
Orally Administered N-Oleoyl Alanine Blocks Acute Opioid Withdrawal Induced-Conditioned Place Preference and Attenuates Somatic Withdrawal following Chronic Opioid Exposure in Rats


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drugs
2.3. Apparatus
2.4. Surgery
2.5. Procedures
2.5.1. Experiment 1
2.5.2. Experiment 2
2.6. Data Analysis
3. Results
3.1. Experiment 1
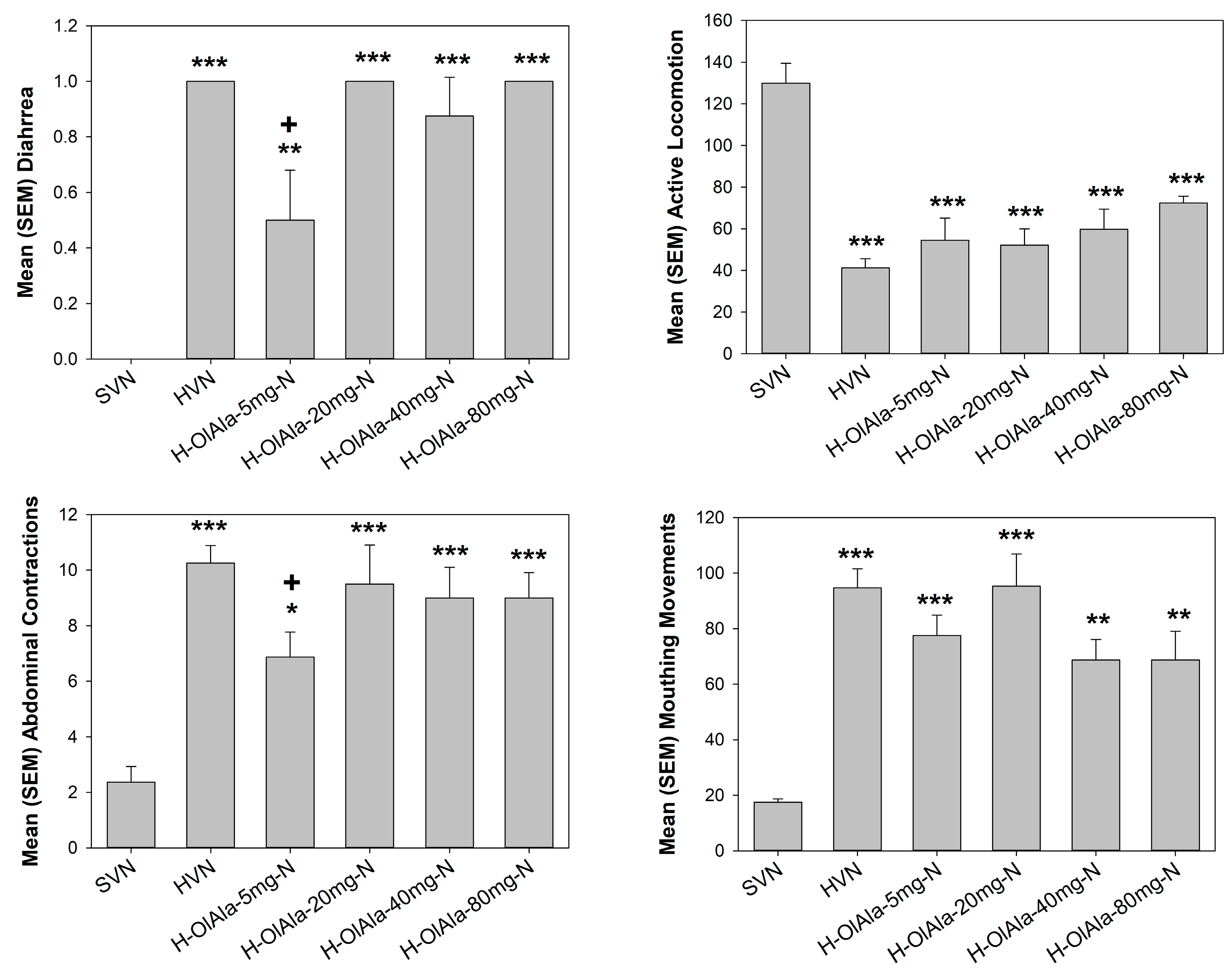
3.2. Experiment 2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burstein, S.H.; McQuain, C.A.; Ross, A.H.; Salmonsen, R.A.; Zurier, R.E. Resolution of inflammation by N-arachidonoylglycine. J. Cell Biochem. 2011, 112, 3227–3233. [Google Scholar] [CrossRef] [PubMed]
- Dludla, P.V.; Nkambule, B.B.; Mazibuko-Mbeje, S.E.; Nyambuya, T.M.; Marcheggiani, F.; Cirilli, I.; Ziqubu, K.; Shabalala, S.C.; Johnson, R.; Louw, J.; et al. N-Acetyl Cysteine Targets Hepatic Lipid Accumulation to Curb Oxidative Stress and Inflammation in NAFLD: A Comprehensive Analysis of the Literature. Antioxidants 2020, 9, 1283. [Google Scholar] [CrossRef] [PubMed]
- Raboune, S.; Stuart, J.M.; Leishman, E.; Takacs, S.M.; Rhodes, B.; Basnet, A.; Jameyfield, E.; McHugh, D.; Widlanski, T.; Bradshaw, H.B. Novel endogenous N-acyl amides activate TRPV1-4 receptors, BV-2 microglia, and are regulated in brain in an acute model of inflammation. Front. Cell Neurosci. 2014, 8, 195. [Google Scholar] [CrossRef] [PubMed]
- Long, J.Z.; Svensson, K.J.; Bateman, L.A.; Lin, H.; Kamenecka, T.; Lokurkar, I.A.; Lou, J.; Rao, R.R.; Chang, M.R.; Jedrychowski, M.P.; et al. The Secreted Enzyme PM20D1 Regulates Lipidated Amino Acid Uncouplers of Mitochondria. Cell 2016, 166, 424–435. [Google Scholar] [CrossRef]
- Wang, S.; Xu, Q.; Shu, G.; Wang, L.; Gao, P.; Xi, Q.; Zhang, Y.; Jiang, Q.; Zhu, X. N-Oleoyl glycine, a lipoamino acid, stimulates adipogenesis associated with activation of CB1 receptor and Akt signaling pathway in 3T3-L1 adipocyte. Biochem. Biophys. Res. Commun. 2015, 466, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.M.; Bisogno, T.; Petros, T.J.; Chang, S.Y.; Zavitsanos, P.A.; Zipkin, R.E.; Sivakumar, R.; Coop, A.; Maeda, D.Y.; De Petrocellis, L.; et al. Identification of a new class of molecules, the arachidonyl amino acids, and characterization of one member that inhibits pain. J. Biol. Chem. 2001, 276, 42639–42644. [Google Scholar] [CrossRef] [PubMed]
- Mostyn, S.N.; Wilson, K.A.; Schumann-Gillett, A.; Frangos, Z.J.; Shimmon, S.; Rawling, T.; Ryan, R.M.; O’Mara, M.L.; Vandenberg, R.J. Identification of an allosteric binding site on the human glycine transporter, GlyT2, for bioactive lipid analgesics. Elife 2019, 8, e47150. [Google Scholar] [CrossRef] [PubMed]
- Vuong, L.A.; Mitchell, V.A.; Vaughan, C.W. Actions of N-arachidonyl-glycine in a rat neuropathic pain model. Neuropharmacology 2008, 54, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, A.; Cipollone, I.; Verde, R.; Kalkan, H.; Moriello, C.; Iannotti, F.A.; Di Marzo, V.; Piscitelli, F. The endocannabinoidome mediator N-oleoylglycine is a novel protective agent against 1-methyl-4-phenyl-pyridinium-induced neurotoxicity. Front. Aging Neurosci. 2022, 14, 926634. [Google Scholar] [CrossRef]
- Donvito, G.; Piscitelli, F.; Muldoon, P.; Jackson, A.; Vitale, R.M.; D’Aniello, E.; Giordano, C.; Ignatowska-Jankowska, B.M.; Mustafa, M.A.; Guida, F.; et al. N-Oleoyl-glycine reduces nicotine reward and withdrawal in mice. Neuropharmacology 2019, 148, 320–331. [Google Scholar] [CrossRef]
- Lin, Q.; Hai, J.; Yao, L.Y.; Lu, Y. Neuroprotective effects of NSTyr on cognitive function and neuronal plasticity in rats of chronic cerebral hypoperfusion. Brain Res. 2010, 1325, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Petrie, G.N.; Wills, K.L.; Piscitelli, F.; Smoum, R.; Limebeer, C.L.; Rock, E.M.; Humphrey, A.E.; Sheppard-Perkins, M.; Lichtman, A.H.; Mechoulam, R.; et al. Oleoyl glycine: Interference with the aversive effects of acute naloxone-precipitated MWD, but not morphine reward, in male Sprague-Dawley rats. Psychopharmacology 2019, 236, 2623–2633. [Google Scholar] [CrossRef] [PubMed]
- Egashira, N.; Shirakawa, A.; Abe, M.; Niki, T.; Mishima, K.; Iwasaki, K.; Oishi, R.; Fujiwara, M. N-acetyl-L-cysteine inhibits marble-burying behavior in mice. J. Pharmacol. Sci. 2012, 119, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, S.M.; Piscitelli, F.; Silvestri, C.; Limebeer, C.L.; Rock, E.M.; Smoum, R.; Farag, M.; de Almeida, H.; Sullivan, M.T.; Lacroix, S.; et al. Spontaneous and Naloxone-Precipitated Withdrawal Behaviors from Chronic Opiates are Accompanied by Changes in N-Oleoylglycine and N-Oleoylalanine Levels in the Brain and Ameliorated by Treatment With These Mediators. Front. Pharmacol. 2021, 12, 706703. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, S.M.; Smoum, R.; Farag, M.; Atwal, H.; Collins, S.A.; Rock, E.M.; Limebeer, C.L.; Piscitelli, F.; Iannotti, F.A.; Lichtman, A.H.; et al. Oleoyl alanine (HU595): A stable monomethylated oleoyl glycine interferes with acute naloxone precipitated morphine withdrawal in male rats. Psychopharmacology 2020, 237, 2753–2765. [Google Scholar] [CrossRef] [PubMed]
- Rock, E.M.; Ayoub, S.M.; Limebeer, C.L.; Gene, A.; Wills, K.L.; DeVuono, M.V.; Smoum, R.; Di Marzo, V.; Lichtman, A.H.; Mechoulam, R.; et al. Acute naloxone-precipitated morphine withdrawal elicits nausea-like somatic behaviors in rats in a manner suppressed by N-oleoylglycine. Psychopharmacology 2020, 237, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Piscitelli, F.; Guida, F.; Luongo, L.; Iannotti, F.A.; Boccella, S.; Verde, R.; Lauritano, A.; Imperatore, R.; Smoum, R.; Cristino, L.; et al. Protective Effects of N-Oleoylglycine in a Mouse Model of Mild Traumatic Brain Injury. ACS Chem. Neurosci. 2020, 11, 1117–1128. [Google Scholar] [CrossRef] [PubMed]
- Shahen-Zoabi, S.; Smoum, R.; Bingor, A.; Grad, E.; Nemirovski, A.; Shekh-Ahmad, T.; Mechoulam, R.; Yaka, R. N-oleoyl glycine and N-oleoyl alanine attenuate alcohol self-administration and preference in mice. Transl. Psychiatry 2023, 13, 273. [Google Scholar] [CrossRef]
- Rock, E.M.; Limebeer, C.L.; Sullivan, M.T.; DeVuono, M.V.; Lichtman, A.H.; Di Marzo, V.; Mechoulam, R.; Parker, L.A. N-Oleoylglycine and N-Oleoylalanine Do Not Modify Tolerance to Nociception, Hyperthermia, and Suppression of Activity Produced by Morphine. Front. Synaptic Neurosci. 2021, 13, 620145. [Google Scholar] [CrossRef]
- Bradshaw, H.B.; Rimmerman, N.; Hu, S.S.; Burstein, S.; Walker, J.M. Novel endogenous N-acyl glycines identification and characterization. Vitam. Horm. 2009, 81, 191–205. [Google Scholar] [CrossRef]
- Abadji, V.; Lin, S.; Taha, G.; Griffin, G.; Stevenson, L.A.; Pertwee, R.G.; Makriyannis, A. (R)-methanandamide: A chiral novel anandamide possessing higher potency and metabolic stability. J. Med. Chem. 1994, 37, 1889–1893. [Google Scholar] [CrossRef] [PubMed]
- Gracy, K.N.; Dankiewicz, L.A.; Koob, G.F. Opiate withdrawal-induced fos immunoreactivity in the rat extended amygdala parallels the development of conditioned place aversion. Neuropsychopharmacology 2001, 24, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Wills, K.L.; Petrie, G.N.; Millett, G.; Limebeer, C.L.; Rock, E.M.; Niphakis, M.J.; Cravatt, B.F.; Parker, L.A. Double Dissociation of Monoacylglycerol Lipase Inhibition and CB1 Antagonism in the Central Amygdala, Basolateral Amygdala, and the Interoceptive Insular Cortex on the Affective Properties of Acute Naloxone-Precipitated Morphine Withdrawal in Rats. Neuropsychopharmacology 2016, 41, 1865–1873. [Google Scholar] [CrossRef] [PubMed]
- Huestis, M.A. Human cannabinoid pharmacokinetics. Chem. Biodivers. 2007, 4, 1770–1804. [Google Scholar] [CrossRef] [PubMed]
- Izzo, A.A.; Sharkey, K.A. Cannabinoids and the gut: New developments and emerging concepts. Pharmacol. Ther. 2010, 126, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Shahen-Zoabi, S.; Smoum, R.; Beiser, T.; Nemirovski, A.; Mechoulam, R.; Yaka, R. N-Oleoyl Glycine and Its Derivatives Attenuate the Acquisition and Expression of Cocaine-Induced Behaviors. Cannabis Cannabinoid Res. 2023, 8, 812–823. [Google Scholar] [CrossRef] [PubMed]
- Dunn, K.E.; Weerts, E.M.; Huhn, A.S.; Schroeder, J.R.; Tompkins, D.A.; Bigelow, G.E.; Strain, E.C. Preliminary evidence of different and clinically meaningful opioid withdrawal phenotypes. Addict. Biol. 2020, 25, e12680. [Google Scholar] [CrossRef] [PubMed]
- Ware, O.D.; Ellis, J.D.; Dunn, K.E.; Hobelmann, J.G.; Finan, P.; Huhn, A.S. The association of chronic pain and opioid withdrawal in men and women with opioid use disorder. Drug Alcohol. Depend. 2022, 240, 109631. [Google Scholar] [CrossRef]
- Bodnar, R.J.; Kest, B. Sex differences in opioid analgesia, hyperalgesia, tolerance and withdrawal: Central mechanisms of action and roles of gonadal hormones. Horm. Behav. 2010, 58, 72–81. [Google Scholar] [CrossRef]
- Bobzean, S.A.M.; Kokane, S.S.; Butler, B.D.; Perrotti, L.I. Sex differences in the expression of morphine withdrawal symptoms and associated activity in the tail of the ventral tegmental area. Neurosci. Lett. 2019, 705, 124–130. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayoub, S.M.; Rock, E.M.; Limebeer, C.L.; DeVuono, M.V.; Parker, L.A. Orally Administered N-Oleoyl Alanine Blocks Acute Opioid Withdrawal Induced-Conditioned Place Preference and Attenuates Somatic Withdrawal following Chronic Opioid Exposure in Rats. Psychoactives 2024, 3, 184-193. https://doi.org/10.3390/psychoactives3020012
Ayoub SM, Rock EM, Limebeer CL, DeVuono MV, Parker LA. Orally Administered N-Oleoyl Alanine Blocks Acute Opioid Withdrawal Induced-Conditioned Place Preference and Attenuates Somatic Withdrawal following Chronic Opioid Exposure in Rats. Psychoactives. 2024; 3(2):184-193. https://doi.org/10.3390/psychoactives3020012
Chicago/Turabian StyleAyoub, Samantha M., Erin M. Rock, Cheryl L. Limebeer, Marieka V. DeVuono, and Linda A. Parker. 2024. "Orally Administered N-Oleoyl Alanine Blocks Acute Opioid Withdrawal Induced-Conditioned Place Preference and Attenuates Somatic Withdrawal following Chronic Opioid Exposure in Rats" Psychoactives 3, no. 2: 184-193. https://doi.org/10.3390/psychoactives3020012
APA StyleAyoub, S. M., Rock, E. M., Limebeer, C. L., DeVuono, M. V., & Parker, L. A. (2024). Orally Administered N-Oleoyl Alanine Blocks Acute Opioid Withdrawal Induced-Conditioned Place Preference and Attenuates Somatic Withdrawal following Chronic Opioid Exposure in Rats. Psychoactives, 3(2), 184-193. https://doi.org/10.3390/psychoactives3020012