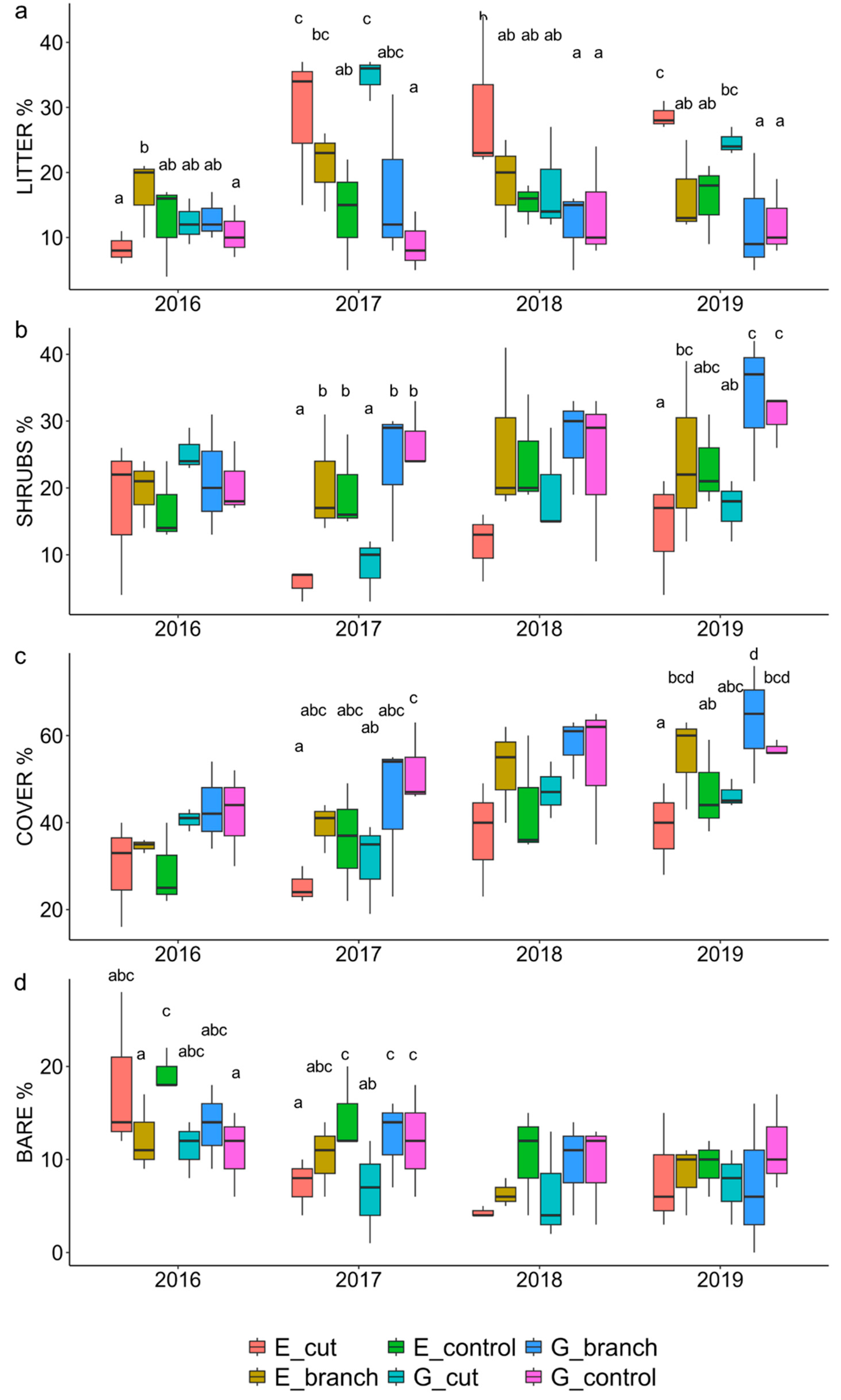
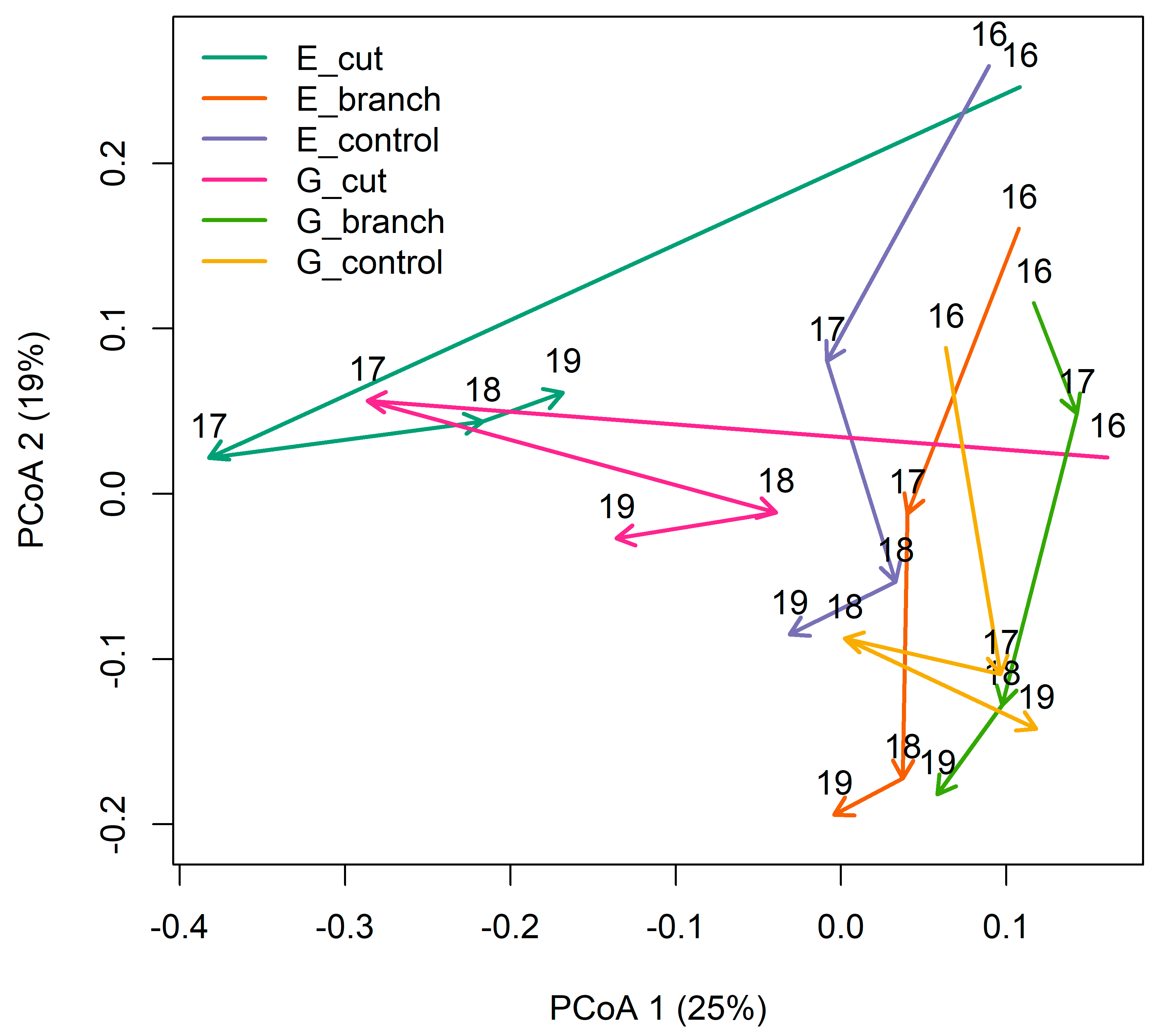
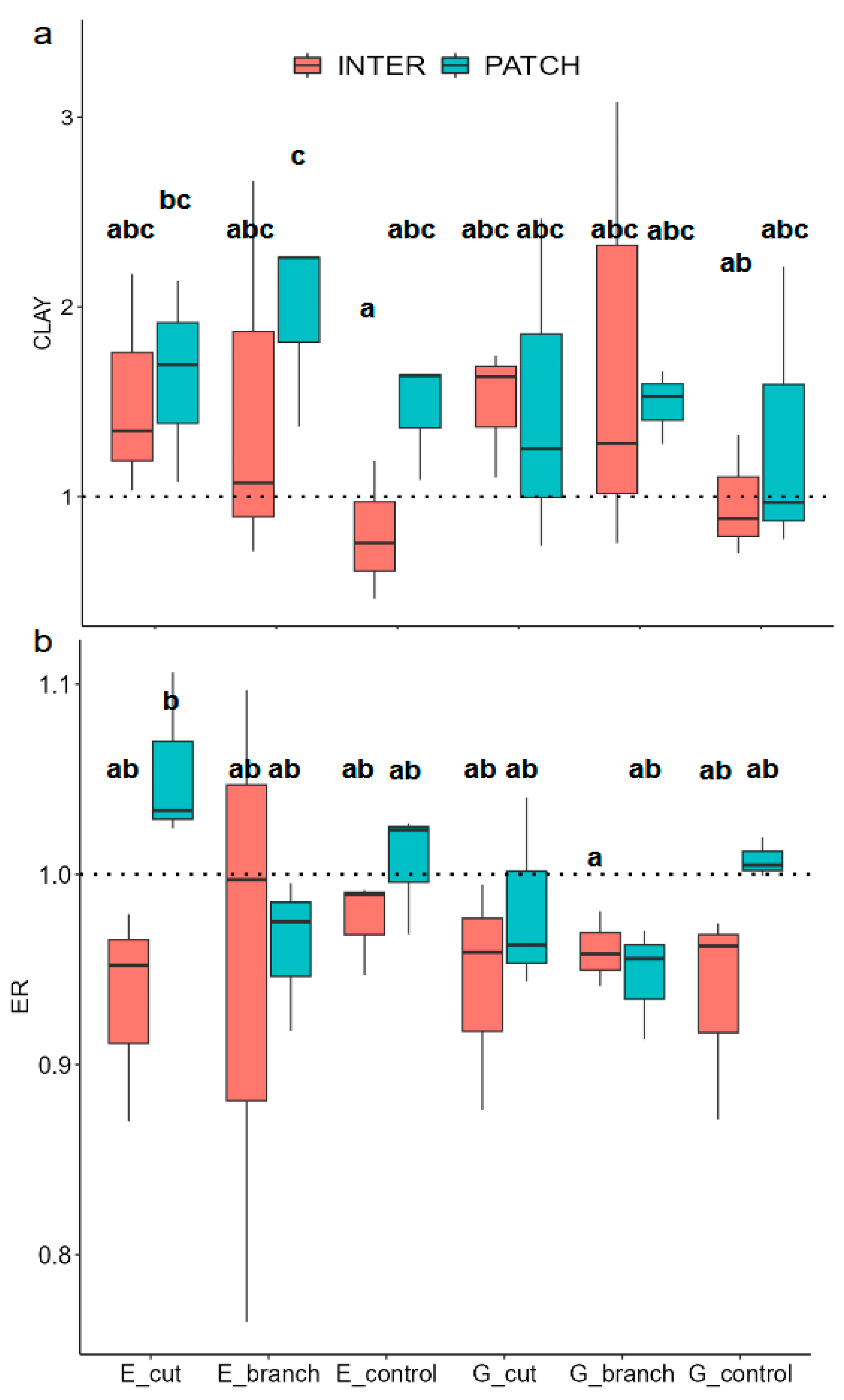
| 2019 | 2018 | 2017 | 2016 | |
|---|---|---|---|---|
| PPT (mm) | 588.6 | 595.2 | 613.5 | 383.3 |
| Stock (AU/ha) | 0.13 | 0.00 | 0.13 | 0.13 |
| GI | 0.61 | 0.00 | 0.15 | 0.77 |
| Condition | Mean | n | s.e | |
|---|---|---|---|---|
| Grazed | 1.42 | 9 | 0.22 | a |
| Enclosed | 2.14 | 9 | 0.22 | b |
| Intervention | 2016 | 2019 | Mean | |
|---|---|---|---|---|
| control_INTER | 4.20 | 3.99 | 0.95 | a |
| cut_INTER | 3.81 | 4.42 | 1.16 | a b |
| cut_PATCH | 4.01 | 5.09 | 1.27 | b |
| branch_PATCH | 3.66 | 5.16 | 1.41 | b |
| control_PATCH | 3.50 | 5.18 | 1.48 | b |
| branch_INTER | 3.08 | 4.32 | 1.40 | b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vogel, B.; Molina, L.; Rostagno, C.M.; La Manna, L. Effects of Grazing and Shrub Management on Species Composition and Soil Properties in Patagonian Grasslands. Grasses 2024, 3, 205-220. https://doi.org/10.3390/grasses3030015
Vogel B, Molina L, Rostagno CM, La Manna L. Effects of Grazing and Shrub Management on Species Composition and Soil Properties in Patagonian Grasslands. Grasses. 2024; 3(3):205-220. https://doi.org/10.3390/grasses3030015
Chicago/Turabian StyleVogel, Braian, Lucia Molina, César M. Rostagno, and Ludmila La Manna. 2024. "Effects of Grazing and Shrub Management on Species Composition and Soil Properties in Patagonian Grasslands" Grasses 3, no. 3: 205-220. https://doi.org/10.3390/grasses3030015
APA StyleVogel, B., Molina, L., Rostagno, C. M., & La Manna, L. (2024). Effects of Grazing and Shrub Management on Species Composition and Soil Properties in Patagonian Grasslands. Grasses, 3(3), 205-220. https://doi.org/10.3390/grasses3030015