
The Source of Melanocytes in Ortho- and Heterotopic Tail Regenerates of Axolotls and the Dependence of the Regenerative Response on the Presence of Neural Tissue

Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experiments
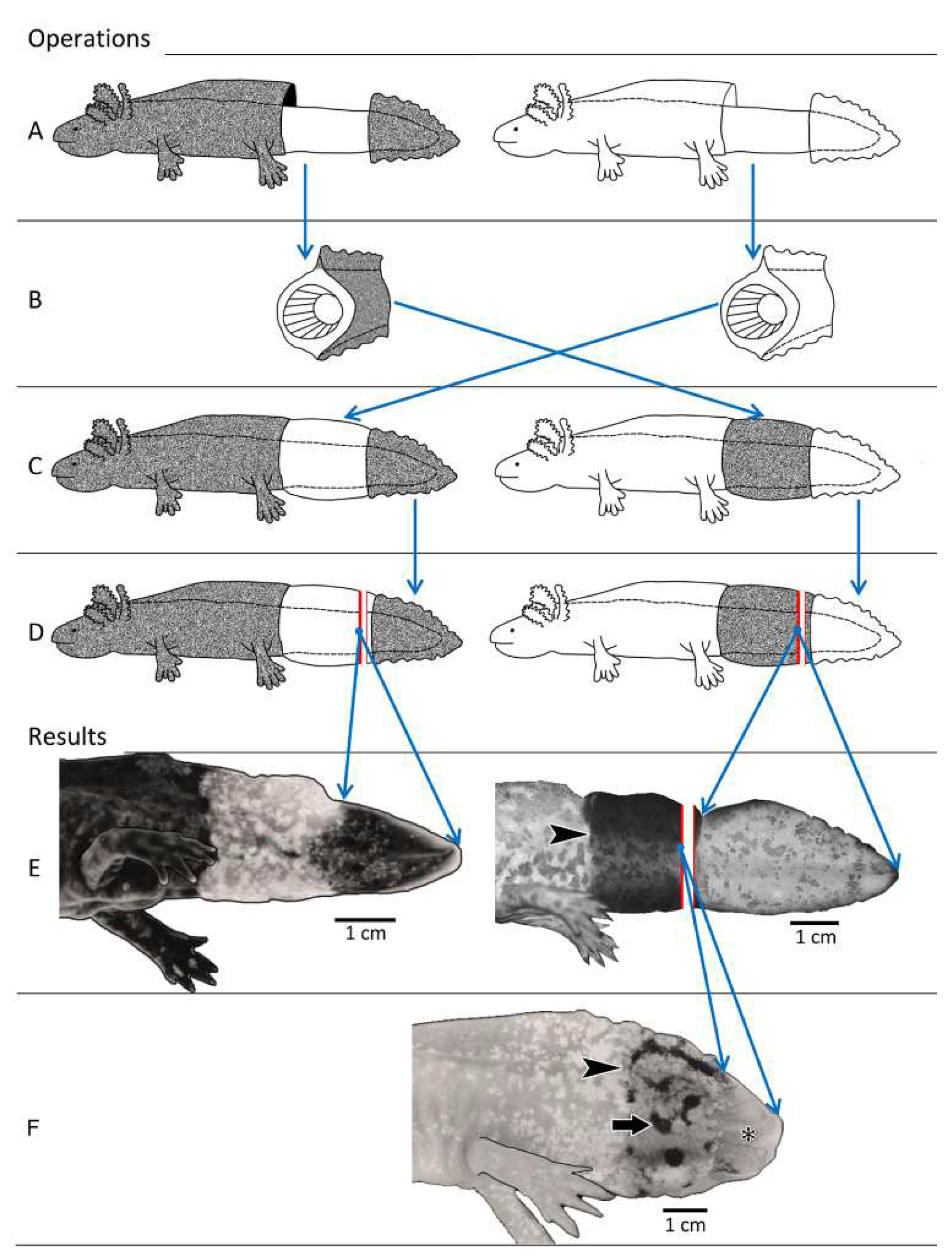
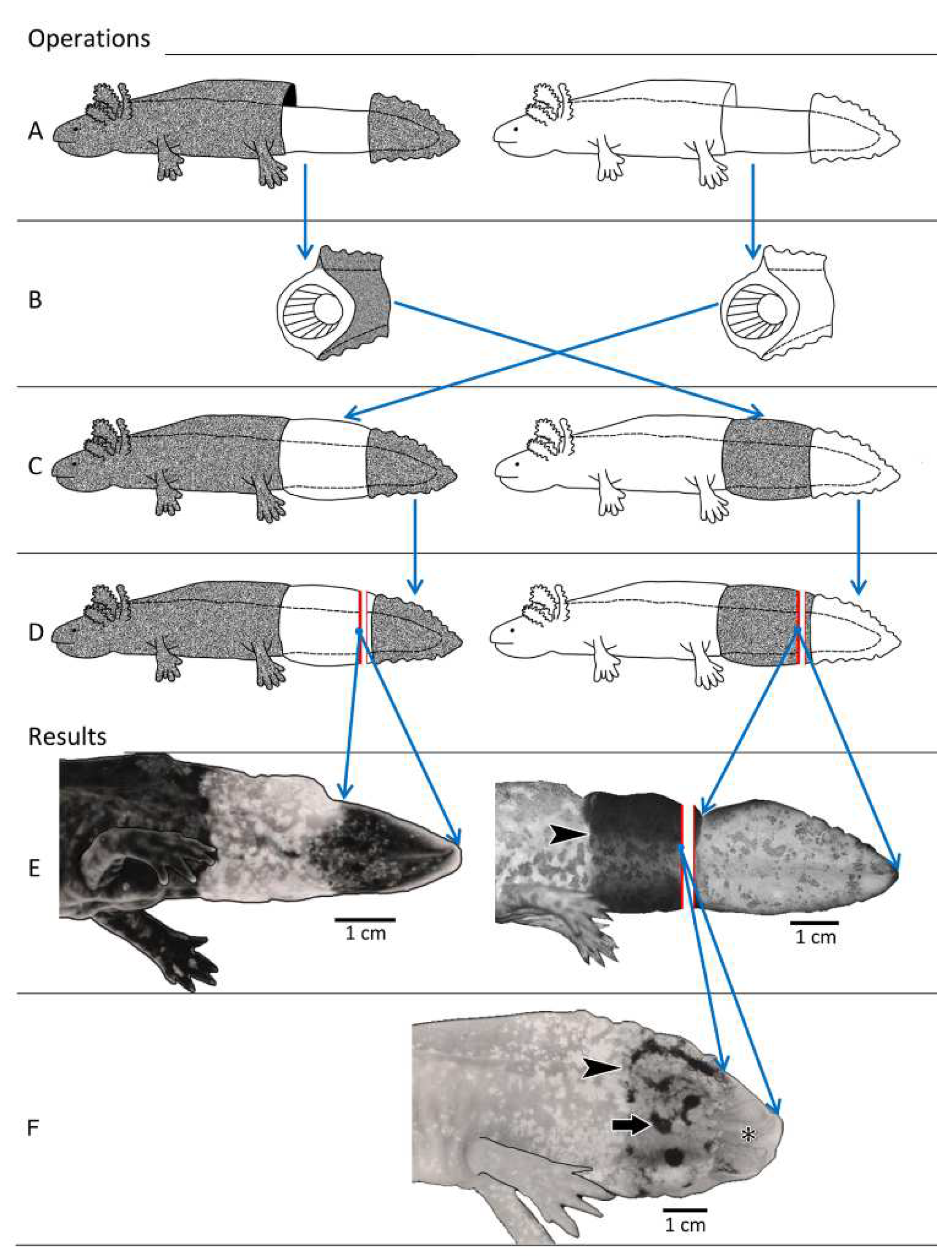
2.2.1. Experiment I: Reciprocal Exchange of Cuffs of Tail Integument between Axolotl Larvae
2.2.2. Experiment II: Transplantation of Cuffs of Tail Integument from Metamorphosed, Dark-Colored Axolotls onto Larvae of Dark and Yellow-Colored Axolotls
2.2.3. Experiment III: Xeno-Transplantation of a Cuff of Tail Integument from an Adult Fire Salamander on a Dark-Colored Axolotl Larva
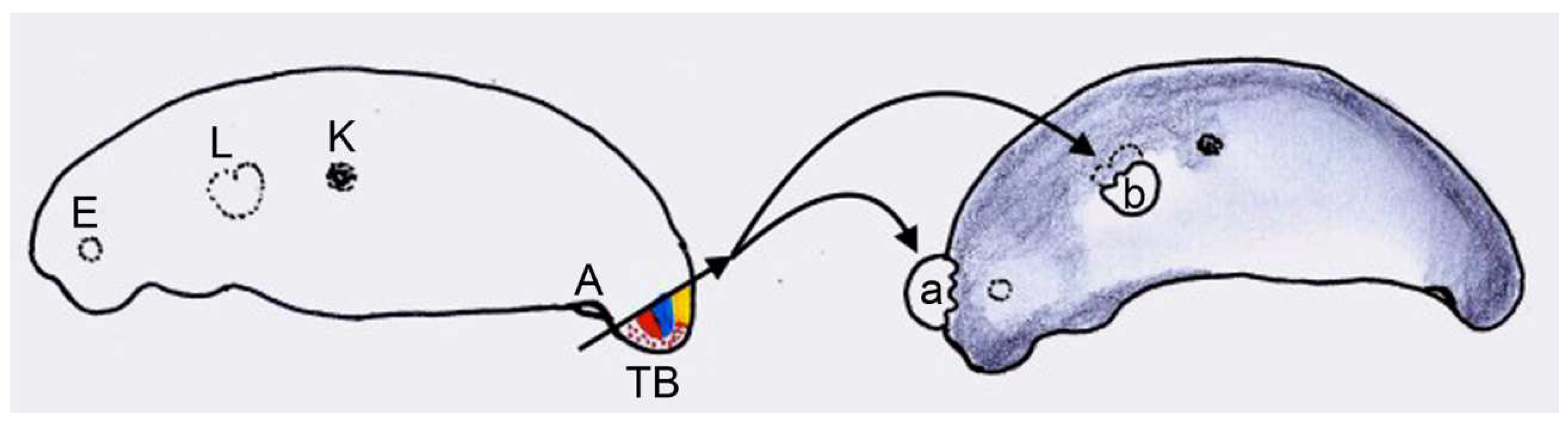
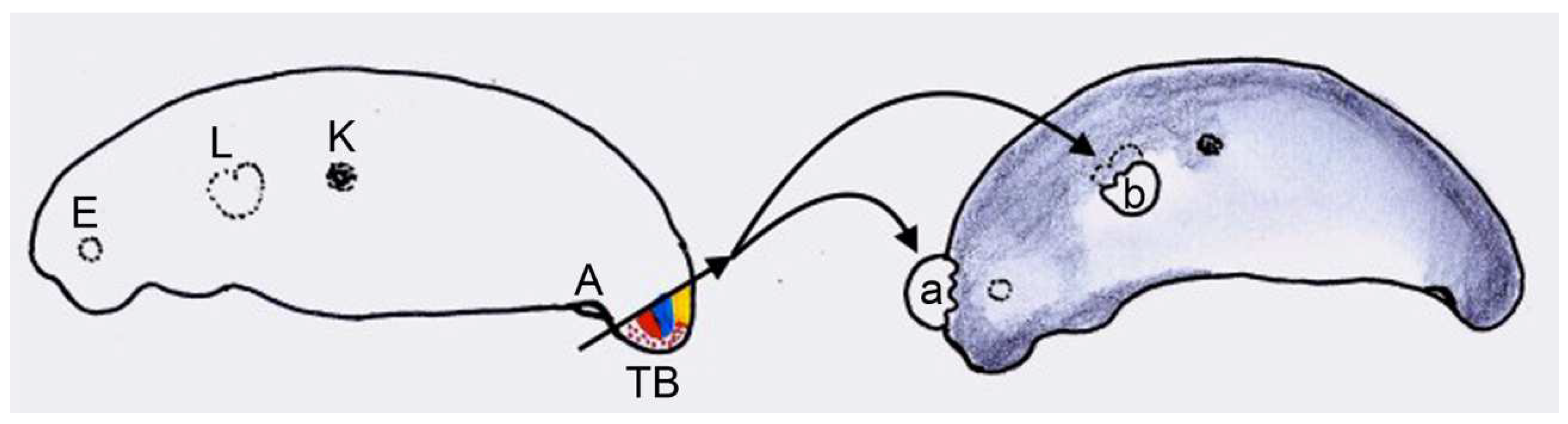
2.2.4. Experiment IV: Heterotopic Transplantation of Tail Buds from Yellow-Colored on Dark-Colored Axolotl Embryos, and Analysis of Tail Regeneration following Resection of the Heterotopic Tails
2.3. Light and Electron Microscopic Methods
2.4. Diagnostic Criteria Used in the Analysis of the Integumental Samples
3. Results
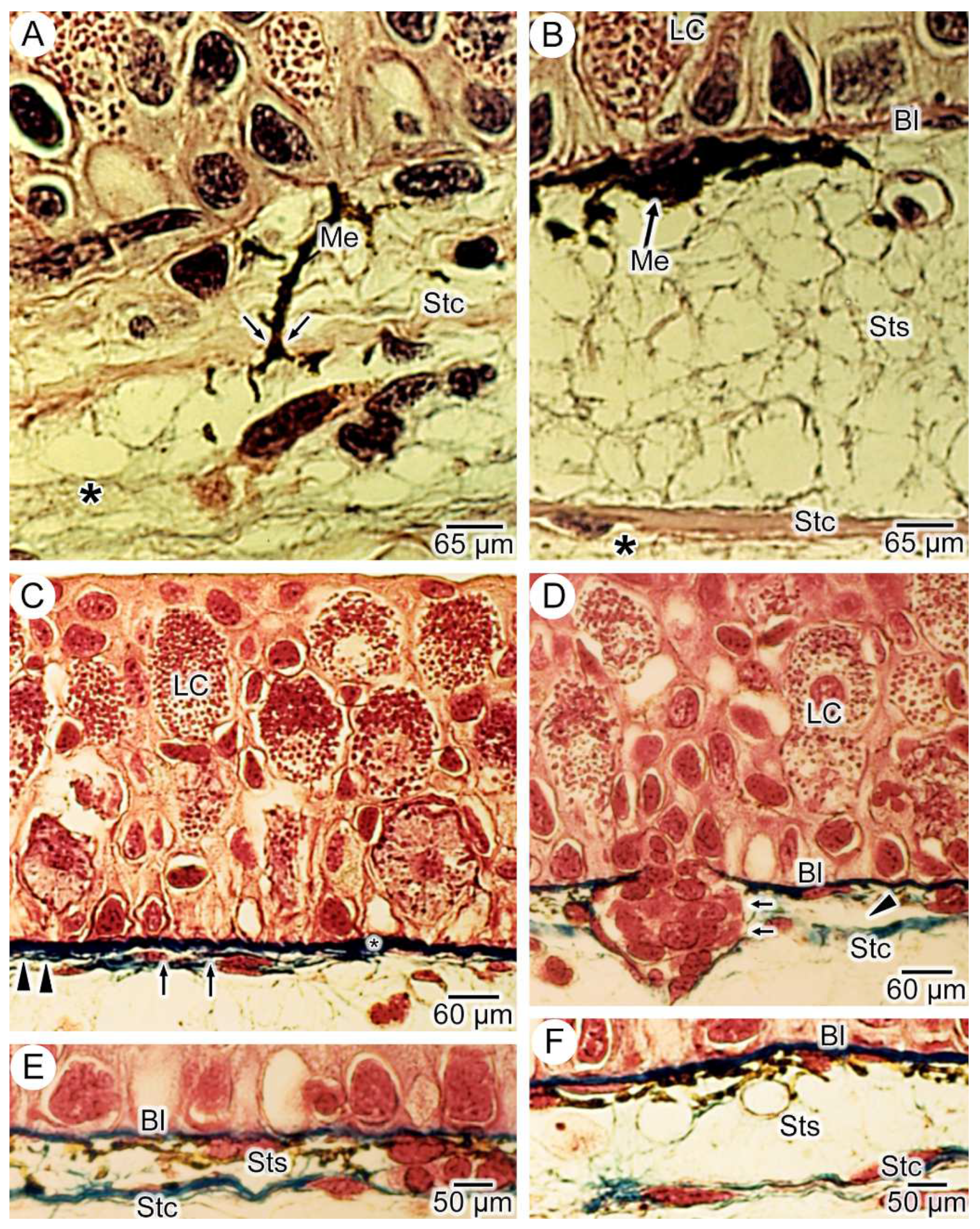
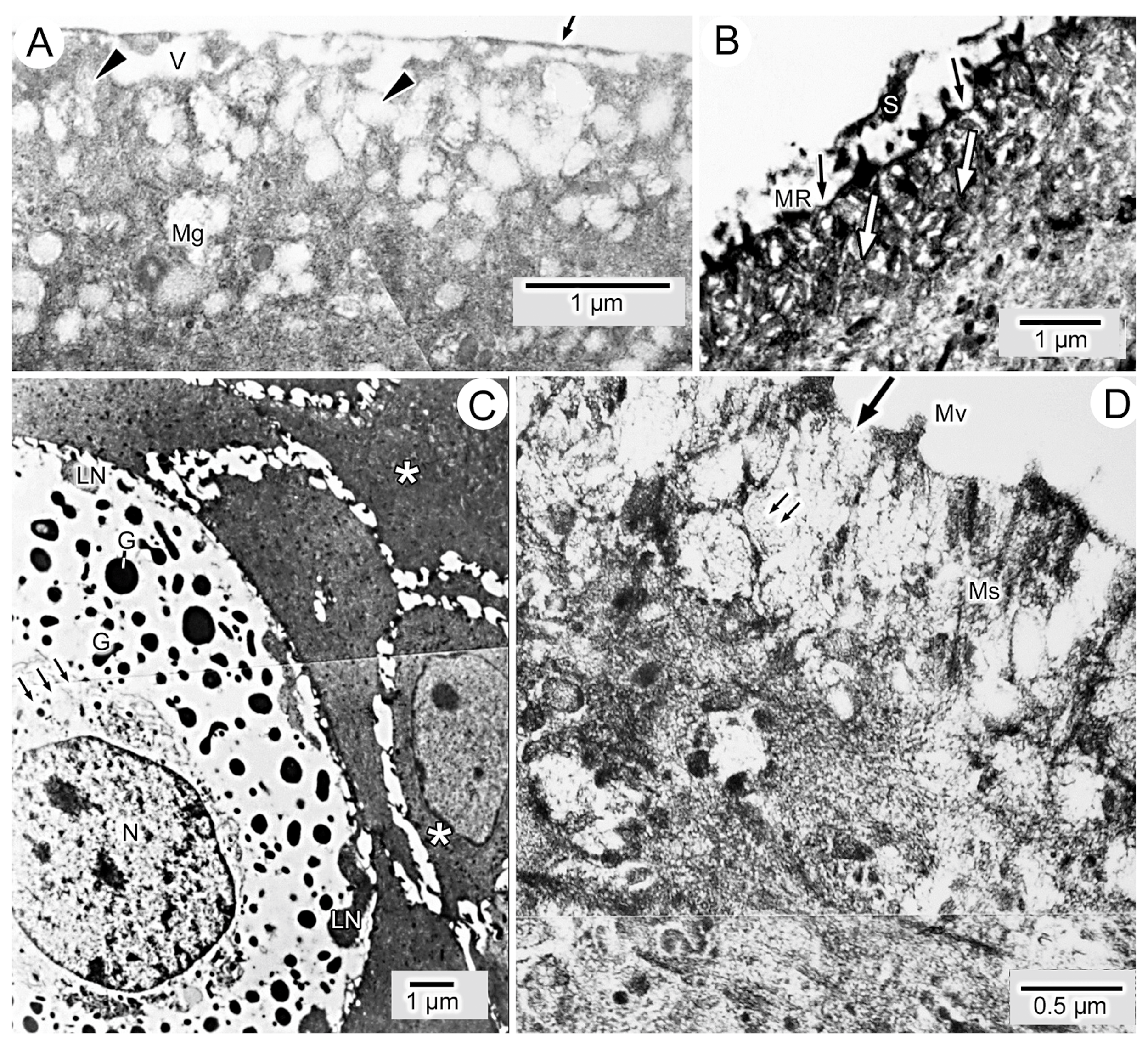
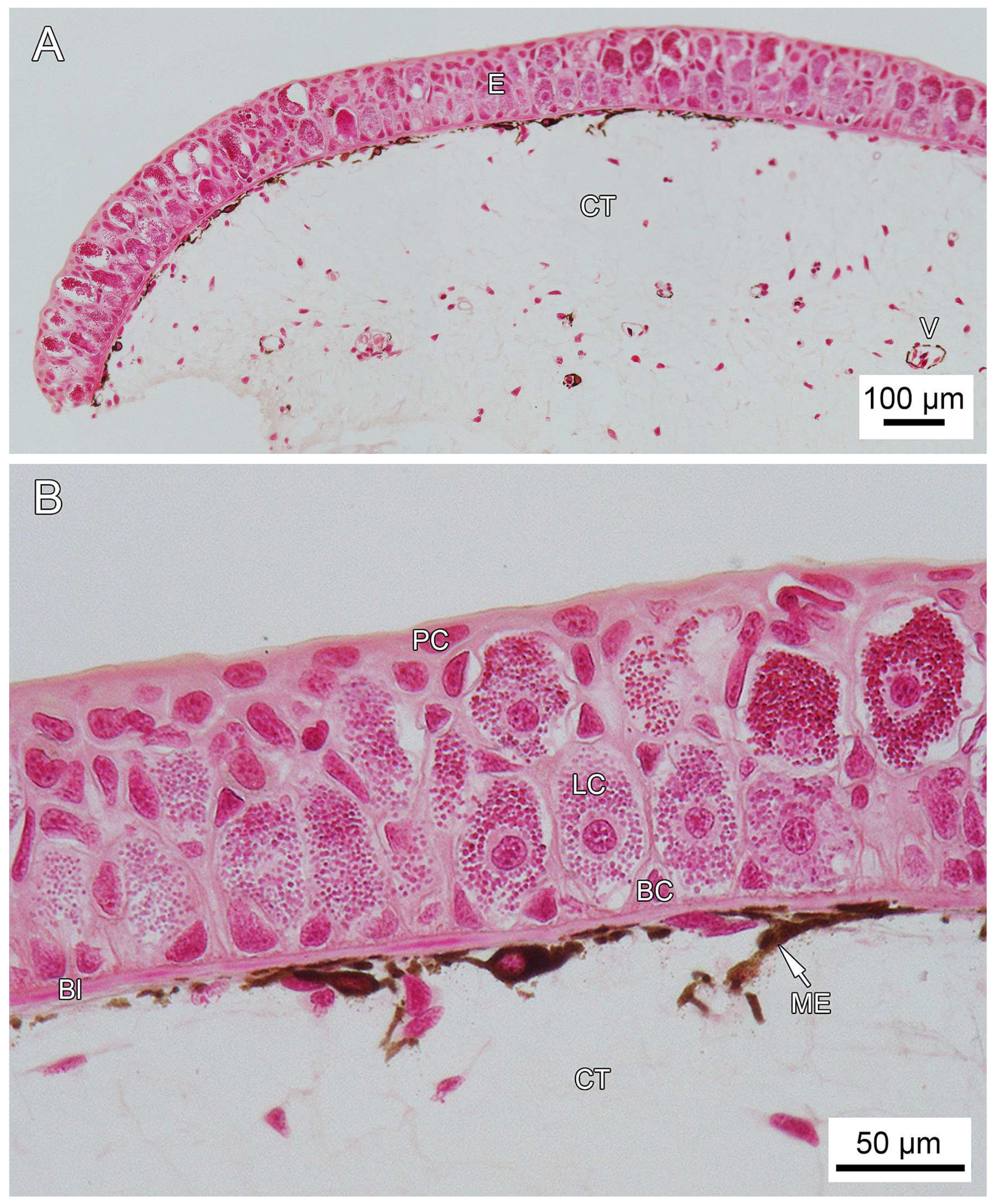
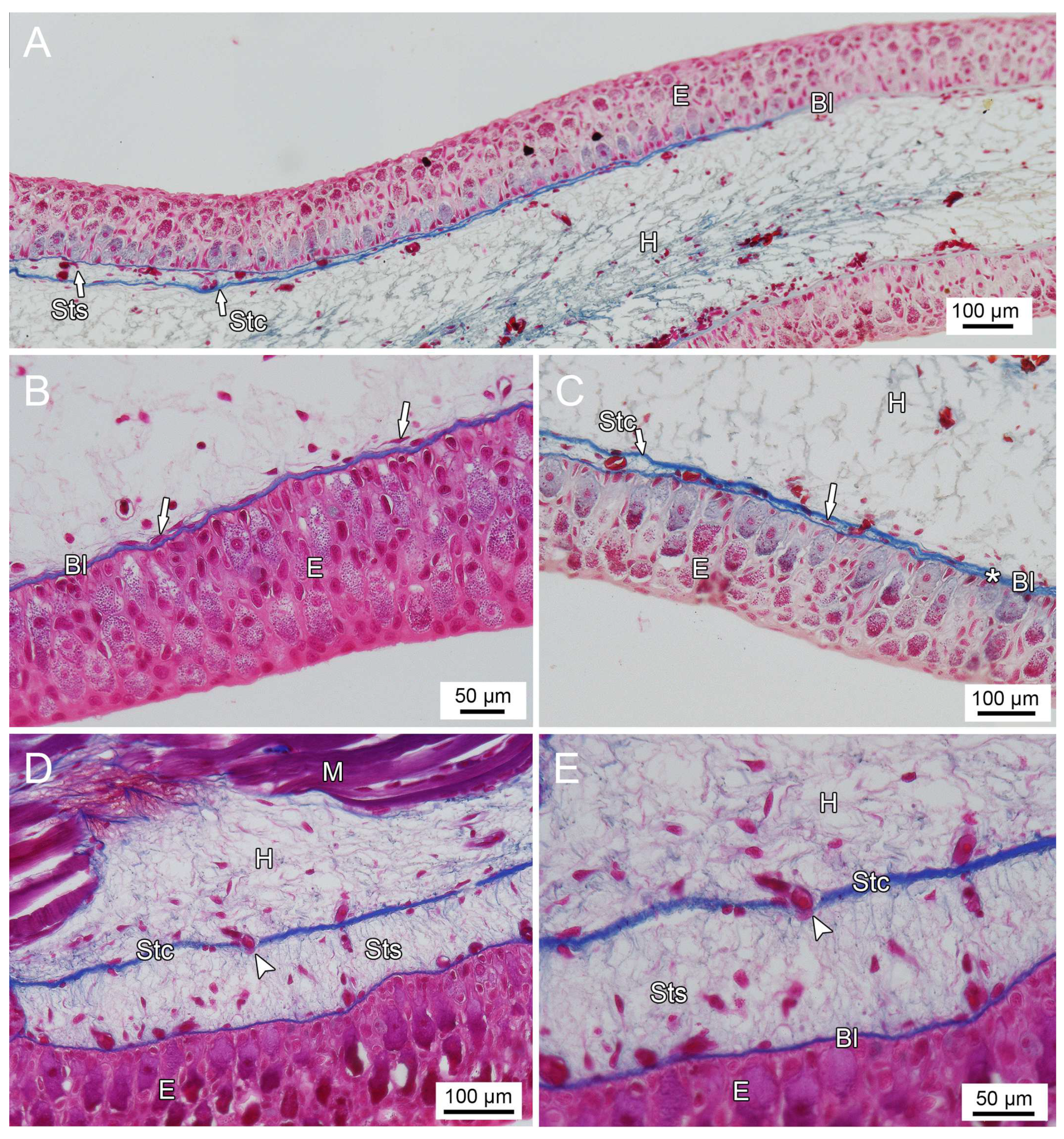
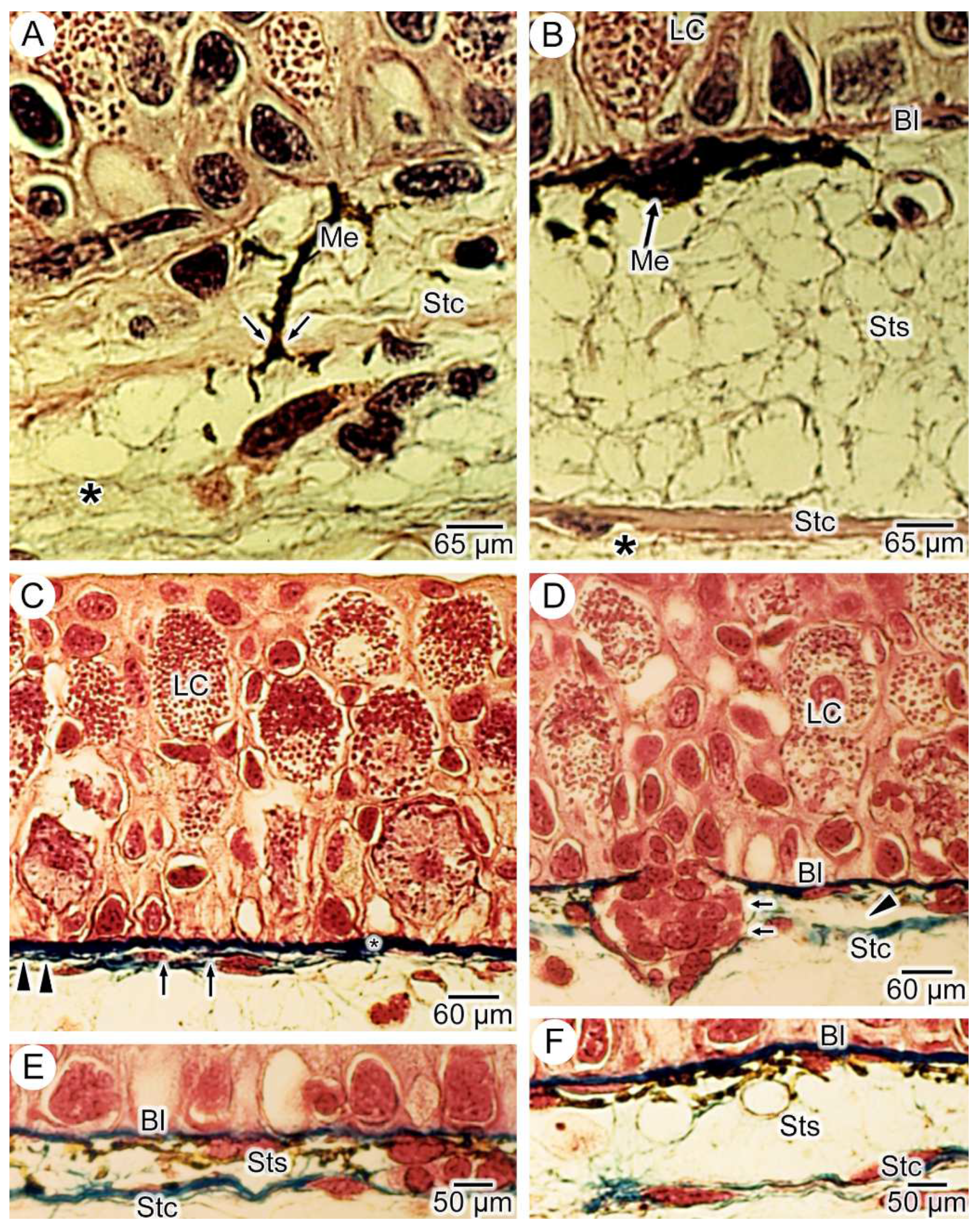
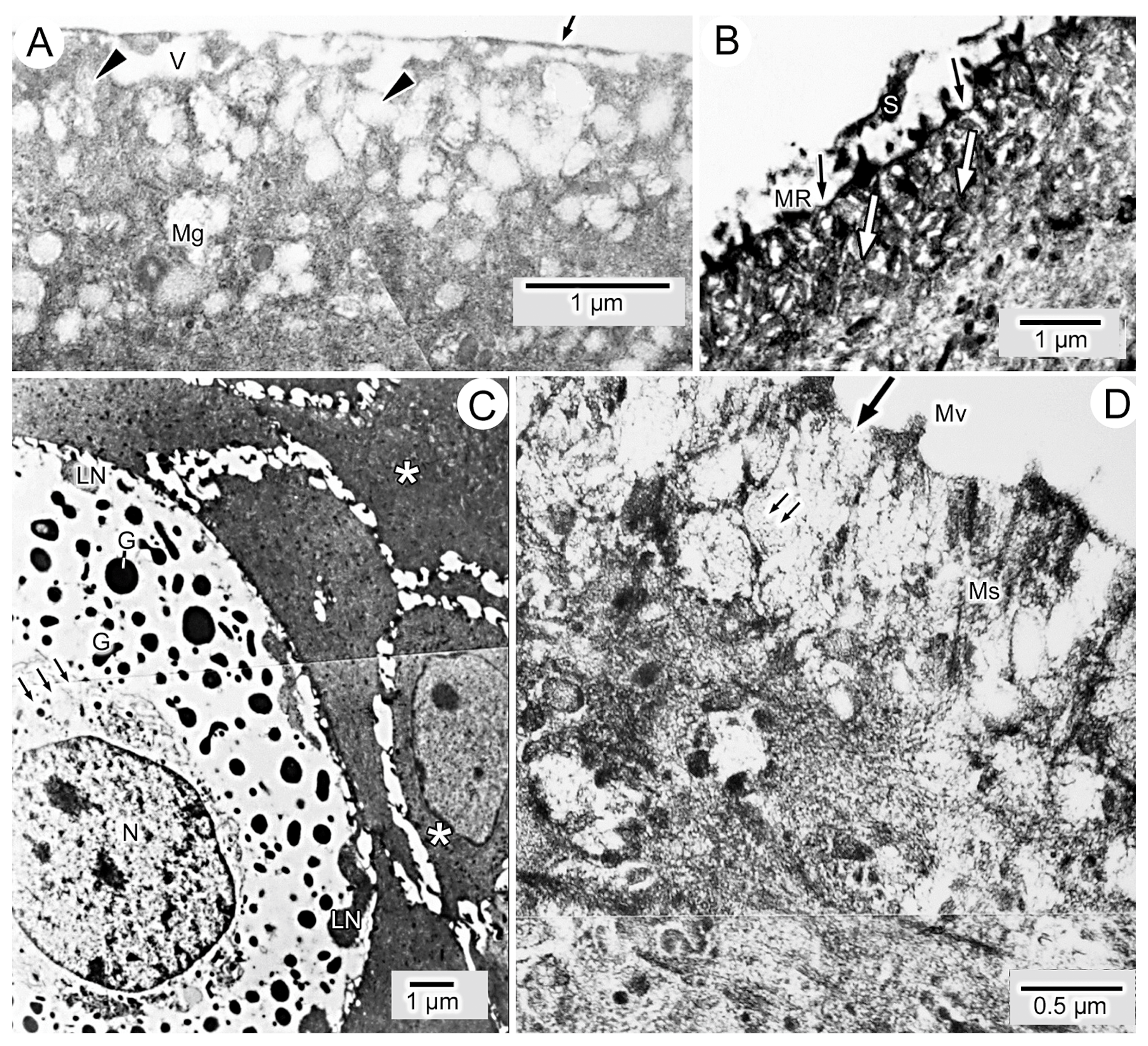
3.1. General Findings on Integumental Structure
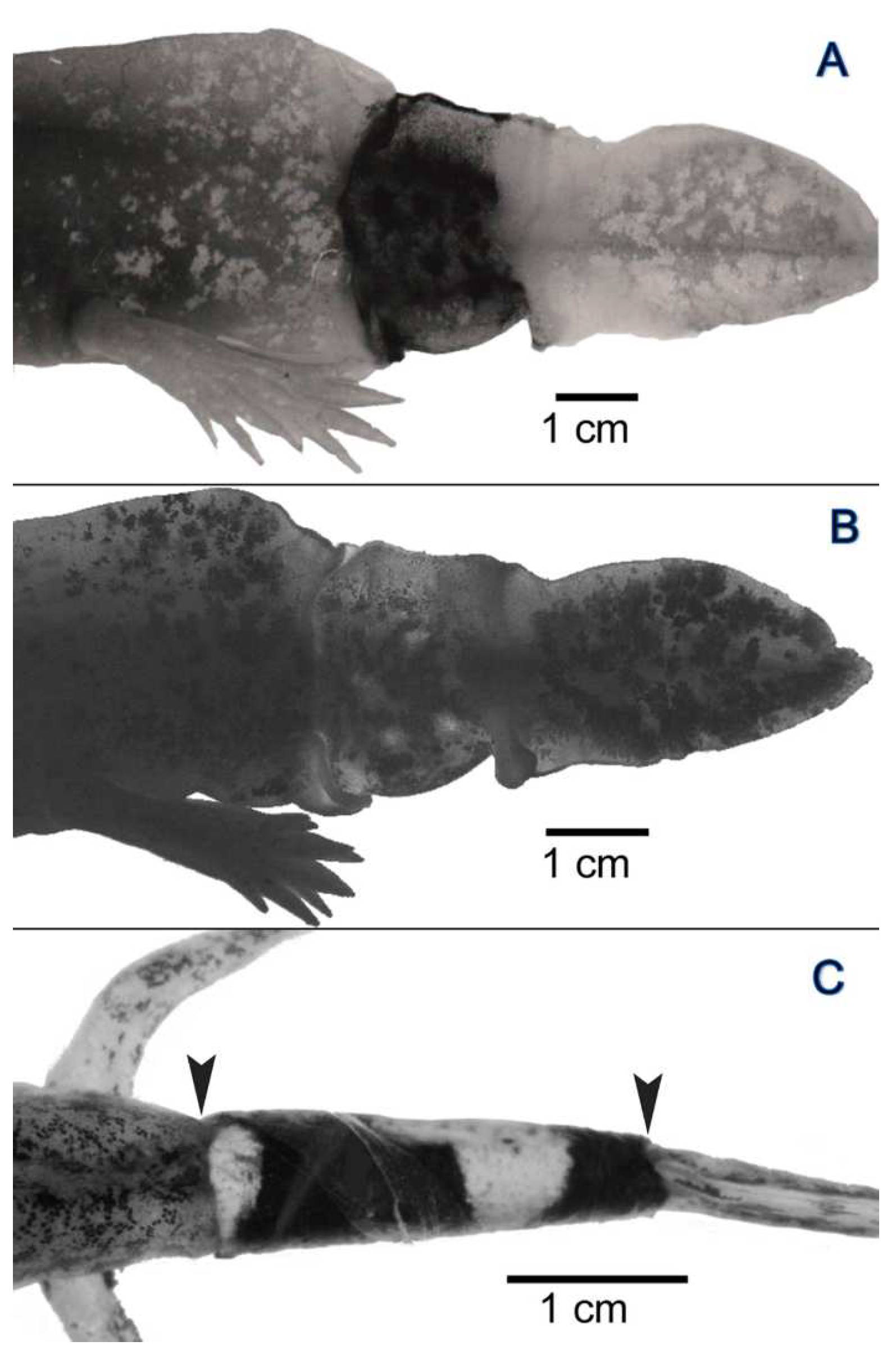
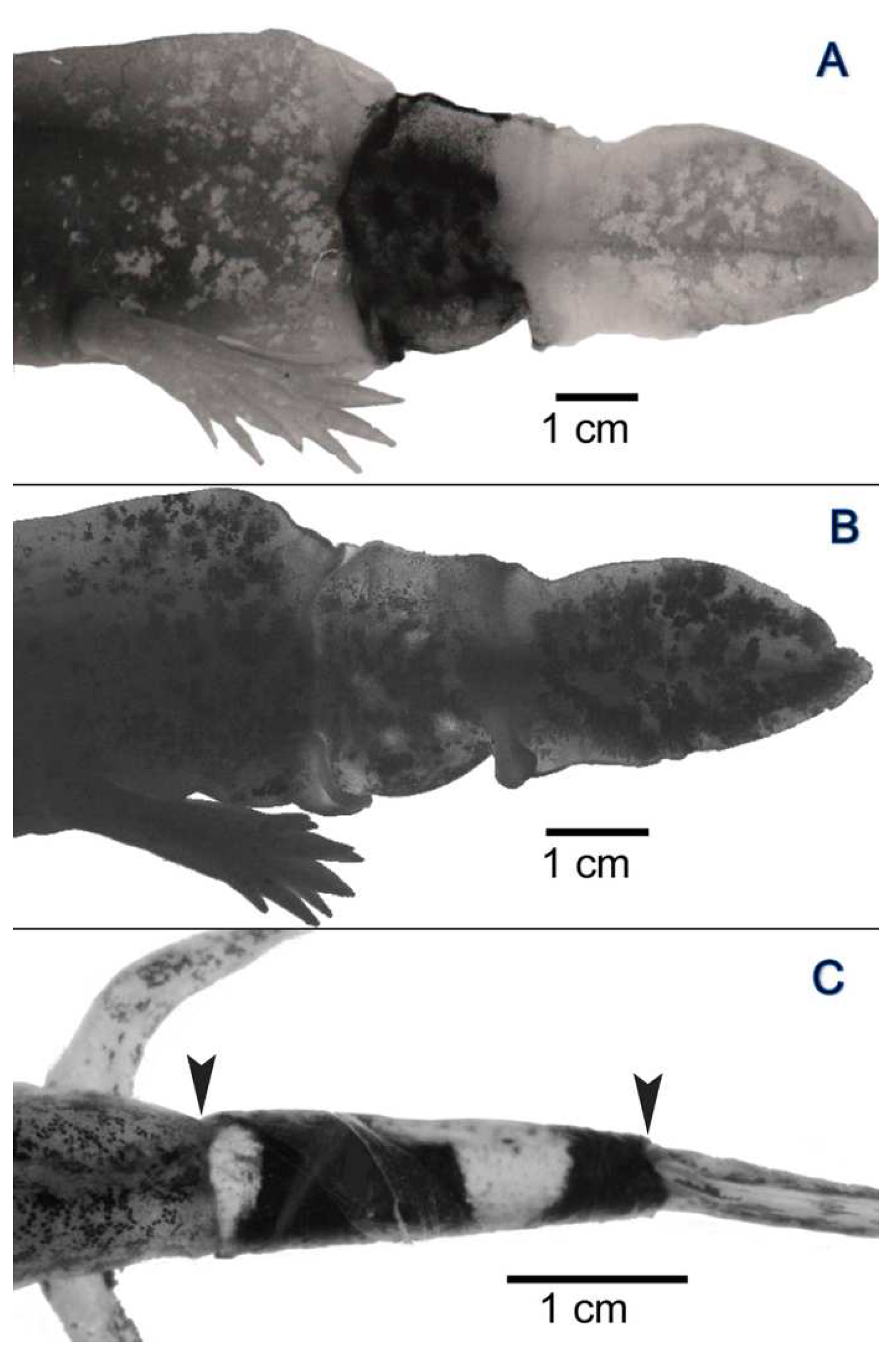
3.2. Experiment I: Reciprocal Exchange of Cuffs of Tail Integument between Axolotl Larvae
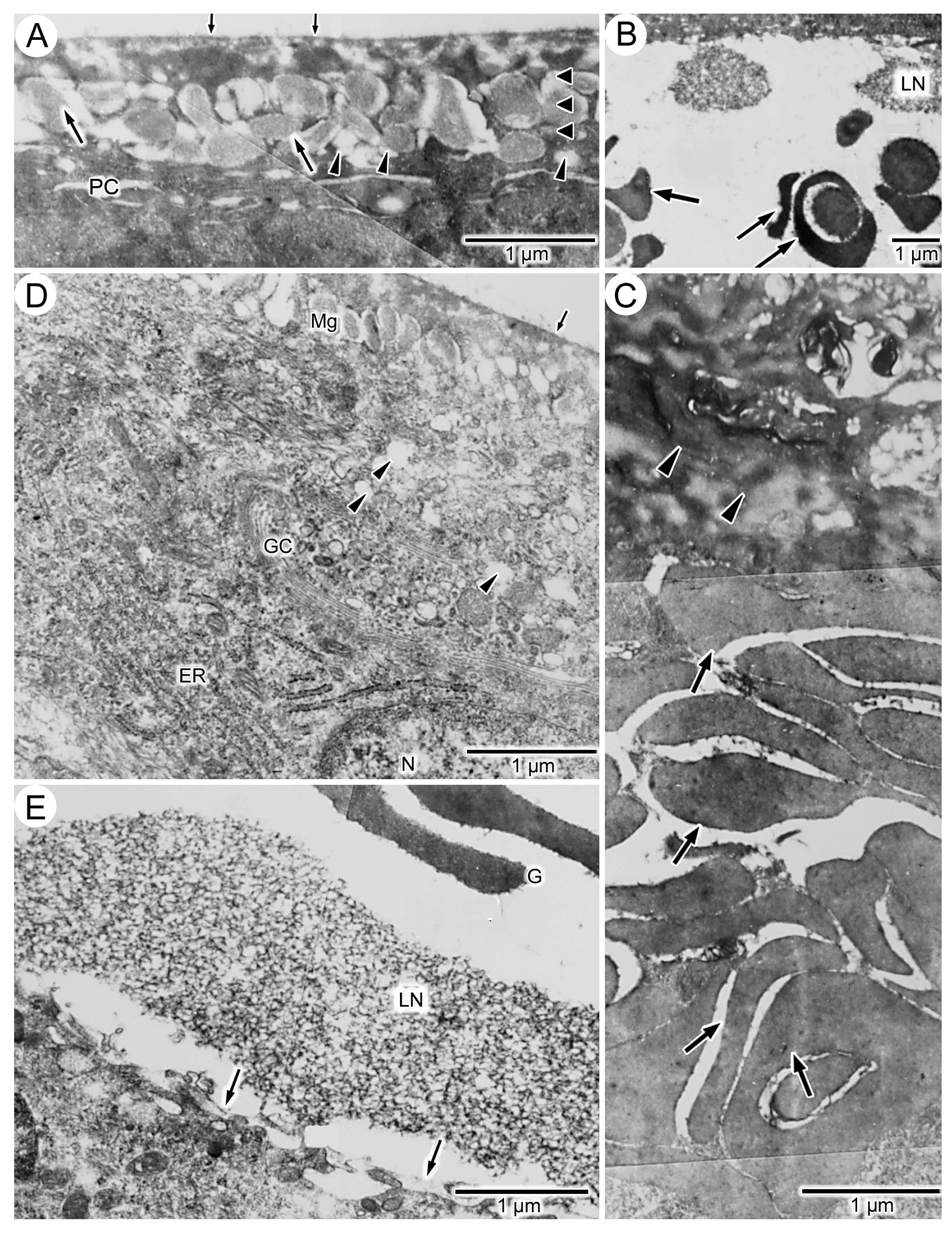
3.3. Experiment II: Transplantation of Cuffs of Tail Integument from Metamorphosed, Dark-Colored Axolotls onto Larvae of Dark-Colored and Yellow-Colored Axolotls
3.4. Experiment III: Xeno-Transplantation of a Cuff of Tail Integument from an Adult Fire Salamander on a Dark-Colored Axolotl Larva
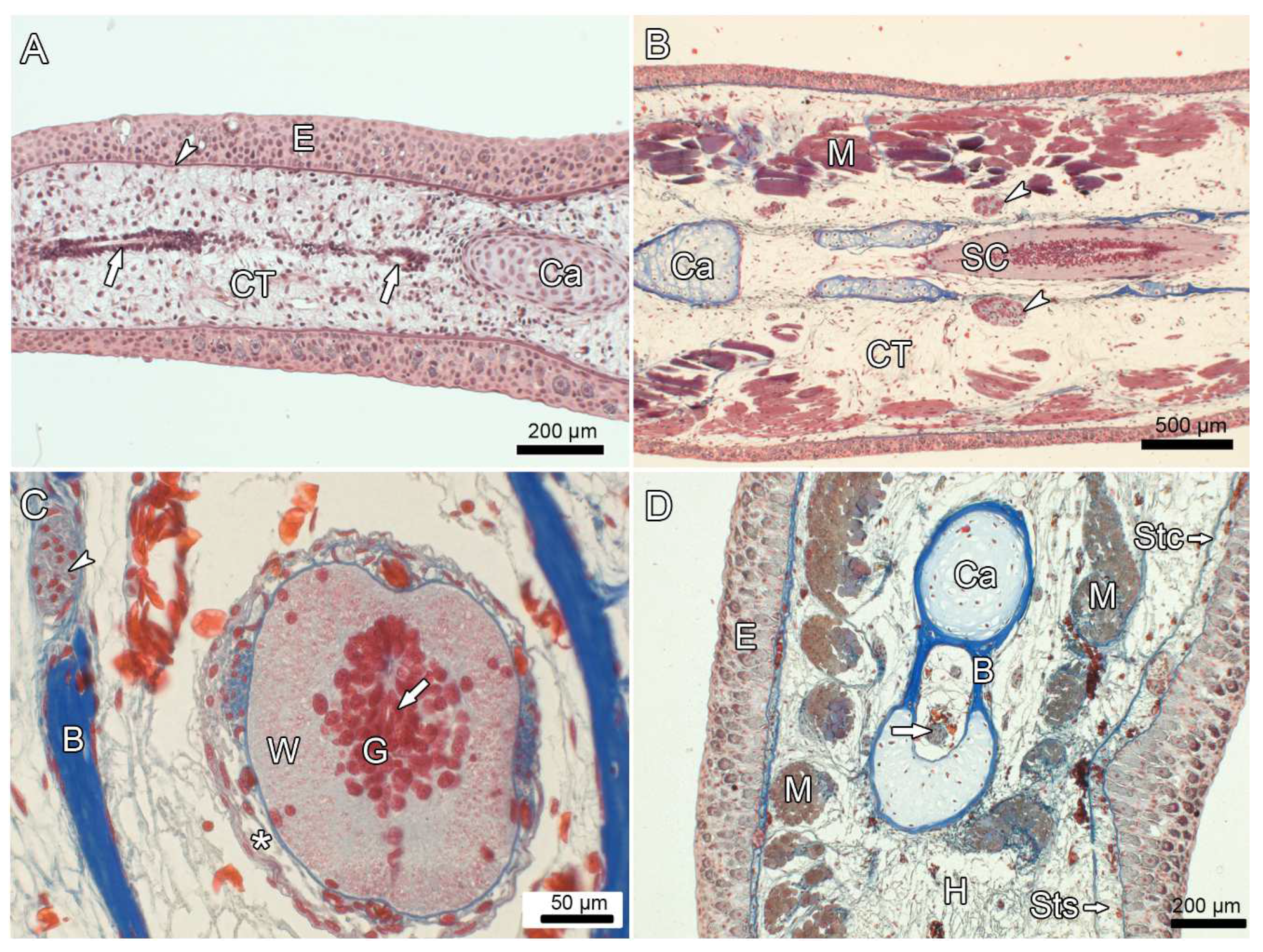
3.5. Experiment IV: Heterotopic Transplantation of Tail Buds from Yellow-Colored on Dark-Colored Axolotl Embryos, and Tail Regeneration following Resection of the Heterotopic Tails

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal No. | Sex (m/f) | Donor Stage | Host Stage | Time between Transplantation and Amputation of Tail (Months) | Body Length at Amputation (cm) | Location of Amputated Tail Transplant (Head, Limb) | Length of Amputated Tail Transplant (cm, % of Total Transplant) | Age at End of Experiment (months) | Body Length at End of Experiment (cm) | Tail Regenerate (yes/no), Length of Tail Regenerate (cm) | Presence of Spinal Cord (sc) or Only Ependymal Tissue (e) in Tail Regenerate | Figures |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | m | 32 | 32 | 24 | 19.5 | head | 3.9 (50) | 40 | 25.0 | yes, 7.0 | sc | Figure 8D |
| 2 | f | 32 | 32 | 24 | 19.0 | limb | 4.0 (50) | 40 | 24.0 | yes, 8.1 | sc | - |
| 3 | m | 32 | 29 | 24 | 18.0 | head | 3.6 (50) | 36 | 24.0 | yes, 7.1 | sc | - |
| 4 | f | 32 | 29 | 24 | 19.0 | limb | 4.1 (50) | 42 | 26.0 | yes, 8.6 | sc | Figure 7A |
| 5 | m | 29 | 29 | 22 | 21.0 | head | 2.1 (25) | 36 | 24.0 | yes, 8.2 | sc | Figure 8B |
| 6 | f | 29 | 33 | 22 | 18.5 | head | 5.5 (75) | 40 | 24.5 | yes, 6.9 | sc | Figure 8C |
| 7 | f | 31 | 33 | 18 | 18.0 | head | 7.4 (95) | 30 | 26.0 | yes, 2.8 | e | Figure 7B and Figure 10A,C |
| 8 | f | 30 | 32 | 24 | 19.0 | limb | 7.6 (95) | 36 | 25.5 | yes, 4.1 | e | Figure 7C and Figure 9A,B |
| 9 | m | 32 | 30 | 18 | 18.5 | head | 7.5 (95) | 28 | 22.5 | yes, 1.7 | e | Figure 8A |
| 10 | m | 29 | 29 | 15 | 18.5 | head | 5.7 (95) | 28 | 20.0 | yes, 2.2 | e | Figure 10B,D,E |
| 11 | f | 28 | 30 | 19 | 18.0 | head | 7.6 (100) | 30 | 22.5 | no * | neither | Figure 7D,E |
| 12 | m | 29 | 32 | 19 | 18.5 | head | 7.4 (100) | 28 | 23.0 | no * | neither | Figure 7F |
| 13 | f | 29 | 32 | 20 | 18.0 | limb | 7.8 (100) | 40 | 26.0 | no * | neither | - |

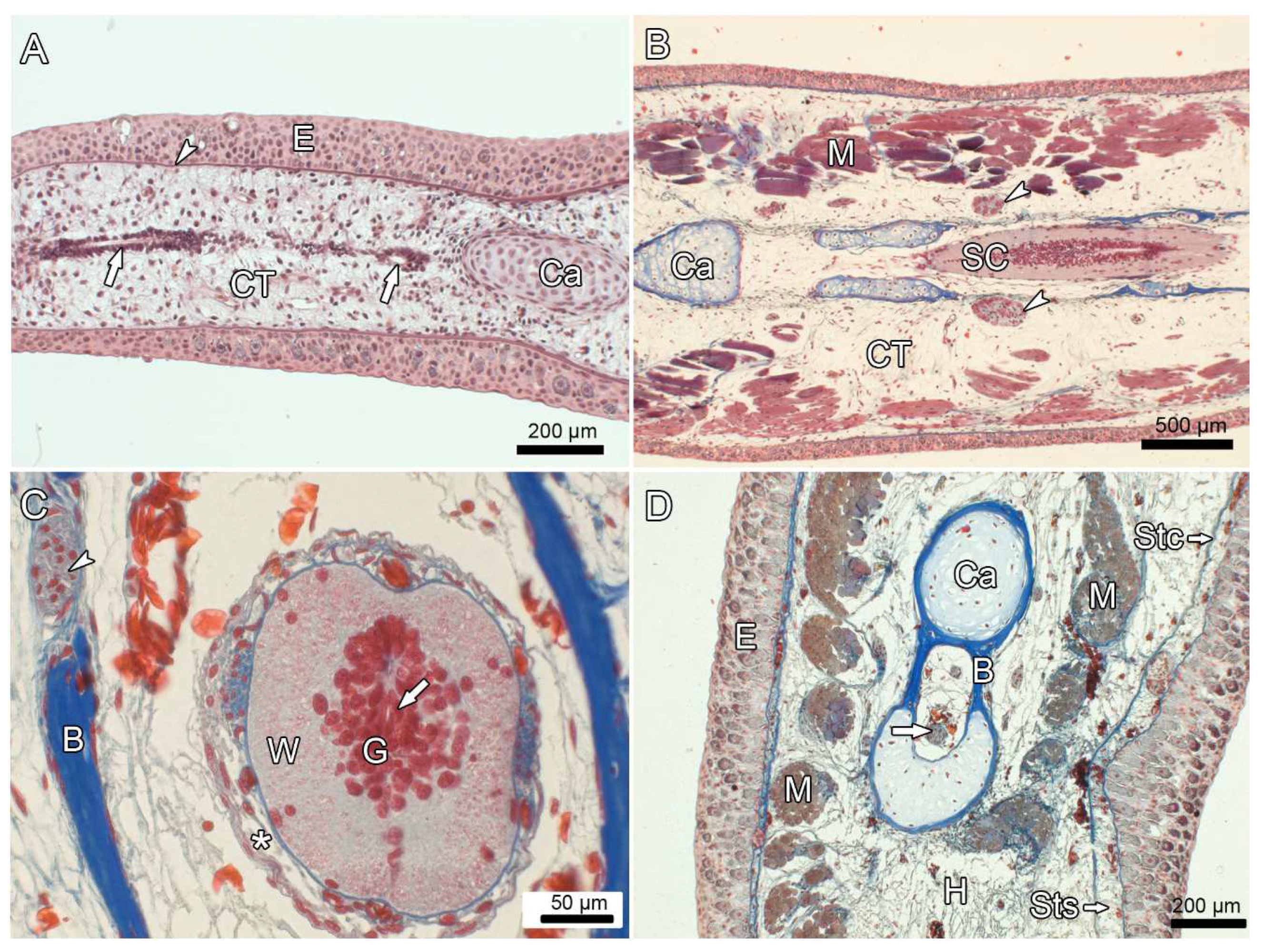
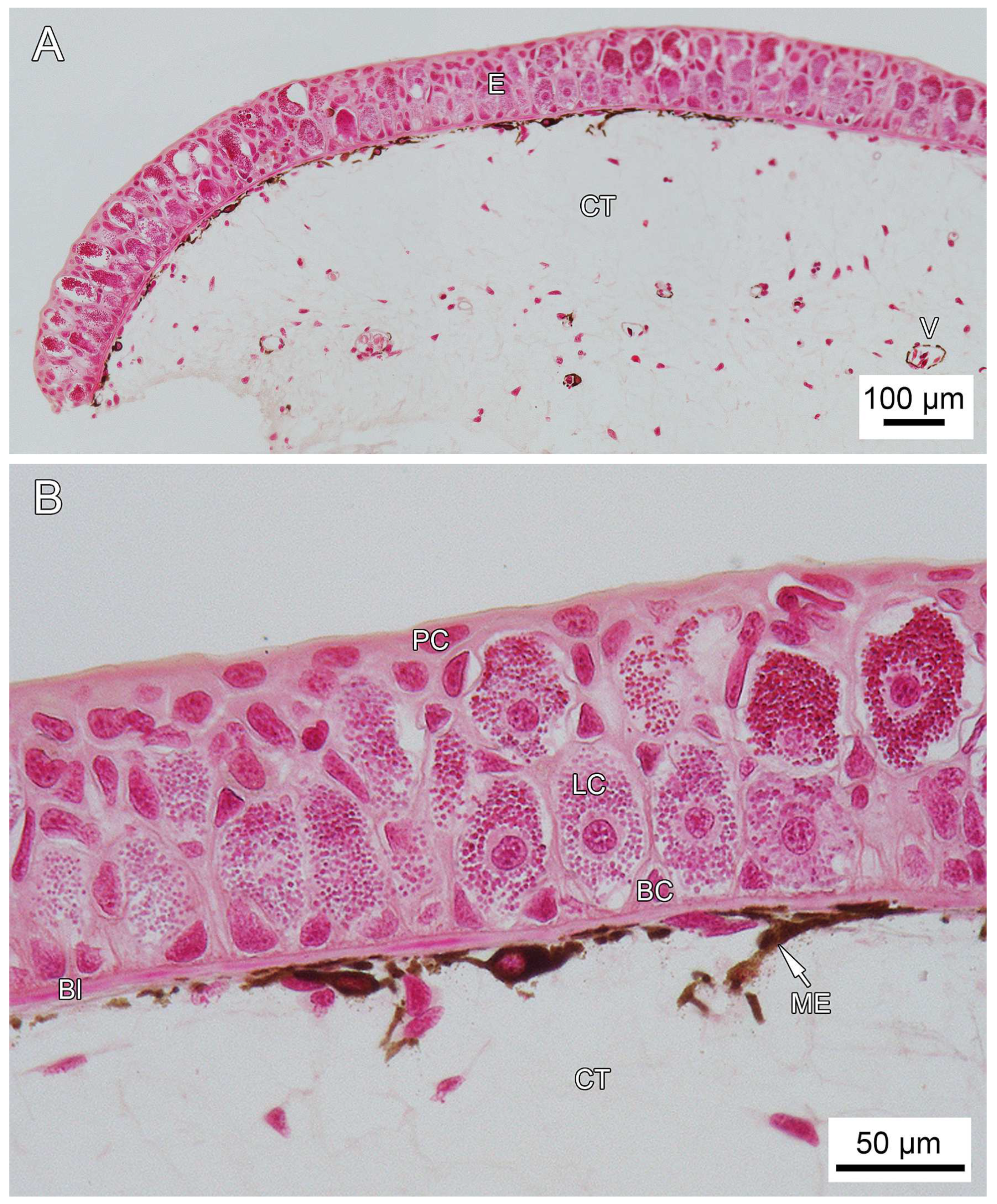
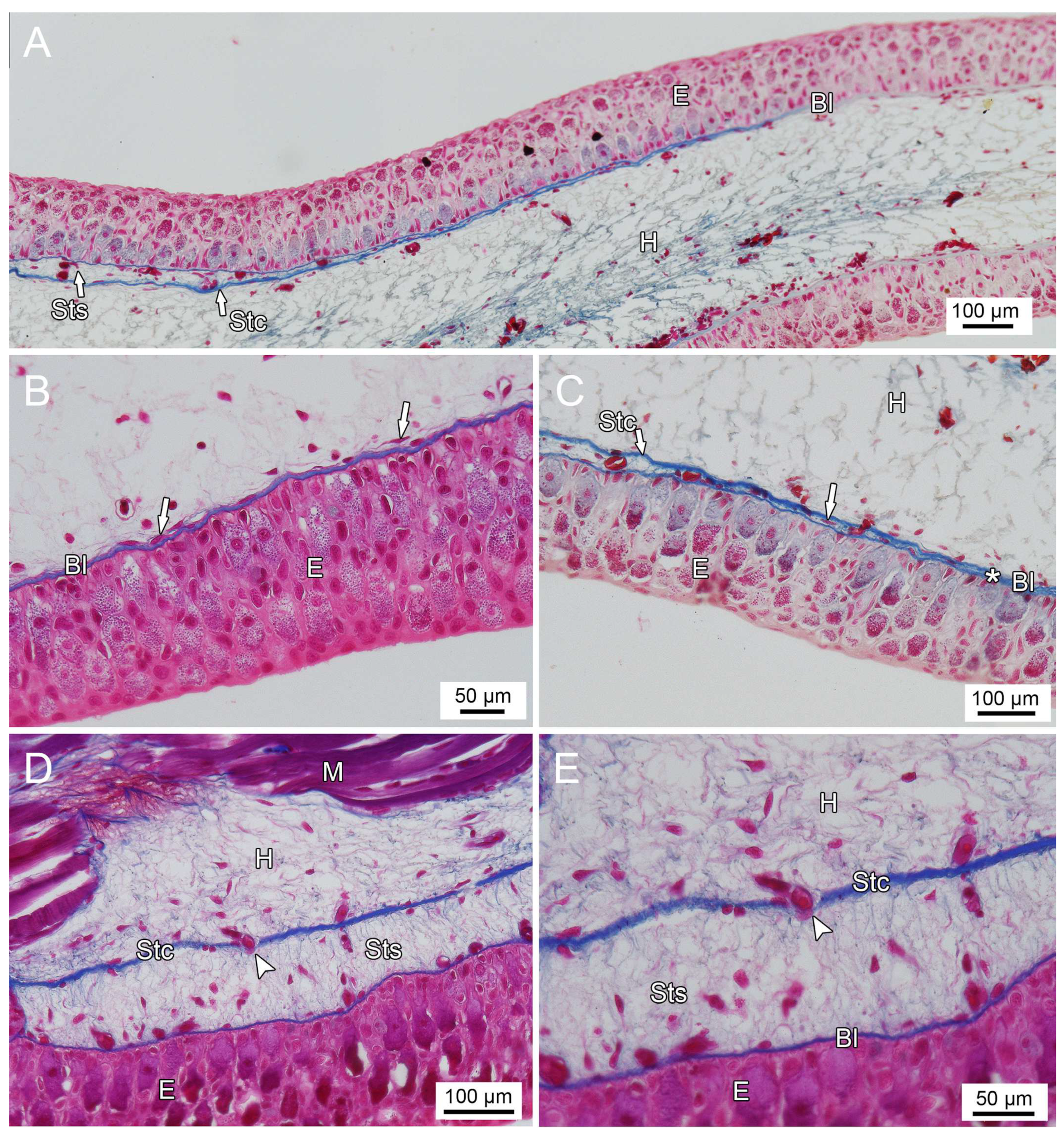
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carlson, B.M. Principles of Regenerative Biology; Elsevier Academic Press: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Stocum, D.L. Regenerative Biology and Medicine, 2nd ed.; Academic Press: Burlington, NJ, USA, 2012. [Google Scholar]
- Murugan, N.J.; Vigran, H.J.; Miller, K.A.; Golding, A.; Pham, Q.L.; Sperry, K.A.; Rasmussen-Ivey, C.; Kane, A.W.; Kaplan, D.L.; Levin, A. Acute multidrug delivery via a wearable bioreactor facilitates long-term limb regeneration and functional recovery in adult Xenopus laevis. Sci. Adv. 2022, 8, eabj2164. [Google Scholar] [CrossRef]
- Alibardi, L. Morphological and cellular aspects of tail and limb regeneration in lizards. In Advances in Anatomy, Embryology and Cell Biology; Springer: Berlin, Germany, 2010; Volume 207. [Google Scholar]
- Kierdorf, U.; Kierdorf, H. Antler regrowth as a form of epimorphic regeneration in vertebrates—A comparative view. Front. Bioscience 2012, E4, 1606–1624. [Google Scholar] [CrossRef]
- Neufeld, D.A.; Zhao, W. Bone regrowth after digit tip amputation in mice is equivalent in adults and neonates. Wound Repair Regen. 1995, 3, 461–466. [Google Scholar] [CrossRef]
- Seifert, A.W.; Muneoka, K. The blastema and epimorphic regeneration in mammals. Dev. Biol. 2018, 433, 190–199. [Google Scholar] [CrossRef]
- Alibardi, L. Review: Limb regeneration in humans: Dream or reality? Ann. Anat. 2018, 217, 1–6. [Google Scholar] [CrossRef]
- Daponte, V.; Tylzanowski, P.; Forlino, A. Appendage regeneration in vertebrates: What makes this possible? Cells 2021, 10, 242. [Google Scholar] [CrossRef]
- Hutchinson, C.; Pilote, M.; Roy, S. The axolotl limb: A model for bone development, regeneration and fracture healing. Bone 2007, 40, 45–56. [Google Scholar] [CrossRef]
- Kragl, M.; Knapp, D.; Nacu, E.; Khattak, S.; Maden, M.; Epperlein, H.H.; Tanaka, E.M. Cells keep a memory of their tissue origin during axolotl limb regeneration. Nature 2009, 460, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, R.R. Albino axolotl from an albino tiger salamander through hybridization. J. Heredity 1967, 58, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Epperlein, H.H.; Löfberg, J. Xanthophores in chromatophore groups of the premigratory neural crest initiate the pigment pattern of the axolotl larva. Roux’s Arch. Dev. Biol. 1984, 193, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Clemen, G.; Greven, H. Morphologische Untersuchungen an der Mundhöhle von Urodelen. III. Die Munddachbezahnung von Ambystoma mexicanum Cope (Ambystomatidae, Amphibia). Zool. Jb. Anat. 1977, 98, 95–136. [Google Scholar]
- Clemen, G. Competence and reactions of early- and late-larval dental laminae in original and not-original dental systems of Ambystoma mexicanum Shaw. Arch. Biol. 1988, 99, 307–324. [Google Scholar]
- Akat, E.; Yenmis, M.; Pombal, M.A.; Molist, P.; Arman, S.; Vesely, M.; Anderson, A.; Ayaz, D. Comparison of vertebrate skin structure at class level: A review. Anat. Rec. 2022, 305, 3543–3608. [Google Scholar] [CrossRef]
- Dupin, E.; Le Douarin, N.M. Development of melanocyte precursors from the vertebrate neural crest. Oncogene 2003, 22, 3016–3023. [Google Scholar] [CrossRef] [PubMed]
- Theis, A. Untersuchungen über die Epidermis im Individualcyclus von Salamandra maculosa. Z. Wiss. Zool. 1932, 14, 356–420. [Google Scholar]
- Fährmann, W. Die Morphodynamik der Epidermis des Axolotls (Siredon mexicanum SHAW) unter dem Einfluß von exogen appliziertem Thyroxin. I. Die Epidermis des neotenen Axolotls. Z. Mikros. Anat. Forsch. Leipzig 1971, 83, 472–506. [Google Scholar]
- Jarial, M.S. Fine structure of the epidermal Leydig cells in the axolotl Ambystoma mexicanum in relation to their function. J. Anat. 1989, 67, 95–102. [Google Scholar]
- Gerling, S.; D’Haese, J.; Greven, H. Number and distribution of Leydig cells (LC) in the epidermis of the growing axolotl, Ambystoma mexicanum (Amphibia: Urodela). Vert. Zool. 2012, 62, 97–111. [Google Scholar] [CrossRef]
- Clemen, G.; Greven, H. Organization of the palate in a spontaneously transforming Mexican axolotl (Ambystoma mexicanum): A case report. Vert Zool. 2015, 65, 351–356. [Google Scholar] [CrossRef]
- Taurog, A.; Oliver, C.; Eskay, R.L.; Porter, J.C.; McKenzie, J.M. The role of TRH in the neoteny of the Mexican axolotl (Ambystoma mexicanum). Gen. Comp. Endocrinol. 1974, 24, 267–279. [Google Scholar] [CrossRef]
- Larras-Regard, E. Hormonal determination of neoteny in facultative neotenic Urodeles. In Metamorphosis; Balls, M., Bownes, M., Eds.; Clarendon Press: Oxford, UK, 1985; pp. 294–311. [Google Scholar]
- Page, R.B.; Monaghan, J.; Walker, J.; Voss, R. A model of transcriptional and morphological changes during thyroid hormone-induced metamorphosis of the axolotl. Gen. Comp. Endocrinol. 2009, 162, 219–232. [Google Scholar] [CrossRef]
- Laudet, V. The origin and evolution of vertebrate metamorphosis. Curr. Biol. 2011, 21, R726–R737. [Google Scholar] [CrossRef]
- Denver, R.J. Neuroendocrinology of amphibian metamorphosis. In Current Topics in Developmental Biology: Animal Metamorphosis; Shi, Y.-B., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; Volume 103, pp. 195–228. [Google Scholar]
- Geigy, R. Die Metamorphose als Folge gewebsspezifischer Determination. Rev. Suisse Zool. 1941, 48, 483–494. [Google Scholar]
- Johnson, C.K.; Voss, S.R. Salamander paedomorphosis: Linking thyroid hormone to life history and life cycle evolution. In Current Topics in Developmental Biology; Shi, Y.-B., Ed.; Academic Press: Burlington, NJ, USA, 2013; Volume 103, pp. 229–258. [Google Scholar]
- Hartwig, H. Metamorphose-Reaktionen auf einen lokalisierten Hormonreiz. Biol. Zentralbl. 1940, 60, 473–478. [Google Scholar]
- Hartwig, H. Gestaltungstendenzen und Metamorphosereaktion bei subcutan transplantierten Hautfragmenten. Naturwissenschaften 1962, 49, 165. [Google Scholar] [CrossRef]
- Clemen, G. Untersuchungen zur Bildung der Vomerspange bei Salamandra salamandra (L.). Roux’s Arch. Dev. Biol. 1979, 185, 305–321. [Google Scholar] [CrossRef] [PubMed]
- Cakir, Y.; Strauch, S.M. Tricaine (MS-222) is a safe anesthetic compound compared to benzocaine and pentobarbital to induce anesthesia in leopard frogs (Rana pipiens). Pharmacol. Rep. 2005, 57, 467–474. [Google Scholar]
- Böck, P. (Ed.) Romeis Histologische Technik, 17th ed.; Urban & Schwarzenberg: München, Germany, 1989. [Google Scholar]
- Schreckenberg, G.M.; Jacobsen, A.G. Normal stages of development of the axolotl Ambystoma mexicanum. Dev. Biol. 1975, 42, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Johannes, M.L.; Klessen, C. Alcianblue/PAS or PAS/Alcianblue? Remarks on a classical technique used in carbohydrate histochemistry. Histochemie 1984, 80, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Kushida, H. A styrene-methacrylate resin embedding method for ultrathin sectioning. J. Electron Microsc. 1961, 10, 16–19. [Google Scholar]
- Gersch, H. Aufbau und Differenzierung des Integuments vom Axolotl auf Grund einer vergleichenden Untersuchung mit Hilfe vitaler und histologischer Färbemethoden. Z. Mikrosk. Anat. Forsch. 1942, 51, 513–554. [Google Scholar]
- Fährmann, W. Die Morphodynamik der Epidermis des Axolotls (Siredon mexicanum SHAW) unter dem Einfluß von exogen appliziertem Thyroxin. II. Die Epidermis während der Metamorphose. Z. Mikrosk. Anat. Forsch. 1971, 83, 535–568. [Google Scholar] [PubMed]
- Fährmann, W. Die Morphodynamik der Epidermis des Axolotls (Siredon mexicanum SHAW) unter dem Einfluß von exogen appliziertem Thyroxin. III. Die Epidermis des metamorphosierten Axolotls. Z. Mikrosk. Anat. Forsch. 1971, 84, 1–25. [Google Scholar]
- Warburg, M.R.; Lewinson, D. Ultrastructure of epidermis of Salamandra salamandra followed throughout ontogenesis. Cell Tissue Res. 1977, 181, 369–393. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, M.; Lewinson, D.; Warburg, M.R. Ultrastructural studies of the epidermal Leydig Cell in larvae of Salamandra salamandra (Caudata, Salamandrida). J. Morphol. 1982, 174, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Kantorek, S.; Clemen, G. The origin and development of Leydig cells in the epithelium of the oropharynx of Ambystoma mexicanum SHAW (Urodela: Ambystomatidae)—An ultrastructural analysis. Zool. Anz. 1991, 227, 13–24. [Google Scholar]
- Echeverri, K.; Tanaka, E.M. Ectoderm to mesoderm lineage switching during axolotl tail regeneration. Science 2002, 298, 1993–1996. [Google Scholar] [CrossRef]
- Wallace, R.B. Regeneration; Academic Press: New York, NY, USA, 1981. [Google Scholar]
- Kragl, M.; Roensch, K.; Nüsslein, I.; Tazaki, A.; Taniguchi, Y.; Tarui, H.; Hayashi, T.; Agata, K.; Tanaka, E.M. Muscle and connective tissue progenitor populations show distinct Twist1 and Twist3 expression profiles during axolotl limb regeneration. Dev. Biol. 2013, 373, 196–204. [Google Scholar] [CrossRef]
- Ferretti, P. Is there a relationship between adult neurogenesis and neuron generation following injury across evolution? Eur. J. Neurosci. 2011, 34, 951–962. [Google Scholar] [CrossRef]
- Seifert, A.W.; Monaghan, J.R.; Smith, M.D.; Pasch, B.; Stier, A.C.; Michonneau, F.; Maden, M. The influence of fundamental traits on mechanisms controlling appendage regeneration. Biol. Rev. 2012, 87, 330–345. [Google Scholar] [CrossRef]
- Norris, D.O.; Platt, J.E. T3- and T4 induced rates of metamorphosis in immature and sexually mature larvae of Ambystoma tigrinum (Amphibia: Caudata). J. Exp. Zool. 1974, 189, 303–310. [Google Scholar] [CrossRef]
- Norris, D.O.; Gern, W.A. Thyroxine-induced activation of hypothalamo-hypophysial axis in neotenic salamander larvae. Science 1976, 194, 525–527. [Google Scholar] [CrossRef]
- Kumar, A.; Godwin, J.W.; Gates, P.B.; Garza-Garcia, A.A.; Brockes, J.P. Molecular basis for the nerve dependence of limb regeneration in an adult vertebrate. Science 2007, 318, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Yin, V.P.; Poss, K.D. New regulators of vertebrate appendage regeneration. Curr. Opin. Genet. Dev. 2008, 18, 381–386. [Google Scholar] [CrossRef]
- Makanae, A.; Mitogawa, K.; Satoh, A. Cooperative inputs of Bmp and Fgf signaling induce tail regeneration in urodele amphibians. Dev. Biol. 2016, 410, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, E.M.; Ferretti, P. Considering the evolution of regeneration in the central nervous system. Nat. Rev. Neurosci. 2009, 10, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, E.A.B.; Vickaryous, M.K. Neural stem/progenitor cells are activated during tail regeneration in the leopard gecko (Eublepharis macularius). J. Comp. Neurol. 2018, 526, 285–309. [Google Scholar] [CrossRef]
- Butler, E.G.; Ward, M.B. Reconstitution of the spinal cord following ablation in urodele larvae. J. Exp. Zool. 1965, 160, 47–66. [Google Scholar] [CrossRef]

| Feature | Ambystoma | Salamandra | ||
|---|---|---|---|---|
| Pre-Metamorphic | Post-Metamorphic | Pre-Metamorphic | Post-Metamorphic | |
| Basal layer of epidermis | Yes, few tonofilaments | Yes, many tonofilaments | Yes, few tonofilaments | Yes, many tonofilaments |
| Intermediate layers of epidermis | Yes, replacement pavement cells | Yes, replacement keratinocytes | Yes, replacement pavement cells | Yes, replacement keratinocytes |
| Superficial layer of epidermis | Apical cell seam (pavement cells) with layers of roundish mucin granules. Apical plasmalemma with microvilli/-ridges. Single or groups of keratinized cells | Layer of living keratinocytes overlain by a stratum corneum | Apical cell seam (pavement cells) with layers of roundish mucin granules. Apical plasmalemma with microvilli/-ridges. Single or groups of keratinized cells | Layer of living keratinocytes overlain by a thick stratum corneum |
| Leydig cells in epidermis | Yes, larger granules than in Salamandra | No | Yes, smaller, more irregularly shaped granules than in Ambystoma | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clemen, G.; Kierdorf, U.; Hermes, M.; Kierdorf, H. The Source of Melanocytes in Ortho- and Heterotopic Tail Regenerates of Axolotls and the Dependence of the Regenerative Response on the Presence of Neural Tissue. Anatomia 2024, 3, 29-49. https://doi.org/10.3390/anatomia3010004
Clemen G, Kierdorf U, Hermes M, Kierdorf H. The Source of Melanocytes in Ortho- and Heterotopic Tail Regenerates of Axolotls and the Dependence of the Regenerative Response on the Presence of Neural Tissue. Anatomia. 2024; 3(1):29-49. https://doi.org/10.3390/anatomia3010004
Chicago/Turabian StyleClemen, Günter, Uwe Kierdorf, Michael Hermes, and Horst Kierdorf. 2024. "The Source of Melanocytes in Ortho- and Heterotopic Tail Regenerates of Axolotls and the Dependence of the Regenerative Response on the Presence of Neural Tissue" Anatomia 3, no. 1: 29-49. https://doi.org/10.3390/anatomia3010004
APA StyleClemen, G., Kierdorf, U., Hermes, M., & Kierdorf, H. (2024). The Source of Melanocytes in Ortho- and Heterotopic Tail Regenerates of Axolotls and the Dependence of the Regenerative Response on the Presence of Neural Tissue. Anatomia, 3(1), 29-49. https://doi.org/10.3390/anatomia3010004