
Beef Cattle Grazing Native Grasslands May Follow Three Different Supplement Response Patterns

 ,
,  , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Database Compilation
2.2. Herbage Dry Matter Intake and Estimations of Substitution Rate
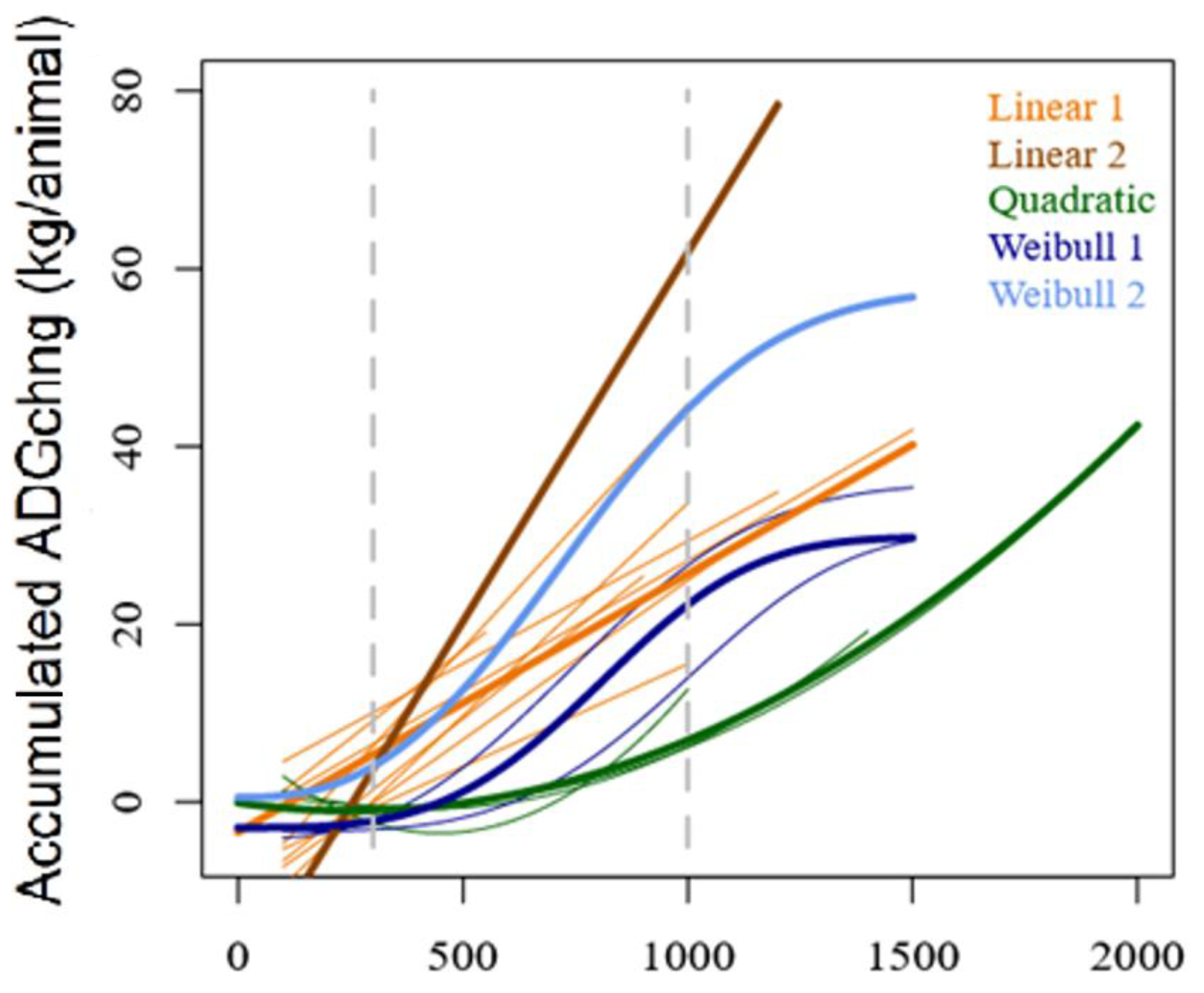
2.3. Phase Identification and the Change in ADGchng vs. Growing Degree Days
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial ID | Location, Year | Supplementation Treatments * | Breed and Category | Duration (Days) | Replicates | Animals/Trial | Source |
|---|---|---|---|---|---|---|---|
| 1 * | La Magnolia, 2013 | TMR with fibre | Braford male calves | 97 | 2 | 40 | [2] |
| 2 * | Glencoe, 2013 | TMR with fibre | Hereford male calves | 120 | 2 | 40 | [2] |
| 3 * | La Magnolia, 2014 | RB (ground and pelleted) | Braford male calves | 68 | 2 | 40 | [38] |
| 4 * | Glencoe, 2014 | RB (ground and pelleted) | Hereford male calves | 108 | 2 | 50 | [38] |
| 5 * | Glencoe, 2015 | Various (maize, expellers and RB) | Hereford male calves | 141 | 2 | 50 | [38] |
| 6 * | Glencoe, 2009 | RB | Hereford male and female calves | 113 | 2 | 48 | [39] |
| 7 * | Glencoe, 2010 | RB | Hereford male calves | 111 | 2 | 48 | [39] |
| 8 * | Glencoe, 2011 | RB | Hereford male calves | 119 | 2 | 48 | [39] |
| 14 * | Ptas del Chuy, 2011 | TMR | British crossbred male calves | 81 | 2 | 48 | [40] |
| 16 * | Glencoe, 2007 | RB | British crossbred male calves | 98 | 2 | 24 | [39] |
| 17 | Palo a Pique, 2012 | TMR with fibre | British crossbred male calves | 77 | 1 | 12 | [41] |
| 24 * | Glencoe, 2004 | Various (RB and expeller) | British crossbred steers | 42 | 2 | 70 | [42] |
| 25 * | Glencoe, 2004 | Various (RB and expeller) | British crossbred steers | 78 | 2 | 70 | [42] |
| 27 | Palo a Pique, 2008 | TMR | British crossbred male calves | 77 | 1 | 56 | [43] |
| 28 * | Cañada del Pueblo, 2008 | DDGS | Hereford female calves | 89 | 2 | 40 | [44] |
| 29 * | Tomás Gomensoro, 2008 | DDGS | British crossbred male calves | 84 | 2 | 40 | [44] |
| Total | 1156 |
Appendix B
| Variable | Min | Mean | Min | SD | CV |
|---|---|---|---|---|---|
| HDMI 45%DMD (kg/animal/d) | 2.47 | 6.48 | 10.30 | 1.69 | 26 |
| HDMI 55%DMD (kg/animal/d) | 1.88 | 4.94 | 7.85 | 1.21 | 24 |
| HDMI 65%DMD (kg/animal/d) | 1.43 | 3.90 | 6.07 | 0.94 | 24 |
| HDMI 45%DMD (%BW) | 0.00 | 2.90 | 5.39 | 1.04 | 36 |
| HDMI 55%DMD (%BW) | 0.00 | 2.21 | 3.69 | 0.75 | 34 |
| HDMI 65%DMD (%BW) | 0.00 | 1.74 | 2.77 | 0.58 | 34 |
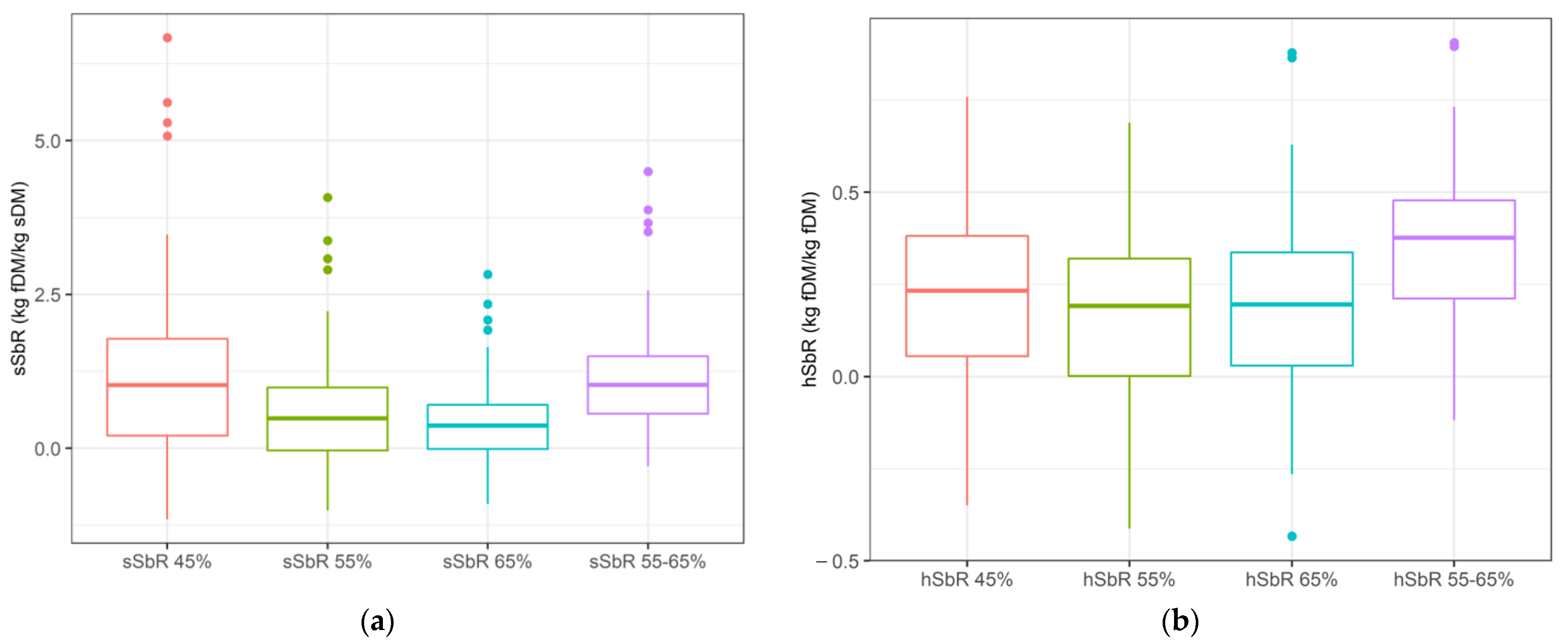
| sSbR 45% | −1.16 | 1.06 | 5.62 | 1.26 | 119 |
| sSbR 55% | −1.01 | 0.50 | 3.38 | 0.81 | 162 |
| sSbR 65% | −0.90 | 0.32 | 2.35 | 0.59 | 185 |
| sSbR 55–65% | −0.29 | 1.09 | 3.88 | 0.76 | 70 |
| hSbR 45% | −0.35 | 0.20 | 0.67 | 0.21 | 108 |
| hSbR 55% | −0.41 | 0.13 | 0.61 | 0.21 | 160 |
| hSbR 65% | −0.43 | 0.12 | 0.60 | 0.22 | 179 |
| hSbR 55–65% | −0.12 | 0.31 | 0.71 | 0.18 | 56 |
References
- Simeone, A.; Beretta, V.; Blasina, M.; Piñeirúa, A.; Renau, M. Winter Response of Weaned Beef Calves to Self-fed Supplementation on Native Pastures. Proc. Aust. Soc. Anim. Prod. 2010, 28, 5. [Google Scholar]
- Cazzuli, F.; Lagomarsino, X.; Boggiano, P.; Saadoun, A.; Montossi, F. Self-feeding improved animal performance of calves grazing native grasslands during winter on extensive livestock production systems. Agro. Sur. 2018, 46, 29–39. [Google Scholar] [CrossRef]
- Cazzuli, F.; De Barbieri, I.; Jaurena, M.; Silveira, C.; Montossi, F. Restricted oats grazing and its frequency of allocation affects calves’ performance on native grasslands. Arch. Zootec. 2019, 68, 220–227. [Google Scholar] [CrossRef]
- Grigera, G.; Oesterheld, M.; Pacín, F. Monitoring forage production for farmers’ decision making. Agric. Syst. 2007, 94, 637–648. [Google Scholar] [CrossRef]
- De Figueiredo, D.M.; De Oliveira, A.S.; Sales, M.F.L.; Paulino, M.F.; Do Vale SM, L.R. Economic analysis of four supplementation strategies for growth and finish beef cattle in a pasture-supplement system. Rev. Bras. Zootec. 2007, 36, 1443–1453. [Google Scholar]
- Cazzuli, F.; Sánchez, J.; Hirigoyen, A.; Rovira, P.; Beretta, V.; Simeone, A.; Jaurena, M.; Durante, M.; Savian, J.V.; Poppi, D.; et al. Supplement feed efficiency of growing beef cattle grazing native Campos grasslands during winter: A collated analysis. Transl. Anim. Sci. 2023, 7, txad028. [Google Scholar] [CrossRef]
- Gekara, O.J.; Prigge, E.C.; Bryan, W.B.; Nestor, E.L.; Seidel, G. Influence of sward height, daily timing of concentrate supplementation, and restricted time for grazing on forage utilization by lactating beef cows. J. Anim. Sci. 2005, 83, 1435–1444. [Google Scholar] [CrossRef]
- Krysl, L.J.; Hess, B.W. Influence of Supplementation on Behaviour of Grazing Cattle. J. Anim. Sci. 1993, 71, 2546–2555. [Google Scholar] [CrossRef]
- Bailey, D.W.; Jensen, D. Method of Supplementation May Affect Cattle Grazing Patterns. Rangel. Ecol. Manag. 2008, 61, 131–135. [Google Scholar] [CrossRef]
- Palma, M.N.; Reis, W.L.; Rodrigues, J.P.; Silva, T.E.; Franco, M.O.; Rennó, L.N.; Detmann, E. Strategies of energy supplementation for cattle fed tropical forage and infrequently supplemented with protein. Anim. Feed. Sci. Technol. 2023, 297, 115599. [Google Scholar] [CrossRef]
- Costa, D.F.A.; Correia, P.S.; Dorea, J.R.R.; De Souza, J.; Congio, G.F.D.S.; Pires, A.V.; Malafaia, P.A.M.; Drouillard, J.; Dias, C.T.D.S.; Luchiari-Filho, A.; et al. Strategic supplementation of growing cattle on tropical pastures improves nutrient use and animal performance, with fewer days required on the finishing phase. Anim. Prod. Sci. 2021, 61, 480. [Google Scholar] [CrossRef]
- Detmann, E.; Valente, E.; Batista, E.D.; Huhtanen, P. An evaluation of the performance and efficiency of nitrogen utilization in cattle fed tropical grass pastures with supplementation. Livest. Sci. 2014, 162, 141–153. [Google Scholar] [CrossRef]
- Caton, J.S.; Dhuyvetter, D.V. Influence of energy supplementation on grazing ruminants: Requirements and responses. J. Anim. Sci. 1997, 75, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Nkrumah, J.D.; Li, C.; Basarab, J.A.; Goonewardene, L.A.; Okine, E.K.; Crews, D.H.; Moore, S.S. Test duration for growth, feed intake, and feed efficiency in beef cattle using the GrowSafe System. J. Anim. Sci. 2006, 84, 2289–2298. [Google Scholar] [CrossRef]
- Stafford, S.D.; Cochran, R.C.; VanZant, E.S.; O Fritz, J. Evaluation of the potential of supplements to substitute for low-quality, tallgrass-prairie forage. J. Anim. Sci. 1996, 74, 639–647. [Google Scholar] [CrossRef]
- Grainger, C.; Mathews, G. Positive relation between substitution rate and pasture allowance for cows receiving concentrates. Aust. J. Exp. Agric. 1989, 29, 355–360. [Google Scholar] [CrossRef]
- Rogers, G.L. Pasture and supplements in the temperate zone. In The Challenge: Efficient Dairy Production; Australian Society of Animal Production: Toowong, Queensland, Australia, 1985; pp. 85–108. [Google Scholar]
- Beever, D.E.; Doyle, P.T. Feed conversion efficiency as a key determinant of dairy herd performance: A review. Aust. J. Exp. Agric. 2007, 47, 645–657. [Google Scholar] [CrossRef]
- Da Trindade, J.K.; Neves, F.P.; Pinto, C.E.; Bremm, C.; Mezzalira, J.C.; Nadin, L.B.; Genro, T.C.; Gonda, H.L.; Carvalho, P.C. Daily Forage Intake by Cattle on Natural Grassland: Response to Forage Allowance and Sward Structure. Rangel. Ecol. Manag. 2016, 69, 59–67. [Google Scholar] [CrossRef]
- CattleExplorer. CattleExplorer Spreadsheet. CSIRO GrazPlan. 2012. Available online: http://www.grazplan.csiro.au/ (accessed on 13 May 2023).
- Osítis, U.; Strikauska, S.; Grundmane, A. Lopbarības Analīžu Rezultātu Apkopojums. LLU SIA Jelgavas Tipogrāfija 2003, 62, 1. [Google Scholar]
- Sparks, A. Nasapower: NASA-POWER Data from R. 2023. Available online: https://cran.r-project.org/web/packages/nasapower/index.html (accessed on 13 May 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.r-project.org/ (accessed on 13 May 2023).
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Versión 2015 [Computer Program] Grupo InfoStat, FCA; Universidad Nacional de Córdoba: Cordoba, Argentina, 2015. [Google Scholar]
- Méndez, M.N.; Chilibroste, P.; Aguerre, M. Pasture dry matter intake per cow in intensive dairy production systems: Effects of grazing and feeding management. Animal 2020, 14, 846–853. [Google Scholar] [CrossRef]
- Kellaway, R.; Harrington, T. Feeding Concentrates: Supplements for Dairy Cows. Sci. Access 2004, 1, 271. [Google Scholar]
- Pérez-Prieto, L.; Delagarde, R. Meta-analysis of the effect of pasture allowance on pasture intake, milk production, and grazing behavior of dairy cows grazing temperate grasslands. J. Dairy Sci. 2013, 96, 6671–6689. [Google Scholar] [CrossRef]
- Penno, J.W.; Macdonald, K.A.; Holmes, C.W.; Davis, S.R.; Wilson, G.F.; Brookes, I.M.; Thom, E.R. Responses to supplementation by dairy cows given low pasture allowances in different seasons 1. Pasture intake and substitution. Anim. Sci. 2006, 82, 661–670. [Google Scholar] [CrossRef][Green Version]
- Dixon, R.M.; Stockdale, C.R. Associative effects between forages and grains: Consequences for feed utilisation. Aust. J. Agric. Res. 1999, 50, 757–774. [Google Scholar] [CrossRef]
- Cazzuli, F.; Bremm, C.; Jaurena, M.; Poppi, D.; Durante, M.; Benvenutti, M.A.; Savian, J.V.; Devincenzi, T.; Rovira, P.; Lagomarsino, X.; et al. The defoliation dynamics of a stockpiled native grassland pasture follow similar patterns between supplemented and unsupplemented beef calves. Grass Forage Sci. 2022, 78, 306–316. [Google Scholar] [CrossRef]
- Bohnert, D.W.; DelCurto, T.; Clark, A.A.; Merrill, M.L.; Falck, S.J.; Harmon, D.L. Protein supplementation of ruminants consuming low-quality cool- or warm-season forage: Differences in intake and digestibility. J. Anim. Sci. 2011, 89, 3707–3717. [Google Scholar] [CrossRef]
- Berretta, J.; Risso, D.; Montossi, F.; Pigurina, G. Campos in Uruguay. In Grassland Ecophysiology and Grazing Ecology; Lemaire, G.F., Hodgon, J., de Moraes, A., Nabinger, C., Carvalho, Eds.; CAB International: Wallingford, UK, 2000; pp. 377–394. [Google Scholar]
- Da Trindade, J.; Pinto, C.; Neves, F.; Mezzalira, J.; Bremm, C.; Genro, T.; Tischler, M.; Nabinger, C.; Gonda, H.; Paulo, C.; et al. Forage Allowance as a Target of Grazing Management: Implications on Grazing Time and Forage Searching. Rangel. Ecol. Manag. 2012, 65, 382–393. [Google Scholar] [CrossRef]
- Mezzalira, J.C.; Carvalho, P.C.D.F.; Da Trindade, J.K.; Bremm, C.; Fonseca, L.; Amaral, M.F.D.; Reffatti, M.V. Produção animal e vegetal em pastagem nativa manejada sob diferentes ofertas de forragem por bovinos. Ciência Rural. 2012, 42, 1264–1270. [Google Scholar] [CrossRef]
- da Cunha, L.L.; Bremm, C.; Savian, J.V.; Zubieta, Á.S.; Rossetto, J.; Carvalho, P.C.d.F. Relevance of sward structure and forage nutrient contents in explaining methane emissions from grazing beef cattle and sheep. Sci. Total Environ. 2023, 869, 161695. [Google Scholar] [CrossRef]
- Lascano, C.E.; Schmidt, A.; Barahona, R. Forage Quality and the Environment. International Grassland Congress, 19 October 2015. 2001, pp. 1–19. Available online: http://www.internationalgrasslands.org/papers/pdfs/tema9_1.pdf (accessed on 13 May 2023).
- Benvenutti, M.A.; Pavetti, D.R.; Poppi, D.P.; Gordon, I.J.; Cangiano, C.A. Defoliation patterns and their implications for the management of vegetative tropical pastures to control intake and diet quality by cattle. Grass Forage Sci. 2015, 71, 424–436. [Google Scholar] [CrossRef]
- Montossi, F. Sistemas de suplementación mediante auto-suministro para la mejora de la recría invernal de terneros sobre campo natural en el norte del Uruguay. In Auto-Suministro y Suplementación Sobre Campo Natural; Montossi, F., Ed.; Serie Técnica 234; INIA: Tacuarembó, Uruguay, 2017; p. 70. [Google Scholar]
- Luzardo, S.; Cuadro, R.; Lagomarsino, X.; Montossi, F.; Brito, G.; La Manna, A. Tecnologías para la intensificación de la recría en el basalto—Uso estratégico de suplementación sobre campo natural y pasturas mejoradas. In Alternativas Tecnológicas Para los Sistemas Ganaderos de Basalto; Berretta, E.J., Montossi, F., Brito, G., Eds.; Serie Técnica 217; INIA: Tacuarembó, Uruguay, 2014; pp. 71–91. [Google Scholar] [CrossRef]
- Esteves, M.; Laxalde, S.; Nario, M. Utilización de Nitrógeno no Proteico en Programas de Suplementación Invernal Basados en Autoconsumo Para Terneros Pastoreando Campo Nativo. Bachelor’s Thesis, FAgro/UdelaR, Montevideo, Uruguay, 2013; p. 119. [Google Scholar]
- Rovira, P.; Velazco, J.; Montossi, F. Desempeño productivo de terneros sobre campo natural suplementados con grano húmedo de sorgo y distintos niveles de nitrógeno de liberación lenta (rumenfeed). In Suplementación de Bovinos con Grano Húmedo de Sorgo y Fuentes Proteicas Sobre Campo Natural; Rovira, P., Ed.; INIA: Montevideo, Uruguay, 2014; pp. 47–58. [Google Scholar]
- Arrieta, G.; Lagomarsino, X.; Olivera, J.; Trindade, G. Incidencia de Diferentes Dietas Sobre el Crecimiento Animal, el Rendimiento Carnicero y la Calidad de Carne. Bachelor’s Thesis, FAgro/UdelaR, Montevideo, Uruguay, 2008; p. 218. [Google Scholar]
- Rovira, P.; Velazco, J.I. Evaluación de un sistema de autoconsumo restringido con distinto contenido de sal en la ración en terneros suplementados sobre campo natural. In Suplementación de Bovinos en Pastoreo: Autoconsumo; Rovira, P., Velazco, J., Eds.; Serie Técnica 199; INIA: Montevideo, Uruguay, 2012; pp. 23–31. [Google Scholar]
- Berretta, V.; Simeone, A.; Castro, G.; Ferrés, M.; Legorburu, G.; Victorica, M. DDGS, campo natural y comederos de autoconsumo: Una alianza estratégica para mejorar la recría en ganado de carne. In Jor- Nada Anual de la Unidad de Producción Intensiva de Carne: Pradera, Campo Natural, Suplementación y Corral: Un Medio Campo Para Ganar el Partido de la Rentabilidad; Simeone, A., Berretta, V., Eds.; UPIC/FAgro/UdelaR: Paysandú, Uruguay, 2019; pp. 32–43. [Google Scholar]





| Variable | Mean | SD | CV | Min | Max |
|---|---|---|---|---|---|
| cADG, kg/animal/day | 0.19 | 0.15 | 79 | −0.02 | 0.52 |
| sADG, kg/animal/day | 0.52 | 0.23 | 45 | 0.07 | 1.24 |
| ADGchng, kg/animal/day | 0.34 | 0.18 | 52 | −0.10 | 1.02 |
| Shrunk average BW, kg | 196 | 36 | 18 | 145 | 282 |
| Stocking rate, kg BW/ha | 428 | 95 | 22 | 239 | 630 |
| Forage allowance (FA), kg DM/kg BW | 5.5 | 3.4 | 63 | 1.1 | 19.1 |
| Green forage allowance, kg green DM/kg BW | 2.2 | 1.4 | 63 | 0.68 | 6.5 |
| Green FA/FA | 0.4 | 0.2 | 42 | 0.2 | 0.9 |
| Sward height, cm | 8.1 | 4.2 | 52 | 2.5 | 19.0 |
| Herbage biomass, kg DM/ha | 2157 | 1062 | 49 | 461 | 6163 |
| Herbage DM, % | 55.1 | 8.7 | 16 | 23.7 | 73.3 |
| Green Herbage, % of DM | 43.0 | 17.4 | 41 | 14.6 | 86.7 |
| Herbage CP, % of DM | 8.3 | 2.4 | 29 | 4.3 | 16.6 |
| Herbage ADF, % of DM | 43.4 | 8.2 | 19 | 23.7 | 59.1 |
| Herbage NDF, % of DM | 64.8 | 9.1 | 14 | 30.7 | 81.1 |
| Herbage energy, MJ ME/kg DM | 8.6 | 1.3 | 15 | 6.5 | 12.1 |
| Herbage DM digestibility, % | 55.3 | 6.5 | 12 | 42.9 | 70.5 |
| Forage ME/CP, 100 MJ/kg CP | 1.1 | 0.2 | 19 | 0.7 | 1.6 |
| Supplementation rate, %BW | 0.8 | 0.3 | 35 | 0.4 | 2.0 |
| Supplement DM intake, kg/animal/day | 1.7 | 0.6 | 34 | 1.1 | 3.9 |
| SFE, ADGchng/kg DM of supplement | 0.20 | 0.08 | 40 | −0.06 | 0.35 |
| SFE CP, ADGchng/kg CP of supplement | 1.11 | 0.55 | 50 | −0.26 | 2.40 |
| SFE ME, ADGchng/MJ of supplement | 0.02 | 0.01 | 40 | −0.00 | 0.03 |
| Supplement ME content, MJ/kg of DM | 11.5 | 0.7 | 6 | 10.1 | 12.8 |
| Supplement CP content, % of DM | 20.2 | 9.2 | 46 | 12.1 | 43.9 |
| Supplement ME/CP, 100 MJ/kg CP | 0.5 | 0.2 | 33 | 0.2 | 0.8 |
| Stocking period, day | 96.0 | 27.1 | 28 | 42.0 | 141.0 |
| Correlation between SFE and | Spearman | p-Value |
|---|---|---|
| HDMI 45%DMD (kg/animal/d) | 0.44 | <0.001 |
| HDMI 55%DMD (kg/animal/d) | 0.46 | <0.001 |
| HDMI 65%DMD (kg/animal/d) | 0.48 | <0.001 |
| HDMI 45%DMD (%BW) | 0.40 | <0.001 |
| HDMI 55%DMD (%BW) | 0.46 | <0.001 |
| HDMI 65%DMD (%BW) | 0.50 | <0.001 |
| Correlation between SFE and | Spearman | p-Value |
|---|---|---|
| sSbR 45% | −0.72 | <0.001 |
| sSbR 55% | −0.76 | <0.001 |
| sSbR 65% | −0.79 | <0.001 |
| sSbR 55–65% | −0.72 | <0.001 |
| hSbR 45% | −0.76 | <0.001 |
| hSbR 55% | −0.78 | <0.001 |
| hSbR 65% | −0.79 | <0.001 |
| hSbR 55–65% | −0.78 | <0.001 |
| Multiple Linear Regression Equation | R2 | Adjusted R2 | p-Value |
|---|---|---|---|
| ADGchng300 = −22.21 + 0.092 × BW + 3.66 × sCP:sME | 0.73 | 0.69 | <0.01 |
| ADGchng1000 = 17.90 × sDMintake + 43.51 × sADG + 45.3 × fCP:fME − 7.01 × fCP | 0.85 | 0.79 | <0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cazzuli, F.; Durante, M.; Hirigoyen, A.; Sánchez, J.; Rovira, P.; Beretta, V.; Simeone, A.; Jaurena, M.; Savian, J.V.; Poppi, D.; et al. Beef Cattle Grazing Native Grasslands May Follow Three Different Supplement Response Patterns. Grasses 2023, 2, 168-184. https://doi.org/10.3390/grasses2030014
Cazzuli F, Durante M, Hirigoyen A, Sánchez J, Rovira P, Beretta V, Simeone A, Jaurena M, Savian JV, Poppi D, et al. Beef Cattle Grazing Native Grasslands May Follow Three Different Supplement Response Patterns. Grasses. 2023; 2(3):168-184. https://doi.org/10.3390/grasses2030014
Chicago/Turabian StyleCazzuli, Fiorella, Martín Durante, Andrés Hirigoyen, Javier Sánchez, Pablo Rovira, Virginia Beretta, Alvaro Simeone, Martín Jaurena, Jean Victor Savian, Dennis Poppi, and et al. 2023. "Beef Cattle Grazing Native Grasslands May Follow Three Different Supplement Response Patterns" Grasses 2, no. 3: 168-184. https://doi.org/10.3390/grasses2030014
APA StyleCazzuli, F., Durante, M., Hirigoyen, A., Sánchez, J., Rovira, P., Beretta, V., Simeone, A., Jaurena, M., Savian, J. V., Poppi, D., Montossi, F., Lagomarsino, X., Luzardo, S., Brito, G., Velazco, J. I., Lattanzi, F. A., & Bremm, C. (2023). Beef Cattle Grazing Native Grasslands May Follow Three Different Supplement Response Patterns. Grasses, 2(3), 168-184. https://doi.org/10.3390/grasses2030014