
Gata3 Insufficiency Accelerates Recanalization of Damaged Lymphatics via Adjusting Collagen Composition

 , , , and
, , , and
Abstract
1. Introduction
2. Results
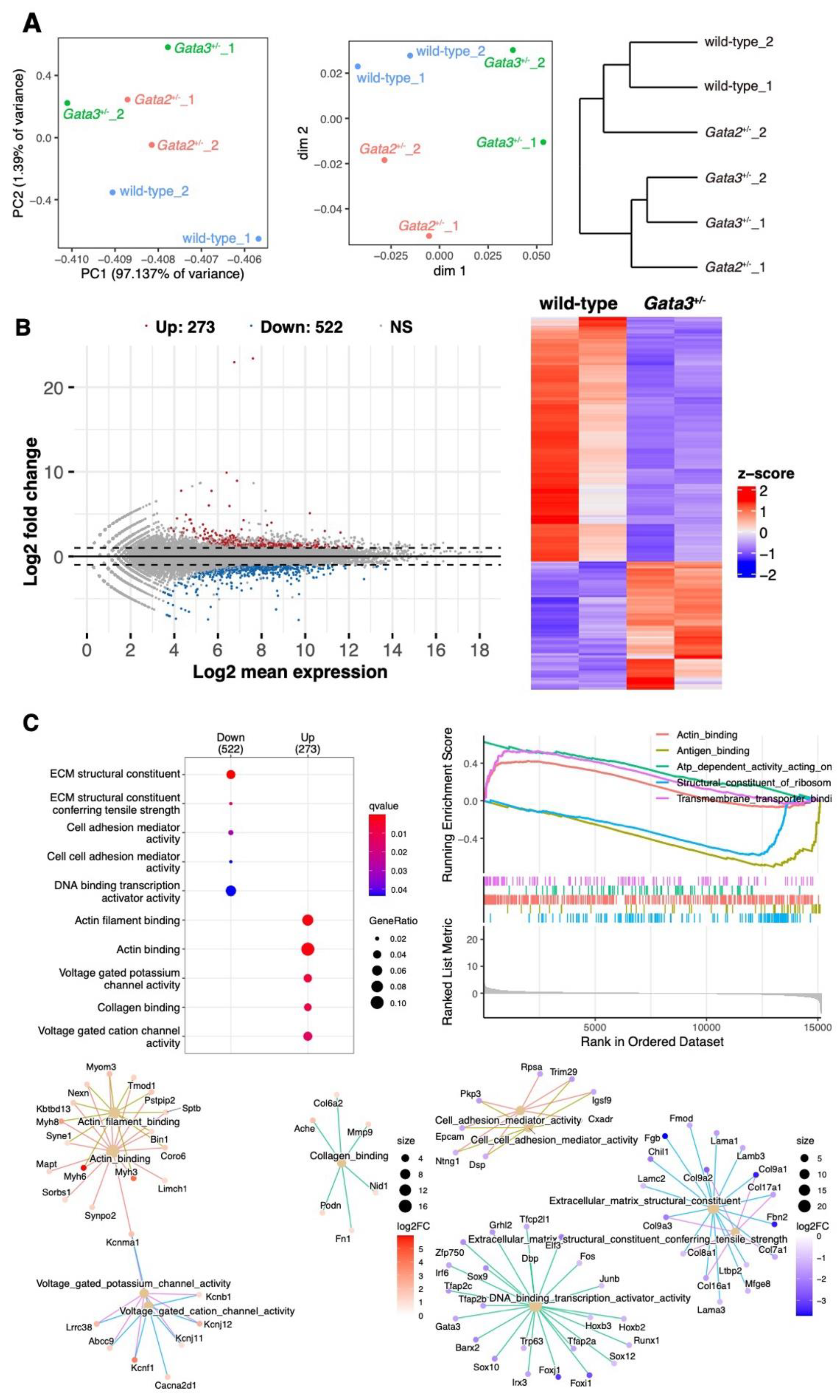
2.1. Different Gene Expression Patterns in the Subcutaneous Tissue of the Popliteal Region of Gata3+/− Mice
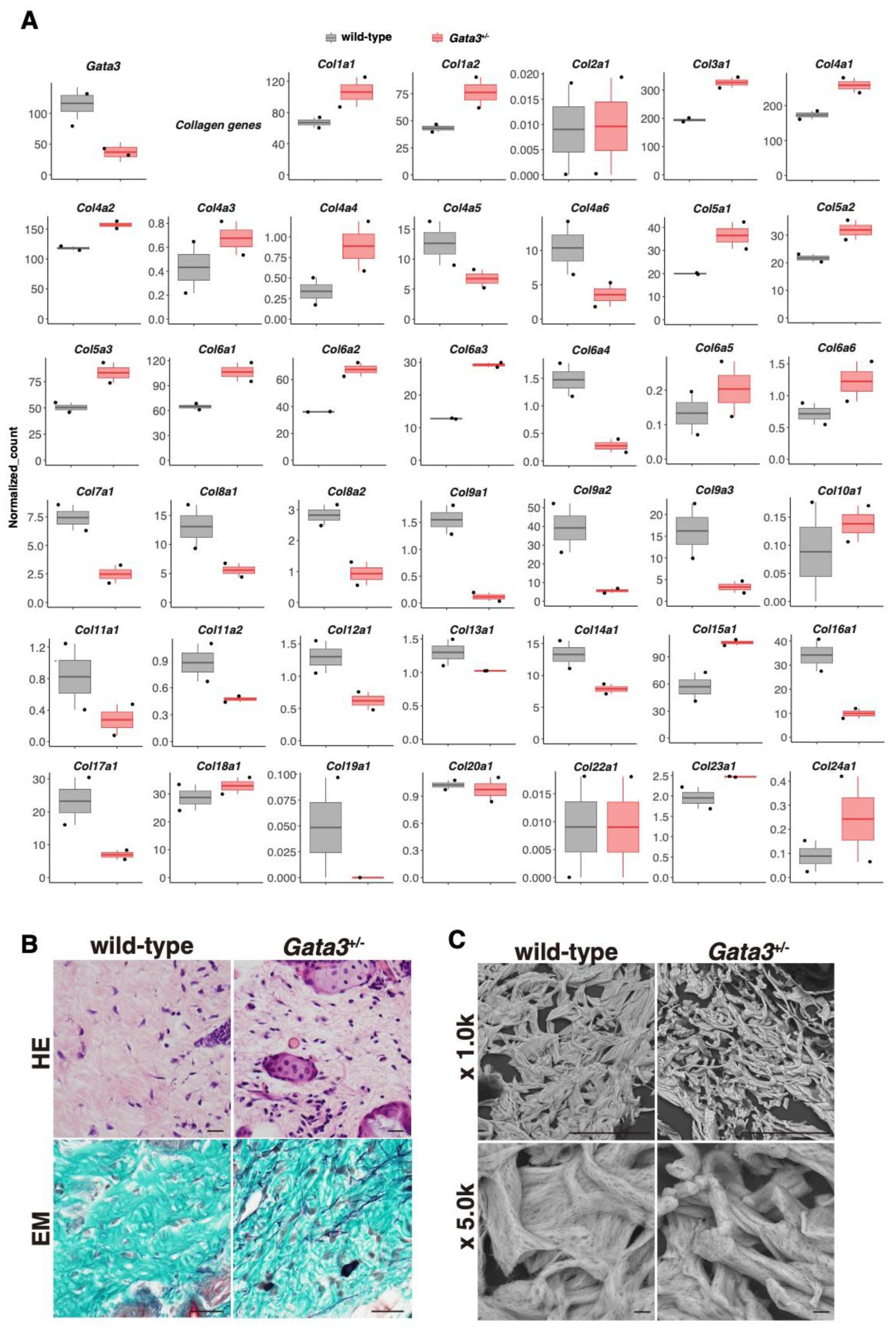
2.2. Altered Expression of the Collagen Gene Cluster and the Presence of Thin Dermal Collagen Fibers in Gata3+/− Skin
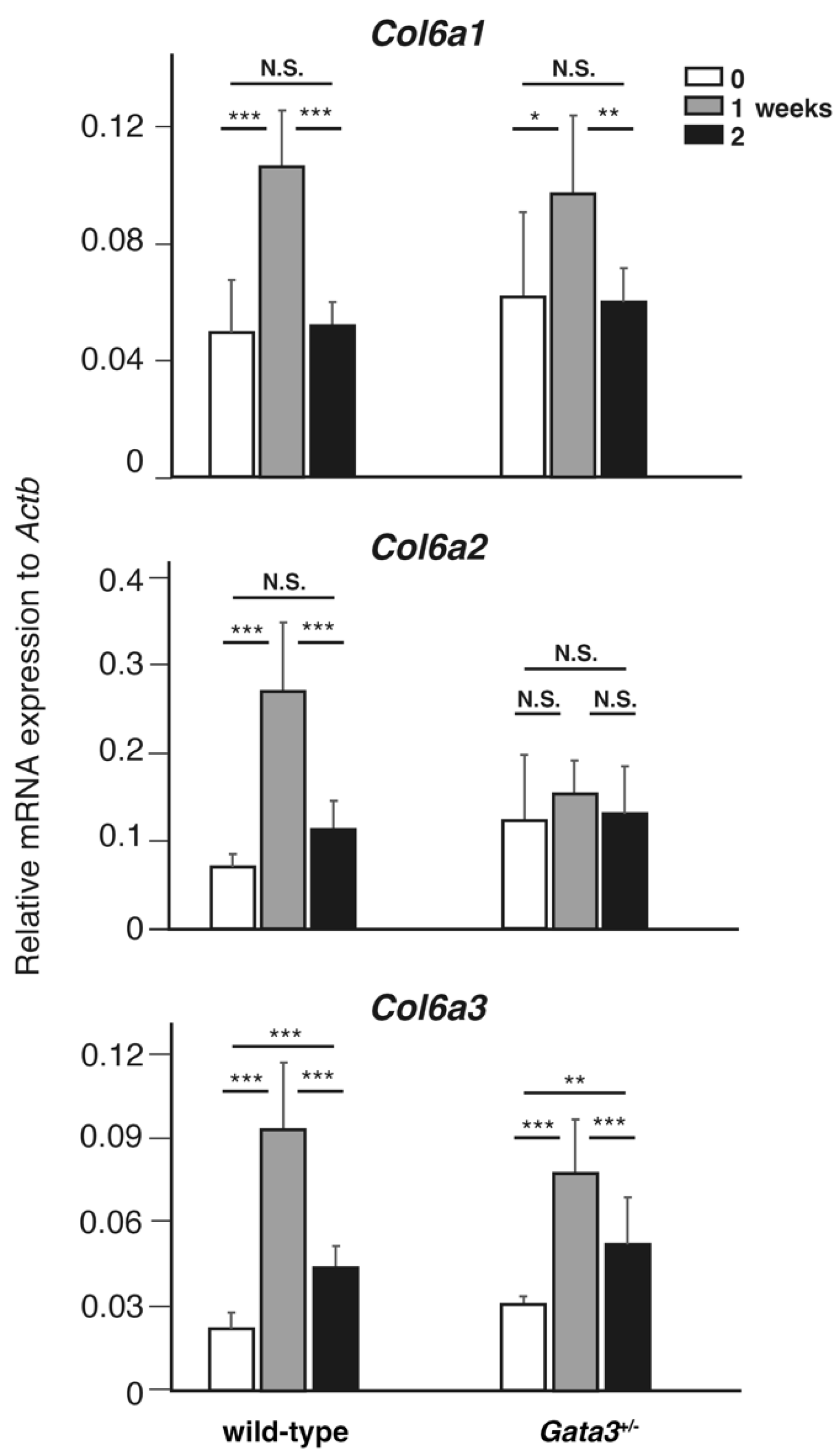
2.3. Transient Upregulation of Col6a Genes During Lymphatic Vessel Recanalization
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Popliteal Lymph Node Extirpation
4.3. RNA-Seq and Bioinformatic Analysis
4.4. Histology
4.5. Scanning Electron Microscopy (SEM)
4.6. cDNA Synthesis and RT-PCR Analysis
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grada, A.A.; Phillips, T.J. Lymphedema: Pathophysiology and clinical manifestations. J. Am. Acad. Dermatol. 2017, 77, 1009–1020. [Google Scholar] [CrossRef] [PubMed]
- Rockson, S.G. Advances in Lymphedema. Circ. Res. 2021, 128, 2003–2016. [Google Scholar] [CrossRef] [PubMed]
- Kataru, R.P.; Wiser, I.; Baik, J.E.; Park, H.J.; Rehal, S.; Shin, J.Y.; Mehrara, B.J. Fibrosis and secondary lymphedema: Chicken or egg? Transl. Res. 2019, 209, 68–76. [Google Scholar] [CrossRef]
- Brown, S.; Dayan, J.H.; Kataru, R.P.; Mehrara, B.J. The Vicious Circle of Stasis, Inflammation, and Fibrosis in Lymphedema. Plast. Reconstr. Surg. 2023, 151, 330e–341e. [Google Scholar] [CrossRef] [PubMed]
- Kazenwadel, J.; Betterman, K.L.; Chong, C.E.; Stokes, P.H.; Lee, Y.K.; Secker, G.A.; Agalarov, Y.; Demir, C.S.; Lawrence, D.M.; Sutton, D.L.; et al. GATA2 is required for lymphatic vessel valve development and maintenance. J. Clin. Investig. 2015, 125, 2979–2994. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ostergaard, P.; Simpson, M.A.; Connell, F.C.; Steward, C.G.; Brice, G.; Woollard, W.J.; Dafou, D.; Kilo, T.; Smithson, S.; Lunt, P.; et al. Mutations in GATA2 cause primary lymphedema associated with a predisposition to acute myeloid leukemia (Emberger syndrome). Nat. Genet. 2011, 43, 929–931. [Google Scholar] [CrossRef] [PubMed]
- Polat, A.; Dinulescu, M.; Fraitag, S.; Nimubona, S.; Toutain, F.; Jouneau, S.; Poullot, E.; Droitcourt, C.; Dupuy, A. Skin manifestations among GATA2-deficient patients. Br. J. Dermatol. 2018, 178, 781–785. [Google Scholar] [CrossRef] [PubMed]
- Watanabe-Asaka, T.; Hayashi, M.; Uemura, S.; Takai, J.; Suzuki, A.; Moriguchi, T.; Kawai, Y. GATA2 participates in the recanalization of lymphatic vessels after surgical lymph node extirpation. Genes Cells 2021, 26, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Watanabe-Asaka, T.; Hayashi, M.; Harada, T.; Uemura, S.; Takai, J.; Nakamura, Y.; Moriguchi, T.; Kawai, Y. Perturbed collagen metabolism underlies lymphatic recanalization failure in Gata2 heterozygous deficient mice. J. Biochem. 2024, 175, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Ho, I.C.; Tai, T.S.; Pai, S.Y. GATA3 and the T-cell lineage: Essential functions before and after T-helper-2-cell differentiation. Nat. Rev. Immunol. 2009, 9, 125–135. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yagi, R.; Zhu, J.; Paul, W.E. An updated view on transcription factor GATA3-mediated regulation of Th1 and Th2 cell differentiation. Int. Immunol. 2011, 23, 415–420. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bovay, E.; Sabine, A.; Prat-Luri, B.; Kim, S.; Son, K.; Willrodt, A.H.; Olsson, C.; Halin, C.; Kiefer, F.; Betsholtz, C.; et al. Multiple roles of lymphatic vessels in peripheral lymph node development. J. Exp. Med. 2018, 215, 2760–2777. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yang, Y.; Oliver, G. Development of the mammalian lymphatic vasculature. J. Clin. Investig. 2014, 124, 888–897. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Frye, M.; Taddei, A.; Dierkes, C.; Martinez-Corral, I.; Fielden, M.; Ortsäter, H.; Kazenwadel, J.; Calado, D.P.; Ostergaard, P.; Salminen, M.; et al. Matrix stiffness controls lymphatic vessel formation through regulation of a GATA2-dependent transcriptional program. Nat. Commun. 2018, 9, 1511. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ross, R. The fibroblast and wound repair. Biol. Rev. Camb. Philos. Soc. 1968, 43, 51–96. [Google Scholar] [CrossRef] [PubMed]
- Knoedler, S.; Broichhausen, S.; Guo, R.; Dai, R.; Knoedler, L.; Kauke-Navarro, M.; Diatta, F.; Pomahac, B.; Machens, H.G.; Jiang, D.; et al. Fibroblasts—The cellular choreographers of wound healing. Front. Immunol. 2023, 14, 1233800. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- van der Rest, M.; Mayne, R.; Ninomiya, Y.; Seidah, N.G.; Chretien, M.; Olsen, B.R. The structure of type IX collagen. J. Biol. Chem. 1985, 260, 220–225. [Google Scholar] [CrossRef] [PubMed]
- van der Rest, M.; Dublet, B.; Champliaud, M.F. Fibril-associated collagens. Biomaterials 1990, 11, 28–31. [Google Scholar] [PubMed]
- Sun, B. The mechanics of fibrillar collagen extracellular matrix. Cell Rep. Phys. Sci. 2021, 2, 100515. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- He, Y.; Sardar, S.; Bay-Jensen, A.C.; Port, H.; Karsdal, M.A. Chapter 9—Type IX collagen. In Biochemistry of Collagens, Laminins and Elastin, 3rd ed.; Karsdal, M.A., Ed.; Academic Press: Cambridge, MA, USA, 2023; pp. 89–95. ISBN 9780443156175. [Google Scholar] [CrossRef]
- Port, H.; He, Y.; Karsdal, M.A.; Madsen, E.A.; Bay-Jensen, A.C.; Willumsen, N.; Holm Nielsen, S. Type IX Collagen Turnover Is Altered in Patients with Solid Tumors. Cancers 2024, 16, 2035. [Google Scholar] [CrossRef]
- Myllyharju, J.; Kivirikko, K.I. Collagens and collagen-related diseases. Ann. Med. 2001, 33, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Arseni, L.; Lombardi, A.; Orioli, D. From Structure to Phenotype: Impact of Collagen Alterations on Human Health. Int. J. Mol. Sci. 2018, 19, 1407. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nielsen, S.H.; Sardar, S.; Karsdal, M.A.; Henriksen, K. Chapter 36—Collagen diseases. In Biochemistry of Collagens, Laminins and Elastin, 3rd ed.; Karsdal, M.A., Ed.; Academic Press: Cambridge, MA, USA, 2023; pp. 371–398. ISBN 9780443156175. [Google Scholar] [CrossRef]
- Salles Rosa Neto, N.; Pereira, I.A.; Sztajnbok, F.R.; Azevedo, V.F. Unraveling the genetic collagen connection: Clinical and therapeutic insights on genetic connective tissue disorders. Adv. Rheumatol. 2024, 64, 32. [Google Scholar] [CrossRef] [PubMed]
- Malfait, F.; Francomano, C.; Byers, P.; Belmont, J.; Berglund, B.; Black, J.; Bloom, L.; Bowen, J.M.; Brady, A.F.; Burrows, N.P.; et al. The 2017 international classification of the Ehlers-Danlos syndromes. Am. J. Med. Genet. C Semin. Med. Genet. 2017, 175, 8–26. [Google Scholar] [CrossRef] [PubMed]
- Malfait, F.; Castori, M.; Francomano, C.A.; Giunta, C.; Kosho, T.; Byers, P.H. The Ehlers-Danlos syndromes. Nat. Rev. Dis. Primers 2020, 6, 64. [Google Scholar] [CrossRef] [PubMed]
- Nixon, T.R.W.; Alexander, P.; Richards, A.; McNinch, A.; Bearcroft, P.W.P.; Cobben, J.; Snead, M.P. Homozygous Type IX collagen variants (COL9A1, COL9A2, and COL9A3) causing recessive Stickler syndrome-Expanding the phenotype. Am. J. Med. Genet. A 2019, 179, 1498–1506. [Google Scholar] [CrossRef] [PubMed]
- Nixon, T.R.W.; Richards, A.J.; Martin, H.; Alexander, P.; Snead, M.P. Autosomal Recessive Stickler Syndrome. Genes 2022, 13, 1135. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Acke, F.R.E.; De Leenheer, E.M.R. Hearing Loss in Stickler Syndrome: An Update. Genes 2022, 13, 1571. [Google Scholar] [CrossRef]
- Bardhan, A.; Bruckner-Tuderman, L.; Chapple, I.L.C.; Fine, J.D.; Harper, N.; Has, C.; Magin, T.M.; Marinkovich, M.P.; Marshall, J.F.; McGrath, J.A.; et al. Epidermolysis bullosa. Nat. Rev. Dis. Primers 2020, 6, 78. [Google Scholar] [CrossRef] [PubMed]
- South, A.P.; Laimer, M.; Gueye, M.; Sui, J.Y.; Eichenfield, L.F.; Mellerio, J.E.; Nyström, A. Type VII Collagen Deficiency in the Oncogenesis of Cutaneous Squamous Cell Carcinoma in Dystrophic Epidermolysis Bullosa. J. Investig. Dermatol. 2023, 143, 2108–2119. [Google Scholar] [CrossRef] [PubMed]
- van der Wees, J.; van Looij, M.A.; de Ruiter, M.M.; Elias, H.; van der Burg, H.; Liem, S.S.; Kurek, D.; Engel, J.D.; Karis, A.; van Zanten, B.G.; et al. Hearing loss following Gata3 haploinsufficiency is caused by cochlear disorder. Neurobiol. Dis. 2004, 16, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Baik, J.E.; Park, H.J.; Kataru, R.P.; Savetsky, I.L.; Ly, C.L.; Shin, J.; Encarnacion, E.M.; Cavali, M.R.; Klang, M.G.; Riedel, E.; et al. TGF-β1 mediates pathologic changes of secondary lymphedema by promoting fibrosis and inflammation. Clin. Transl. Med. 2022, 12, e758. [Google Scholar] [CrossRef] [PubMed]
- Sano, M.; Hirakawa, S.; Sasaki, T.; Inuzuka, K.; Katahashi, K.; Kayama, T.; Yamanaka, Y.; Tsuyuki, H.; Endo, Y.; Naruse, E.; et al. Role of Subcutaneous Adipose Tissues in the Pathophysiology of Secondary Lymphedema. Lymphat. Res. Biol. 2022, 20, 593–599. [Google Scholar] [CrossRef]
- Duhon, B.H.; Phan, T.T.; Taylor, S.L.; Crescenzi, R.L.; Rutkowski, J.M. Current Mechanistic Understandings of Lymphedema and Lipedema: Tales of Fluid, Fat, and Fibrosis. Int. J. Mol. Sci. 2022, 23, 6621. [Google Scholar] [CrossRef] [PubMed]
- Hayashida, K.; Yoshida, S.; Yoshimoto, H.; Fujioka, M.; Saijo, H.; Migita, K.; Kumaya, M.; Akita, S. Adipose-Derived Stem Cells and Vascularized Lymph Node Transfers Successfully Treat Mouse Hindlimb Secondary Lymphedema by Early Reconnection of the Lymphatic System and Lymphangiogenesis. Plast. Reconstr. Surg. 2017, 139, 639–651. [Google Scholar] [CrossRef]
- Will, P.A.; Kilian, K.; Bieback, K.; Fricke, F.; Berner, J.E.; Kneser, U.; Hirche, C. Lymphedema-Associated Fibroblasts Are Related to Fibrosis and Stage Progression in Patients and a Murine Microsurgical Model. Plast. Reconstr. Surg. 2024, 154, 688e–700e. [Google Scholar] [CrossRef]
- Son, H.; Lee, S.; Kim, K.; Koo, K.I.; Hwang, C.H. Deep learning-based quantitative estimation of lymphedema-induced fibrosis using three-dimensional computed tomography images. Sci. Rep. 2022, 12, 15371. [Google Scholar] [CrossRef] [PubMed]
- Pandolfi, P.P.; Roth, M.E.; Karis, A.; Leonard, M.W.; Dzierzak, E.; Grosveld, F.G.; Engel, J.D.; Lindenbaum, M.H. Targeted disruption of the GATA3 gene causes severe abnormalities in the nervous system and in fetal liver haematopoiesis. Nat. Genet. 1995, 11, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Moriguchi, T.; Takako, N.; Hamada, M.; Maeda, A.; Fujioka, Y.; Kuroha, T.; Huber, R.E.; Hasegawa, S.L.; Rao, A.; Yamamoto, M.; et al. Gata3 participates in a complex transcriptional feedback network to regulate sympathoadrenal differentiation. Development 2006, 133, 3871–3881. [Google Scholar] [CrossRef] [PubMed]
- Kawai, S.; Takagi, Y.; Kaneko, S.; Kurosawa, T. Effect of three types of mixed anesthetic agents alternate to ketamine in mice. Exp. Anim. 2011, 60, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Cock, P.J.; Fields, C.J.; Goto, N.; Heuer, M.L.; Rice, P.M. The Sanger FASTQ file format for sequences with quality scores, and the Solexa/Illumina FASTQ variants. Nucleic Acids Res. 2010, 38, 1767–1771. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Etoh, K.; Nakao, M. A web-based integrative transcriptome analysis, RNAseqChef, uncovers the cell/tissue type-dependent action of sulforaphane. J. Biol. Chem. 2023, 299, 104810. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yabe, Y.; Hagiwara, Y.; Ando, A.; Tsuchiya, M.; Minowa, T.; Takemura, T.; Honda, M.; Hatori, K.; Sonofuchi, K.; Kanazawa, K.; et al. Chondrogenic and fibrotic process in the ligamentum flavum of patients with lumbar spinal canal stenosis. Spine 2015, 40, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Yabe, Y.; Hagiwara, Y.; Tsuchiya, M.; Honda, M.; Hatori, K.; Sonofuchi, K.; Kanazawa, K.; Koide, M.; Sekiguchi, T.; Itaya, N.; et al. Decreased elastic fibers and increased proteoglycans in the ligamentum flavum of patients with lumbar spinal canal stenosis. J. Orthop. Res. 2016, 34, 1241–1247. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Gene | Sense Primer | Antisense Primer | Assay |
|---|---|---|---|
| Gata3LacZ | TTCGCCAGCTGGCGTAATAGCGAAGAGGC | TAGGTCACGTTGGTGTAGATGGGCGCATCG | genotyping |
| Col6a1 | AACAGGAATAGGAAATGTGACCC | ACACCACGGATAGGTTAGGGG | qPCR |
| Col6a2 | AAGGCCCCATTGGATTCCC | CTCCCTTCCGACCATCCGAT | qPCR |
| Col6a3 | GCTGCGGAATCACTTTGTGC | CACCTTGACACCTTTCTGGGT | qPCR |
| β-actin | AGATCAAGATCATTGCTCCTCCT | ACGCAGCTCAGTAACAGTCC | qPCR |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayashi, M.; Harada, T.; Takai, J.; Uemura, S.; Moriguchi, T.; Watanabe-Asaka, T.; Kawai, Y. Gata3 Insufficiency Accelerates Recanalization of Damaged Lymphatics via Adjusting Collagen Composition. Lymphatics 2025, 3, 7. https://doi.org/10.3390/lymphatics3010007
Hayashi M, Harada T, Takai J, Uemura S, Moriguchi T, Watanabe-Asaka T, Kawai Y. Gata3 Insufficiency Accelerates Recanalization of Damaged Lymphatics via Adjusting Collagen Composition. Lymphatics. 2025; 3(1):7. https://doi.org/10.3390/lymphatics3010007
Chicago/Turabian StyleHayashi, Moyuru, Takuya Harada, Jun Takai, Satoshi Uemura, Takashi Moriguchi, Tomomi Watanabe-Asaka, and Yoshiko Kawai. 2025. "Gata3 Insufficiency Accelerates Recanalization of Damaged Lymphatics via Adjusting Collagen Composition" Lymphatics 3, no. 1: 7. https://doi.org/10.3390/lymphatics3010007
APA StyleHayashi, M., Harada, T., Takai, J., Uemura, S., Moriguchi, T., Watanabe-Asaka, T., & Kawai, Y. (2025). Gata3 Insufficiency Accelerates Recanalization of Damaged Lymphatics via Adjusting Collagen Composition. Lymphatics, 3(1), 7. https://doi.org/10.3390/lymphatics3010007