
The Inhibition of Pancreatic α-Amylase by Monomeric, Dimeric and Trimeric Procyanidins Is Dependent upon the Structural Characteristics of Inhibitors and Substrates

 ,
,  and
and
Abstract
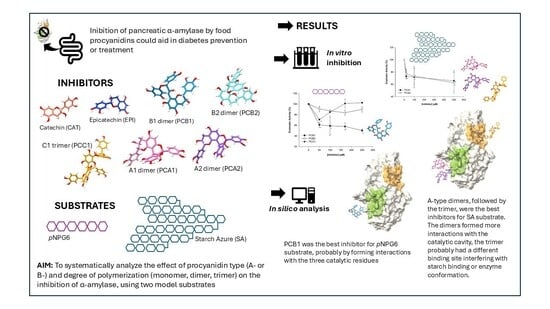
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Enzyme Activity of Pancreatic α-Amylase
2.2.1. p-NPG6 as Substrate
2.2.2. Starch Azure as Substrate
2.3. Inhibition of Pancreatic α-Amylase Enzyme Activity
2.3.1. Determination of the Percent Inhibition with p-NPG6 as Substrate
2.3.2. Determination of the Percent Inhibition with Starch Azure as Substrate
2.4. Molecular Docking Analysis
2.5. Data Analysis
3. Results and Discussion
3.1. Enzyme Activity of α-Amylase Towards Two Model Substrates
3.2. Inhibition of α-Amylase Using p-NPG6 as Substrate
3.3. Inhibition of α-Amylase Using Starch Azure as Substrate
3.4. Molecular Docking
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| p-NPG6 | p-nitrophenyl-α-D-maltohexaoside |
| CAT | Catechin |
| EPI | Epicatechin |
| QUE | Quercetin |
| PCA1 | Procyanidin A1 |
| PCA2 | Procyanidin A2 |
| PCB1 | Procyanidin B1 |
| PCB2 | Procyanidin B2 |
| PCC1 | Procyanidin C1 |
References
- Kanter, J.E.; Bornfeldt, K.E. Impact of Diabetes Mellitus. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1049–1053. [Google Scholar] [CrossRef]
- Kaur, N.; Kumar, V.; Nayak, S.K.; Wadhwa, P.; Kaur, P.; Sahu, S.K. Alpha-amylase as Molecular Target for Treatment of Diabetes Mellitus: A Comprehensive Review. Chem. Biol. Drug Des. 2021, 98, 539–560. [Google Scholar] [CrossRef]
- Martinez-Gonzalez, A.I.; Díaz-Sánchez, Á.G.; de la Rosa, L.A.; Bustos-Jaimes, I.; Alvarez-Parrilla, E. Inhibition of α-Amylase by Flavonoids: Structure Activity Relationship (SAR). Spectrochim. Acta A Mol. Biomol. Spectrosc. 2019, 206, 437–447. [Google Scholar] [CrossRef]
- de la Rosa, L.A.; Moreno-Escamilla, J.O.; Rodrigo-García, J.; Alvarez-Parrilla, E. Phenolic Compounds. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Yahia, E., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 253–271. [Google Scholar]
- Yang, H.; Tuo, X.; Wang, L.; Tundis, R.; Portillo, M.P.; Simal-Gandara, J.; Yu, Y.; Zou, L.; Xiao, J.; Deng, J. Bioactive Procyanidins from Dietary Sources: The Relationship between Bioactivity and Polymerization Degree. Trends Food Sci. Technol. 2021, 111, 114–127. [Google Scholar] [CrossRef]
- Cires, M.J.; Wong, X.; Carrasco-Pozo, C.; Gotteland, M. The Gastrointestinal Tract as a Key Target Organ for the Health-Promoting Effects of Dietary Proanthocyanidins. Front. Nutr. 2017, 3, 57. [Google Scholar] [CrossRef]
- Lam, T.-P.; Tran, N.-V.N.; Pham, L.-H.D.; Lai, N.V.-T.; Dang, B.-T.N.; Truong, N.-L.N.; Nguyen-Vo, S.-K.; Hoang, T.-L.; Mai, T.T.; Tran, T.-D. Flavonoids as Dual-Target Inhibitors against α-Glucosidase and α-Amylase: A Systematic Review of in Vitro Studies. Nat. Prod. Bioprospect 2024, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Visvanathan, R.; Houghton, M.J.; Barber, E.; Williamson, G. Structure-Function Relationships in (Poly)Phenol-Enzyme Binding: Direct Inhibition of Human Salivary and Pancreatic α-Amylases. Food Res. Int. 2024, 188, 114504. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Flores, A.A.; Martinez-Gonzalez, A.I.; Alvarez-Parrilla, E.; Díaz-Sánchez, Á.G.; de la Rosa, L.A.; González-Aguilar, G.A.; Aguilar, C.N. Proanthocyanidins with a Low Degree of Polymerization Are Good Inhibitors of Digestive Enzymes Because of Their Ability to Form Specific Interactions: A Hypothesis. J. Food Sci. 2018, 83, 2895–2902. [Google Scholar] [CrossRef]
- Le Bourvellec, C.; Renard, C.M.G.C. Interactions between Polyphenols and Macromolecules: Effect of Tannin Structure. In Encyclopedia of Food Chemistry; Elsevier: Amsterdam, The Netherlands, 2019; pp. 515–521. [Google Scholar]
- Jiang, C.; Chen, Y.; Ye, X.; Wang, L.; Shao, J.; Jing, H.; Jiang, C.; Wang, H.; Ma, C. Three Flavanols Delay Starch Digestion by Inhibiting α-Amylase and Binding with Starch. Int. J. Biol. Macromol. 2021, 172, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lu, J.; Kan, J.; Wen, X.; Jin, C. Synthesis, Characterization and in Vitro Anti-Diabetic Activity of Catechin Grafted Inulin. Int. J. Biol. Macromol. 2014, 64, 76–83. [Google Scholar] [CrossRef]
- Wang, H.; Du, Y.-J.; Song, H.-C. α-Glucosidase and α-Amylase Inhibitory Activities of Guava Leaves. Food Chem. 2010, 123, 6–13. [Google Scholar] [CrossRef]
- Tao, W.; Pan, H.; Jiang, H.; Wang, M.; Ye, X.; Chen, S. Extraction and Identification of Proanthocyanidins from the Leaves of Persimmon and Loquat. Food Chem. 2022, 372, 130780. [Google Scholar] [CrossRef]
- Darnis, S.; Juge, N.; Guo, X.-J.; Marchis-Mouren, G.; Puigserver, A.; Chaix, J.-C. Molecular Cloning and Primary Structure Analysis of Porcine Pancreatic α-Amylase. BBA-Protein Struct. Mol. Enzymol. 1999, 1430, 281–289. [Google Scholar] [CrossRef]
- Baks, T.; Janssen, A.E.M.; Boom, R.M. A Kinetic Model to Explain the Maximum in A-amylase Activity Measurements in the Presence of Small Carbohydrates. Biotechnol. Bioeng. 2006, 94, 431–440. [Google Scholar] [CrossRef]
- Gaquere-Parker, A.; Taylor, T.; Hutson, R.; Rizzo, A.; Folds, A.; Crittenden, S.; Zahoor, N.; Hussein, B.; Arruda, A. Low Frequency Ultrasonic-Assisted Hydrolysis of Starch in the Presence of α-Amylase. Ultrason. Sonochem 2018, 41, 404–409. [Google Scholar] [CrossRef]
- Rinderknecht, H.; Wilding, P.; Haverback, B.J. A New Method for the Determination of α-Amylase. Experientia 1967, 23, 805. [Google Scholar] [CrossRef] [PubMed]
- Abeliovich, H. On Hill Coefficients and Subunit Interaction Energies. J. Math. Biol. 2016, 73, 1399–1411. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.N. The Hill Equation Revisited: Uses and Misuses. FASEB J. 1997, 11, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Cappiello, M.; Balestri, F.; Moschini, R.; Del-Corso, A.; Mura, U. Apparent Cooperativity and Apparent Hyperbolic Behavior of Enzyme Mixtures Acting on the Same Substrate. J. Enzyme Inhib. Med. Chem. 2016, 31, 1556–1559. [Google Scholar] [CrossRef]
- Ishikura, K.; Nitta, Y.; Watanabe, T. Hydrolysis of Phenyl β-Maltoside Catalyzed by Saccharifying α-Amylase from Bacillus subtilis. J. Biochem. 1977, 81, 1187–1192. [Google Scholar] [PubMed]
- Yoshida, H.; Hiromi, K.; Ono, S. Kinetics and Mechanism of Hydrolysis of Phenyl α-Maltoside by Saccharifying α-Amylase of Bacillus subtilis. J. Biochem. 1969, 65, 741–750. [Google Scholar] [CrossRef]
- Nielsen, J.W.; Kramhøft, B.; Bozonnet, S.; Abou Hachem, M.; Stipp, S.L.S.; Svensson, B.; Willemoës, M. Degradation of the Starch Components Amylopectin and Amylose by Barley α-Amylase 1: Role of Surface Binding Site 2. Arch. Biochem. Biophys. 2012, 528, 1–6. [Google Scholar] [CrossRef]
- Nielsen, M.M.; Seo, E.-S.; Bozonnet, S.; Aghajari, N.; Robert, X.; Haser, R.; Svensson, B. Multi-site Substrate Binding and Interplay in Barley A-amylase 1. FEBS Lett. 2008, 582, 2567–2571. [Google Scholar] [CrossRef]
- Apar, D.K.; Özbek, B. Estamation of Kinetic Parameters for Rice Starch Hydrolysis Inhibited by Added Materials. Chem. Eng. Commun. 2007, 194, 334–344. [Google Scholar] [CrossRef]
- Yoshino, M.; Murakami, K. Analysis of the Substrate Inhibition of Complete and Partial Types. Springerplus 2015, 4, 292. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, C.; Wang, G.; Cao, J.; Yang, X.; Liu, X.; Sun, L. α-Amylase Inhibition of a Certain Dietary Polyphenol Is Predominantly Affected by the Concentration of α-1, 4-Glucosidic Bonds in Starchy and Artificial Substrates. Food Res. Int. 2022, 157, 111210. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, M.; Lu, J.; Li, W.; Wolynes, P.G.; Wang, W. Frustration and the Kinetic Repartitioning Mechanism of Substrate Inhibition in Enzyme Catalysis. J. Phys. Chem. B 2022, 126, 6792–6801. [Google Scholar] [CrossRef] [PubMed]
- Kong, B.-W.; Kim, J.-I.; Kim, M.-J.; Kim, J.C. Porcine Pancreatic α-Amylase Hydrolysis of Native Starch Granules as a Function of Granule Surface Area. Biotechnol. Prog. 2008, 19, 1162–1166. [Google Scholar] [CrossRef]
- Lehoczki, G.; Kandra, L.; Gyémánt, G. The Use of Starch Azure for Measurement of Alpha-Amylase Activity. Carbohydr. Polym. 2018, 183, 263–266. [Google Scholar] [CrossRef]
- Guerra, N.P.; Pastrana Castro, L. Modelling the Effects of Ageing Time of Starch on the Enzymatic Activity of Three Amylolytic Enzymes. Sci. World J. 2012, 2012, 1–13. [Google Scholar] [CrossRef]
- Zhao, L.; Wen, L.; Lu, Q.; Liu, R. Interaction Mechanism between α-Glucosidase and A-Type Trimer Procyanidin Revealed by Integrated Spectroscopic Analysis Techniques. Int. J. Biol. Macromol. 2020, 143, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Barber, E.; Houghton, M.J.; Williamson, G. Flavonoids as Human Intestinal α-Glucosidase Inhibitors. Foods 2021, 10, 1939. [Google Scholar] [CrossRef]
- Shen, H.; Wang, J.; Ao, J.; Hou, Y.; Xi, M.; Cai, Y.; Li, M.; Luo, A. Structure-Activity Relationships and the Underlying Mechanism of α-Amylase Inhibition by Hyperoside and Quercetin: Multi-Spectroscopy and Molecular Docking Analyses. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2023, 285, 121797. [Google Scholar] [CrossRef]
- Peng, Q.; Ma, Y.; Wang, Z.; Wang, J. Inhibition Mechanism of Different Structural Polyphenols against α-Amylase Studied by Solid-State NMR and Molecular Docking. Int. J. Biol. Macromol. 2024, 275, 133757. [Google Scholar] [CrossRef]
- Deogratias, G.; Shadrack, D.M.; Munissi, J.J.E.; Kinunda, G.A.; Jacob, F.R.; Mtei, R.P.; Masalu, R.J.; Mwakyula, I.; Kiruri, L.W.; Nyandoro, S.S. Hydrophobic π-π Stacking Interactions and Hydrogen Bonds Drive Self-Aggregation of Luteolin in Water. J. Mol. Graph. Model. 2022, 116, 108243. [Google Scholar] [CrossRef]
- Barrett, A.H.; Farhadi, N.F.; Smith, T.J. Slowing Starch Digestion and Inhibiting Digestive Enzyme Activity Using Plant Flavanols/Tannins—A Review of Efficacy and Mechanisms. LWT Food Sci. Technol. 2018, 87, 394–399. [Google Scholar] [CrossRef]
- Rue, E.A.; Rush, M.D.; van Breemen, R.B. Procyanidins: A Comprehensive Review Encompassing Structure Elucidation via Mass Spectrometry. Phytochem. Rev. 2018, 17, 1–16. [Google Scholar] [CrossRef]
- Spiegler, V. Anthelmintic A-Type Procyanidins and Further Characterization of the Phenolic Composition of a Root Extract from Paullinia pinnata. Molecules 2020, 25, 2287. [Google Scholar] [CrossRef] [PubMed]
- Prigent, S.V.E.; Voragen, A.G.J.; van Koningsveld, G.A.; Baron, A.; Renard, C.M.G.C.; Gruppen, H. Interactions between Globular Proteins and Procyanidins of Different Degrees of Polymerization. J. Dairy. Sci. 2009, 92, 5843–5853. [Google Scholar] [CrossRef]
- Li, B.; Fu, R.; Tan, H.; Zhang, Y.; Teng, W.; Li, Z.; Tian, J. Characteristics of the Interaction Mechanisms of Procyanidin B1 and Procyanidin B2 with Protein Tyrosine Phosphatase-1B: Analysis by Kinetics, Spectroscopy Methods and Molecular Docking. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2021, 259, 119910. [Google Scholar] [CrossRef]
- de Freitas, V.; Mateus, N. Structural Features of Procyanidin Interactions with Salivary Proteins. J. Agric. Food Chem. 2001, 49, 940–945. [Google Scholar] [CrossRef]
- Sharma, P.; Joshi, T.; Joshi, T.; Chandra, S.; Tamta, S. Molecular Dynamics Simulation for Screening Phytochemicals as α-Amylase Inhibitors from Medicinal Plants. J. Biomol. Struct. Dyn. 2020, 39, 6524–6538. [Google Scholar] [CrossRef]
- Takahama, U.; Hirota, S. Interactions of Flavonoids with α-Amylase and Starch Slowing down Its Digestion. Food Funct. 2018, 9, 677–687. [Google Scholar] [CrossRef]
- Barros, F.; Awika, J.M.; Rooney, L.W. Interaction of Tannins and Other Sorghum Phenolic Compounds with Starch and Effects on in Vitro Starch Digestibility. J. Agric. Food Chem. 2012, 60, 11609–11617. [Google Scholar] [CrossRef]
- Tadera, K.; Minami, Y.; Takamatsu, K.; Matsuoka, T. Inhibition of α-Glucosidase and α-Amylase by Flavonoids. J. Nutr. Sci. Vitaminol. 2006, 52, 149–153. [Google Scholar] [CrossRef]
- Bozonnet, S.; Jensen, M.T.; Nielsen, M.M.; Aghajari, N.; Jensen, M.H.; Kramhøft, B.; Willemoës, M.; Tranier, S.; Haser, R.; Svensson, B. The ‘Pair of Sugar Tongs’ Site on the Non-catalytic Domain C of Barley A-amylase Participates in Substrate Binding and Activity. FEBS J. 2007, 274, 5055–5067. [Google Scholar] [CrossRef]
- Agu, K.C.; Eluehike, N.; Ofeimun, R.O.; Abile, D.; Ideho, G.; Ogedengbe, M.O.; Onose, P.O.; Elekofehinti, O.O. Possible Anti-Diabetic Potentials of Annona muricata (Soursop): Inhibition of α-Amylase and α-Glucosidase Activities. Clin. Phytosci. 2019, 5, 21. [Google Scholar] [CrossRef]
- Fan, J.; Fu, A.; Zhang, L. Progress in Molecular Docking. Quant. Biol. 2019, 7, 83–89. [Google Scholar] [CrossRef]
- Nada, A.A.; Metwally, A.M.; Asaad, A.M.; Celik, I.; Ibrahim, R.S.; Eldin, S.M.S. Synergistic Effect of Potential Alpha-Amylase Inhibitors from Egyptian Propolis with Acarbose Using in Silico and in Vitro Combination Analysis. BMC Complement. Med. Ther. 2024, 24, 65. [Google Scholar] [CrossRef] [PubMed]
- Dai, T.; Chen, J.; McClements, D.J.; Li, T.; Liu, C. Investigation the Interaction between Procyanidin Dimer and α-Glucosidase: Spectroscopic Analyses and Molecular Docking Simulation. Int. J. Biol. Macromol. 2019, 130, 315–322. [Google Scholar] [CrossRef]
- Proença, C.; Freitas, M.; Ribeiro, D.; Tomé, S.M.; Oliveira, E.F.T.; Viegas, M.F.; Araújo, A.N.; Ramos, M.J.; Silva, A.M.S.; Fernandes, P.A.; et al. Evaluation of a Flavonoids Library for Inhibition of Pancreatic α-Amylase towards a Structure–Activity Relationship. J. Enzyme Inhib. Med. Chem. 2019, 34, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G. Possible Effects of Dietary Polyphenols on Sugar Absorption and Digestion. Mol. Nutr. Food Res. 2013, 57, 48–57. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Fit | Vmax | KM | R2 | h |
|---|---|---|---|---|---|
| p-NPG6 | Hill | 0.0383 ± 0.0038 mM min−1 | 3.1447 ± 0.4526 mM | 0.9745 | 1.871 ± 0.2965 |
| Michaelis–Menten | 0.0698 ± 0.0144 mM min−1 | 9.1077 ± 2.9978 mM | 0.9444 | NA | |
| Starch azure | Hill | 0.0167 ± 0.0018 AU min−1 | 1.3601 ± 0.345 mg mL−1 | 0.8983 | 2.3454 ± 1.3201 |
| Michaelis–Menten | 0.0201 ± 0.0036 AU min−1 | 1.6591 ± 1.0779 mg mL−1 | 0.7901 | NA |
| Ligand | ΔG (Kcal mol−1) | Molecular Structure | H-Bonds | π-π Stacking | ||
|---|---|---|---|---|---|---|
| Residues | d (Å) | Residues | d (Å) | |||
| PCA1 | −8.88 |  | Glu352 | 1.88–2.94 | Trp59 | 4.47–4.63 |
| Gln63 | 3.27–3.29 | |||||
| Asp197 | 1.81–2.54 | |||||
| Asp300 | 3.81 | |||||
| PCA2 | −8.42 |  | Arg195 | 2.71 | Trp59 | 3.91–4.41 |
| Asp197 | 2.64 | Tyr62 | 3.66–4.67 | |||
| Asp300 | 1.70 | |||||
| Hse305 | 2.77 | |||||
| Gln63 | 2.98 | |||||
| PCB1 | −8.85 |  | Asp300 | 1.77 | Trp59 | 3.85–4.95 |
| Glu233 | 2.06 | |||||
| Asp197 | 1.94 | |||||
| Tyr151 | 3.51 | |||||
| Lys200 | 2.38 | |||||
| PCB2 | −8.68 |  | Hse305 | 2.34–2.69 | Tyr62 | 3.67–4.50 |
| Asp356 | 2.36 | Trp59 | 3.94–4.26 | |||
| Asp300 | 1.94 | |||||
| Asp197 | 2.69 | |||||
| PCC1 | −9.41 |  | Asp290 | 3.65 | Phe406 | 4.59–4.59 |
| Glu282 | 2.12–2.53 | Tyr2 | 4.60–4.84 | |||
| Asp402 | 2.78 | |||||
| Gly403 | 2.14 | |||||
| Arg421 | 1.97 | |||||
| CAT | −7.58 |  | Asp197 | 1.93–2.11 | Tyr62 | 3.79 |
| Hse101 | 2.37 | Trp59 | 3.97–4.20 | |||
| Asp300 | 1.84 | |||||
| EPI | −7.56 |  | Glu233 | 1.98 | Tyr62 | 4.35 |
| Tyr151 | 2.53 | |||||
| Asp197 | 2.77 | |||||
| Gln63 | 2.41 | |||||
| QUE | −7.74 |  | Hse305 | 2.59 | Trp58 | 3.50–4.29 |
| Asp356 | 3.61 | Tyr62 | 3.72–4.21 | |||
| Gln63 | 2.78 | |||||
| Asp197 | 2.24–2.37 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguilar-López, J.V.; Arras-Gardea, A.V.; Martinez-Gonzalez, A.I.; Alvarez-Parrilla, E.; de la Rosa, L.A. The Inhibition of Pancreatic α-Amylase by Monomeric, Dimeric and Trimeric Procyanidins Is Dependent upon the Structural Characteristics of Inhibitors and Substrates. Appl. Biosci. 2025, 4, 49. https://doi.org/10.3390/applbiosci4040049
Aguilar-López JV, Arras-Gardea AV, Martinez-Gonzalez AI, Alvarez-Parrilla E, de la Rosa LA. The Inhibition of Pancreatic α-Amylase by Monomeric, Dimeric and Trimeric Procyanidins Is Dependent upon the Structural Characteristics of Inhibitors and Substrates. Applied Biosciences. 2025; 4(4):49. https://doi.org/10.3390/applbiosci4040049
Chicago/Turabian StyleAguilar-López, Jocelin Violeta, Ana V. Arras-Gardea, Alejandra I. Martinez-Gonzalez, Emilio Alvarez-Parrilla, and Laura A. de la Rosa. 2025. "The Inhibition of Pancreatic α-Amylase by Monomeric, Dimeric and Trimeric Procyanidins Is Dependent upon the Structural Characteristics of Inhibitors and Substrates" Applied Biosciences 4, no. 4: 49. https://doi.org/10.3390/applbiosci4040049
APA StyleAguilar-López, J. V., Arras-Gardea, A. V., Martinez-Gonzalez, A. I., Alvarez-Parrilla, E., & de la Rosa, L. A. (2025). The Inhibition of Pancreatic α-Amylase by Monomeric, Dimeric and Trimeric Procyanidins Is Dependent upon the Structural Characteristics of Inhibitors and Substrates. Applied Biosciences, 4(4), 49. https://doi.org/10.3390/applbiosci4040049