

Can Caenorhabditis elegans Serve as a Reliable Model for Drug and Nutraceutical Discovery?


Abstract
1. Introduction
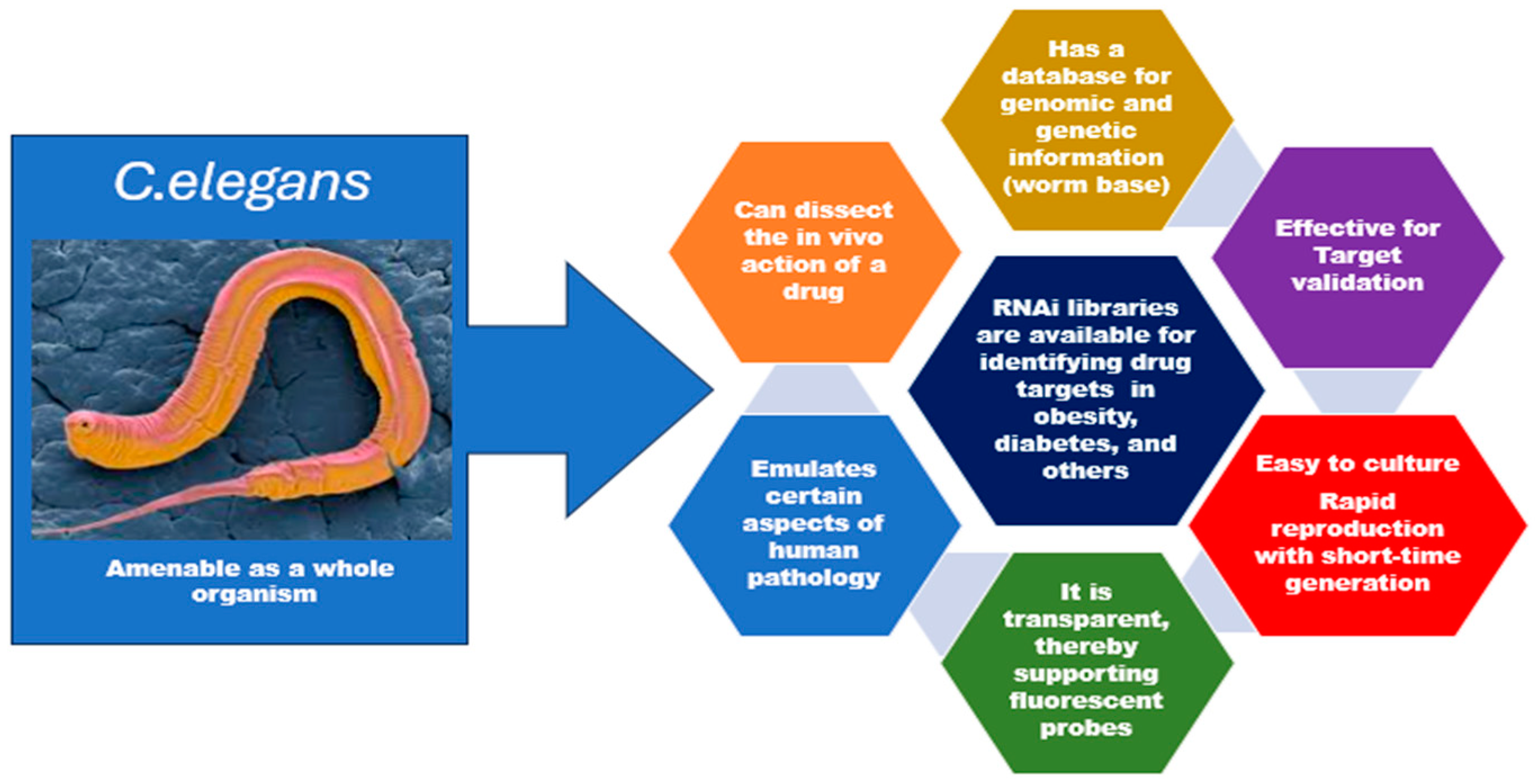
1.1. Molecular Basis for Using C. elegans in Biomedical Research
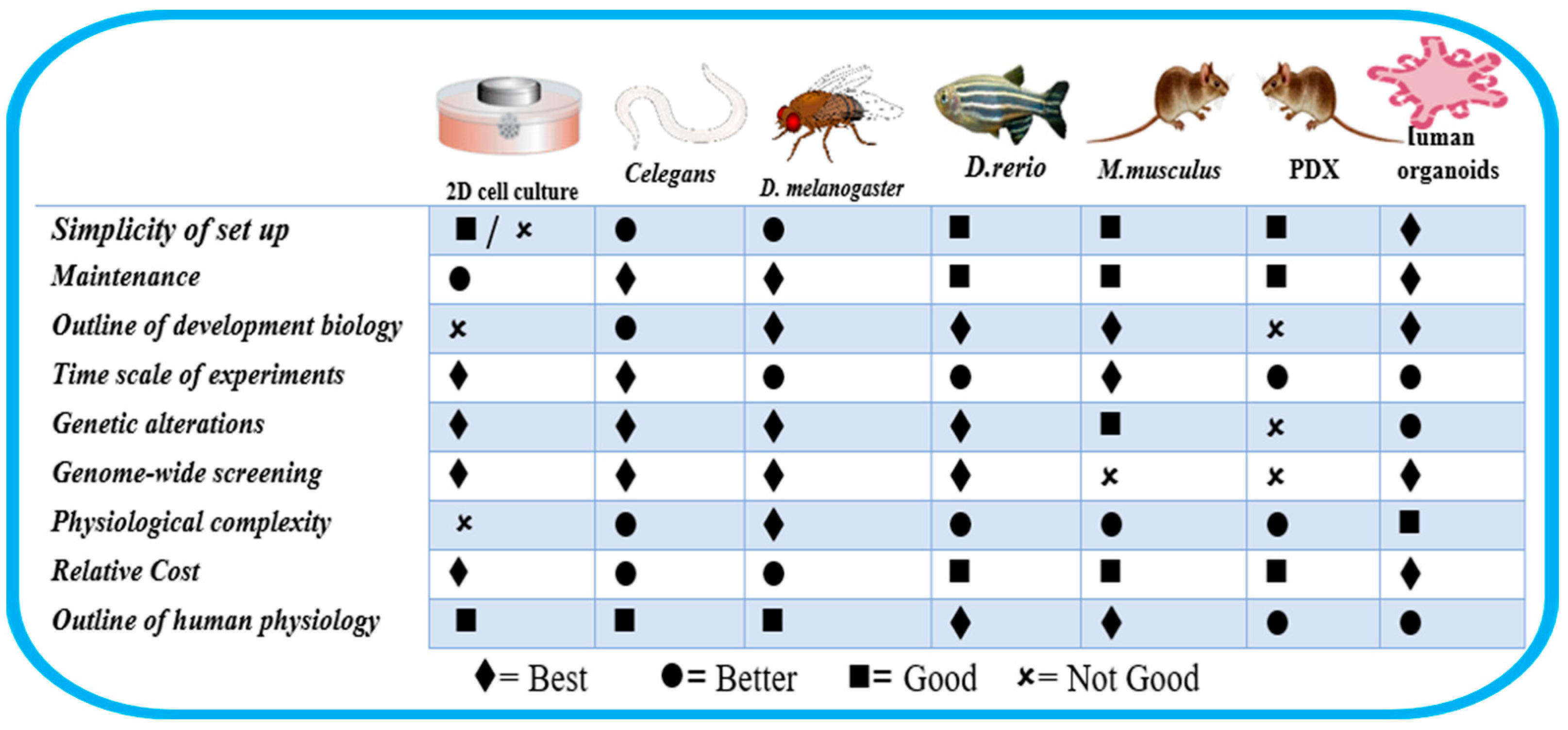
1.2. Is C. elegans Suitable as a Reliable Model?
2. C. elegans for Early Drug Development and Target Identification
3. C. elegans for Evaluation of Drug Effects and Possible Action Mechanisms
4. C. elegans in Pharmacokinetic Studies

{kind=link}
{kind=link}
| S/N | Technical Approaches | Applications | Drug Class | Limitations | Ref |
|---|---|---|---|---|---|
| 1 | Fluorescence microscopy and plate reader | Quantifying organic anion uptake and accumulation | Organic anions (Fluorescein) | Limited to fluorescent compounds | [51] |
| 2 | Epifluorescent microscopy | Quantifying organic anion uptake and accumulation | Anticancer (Doxorubicin) | Limited to fluorescent compounds | [52] |
| 3 | In-silico docking and ADMET analysis | Predicting drug interactions and properties | Herbicide (Paraquat), Antiparkinson (Levodopa) | Requires experimental validation | [81,82] |
| 4 | High-Performance Liquid Chromatography-Tandem Mass Spectrometry (HPLC-MS/MS) | Metabolite identification and characterization | Anthelmintic (Albendazole) | Requires specialized equipment | [83] |
| 5 | Whole genome microarrays, Reverse Transcription Quantitative Polymerase Chain Reaction (RT-QPCR), HPLC-MS | Gene expression analysis and metabolite identification | Anthelmintics (Ivermectin, Albendazole) | Complex data interpretation | [84] |
| 6 | Liquid Chromatography with Diode Array Detection (LC-DAD) and LC-MS/MS | Detailed metabolite characterization | Isoflavone (Genistein) | Requires specialized equipment | [85] |
| 7 | Toxicokinetics experiments, one-compartment model | Studying drug absorption, distribution, metabolism, and excretion | Antidepressant (Fluoxetine) | Model assumptions may not fully represent biological complexity | [79,82] |
| 8 | Behavioral analysis | Assessing drug effects on nematode locomotion, and potential resistance. | Anthelmintic (Pyrantel) | Indirect measure of pharmacokinetics | [76,85] |
5. C. elegans and Nutraceutical Development
6. C. elegans in Pharmaceutical Development in Anti-Aging Research
6.1. C. elegans in the Identification of Genes That Affect Lifespan
6.2. Dietary Restriction and Specific Gene Mutation
6.3. Natural Chemicals and Cellular Pathways
6.4. Developing New Drugs to Improve Age-Linked Diseases
7. C. elegans in Screening Novel Antimicrobial Compounds
8. C. elegans in Lipid Quantification and Antiobesity
8.1. Genetic Regulators of Lipid Metabolism
8.2. Experimental Approaches and Applications of C. elegans in Lipid Antiobesity Studies
8.3. Applications in Obesity Research
9. C. elegans in Neurodegenerative Diseases
10. C. elegans in the Study of Gut Health Permeability
11. Limitations of Using C. elegans Models in Pharmaceutical Research
Model Variation and Reproducibility
12. C. elegans and Toxicity Issues: A Comparison with Mammals
13. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Venkatesan, P. Re-emergence of infectious diseases associated with the past. Lancet Microbe 2021, 2, e140. [Google Scholar] [CrossRef]
- Puri, V.; Nagpal, M.; Singh, I.; Singh, M.; Dhingra, G.A.; Huanbutta, K.; Dheer, D.; Sharma, A.; Sangnim, T. A comprehensive review on nutraceuticals: Therapy support and formulation challenges. Nutrients 2022, 14, 4637. [Google Scholar] [CrossRef] [PubMed]
- Esther, B.; Rapport, L.; Lockwood, B. 1. What is a nutraceutical. Stroke 2018, 13, 57. [Google Scholar]
- Sharifi-Rad, J.; El Rayess, Y.; Rizk, A.A.; Sadaka, C.; Zgheib, R.; Zam, W.; Sestito, S.; Rapposelli, S.; Neffe-Skocińska, K.; Zielińska, D.; et al. Turmeric and its major compound curcumin on health: Bioactive effects and safety profiles for food, pharmaceutical, biotechnological and medicinal applications. Front. Pharmacol. 2020, 11, 550909. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda-Crespo, D.; Reguera, R.M.; Rojo-Vázquez, F.; Balaña-Fouce, R.; Martínez-Valladares, M. Drug discovery technologies: Caenorhabditis elegans as a model for anthelmintic therapeutics. Med. Res. Rev. 2020, 40, 1715–1753. [Google Scholar] [CrossRef]
- Huang, Y.; Huang, Z.; Tang, Z.; Chen, Y.; Huang, M.; Liu, H.; Huang, W.; Ye, Q.; Jia, B. Research progress, challenges, and breakthroughs of organoids as disease models. Front. Cell Dev. Biol. 2021, 9, 574. [Google Scholar] [CrossRef] [PubMed]
- Carretero, M.; Solis, G.M.; Petrascheck, M. C. elegans as a model for drug discovery. Curr. Top. Med. Chem. 2017, 17, 2067–2076. [Google Scholar] [CrossRef]
- Morgan, P.; Brown, D.G.; Lennard, S.; Anderton, M.J.; Barrett, J.C.; Eriksson, U.; Fidock, M.; Hamrén, B.; Johnson, A.; March, R.E.; et al. Impact of a five-dimensional framework on R&D productivity at AstraZeneca. Nat. Rev. Drug Discov. 2018, 17, 167–181. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Kaneko, M.; Narukawa, M. Approval success rates of drug candidates based on target, action, modality, application, and their combinations. Clin. Transl. Sci. 2021, 14, 1113–1122. [Google Scholar] [CrossRef]
- Grimstein, M.; Yang, Y.; Zhang, X.; Grillo, J.; Huang, S.-M.; Zineh, I.; Wang, Y. Physiologically based pharmacokinetic modeling in regulatory science: An update from the US Food and Drug Administration’s Office of Clinical Pharmacology. J. Pharm. Sci. 2019, 108, 21–25. [Google Scholar] [CrossRef]
- Pognan, F.; Beilmann, M.; Boonen, H.C.M.; Czich, A.; Dear, G.; Hewitt, P.; Mow, T.; Oinonen, T.; Roth, A.; Steger-Hartmann, T.; et al. The evolving role of investigative toxicology in the pharmaceutical industry. Nat. Rev. Drug Discov. 2023, 22, 317–335. [Google Scholar] [CrossRef]
- Antoni, D.; Burckel, H.; Josset, E.; Noel, G. Three-dimensional cell culture: A breakthrough in vivo. Int. J. Mol. Sci. 2015, 16, 5517–5527. [Google Scholar] [CrossRef]
- Kapałczyńska, M.; Kolenda, T.; Przybyła, W.; Zajączkowska, M.; Teresiak, A.; Filas, V.; Ibbs, M.; Bliźniak, R.; Łuczewski, L.; Lamperska, K. 2D and 3D cell cultures—A comparison of different types of cancer cell cultures. Arch. Med. Sci. 2018, 14, 910–919. [Google Scholar] [CrossRef] [PubMed]
- Naranjo-Galindo, F.J.; Ai, R.; Fang, E.F.; Nilsen, H.L.; SenGupta, T. C. elegans as an animal model to study the intersection of DNA repair, aging and neurodegeneration. Front. Aging 2022, 3, 916118. [Google Scholar] [CrossRef] [PubMed]
- Cheruku, G.R.; Wilson, C.V.; Raviendran, S.; Xiao, Q. Recent Advances and Future Perspectives in Vascular Organoids and Vessel-on-Chip. Organoids 2024, 3, 203–246. [Google Scholar] [CrossRef]
- Gerli, M.F.M.; Calà, G.; Beesley, M.A.; Sina, B.; Tullie, L.; Sun, K.Y.; Panariello, F.; Michielin, F.; Davidson, J.R.; Russo, F.M.; et al. Single-cell guided prenatal derivation of primary fetal epithelial organoids from human amniotic and tracheal fluids. Nat. Med. 2024, 30, 875–887. [Google Scholar] [CrossRef]
- Singh, R.; Gholipourmalekabadi, M.; Shafikhani, S.H. Animal models for type 1 and type 2 diabetes: Advantages and limitations. Front. Endocrinol. 2024, 15, 1359685. [Google Scholar] [CrossRef]
- Simon, F.; Oberhuber, A.; Schelzig, H. Advantages and disadvantages of different animal models for studying ischemia/reperfusion injury of the spinal cord. Eur. J. Vasc. Endovasc. Surg. 2015, 49, 744. [Google Scholar] [CrossRef]
- O’Reilly, L.P.; Luke, C.J.; Perlmutter, D.H.; Silverman, G.A.; Pak, S.C. C. elegans in high-throughput drug discovery. Adv. Drug Deliv. Rev. 2014, 69–70, 247–253. [Google Scholar] [CrossRef]
- Kim, J.; Koo, B.-K.; Knoblich, J.A. Human organoids: Model systems for human biology and medicine. Nat. Rev. Mol. Cell Biol. 2020, 21, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Ma, L.; Mishra, A.; Turnbull, J.E.; Tu, H. Editorial: C. elegans as an emerging model of pharmacological innovation. Front. Pharmacol. 2022, 13, 1029752. [Google Scholar] [CrossRef] [PubMed]
- Zarroug, S.H.O.; Bajaman, J.S.; Hamza, F.N.; Saleem, R.A.; Abdalla, H.K. Caenorhabditis elegans as an in vivo model for the discovery and development of natural plant-based antimicrobial compounds. Pharmaceuticals 2023, 16, 1070. [Google Scholar] [CrossRef]
- Giunti, S.; Andersen, N.; Rayes, D.; De Rosa, M.J. Drug discovery: Insights from the invertebrate Caenorhabditis elegans. Pharmacol. Res. Perspect. 2021, 9, e00721. [Google Scholar] [CrossRef] [PubMed]
- Kropp, P.A.; Bauer, R.; Zafra, I.; Graham, C.; Golden, A. Caenorhabditis elegans for rare disease modeling and drug discovery: Strategies and strengths. Dis. Model. Mech. 2021, 14, dmm049010. [Google Scholar] [CrossRef]
- Kim, Y.; Park, Y.; Hwang, J.; Kwack, K. Comparative genomic analysis of the human and nematode Caenorhabditis elegans uncovers potential reproductive genes and disease associations in humans. Physiol. Genom. 2018, 50, 1002–1014. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhou, B.; Oshiro-Rapley, N.; Li, M.; Paulo, J.A.; Webster, C.M.; Mou, F.; Kacergis, M.C.; Talkowski, M.E.; Carr, C.E.; et al. An ancient, unified mechanism for metformin growth inhibition in C. elegans and cancer. Cell 2016, 167, 1705–1718.e13. [Google Scholar] [CrossRef]
- Paschinger, K.; Yan, S.; Wilson, I.B.H. N-glycomic Complexity in Anatomical Simplicity: Caenorhabditis elegans as a Non-model Nematode? Front. Mol. Biosci. 2019, 6, 9. [Google Scholar] [CrossRef]
- Zhang, S.; Li, F.; Zhou, T.; Wang, G.; Li, Z. Caenorhabditis elegans as a useful model for studying aging mutations. Front. Endocrinol. 2020, 11, 554994. [Google Scholar] [CrossRef]
- Roussos, A.; Kitopoulou, K.; Borbolis, F.; Palikaras, K. Caenorhabditis elegans as a model system to study human neurodegenerative disorders. Biomolecules 2023, 13, 478. [Google Scholar] [CrossRef]
- Athar, F.; Templeman, N.M. C. elegans as a model organism to study female reproductive health. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2022, 266, 111152. [Google Scholar] [CrossRef]
- Higgins, D.P.; Weisman, C.M.; Lui, D.S.; A D’agostino, F.; Walker, A.K. Defining characteristics and conservation of poorly annotated genes in Caenorhabditis elegans using WormCat 2.0. Genetics 2022, 221, iyac085. [Google Scholar] [CrossRef] [PubMed]
- A Calarco, J.; Taylor, S.R.; Miller, D.M. Detecting gene expression in Caenorhabditis elegans. Genetics 2024, 229, iyae167. [Google Scholar] [CrossRef]
- Li, G.; Gong, J.; Liu, J.; Liu, J.; Li, H.; Hsu, A.-L.; Liu, J.; Xu, X.S. Genetic and pharmacological interventions in the aging motor nervous system slow motor aging and extend life span in C. elegans. Sci. Adv. 2019, 5, eaau5041. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.; Alvarez-Illera, P.; Santo-Domingo, J.; Fonteriz, R.I.; Montero, M. Modeling Alzheimer’s Disease in Caenorhabditis elegans. Biomedicines 2022, 10, 288. [Google Scholar] [CrossRef]
- Romussi, S.; Giunti, S.; Andersen, N.; De Rosa, M.J. C. elegans: A prominent platform for modeling and drug screening in neurological disorders. Expert Opin. Drug Discov. 2024, 19, 565–585. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.K.; Priya, A.; Malik, Z.; Thanaraj, T.A.; Singh, A.K.; Mago, P.; Ghosh, C.; Shalimar; Tandon, R.; Chaturvedi, R. A bioinformatics approach to elucidate conserved genes and pathways in C. elegans as an animal model for cardiovascular research. Sci. Rep. 2024, 14, 7471. [Google Scholar] [CrossRef]
- Xia, X. Bioinformatics and Drug Discovery. Curr. Top. Med. Chem. 2017, 17, 1709–1726. [Google Scholar] [CrossRef]
- Meissner, F.; Geddes-McAlister, J.; Mann, M.; Bantscheff, M. The emerging role of mass spectrometry-based proteomics in drug discovery. Nat. Rev. Drug Discov. 2022, 21, 637–654. [Google Scholar] [CrossRef]
- Sun, D.; Gao, W.; Hu, H.; Zhou, S. Why 90% of clinical drug development fails and how to improve it? Acta Pharm. Sin. B 2022, 12, 3049–3062. [Google Scholar] [CrossRef]
- Caldwell, K.A.; Willicott, C.W.; Caldwell, G.A. Modeling neurodegeneration in Caenorhabditis elegans. Dis. Model. Mech. 2020, 13, dmm046110. [Google Scholar] [CrossRef]
- Corsi, A.K.; Wightman, B.; Chalfie, M. A transparent window into biology: A primer on Caenorhabditis elegans. Genetics 2015, 200, 387–407. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhu, Y.; Song, B.; Fu, R.; Zhou, Y. The In vivo toxicity assessments of water-dispersed fluorescent silicon nanoparticles in Caenorhabditis elegans. Int. J. Environ. Res. Public Health 2022, 19, 4101. [Google Scholar] [CrossRef] [PubMed]
- Sofela, S.; Sahloul, S.; Song, Y.-A. Biophysical analysis of drug efficacy on C. elegans models for neurodegenerative and neuromuscular diseases. PLoS ONE 2021, 16, e0246496. [Google Scholar] [CrossRef]
- Meneely, P.M.; Dahlberg, C.L.; Rose, J.K. Working with worms: Caenorhabditis elegans as a model organism. Curr. Protoc. Essent. Lab. Tech. 2019, 19, e35. [Google Scholar] [CrossRef]
- Hashmi, S.; Wang, Y.; Parhar, R.S.; Collison, K.S.; Conca, W.; Al-Mohanna, F.; Gaugler, R. A C. elegans model to study human metabolic regulation. Nutr. Metab. 2013, 10, 31. [Google Scholar] [CrossRef] [PubMed]
- E Escobar, I.; White, A.; Kim, W.; Mylonakis, E. New antimicrobial bioactivity against multidrug-resistant gram-positive bacteria of kinase inhibitor IMD0354. Antibiotics 2020, 9, 665. [Google Scholar] [CrossRef]
- Jim, V.; Slauter, R. Overview of ADME Science. In A Comprehensive Guide to Toxicology in Nonclinical Drug Development; Academic Press: Cambridge, MA, USA, 2024; pp. 49–82. [Google Scholar]
- van der Most, M.A.; Estruch, I.M.; Brink, N.W.v.D. Contrasting dose response relationships of neuroactive antidepressants on the behavior of C. elegans. Ecotoxicol. Environ. Saf. 2023, 250, 114493. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.; Zheng, F.; Zhang, W.; Li, L.; Li, Y.; Hu, H.; Wu, Y.; Bao, W.; Li, G.; Wang, Q.; et al. Oxidation and antioxidation of natural products in the model organism Caenorhabditis elegans. Antioxidants 2022, 11, 705. [Google Scholar] [CrossRef] [PubMed]
- Harold, A.L.; Woock, A.E.; Bridges, A.A.; Cecile, J.P. Organic Anion Transport in Caenorhabditis elegans. FASEB J. 2013, 27, 732.1. [Google Scholar] [CrossRef]
- Amirthagunabalasingam, S.; Papaluca, A.; Harihar, T.; Ramotar, D. Imaging the Pharynx to Measure the Uptake of Doxorubicin in Caenorhabditis elegans. Bio-Protocol 2017, 7, e2291. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, H.; Jin, S.; Lu, Y.; Peng, Y.; Zhao, L.; Wang, X. Cannabidiol protects against Alzheimer’s disease in C. elegans via ROS scavenging activity of its phenolic hydroxyl groups. Eur. J. Pharmacol. 2022, 919, 174829. [Google Scholar] [CrossRef]
- Blackwell, T.K.; Sewell, A.K.; Wu, Z.; Han, M. TOR signaling in Caenorhabditis elegans development, metabolism, and aging. Genetics 2019, 213, 329–360. [Google Scholar] [CrossRef]
- Suárez, G.; Alcántara, I.; Salinas, G. Caenorhabditis elegans as a valuable model for the study of anthelmintic pharmacodynamics and drug-drug interactions: The case of ivermectin and eprinomectin. Front. Pharmacol. 2022, 13, 984905. [Google Scholar] [CrossRef]
- Sohrabi, S.; Mor, D.E.; Kaletsky, R.; Keyes, W.; Murphy, C.T. High-throughput behavioral screen in C. elegans reveals Parkinson’s disease drug candidates. Commun. Biol. 2021, 4, 203. [Google Scholar] [CrossRef] [PubMed]
- Chrienova, Z.; Rysanek, D.; Oleksak, P.; Stary, D.; Bajda, M.; Reinis, M.; Mikyskova, R.; Novotny, O.; Andrys, R.; Skarka, A.; et al. Discovery of small molecule mechanistic target of rapamycin inhibitors as anti-aging and anti-cancer therapeutics. Front. Aging Neurosci. 2022, 14, 1048260. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-P.; Ma, X.; Liu, J.-L.; Liu, A.-L. Exploring the potential use of Caenorhabditis elegans as an animal model for evaluating chemical-induced intestinal dysfunction. Toxicol. Appl. Pharmacol. 2024, 493, 117140. [Google Scholar] [CrossRef] [PubMed]
- Eric, D.L.; Morishita, M.; Hirotsu, T. C. elegans as a Powerful Tool for Cancer Screening. Biomedicines 2022, 10, 2371. [Google Scholar] [CrossRef]
- Hussar, P. Apoptosis regulators bcl-2 and caspase-3. Encyclopedia 2022, 2, 1624–1636. [Google Scholar] [CrossRef]
- Horowitz, L.B.; Shaham, S. Apoptotic and Nonapoptotic Cell Death in Caenorhabditis elegans Development. Annu. Rev. Genet. 2024, 58, 113–134. [Google Scholar] [CrossRef]
- La Marca, J.E.; Kelly, G.L.; Strasser, A.; Diepstraten, S.T. Don’t fear the reaper: The role of regulated cell death in tumorigenesis and BH3-mimetics for cancer therapy. Dev. Cell 2024, 59, 2532–2548. [Google Scholar] [CrossRef]
- Kovalchuk, I. Genome stability in Caenorhabditis elegans. In Genome Stability, 2nd ed.; Academic Press: Cambridge, MA, USA, 2021; pp. 177–200. [Google Scholar] [CrossRef]
- Eldesouky, H.E.; Salama, E.A.; Hazbun, T.R.; Mayhoub, A.S.; Seleem, M.N. Ospemifene displays broad-spectrum synergistic interactions with itraconazole through potent interference with fungal efflux activities. Sci. Rep. 2020, 10, 6089. [Google Scholar] [CrossRef]
- Jamiu, A.T.; Albertyn, J.; Sebolai, O.; Gcilitshana, O.; Pohl, C.H. Inhibitory effect of polyunsaturated fatty acids alone or in combination with fluconazole on Candida krusei biofilms in vitro and in Caenorhabditis elegans. Med. Mycol. 2021, 59, 1225–1237. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Zhang, Y.; Liu, M.; Huang, Y.; Wang, S.; An, J.; Wang, Y.; Shang, Y. The joint effects of nanoplastics and TBBPA on neurodevelopmental toxicity in Caenorhabditis elegans. Toxicol. Res. 2023, 12, 76–85. [Google Scholar] [CrossRef]
- Ikenaka, K.; Tsukada, Y.; Giles, A.C.; Arai, T.; Nakadera, Y.; Nakano, S.; Kawai, K.; Mochizuki, H.; Katsuno, M.; Sobue, G.; et al. A behavior-based drug screening system using a Caenorhabditis elegans model of motor neuron disease. Sci. Rep. 2019, 9, 10104. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Quan, J.I.; Li, L.; Kinghorn, K.J.; Ivanov, D.K.; Tain, L.S.; Slack, C.; Kerr, F.; Nespital, T.; Thornton, J.; Hardy, J.; et al. Lithium Promotes Longevity through GSK3/NRF2-Dependent Hormesis. Cell Rep. 2016, 15, 638–650. [Google Scholar] [CrossRef] [PubMed]
- Cabreiro, F.; Au, C.; Leung, K.Y.; Vergara-Irigaray, N.; Cochemé, H.M.; Noori, T.; Weinkove, D.; Schuster, E.; Greene, N.D.; Gems, D. Metformin retards aging in C. elegans by altering microbial folate and methionine metabolism. Cell 2013, 153, 22839. [Google Scholar] [CrossRef]
- Chen, J.; Ou, Y.; Li, Y.; Hu, S.; Shao, L.-W.; Liu, Y. Metformin extends C. elegans lifespan through lysosomal pathway. eLife 2017, 6, e31268. [Google Scholar] [CrossRef]
- Ahmad, W.; Ebert, P.R. Metformin Attenuates Aβ Pathology Mediated Through Levamisole Sensitive Nicotinic Acetylcholine Receptors in a C. elegans Model of Alzheimer’s Disease. Mol. Neurobiol. 2017, 54, 5427–5439. [Google Scholar] [CrossRef]
- Saewanee, N.; Praputpittaya, T.; Malaiwong, N.; Chalorak, P.; Meemon, K. Neuroprotective effect of metformin on dopaminergic neurodegeneration and α-synuclein aggregation in C. elegans model of Parkinson’s disease. Neurosci. Res. 2021, 162, 13–21. [Google Scholar] [CrossRef]
- Udono, H.; Nishida, M. Metformin-ROS-Nrf2 connection in the host defense mechanism against oxidative stress, apoptosis, cancers, and ageing. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2022, 1866, 130171. [Google Scholar] [CrossRef]
- Markaki, M.; Tavernarakis, N. Caenorhabditis elegans as a model system for human diseases. Curr. Opin. Biotechnol. 2020, 63, 118–125. [Google Scholar] [CrossRef]
- Göethel, G.; Augsten, L.V.; das Neves, G.M.; Gonçalves, I.L.; de Souza, J.P.S.; Garcia, S.C.; Eifler-Lima, V.L. The role of alternative toxicological trials in drug discovery programs. The case of Caenorhabditis elegans and other methods. Curr. Med. Chem. 2022, 29, 5270–5288. [Google Scholar] [CrossRef] [PubMed]
- Hartman, J.H.; Widmayer, S.J.; Bergemann, C.M.; King, D.E.; Morton, K.S.; Romersi, R.F.; Jameson, L.E.; Leung, M.C.K.; Andersen, E.C.; Taubert, S.; et al. Xenobiotic metabolism and transport in Caenorhabditis elegans. J. Toxicol. Environ. Health Part B 2021, 24, 51–94. [Google Scholar] [CrossRef]
- Sharma, N.; Au, V.; Martin, K.; Edgley, M.L.; Moerman, D.; Mains, P.E.; Gilleard, J.S. Multiple UDP glycosyl-transferases modulate benzimidazole drug sensitivity in the nematode Caenorhabditis elegans in an additive manner. Int. J. Parasitol. 2024, 54, 535–549. [Google Scholar] [CrossRef]
- Raza, A.; Williams, A.R.; Abeer, M.M. Importance of ABC Transporters in the Survival of Parasitic Nematodes and the Prospect for the Development of Novel Control Strategies. Pathogens 2023, 12, 755. [Google Scholar] [CrossRef] [PubMed]
- Laing, S.T.; Ivens, A.; Butler, V.; Ravikumar, S.P.; Laing, R.; Woods, D.J.; Gilleard, J.S. The transcriptional response of Caenorhabditis elegans to ivermectin exposure identifies novel genes involved in the response to reduced food intake. PLoS ONE 2012, 7, e31367. [Google Scholar] [CrossRef]
- van der Most, M.A.; Bakker, W.; Wesseling, S.; Brink, N.W.v.D. Toxicokinetics of the Antidepressant Fluoxetine and Its Active Metabolite Norfluoxetine in Caenorhabditis elegans and Their Comparative Potency. Environ. Sci. Technol. 2024, 58, 3129–3140. [Google Scholar] [CrossRef]
- Shen, M. Pain Chemogenomics Knowledgebase (PAIN-CKB) for Systems Pharmacology Target Mapping and PBPK Modeling Investigation of Opioid Drug-Drug Interactions. Doctoral Dissertation, University of Pittsburgh, Pittsburgh, PA, USA, 2020. [Google Scholar]
- Jatti, T.; Maniyal, N.; Mouli, S.; Shenoy, R.; Bhat, S. Molecular Docking and ADMET of Levodopa against Leucine-Rich Repeat Kinases, and In-Vitro Mobility Analysis in C. elegans for Parkinson’s Disease. In Proceedings of the 2023 IEEE 23rd International Conference on Bioinformatics and Bioengineering (BIBE), Dayton, OH, USA, 4–6 December 2023. [Google Scholar]
- Stasiuk, S.J.; MacNevin, G.; Workentine, M.L.; Gray, D.; Redman, E.; Bartley, D.; Morrison, A.; Sharma, N.; Colwell, D.; Ro, D.K.; et al. Similarities and differences in the biotransformation and transcriptomic responses of Caenorhabditis elegans and Haemonchus contortus to five different benzimidazole drugs. Int. J. Parasitol. Drugs Drug Resist. 2019, 11, 13–29. [Google Scholar] [CrossRef]
- Kim, D.K.; Park, J.Y.; Kang, Y.J.; Khang, D. Drug repositioning of metformin encapsulated in PLGA combined with photothermal therapy ameliorates rheumatoid arthritis. Int. J. Nanomed. 2023, 18, 7267–7285. [Google Scholar] [CrossRef]
- Soukup, S.T.; Spanier, B.; Grünz, G.; Bunzel, D.; Daniel, H.; Kulling, S.E. Formation of Phosphoglycosides in Caenorhabditis elegans: A Novel Biotransformation Pathway. PLoS ONE 2012, 7, e46914. [Google Scholar] [CrossRef]
- Sheeja, M.D.; Prasanth, M.I.; Brimson, J.M.; Verma, K.; Prasansuklab, A.; Tencomnao, T. Hibiscus sabdariffa extract protects HT-22 cells from glutamate-induced neurodegeneration by upregulating glutamate transporters and exerts lifespan extension in C. elegans via DAF-16 mediated pathway. Nutr. Healthy Aging 2021, 6, 229–247. [Google Scholar] [CrossRef]
- Ha, N.M.; Tran, S.H.; Shim, Y.-H.; Kang, K. Caenorhabditis elegans as a powerful tool in natural product bioactivity research. Appl. Biol. Chem. 2022, 65, 18. [Google Scholar] [CrossRef]
- Awuchi, C.G.; Okpala, C.O.R. Natural nutraceuticals, especially functional foods, their major bioactive components, formulation, and health benefits for disease prevention: An overview. J. Food Bioact. 2022, 19, 97–123. [Google Scholar] [CrossRef]
- Loo, J.; Bana, M.A.F.S.; Tan, J.K.; Goon, J.A. Effect of dietary restriction on health span in Caenorhabditis elegans: A systematic review. Exp. Gerontol. 2023, 182, 112294. [Google Scholar] [CrossRef]
- Long, N.P.; Kang, J.S.; Kim, H.M. Caenorhabditis elegans: A model organism in the toxicity assessment of environmental pollutants. Environ. Sci. Pollut. Res. 2023, 30, 39273–39287. [Google Scholar] [CrossRef]
- Nitin, V.; Sharma, S.; Thakur, N.; Kaur, N.; Dua, K. Nanotherapeutics for Alz-heimer’s disease using metal nanocomposites. Met. Nanocompos. Nanother. Oxidative Stress-Induc. Metab. Disord. 2024, 1, 372–391. [Google Scholar]
- Aranaz, P.; Peña, A.; Vettorazzi, A.; Fabra, M.J.; Martínez-Abad, A.; López-Rubio, A.; Pera, J.; Parladé, J.; Castellari, M.; Milagro, F.I.; et al. Grifola frondosa (Maitake) Extract Reduces Fat Accumulation and Improves Health Span in C. elegans through the DAF-16/FOXO and SKN-1/NRF2 Signalling Pathways. Nutrients 2021, 13, 3968. [Google Scholar] [CrossRef]
- Lippi, L.; Uberti, F.; Folli, A.; Turco, A.; Curci, C.; d’Abrosca, F.; de Sire, A.; Invernizzi, M. Impact of nutraceuticals and dietary supplements on mitochondria modifications in healthy aging: A systematic review of randomized controlled trials. Aging Clin. Exp. Res. 2022, 34, 2659–2674. [Google Scholar] [CrossRef]
- Deledda, A.; Giordano, E.; Velluzzi, F.; Flore, G.; Franceschelli, S.; Speranza, L.; Ripari, P. Mitochondrial aging and senolytic natural products with protective potential. Int. J. Mol. Sci. 2022, 23, 16219. [Google Scholar] [CrossRef]
- Iwanir, S.; Ruach, R.; Itskovits, E.; Pritz, C.O.; Bokman, E.; Zaslaver, A. Irrational behavior in C. elegans arises from asymmetric modulatory effects within single sensory neurons. Nat. Commun. 2019, 10, 3202. [Google Scholar] [CrossRef]
- Shen, P.; Zhang, R.; McClements, D.J.; Park, Y. Nanoemulsion-based delivery systems for testing nutraceutical efficacy using Caenorhabditis elegans: Demonstration of curcumin bioaccumulation and body-fat reduction. Food Res. Int. 2019, 120, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Huang, X.; Dou, L.; Yan, M.; Shen, T.; Tang, W.; Li, J. Aging and aging-related diseases: From molecular mechanisms to interventions and treatments. Signal Transduct. Target. Ther. 2022, 7, 391. [Google Scholar] [CrossRef]
- Uno, M.; Nishida, E. Lifespan-regulating genes in C. elegans. Npj Aging Mech. Dis. 2016, 2, 16010. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, H.; Hu, Q.; Wang, L.; Liu, J.; Zheng, Z.; Zhang, W.; Ren, J.; Zhu, F.; Liu, G.-H. Epigenetic regulation of aging: Implications for interventions of aging and diseases. Signal Transduct. Target. Ther. 2022, 7, 374. [Google Scholar] [CrossRef]
- Pedro, F.J.; Pitt, B.; Zannad, F. Histone deacetylase inhibitors for cardiovascular conditions and healthy longevity. Lancet Healthy Longev. 2021, 2, e371–e379. [Google Scholar] [CrossRef] [PubMed]
- Revtovich, A.V.; Lee, R.; Kirienko, N.V. Interplay between mitochondria and diet mediates pathogen and stress resistance in Caenorhabditis elegans. PLoS Genet. 2019, 15, e1008011. [Google Scholar] [CrossRef]
- Greer, E.L.; Brunet, A. Different dietary restriction regimens extend lifespan by both independent and overlapping genetic pathways in C. elegans. Aging Cell 2009, 8, 113–127. [Google Scholar] [CrossRef]
- Ewald, C.Y.; Castillo-Quan, J.I.; Blackwell, T.K. Untangling longevity, dauer, and healthspan in Caenorhabditis elegans insulin/IGF-1-signalling. Gerontology 2017, 64, 96–104. [Google Scholar] [CrossRef]
- Hahm, J.-H.; Seo, H.-D.; Jung, C.H.; Ahn, J. Longevity through diet restriction and immunity. BMB Rep. 2023, 56, 537–544. [Google Scholar] [CrossRef]
- Zia, A.; Farkhondeh, T.; Pourbagher-Shahri, A.M.; Samarghandian, S. The role of curcumin in aging and senescence: Molecular mechanisms. Biomed. Pharmacother. 2021, 134, 111119. [Google Scholar] [CrossRef]
- Green, C.L.; Lamming, D.W.; Fontana, L. Molecular mechanisms of dietary restriction promoting health and longevity. Nat. Rev. Mol. Cell Biol. 2021, 23, 56–73. [Google Scholar] [CrossRef] [PubMed]
- Cuong, V.T.; Chen, W.; Shi, J.; Zhang, M.; Yang, H.; Wang, N.; Yang, S.; Li, J.; Yang, P.; Fei, J. The anti-oxidation and anti-aging effects of Ganoderma lucidum in Caenorhabditis elegans. Exp. Gerontol. 2019, 117, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Chen, Y.; Lin, Y.; Wang, X.; Hu, L.; Cao, Y.; Chen, Y. Antistress and anti-aging activities of Caenorhabditis elegans were enhanced by Momordica saponin extract. Eur. J. Nutr. 2020, 60, 1819–1832. [Google Scholar] [CrossRef]
- Bahrami, A.; Montecucco, F.; Carbone, F.; Sahebkar, A. Effects of curcumin on aging: Molecular mechanisms and experimental evidence. BioMed. Res. Int. 2021, 2021, 8972074. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Lin, Y.; Xiao, J.; Lan, Y.; Cao, Y.; Chen, Y. Effect of Momordica saponin and Cyclocarya paliurus pol-ysaccharide-enriched beverages on oxidative stress and fat accumulation in Caenorhabditis elegans. J. Sci. Food Agric. 2021, 101, 3366–3375. [Google Scholar] [CrossRef] [PubMed]
- Khayatan, D.; Razavi, S.M.; Arab, Z.N.; Hosseini, Y.; Niknejad, A.; Momtaz, S.; Abdolghaffari, A.H.; Sathyapalan, T.; Jamialahmadi, T.; Kesharwani, P.; et al. Superoxide dismutase: A key target for the neuroprotective effects of curcumin. Mol. Cell. Biochem. 2023, 479, 693–705. [Google Scholar] [CrossRef]
- Turer, B.Y.; Sanlier, N. Relationship of Curcumin with Aging and Alzheimer and Parkinson Disease, the Most Prevalent Age-Related Neurodegenerative Diseases: A Narrative Review. Nutr. Rev. 2024, 83, e1243–e1258. [Google Scholar] [CrossRef]
- Park, H.-E.H.; Hwang, W.; Ham, S.; Kim, E.; Altintas, O.; Park, S.; Son, H.G.; Lee, Y.; Lee, D.; Heo, W.D.; et al. A PTEN variant uncouples longevity from impaired fitness in Caenorhabditis elegans with reduced insulin/IGF-1 signaling. Nat. Commun. 2021, 12, 5631. [Google Scholar] [CrossRef]
- Yoon, D.S.; Cha, D.S.; Choi, Y.; Lee, J.W.; Lee, M. MPK-1/ERK is required for the full activity of resveratrol in extended lifespan and reproduction. Aging Cell 2018, 18, e12867. [Google Scholar] [CrossRef]
- Miller, B.C.; Mathai, M.; Yadav, H.; Jain, S. Geroprotective potential of microbiome modulators in the Caenorhabditis elegans model. GeroScience 2023, 46, 129–151. [Google Scholar] [CrossRef]
- Solis, G.M.; Kardakaris, R.; Valentine, E.R.; Bar-Peled, L.; Chen, A.L.; Blewett, M.M.; A McCormick, M.; Williamson, J.R.; Kennedy, B.; Cravatt, B.F.; et al. Translation attenuation by minocycline enhances longevity and proteostasis in old post-stress-responsive organisms. eLife 2018, 7, e40314. [Google Scholar] [CrossRef]
- Janssens, G.E.; Lin, X.-X.; Millan-Ariño, L.; Kavšek, A.; Sen, I.; Seinstra, R.I.; Stroustrup, N.; Nollen, E.A.; Riedel, C.G. Transcriptomics-based screening identifies pharmacological inhibition of Hsp90 as a means to defer aging. Cell Rep. 2019, 27, 467–480.e6. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Chen, X.; Kong, X.-M.; Liu, T.-W.; Feng, X.-Q.; Chen, F.-E.; Zhuang, Z.-H. Anti-aging effect of methylurolithin A and its amide derivatives on nematode Caenorhabditis elegans. Tetrahedron Lett. 2023, 118, 154389. [Google Scholar] [CrossRef]
- Bonuccelli, G.; Brooks, D.R.; Shepherd, S.; Sotgia, F.; Lisanti, M.P. Antibiotics that target mitochondria extend lifespan in C. elegans. Aging 2023, 15, 11764–11781. [Google Scholar] [CrossRef]
- Yu, X.; Li, H.; Lin, D.; Guo, W.; Xu, Z.; Wang, L.; Guan, S. Ginsenoside prolongs the lifespan of C. elegans via lipid metabolism and activating the stress response signaling pathway. Int. J. Mol. Sci. 2021, 22, 9668. [Google Scholar] [CrossRef]
- Snell, T.W.; Johnston, R.K.; Matthews, A.B.; Zhou, H.; Gao, M.; Skolnick, J. Repurposed FDA-approved drugs targeting genes influencing aging can extend lifespan and healthspan in rotifers. Biogerontology 2018, 19, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Bar-Ziv, R.; Frakes, A.E.; Higuchi-Sanabria, R.; Bolas, T.; Frankino, P.A.; Gildea, H.K.; Metcalf, M.G.; Dil-lin, A. Measurements of Physiological Stress Responses in C. elegans. J. Vis. Exp. 2020, 21, e61001. [Google Scholar]
- Dues, D.J.; Andrews, E.K.; Schaar, C.E.; Bergsma, A.L.; Senchuk, M.M.; Van Raamsdonk, J.M. Aging causes decreased resistance to multiple stresses and a failure to activate specific stress response pathways. Aging 2016, 8, 777–795. [Google Scholar] [CrossRef]
- Kyriakou, E.; Taouktsi, E.; Syntichaki, P. The thermal stress coping network of the nematode Caenorhabditis elegans. Int. J. Mol. Sci. 2022, 23, 14907. [Google Scholar] [CrossRef]
- Szigeti, B.; Gleeson, P.; Vella, M.; Khayrulin, S.; Palyanov, A.; Hokanson, J.; Currie, M.; Cantarelli, M.; Idili, G.; Larson, S. OpenWorm: An open-science approach to modeling Caenorhabditis elegans. Front. Comput. Neurosci. 2014, 8, 137. [Google Scholar] [CrossRef]
- Norris, D.D. Design and Application of Alternative Transgenic Reporter Systems in Caenorhabditis elegans. Master’s Thesis, Michigan Technological University, Houghton, MI, USA, 2024. [Google Scholar]
- Nance, J.; Frøkjær-Jensen, C. The Caenorhabditis elegans transgenic toolbox. Genetics 2019, 212, 959–990. [Google Scholar] [CrossRef] [PubMed]
- Ayon, N.J. High-throughput screening of natural product and synthetic molecule libraries for antibacterial drug discovery. Metabolites 2023, 13, 625. [Google Scholar] [CrossRef]
- Foster, K.J.; Cheesman, H.K.; Liu, P.; Peterson, N.D.; Anderson, S.M.; Pukkila-Worley, R. Innate immunity in the C. elegans intestine is programmed by a neuronal regulator of AWC olfactory neuron development. Cell Rep. 2020, 31, 107478. [Google Scholar] [CrossRef] [PubMed]
- Mahesh, R.; Ilangovan, P.; Nongbri, D.; Suchiang, K. Probiotics interactions and the modulation of major signalling pathways in host model organism Caenorhabditis elegans. Indian J. Microbiol. 2021, 61, 404–416. [Google Scholar] [CrossRef] [PubMed]
- Midkiff, D.; San-Miguel, A. Microfluidic technologies for high throughput screening through sorting and on-chip culture of C. elegans. Molecules 2019, 24, 4292. [Google Scholar] [CrossRef]
- Samsudin, S.; Al-Talib, H.; Zain, Z.M.; Murugaiah, C. Panton-Valentine Leukocidin-Positive Staphylococcus aureus: A successful infectious enemy? Int. Med. J. 2020, 27, 540–544. [Google Scholar]
- Niu, H.; Gu, J.; Zhang, Y. Bacterial persisters: Molecular mechanisms and therapeutic development. Signal Transduct. Target. Ther. 2024, 9, 174. [Google Scholar] [CrossRef]
- Moy, T.I.; Conery, A.L.; Larkins-Ford, J.; Wu, G.; Mazitschek, R.; Casadei, G.; Lewis, K.; Carpenter, A.E.; Ausubel, F.M. High-throughput screen for novel antimicrobials using a whole animal infection model. ACS Chem. Biol. 2009, 4, 527–533. [Google Scholar] [CrossRef]
- Tharmalingam, N.; Rajmuthiah, R.; Kim, W.; Fuchs, B.B.; Jeyamani, E.; Kelso, M.J.; Mylonakis, E. Antibacterial properties of four novel hit compounds from a methicillin-resistant Staphylococcus aureus–Caenorhabditis elegans high-throughput screen. Microb. Drug Resist. 2018, 24, 666–674. [Google Scholar] [CrossRef]
- Xie, H.; Zhan, Y.; Chen, X.; Zeng, Q.; Chen, D.; Liang, J. Brevinin-2 drug family—New applied peptide candidates against methicillin-resistant Staphylococcus aureus and their effects on Lys-7 expression of innate immune pathway DAF-2/DAF-16 in Caenorhabditis elegans. Appl. Sci. 2018, 8, 2627. [Google Scholar] [CrossRef]
- Iatsenko, J.; Yim, J.; Schroeder, F.; Iatsenko, R.S. B. subtilis GS67 protects C. elegans from Gram-positive pathogens via fengycin-mediated microbial antagonism. Curr. Biol. 2014, 24, 2720–2727. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.; A Yehye, W.; Rahman, N.A.; Tan, M.-W.; Nathan, S. Discovery of potential anti-infectives against Staphylococcus aureus using a Caenorhabditis elegans infection model. BMC Complement. Altern. Med. 2014, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Moy, T.I.; Ball, A.R.; Anklesaria, Z.; Casadei, G.; Lewis, K.; Ausubel, F.M. Identification of novel antimicrobials using a live-animal infection model. Proc. Natl. Acad. Sci. USA 2006, 103, 10414–10419. [Google Scholar] [CrossRef] [PubMed]
- Rajamuthiah, R.; Fuchs, B.B.; Jayamani, E.; Kim, Y.; Larkins-Ford, J.; Conery, A.; Ausubel, F.M.; Mylonakis, E. Whole animal automated platform for drug discovery against multi-drug-resistant Staphylococcus aureus. PLoS ONE 2014, 9, e89189. [Google Scholar] [CrossRef]
- Kim, W.; Zhu, W.; Hendricks, G.L.; Van Tyne, D.; Steele, A.D.; Keohane, C.E.; Fricke, N.; Conery, A.L.; Shen, S.; Pan, W.; et al. A new class of synthetic retinoid antibiotics effective against bacterial persisters. Nature 2018, 556, 103–107. [Google Scholar] [CrossRef]
- Wu, K.; E Simor, A.; Vearncombe, M.; McClure, J.-A.; Zhang, K. A Caenorhabditis elegans host model correlates with invasive disease caused by Staphylococcus aureus recovered during an outbreak in neonatal intensive care. Can. J. Infect. Dis. Med. Microbiol. 2012, 23, 130–134. [Google Scholar] [CrossRef]
- Spreafico, R.; Soriaga, L.B.; Grosse, J.; Virgin, H.W.; Telenti, A. Advances in genomics for drug development. Genes 2020, 11, 942. [Google Scholar] [CrossRef]
- Ke, W.; Reed, J.N.; Yang, C.; Higgason, N.; Rayyan, L.; Wählby, C.; Carpenter, A.E.; Civelek, M.; O’rourke, E.J. Genes in human obesity loci are causal obesity genes in C. elegans. PLoS Genet. 2021, 17, e1009736. [Google Scholar] [CrossRef]
- Subhadra, M.; Mir, D.A.; Ankita, K.; Sindunathy, M.; Kishore, H.D.; Ravichandiran, V.; Balamurugan, K. Exploring diabesity pathophysiology through proteomic analysis using Caenorhabditis elegans. Front. Endocrinol. 2024, 15, 1383520. [Google Scholar] [CrossRef]
- Goh, G.Y.S.; Winter, J.J.; Bhanshali, F.; Doering, K.R.S.; Lai, R.; Lee, K.; Veal, E.A.; Taubert, S. NHR-49/HNF4 integrates regulation of fatty acid metabolism with a protective transcriptional response to oxidative stress and fasting. Aging Cell 2018, 17, e12743. [Google Scholar] [CrossRef]
- Witting, M.; Schmitt-Kopplin, P. The Caenorhabditis elegans lipidome: A primer for lipid analysis in Caenorhabditis elegans. Arch. Biochem. Biophys. 2016, 589, 27–37. [Google Scholar] [CrossRef]
- Shen, P.; Yue, Y.; Zheng, J.; Park, Y. Caenorhabditis elegans: A convenient in vivo model for assessing the impact of food bioactive compounds on obesity, aging, and Alzheimer’s disease. Annu. Rev. Food Sci. Technol. 2018, 9, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.C.; Dunbar, T.L.; Nargund, A.M.; Haynes, C.M.; Troemel, E.R. The C. elegans CCAAT-enhancer-binding protein gamma is required for surveillance immunity. Cell Rep. 2016, 14, 1581–1589. [Google Scholar] [CrossRef]
- Xu, X.-Y.; Hu, J.-P.; Wu, M.-M.; Wang, L.-S.; Fang, N.-Y. CCAAT/enhancer-binding protein CEBP-2 controls fat consumption and fatty acid desaturation in Caenorhabditis elegans. Biochem. Biophys. Res. Commun. 2015, 468, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Maulik, M.; Mitra, S.; Basmayor, A.M.; Lu, B.; Taylor, B.E.; Bult-Ito, A. Genetic silencing of fatty acid desaturases modulates α-synuclein toxicity and neuronal loss in Parkinson-like models of C. elegans. Front. Aging Neurosci. 2019, 11, 207. [Google Scholar] [CrossRef]
- An, L.; Fu, X.; Chen, J.; Ma, J. Application of Caenorhabditis elegans in lipid metabolism research. Int. J. Mol. Sci. 2023, 24, 1173. [Google Scholar] [CrossRef] [PubMed]
- Barros, A.G.d.A.; Bridi, J.C.; de Souza, B.R.; Júnior, C.d.C.; Torres, K.C.d.L.; Malard, L.; Jorio, A.; de Miranda, D.M.; Ashrafi, K.; Romano-Silva, M.A. Dopamine signaling regulates fat content through β-oxidation in Caenorhabditis elegans. PLoS ONE 2014, 9, e85874. [Google Scholar] [CrossRef]
- Berthoud, H.; Seeley, R.J.; Roberts, S.B. Physiology of energy intake in the weight-reduced state. Obesity 2021, 29, S25–S30. [Google Scholar] [CrossRef]
- Ekraminasab, S.; Dolatshahi, M.; Sabahi, M.; Mardani, M.; Rashedi, S. The interactions between adipose tissue secretions and Parkinson’s disease: The role of leptin. Eur. J. Neurosci. 2022, 55, 873–891. [Google Scholar] [CrossRef]
- Soares, M.V.; Viçozzi, G.P.; Kuhn, E.C.; Weishaupt, A.-K.; Kubens, L.; Bornhorst, J.; Avila, D.S. Neurotoxicology of organic environmental toxicants using Caenorhabditis elegans as a model. In Advances in Neurotoxicology; Academic Press: Cambridge, MA, USA, 2023; Volume 9, pp. 149–180. [Google Scholar] [CrossRef]
- Dowden, H.; Munro, J. Trends in clinical success rates and therapeutic focus. Nat. Rev. Drug Discov. 2019, 18, 495–496. [Google Scholar] [CrossRef]
- Liang, J.J.H.; McKinnon, I.A.; Rankin, C.H. The contribution of C. elegans neurogenetics to understanding neurodegenerative diseases. In Nature’s Gift to Neuroscience; CRC Press: Boca Raton, FL, USA, 2022; pp. 317–338. [Google Scholar]
- Rani, N.; Alam, M.M.; Jamal, A.; Bin Ghaffar, U.; Parvez, S. Caenorhabditis elegans: A transgenic model for studying age-associated neurodegenerative diseases. Ageing Res. Rev. 2023, 91, 102036. [Google Scholar] [CrossRef]
- Teo, M.Y.; Rathkopf, D.E.; Kantoff, P. Treatment of advanced prostate cancer. Annu. Rev. Med. 2019, 70, 479–499. [Google Scholar] [CrossRef]
- Ahmad, W. Glucose enrichment impair neurotransmission and induce Aβ oligomerization that cannot be reversed by manipulating O-β-GlcNAcylation in the C. elegans model of Alzheimer’s disease. J. Nutr. Biochem. 2022, 108, 109100. [Google Scholar] [CrossRef]
- Chau, D.D.-L.; Ng, L.L.-H.; Zhai, Y.; Lau, K.-F. Amyloid precursor protein and its interacting proteins in neurodevelopment. Biochem. Soc. Trans. 2023, 51, 1647–1659. [Google Scholar] [CrossRef]
- Jing, Y. Regulation of Caenorhabditis elegans model in Alzheimer’s Disease. In E3S Web of Conferences; EDP Sciences: Les Ulis, France, 2020; Volume 185, p. 03043. [Google Scholar] [CrossRef]
- He, C.-L.; Tang, Y.; Wu, J.-M.; Long, T.; Yu, L.; Teng, J.-F.; Qiu, W.-Q.; Pan, R.; Yu, C.-L.; Qin, D.-L.; et al. Chlorogenic acid delays the progression of Parkinson’s disease via autophagy induction in Caenorhabditis elegans. Nutr. Neurosci. 2021, 26, 11–24. [Google Scholar] [CrossRef]
- Abusrair, A.H.; Elsekaily, W.; Bohlega, S. Tremor in Parkinson’s Disease: From Pathophysiology to Advanced Therapies. Tremor. Other Hyperkinetic Mov. 2022, 12, 29. [Google Scholar] [CrossRef]
- Poewe, W.; Volc, D.; Seppi, K.; Medori, R.; Lührs, P.; Kutzelnigg, A.; Djamshidian, A.; Thun-Hohenstein, C.; Meissner, W.G.; Rascol, O.; et al. Safety and tolerability of active immunotherapy targeting α-synuclein with PD03A in patients with early Parkinson’s disease: A randomized, placebo-controlled, phase 1 study. J. Park. Dis. 2021, 11, 1079–1089. [Google Scholar] [CrossRef]
- Ma, L.; Zhao, Y.; Chen, Y.; Cheng, B.; Peng, A.; Huang, K. Caenorhabditis elegans as a model system for target identification and drug screening against neurodegenerative diseases. Eur. J. Pharmacol. 2018, 819, 169–180. [Google Scholar] [CrossRef]
- Gaeta, A.L.; Caldwell, K.A.; Caldwell, G.A. Found in translation: The utility of C. elegans alpha-synuclein models of Parkinson’s disease. Brain Sci. 2019, 9, 73. [Google Scholar] [CrossRef]
- Fan, H.-C.; Ho, L.-I.; Chi, C.-S.; Chen, S.-J.; Peng, G.-S.; Chan, T.-M.; Lin, S.-Z.; Harn, H.-J. Polyglutamine (PolyQ) Diseases: Genetics to Treatments. Cell Transplant. 2014, 23, 441–458. [Google Scholar] [CrossRef]
- Zanni, E.; De Bellis, G.; Bracciale, M.P.; Broggi, A.; Santarelli, M.L.; Sarto, M.S.; Palleschi, C.; Uccelletti, D. Graphite Nanoplatelets and Caenorhabditis elegans: Insights from an in Vivo Model. Nano Lett. 2012, 12, 2740–2744. [Google Scholar] [CrossRef]
- Golegaonkar, S.; Tabrez, S.S.; Pandit, A.; Sethurathinam, S.; Jagadeeshaprasad, M.G.; Bansode, S.; Sampathkumar, S.; Kulkarni, M.J.; Mukhopadhyay, A. Rifampicin reduces advanced glycation end products and activates DAF-16 to increase lifespan in Caenorhabditis elegans. Aging Cell 2015, 14, 463–473. [Google Scholar] [CrossRef]
- Abu-Elfotuh, K.; Al-Najjar, A.H.; Mohammed, A.A.; Aboutaleb, A.S.; Badawi, G.A. Fluoxetine ameliorates Alzheimer’s disease progression and prevents the exacerbation of cardiovascular dysfunction of socially isolated depressed rats through activation of Nrf2/HO-1 and hindering TLR4/NLRP3 inflammasome signaling pathway. Int. Immunopharmacol. 2022, 104, 108488. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef]
- Link, P. Natural Products Against Neurodegenerative Diseases: Effects in the Model Organism Caenorhabditis elegans. Ph.D. Thesis, Ruperto-Carola University of Heidelberg, Heidelberg, Germany, 2018. [Google Scholar] [CrossRef]
- Sadananda, G.; Velmurugan, J.D.; Subramaniam, J.R. DMSO Delays Alzheimer Disease Causing Aβ-induced Paralysis in C. elegans Through Modulation of Glutamate/Acetylcholine Neurotransmission. Ann. Neurosci. 2021, 28, 55–64. [Google Scholar] [CrossRef]
- Pandey, T.; Smita, S.S.; Mishra, A.; Sammi, S.R.; Pandey, R. Swertiamarin, a secoiridoid glycoside modulates nAChR and AChE activity. Exp. Gerontol. 2020, 138, 111010. [Google Scholar] [CrossRef]
- Dirksen, P.; Assié, A.; Zimmermann, J.; Zhang, F.; Tietje, A.-M.; Marsh, S.A.; Félix, M.-A.; Shapira, M.; Kaleta, C.; Schulenburg, H.; et al. CeMbio-The Caenorhabditis elegans microbiome resource. G3 Genes Genomes Genet. 2020, 10, 3025–3039. [Google Scholar] [CrossRef]
- Cheddadi, R.; Yeramilli, V.; Gamra, I.; Davies, J.; Tanner, S.; Martin, C. Intestinal Development and Gut Disease: Contributions From the Caenorhabditis elegans Model. J. Surg. Res. 2024. [Google Scholar] [CrossRef]
- Dirksen, P.; Marsh, S.A.; Braker, I.; Heitland, N.; Wagner, S.; Nakad, R.; Mader, S.; Petersen, C.; Kowallik, V.; Rosenstiel, P.; et al. The native microbiome of the nematode Caenorhabditis elegans: Gateway to a new host-microbiome model. BMC Biol. 2016, 14, 38. [Google Scholar] [CrossRef]
- Zhang, J.; Holdorf, A.D.; Walhout, A.J. C. elegans and its bacterial diet as a model for systems-level understanding of host–microbiota interactions. Curr. Opin. Biotechnol. 2017, 46, 74–80. [Google Scholar] [CrossRef]
- Singh, A.; Luallen, R.J. Understanding the factors regulating host–microbiome interactions using Caenorhabditis elegans. Philos. Trans. R. Soc. B Biol. Sci. 2024, 379, 20230059. [Google Scholar] [CrossRef]
- Sim, S.; Hibberd, M.L. Caenorhabditis elegans susceptibility to gut Enterococcus faecalis infection is associated with fat metabolism and epithelial junction integrity. BMC Microbiol. 2016, 16, 6. [Google Scholar] [CrossRef]
- Kumar, A.; Baruah, A.; Tomioka, M.; Iino, Y.; Kalita, M.C.; Khan, M. Caenorhabditis elegans: A model to understand host–microbe interactions. Cell. Mol. Life Sci. 2019, 77, 1229–1249. [Google Scholar] [CrossRef]
- Tsolis, R.M.; Bäumler, A.J. Gastrointestinal host-pathogen interaction in the age of microbiome research. Curr. Opin. Microbiol. 2020, 53, 78–89. [Google Scholar] [CrossRef]
- Ren, M.; Zhao, L.; Ding, X.; Krasteva, N.; Rui, Q.; Wang, D. Developmental basis for an intestinal barrier against the toxicity of graphene oxide. Part. Fibre Toxicol. 2018, 15, 26. [Google Scholar] [CrossRef]
- Vancamelbeke, M.; Vermeire, S. The intestinal barrier: A fundamental role in health and disease. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef]
- Cheng, C.K.; Huang, Y. The gut-cardiovascular connection: New era for cardiovascular therapy. Med. Rev. 2021, 1, 23–46. [Google Scholar] [CrossRef]
- Massier, L.; Chakaroun, R.; Kovacs, P.; Heiker, J.T. Blurring the picture in leaky gut research: How shortcomings of zonulin as a biomarker mislead the field of intestinal permeability. Gut 2020, 70, 1801–1802. [Google Scholar] [CrossRef]
- Kim, M.R.; Cho, S.-Y.; Lee, H.J.; Kim, J.Y.; Nguyen, U.T.T.; Ha, N.M.; Choi, K.Y.; Cha, K.H.; Kim, J.-H.; Kim, W.K.; et al. Schisandrin C improves leaky gut conditions in intestinal cell monolayer, organoid, and nematode models by increasing tight junction protein expression. Phytomedicine 2022, 103, 154209. [Google Scholar] [CrossRef]
- Rutter, J.W.; Ozdemir, T.; Galimov, E.R.; Quintaneiro, L.M.; Rosa, L.; Thomas, G.M.; Cabreiro, F.; Barnes, C.P. Detecting Changes in the Caenorhabditis elegans Intestinal Environment Using an Engineered Bacterial Biosensor. ACS Synth. Biol. 2019, 8, 2620–2628. [Google Scholar] [CrossRef]
- Wang, S.; Ahmadi, S.; Nagpal, R.; Jain, S.; Mishra, S.P.; Kavanagh, K.; Zhu, X.; Wang, Z.; McClain, D.A.; Kritchevsky, S.B.; et al. Lipoteichoic acid from the cell wall of a heat killed Lactobacillus paracasei D3-5 ameliorates aging-related leaky gut, inflammation and improves physical and cognitive functions: From C. elegans to mice. GeroScience 2019, 42, 333–352. [Google Scholar] [CrossRef]
- Kim, J.; Moon, Y. Worm-based alternate assessment of probiotic intervention against gut barrier infection. Nutrients 2019, 11, 2146. [Google Scholar] [CrossRef]
- Raposo, V.L. Safe Drugs Versus Innovative Drugs (Can We Have Both?). Adv. Pharm. Bull. 2019, 10, 334–337. [Google Scholar] [CrossRef]
- Anupama, S.; Kaduskar, B.; Reddy, K.C.; Kumsta, C.; Bier, E.; Hansen, M.; Chalasani, S.H. Conserved neuropeptidergic regulation of intestinal integrity in invertebrate models of aging. bioRxiv 2022. [Google Scholar] [CrossRef]
- Xiong, H.; Pears, C.; Woollard, A. An enhanced C. elegans based platform for toxicity assessment. Sci. Rep. 2017, 7, 9839. [Google Scholar] [CrossRef]
- Calahorro, F.; Holden-Dye, L.; O’connor, V. Impact of drug solvents on C. elegans pharyngeal pumping. Toxicol. Rep. 2021, 8, 1240–1247. [Google Scholar] [CrossRef]
- Zhang, A.; Hsiung, K.C.; Kern, C.C.; Wang, Y.; Girtle, A.L.; Xu, N.; Gems, D. Unraveling effects of anti-aging drugs on C. elegans using liposomes. GeroScience 2023, 45, 1583–1603. [Google Scholar] [CrossRef]
- Flavel, M.R.; Mechler, A.; Shahmiri, M.; Mathews, E.R.; E Franks, A.; Chen, W.; Zanker, D.; Xian, B.; Gao, S.; Luo, J.; et al. Growth of Caenorhabditis elegans in defined media is dependent on presence of particulate matter. G3 Genes|Genomes|Genet. 2018, 8, 567–575. [Google Scholar] [CrossRef]
- Ramage, G.; Borghi, E.; Rodrigues, C.F.; Kean, R.; Williams, C.; Lopez-Ribot, J. Our current clinical under-standing of Candida biofilms: Where are we two decades on? APMIS 2023, 131, 636–653. [Google Scholar] [CrossRef]
- Wang, E.; Jiang, Y.; Zhao, C. Structural and physiological functions of Caenorhabditis elegans epidermis. Heliyon 2024, 10, e38680. [Google Scholar] [CrossRef]
- Sundaram, M.V.; Pujol, N. The Caenorhabditis elegans cuticle and precuticle: A model for studying dynamic apical extracellular matrices in vivo. Genetics 2024, 227, iyae072. [Google Scholar] [CrossRef]
- Hunt, P.R. The C. elegans model in toxicity testing. J. Appl. Toxicol. 2016, 37, 50–59. [Google Scholar] [CrossRef]
- Sarma, G.P.; Lee, C.W.; Portegys, T.; Ghayoomie, V.; Jacobs, T.; Alicea, B.; Cantarelli, M.; Currie, M.; Gerkin, R.C.; Gingell, S.; et al. OpenWorm: Overview and recent advances in integrative biological simulation of Caenorhabditis elegans. Philos. Trans. R. Soc. B 2018, 373, 20170382. [Google Scholar] [CrossRef]
- Petrascheck, M.; Miller, D.L. Computational Analysis of Lifespan Experiment Reproducibility. Front. Genet. 2017, 8, 92. [Google Scholar] [CrossRef]
- FDA. The C. elegans Model in Toxicity Testing. Toxicology Research. 2022. Available online: https://www.fda.gov/food/toxicology-research/c-elegans-model-toxicity-testing#:~:text=elegans%20tests%20is%20comparable%20to,%2C%20sensory%2C%20and%20neuromuscular%20systems (accessed on 15 March 2025).
- Urban, N.D.; Cavataio, J.P.; Berry, Y.; Vang, B.; Maddali, A.; Sukpraphrute, R.J.; Schnell, S.; Truttmann, M.C. Explaining inter-lab variance in C. elegans N2 lifespan: Making a case for standardized reporting to enhance reproducibility. Exp. Gerontol. 2021, 156, 111622. [Google Scholar] [CrossRef]
- van der Zanden, T.M.; Mooij, M.G.; Vet, N.J.; Neubert, A.; Rascher, W.; Lagler, F.B.; Male, C.; Grytli, H.; Halvorsen, T.; de Hoog, M.; et al. Benefit-Risk Assessment of Off-Label Drug Use in Children: The Bravo Framework. Clin. Pharmacol. Ther. 2021, 110, 952–965. [Google Scholar] [CrossRef]
- van Meer, P.J.; Graham, M.L.; Schuurman, H.-J. The safety, efficacy and regulatory triangle in drug development: Impact for animal models and the use of animals. Eur. J. Pharmacol. 2015, 759, 3–13. [Google Scholar] [CrossRef]
- van der Voet, M.; Teunis, M.; de Haar, J.L.-V.; Stigter, N.; Bhalla, D.; Rooseboom, M.; Wever, K.E.; Krul, C.; Pieters, R.; Wildwater, M.; et al. Towards a reporting guideline for developmental and reproductive toxicology testing in C. elegans and other nematodes. Toxicol. Res. 2021, 10, 1202–1210. [Google Scholar] [CrossRef]
| Study Design | Bacterial Strain Used | Screening Method | Key Findings | Compound Class | In Vitro Activity | In Vivo Activity | Clinical Potential | Ref |
|---|---|---|---|---|---|---|---|---|
| HTS | MRSA, VISA, VRSA, VRE | C. elegans-MRSA infection model in 384-well plates | Identified IMD0354 as a potent bacteriostatic drug against MRSA and VRE | Kinase inhibitor (IMD0354) | Minimum Inhibitory Concentration (MIC): 0.06–0.25 µg/mL against MRSA and VRE | Complete protection of C. elegans at 7.14 µg/mL | Promising lead compound for multidrug-resistant bacteria | [46] |
| C. elegans infection model | B. subtilis GS67 | N/A | B. subtilis GS67 increased C. elegans resistance to Gram-positive pathogens, through the production of fengycin | Lipopeptide (Fengycin) | - | Increased worm resistance to pathogens | Potential for commensal mediated protection | [136] |
| Liquid-based screening assay | S. aureus NCTC8325-4, MSSA, ATCC33591 (MRSA) | Liquid-based C. elegans-S. aureus assay in 24-well plates | Identification of 14 natural compounds and 14 synthetic compounds that prolonged survival of infected worms | Natural extracts and synthetic compounds | >50% survival of infected worms | 8/28 hits had no effect on S. aureus growth | Potential for identifying host-defense modulators | [138] |
| Automated HTS | E. faecalis MMH594 | Automated C. elegans-E. faecalis model in 384-well plates | Identified 28 compounds not without prior antimicrobial properties | Various compounds and extract | Promoted nematode survival | Some compounds had little or no effect on in vitro bacterial growth | Potential for identifying compounds targeting bacterial virulence or host defense | [139] |
| HTS | Automated HTS | E. faecalis MMH594 | Identified 28 compounds without prior antimicrobial properties | Various compounds and extracts | Enhanced survival of infected worms | Six structural classes had a positive effect on infected C. elegans but had no effect on pathogen growth in vitro | Novel antimicrobial compounds with potential host-mediated effects | [134] |
| High-throughput liquid screening | MRSA MW2 BAA-1707, VRSA VRS1 | C. elegans-MRSA liquid assay in 384-well plates | Identified 27 out of 30 clinically used antibiotics and the antihelminthic drug closantel as hits | FDA-approved drugs | Increased C. elegans survival | Identified known antibiotics and novel antimicrobial compound (closantel) | Potential for repurposing existing drugs for antimicrobial use | [135] |
| C. elegans infection screen | MRSA | C. elegans-MRSA infection screen (method not specified) | Identified two synthetic retinoids, CD437 and CD1530, effective against MRSA and persister cells | Synthetic retinoids | Enhanced survival of infected worms | Effective against both growing and persister MRSA cells | Promising new class of antimicrobials for difficult-to-treat infections | [140] |
| HTS | MRSA MW2, E. faecium ATCC E007, K. pneumoniae ATCC 77326, A. baumannii ATCC 17978, P. aeruginosa PA14, E. aerogenes EAE 2625 | C. elegans-MRSA HTS in 384-well plates | Identified four novel antistaphylococcal compounds with MICs from 2 to 8 ffg/mL against MRSA | Novel synthetic compounds | Improved survival of MRSA-infected worms | MICs: 2–8 ffg/mL against MRSA and E. faecium | New antistaphylococcal compounds with potential for further development | [136] |
| C. elegans infection model | MSSA (type-C and type-G strains) | C. elegans-MSSA infection model | Demonstrated correlation between MSSA strain virulence in C. elegans and clinical manifestations | Virulence assessment | N/A | Correlated with clinical manifestations | Potential for using C. elegans to predict clinical virulence of S. aureus strains | [141] |
| C. elegans infection assay | MRSA | C. elegans-MRSA assay | Identified four Brevinin-2 family peptides that improved survival of MRSA-infected worms | Brevinin-2 peptides | Improved survival of MRSA-infected worms | N/A | Potential new applied peptide candidates against MRSA | [135] |
| SN | Compounds | Class of Drugs | Diseases | Action Mechanisms | Effects | Ref. |
|---|---|---|---|---|---|---|
| 1 | Cannabidiol (CBD) | nonpsychoactive phytocannabinoid | AD | It employs an antioxidative mechanism reliant on the classical transcription factors DAF-16 and SKN-1. | Reduces ROS in vivo, but does not overexpress genes related to anti-oxidation. | [169] |
| 2 | Cannabidiol (CBD) and dihydroartemisinin (DHA) | Drug–drug conjugate | AD | Conjugates suppressed NF-B activation but not MAPK activation in BV-2 cells, hence preventing LPS-induced neuroinflammation. | They exhibit cytotoxicity, significantly limiting their therapeutic windows. The CBD-DHA conjugate C3D reduced neurotoxicity while maintaining anti-neuroinflammation action. | [124] |
| 3 | Dihydroartemisinin (DHA) | Artemisinin | AD | Artemisinin alleviates neuroinflammatory responses in the central nervous system (CNS). | [124] | |
| 4 | Sorafenib | A Raf Kinase inhibitor | PD | Inhibition of leucine-rich repeat kinase 2 (LRRK2). | Increased survival and reduced dopaminergic neuron degeneration in G2019S-LRRK2 transgenic C. elegans. | [171] |
| 5 | GW5074 | an indoline compound | PD | Strongly suppresses the autophosphorylation of LRRK2 and G2019S LRRK2. | Enhanced survival and decreased degeneration of dopaminergic neurons in G2019S-LRRK2 transgenic C. elegans. | [171] |
| 6 | Rifampicin | Glycation inhibitor | RIF plays a double role by reducing glycation in vivo while simultaneously activating pro-longevity processes through DAF-16. | Extends the lifespan of C. elegans by up to 60%. | [172] | |
| 7 | Fluoxetine | Selective serotonin reuptake inhibitor | Antidepressant, AD | Possesses antioxidative properties while targeting the Nrf2 signaling pathway. It also exhibits anti-inflammatory/antioxidant actions via inhibiting TLR4/NLRP3 inflammasome signaling pathways. | Resulted in improved antioxidant defense mechanism, decreases in AChE activity, amyloid, Tau protein, MDA, TNF-, and IL1 contents. It also showed cardioprotective benefits by improving lipid profile and significantly lowering cardiac enzymes such as CK-MB, troponin, and MEF2. | [171] |
| 8 | Galantamine, haemanthidine | A benzylphenethylamine alkaloid | Moderate dementia (AD) | It is a cholinesterase inhibitor with a dual-action mechanism. As a reversible acetylcholinesterase inhibitor, it enhances acetylcholine’s action on nicotinic receptors, improving cholinergic neurotransmission in the CNS. | Delayed paralysis and improved lifespan of transgenic worms. | [173] |
| 9 | DMSO | A universal solvent | Aβ toxicity | Both the presynaptic acetylcholinesterase inhibitor (AChEi) aldicarb and the postsynaptic nicotinic acetylcholine receptor (iAChR) agonist levamisole induced paralysis in CL4176 worms, with approximately 70% paralysis observed. DMSO (2%) delayed acetylcholinesterase neurotransmission and appeared to alter Ca2+ ion permeability, which is essential for neurotransmission. Interestingly, both DMSO and ethylenediaminetetraacetic acid (EDTA) provided similar resistance to aldicarb, whether used separately or together. | Delayed paralysis by 48% and 90%, and provided protective effects against Aβ toxicity. | [174] |
| 10 | Swertiamarin | A secoiridoidial glycoside | Cholinergic dysfunction | It offers neuroprotection by modulating ROS scavenging genes and enhancing cholinergic neurotransmission through the regulation of synaptic AChE. | Amelioration of cholinergic dysfunction. | [175] |
| 11 | Glycyrrhizic acid (GA), glycyrrhetinic acid (GRA), liquiritigenin (LG), and isoliquiritigenin (ILG) | Compounds from traditional Chinese medicine (the most active plant for the subject of study) G. uralensis | AD biomarkers, Aβ aggregates and oxidative damage | These substances may activate the transcription factor DAF-16. | Reduced Aβ accumulation. It also had positive effects in counteracting protein aggregation and oxidative stress. | [176] |
| S/N | Compound | Drug Class | Action Mechanism | Effect | Ref. |
|---|---|---|---|---|---|
| 1 | Schisandrin C | A dibenzocyclooctadiene lignan | Schisandrin C improved intestinal permeability dysfunction and suppressed NF-κB and p38 MAPK signaling in Caco-2 cells, which regulate MLCK expression and the structural remodeling of the Tight Junction complex. In both intestinal organoids and C. elegans, Schisandrin C significantly improved aberrant FITC–dextran permeabilization. | Schisandrin C reduces abnormal intestinal permeability and regulates the production of tight junction (TJ) proteins, along with MLCK, p-MLC, and inflammation-related proteins, all of which are associated with leaky gut symptoms and the development of IBD. As a result, it shows potential as a treatment for leaky gut syndrome. | [188] |
| 2 | Lipoteichoic acid | Extract from the probiotic organism Lactobacillus paracasei strain D3-5 | Regulates the TLR-2/p38-MAPK/NF-kB pathway. | Displays strong activity to stimulate mucin production and lower aging-related leaky gut and inflammation | [190] |
| 3 | Lipoteichoic acid | Lipoteichoic acid from a heat-killed Lactobacillus strain | - | The lipoteichoic acid (LTA) element stimulates mucin production through the TLR-2/p38-MAPK/NF-kB pathway, strengthening the reduction of age-related intestinal permeability and inflammation. | [166] |
| 4 | Biacalin | Flavone glycoside | Baicalin (10, 100 mol/L) increased survival and restored behavioral abnormalities in LPS-stimulated C. elegans. It also reduced oxidative stress caused by LPS damage by lowering MDA levels and enhancing SOD activity. | [188] | |
| 5 | Probiotic cocktail containing five Lactobacillus and five Enterococcus strains | Probiotics derived from human-origin sources | Probiotics modified the microbiota in a way that enhanced bile salt hydrolase activity, increasing taurine levels in the gut, which promoted tight junction formation and reduced intestinal permeability. | Probiotic-modulated gut microbiota primarily alleviated leaky gut by enhancing tight junctions, which subsequently reduced inflammation. Taurine restored intestinal integrity and improved health markers in C. elegans. | [190] |
| 6 | E. coli Nissle 1917 | A probiotic | Enteropathogenic E. coli (EPEC) infection reduced the lifespan of C. elegans, but pretreatment with the probiotic E. coli Nissle 1917 (EcN) significantly enhanced the survival of worms infected with EPEC. | Increased the expression of tight junction proteins such as ZO-1 in C. elegans, suggesting that EcN enhances gut barrier integrity. | [191] |
| 7 | 3,3′-Diindolylmethane (DIM) | A digestive byproduct | DIM significantly improved the intestinal permeability of differentiated Caco-2 cells by increasing the expression of tight junction proteins, such as occludin and ZO-1. | It shows potential as a nutraceutical candidate for treating leaky gut syndrome by enhancing intestinal permeability. | [192] |
| 8 | Neuronal peptide | Neuropeptigenic regulators | Neuron-specific knockdown of CAPS and PC2 decreased intestinal leakiness occurring due to age, and extended lifespan in C. elegans and D. melanogaster. | Neuronal knockdowns of insulin-like peptides daf-28, ins-7, dilp2, dilp3, and dilp5 antagonized intestinal barrier integrity and lifespan. Furthermore, it reduces insulin signaling in the intestine through the knockdown of the insulin receptor. | [193] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deji-Oloruntoba, O.O.; Elufioye, T.O.; Adefegha, S.A.; Jang, M. Can Caenorhabditis elegans Serve as a Reliable Model for Drug and Nutraceutical Discovery? Appl. Biosci. 2025, 4, 23. https://doi.org/10.3390/applbiosci4020023
Deji-Oloruntoba OO, Elufioye TO, Adefegha SA, Jang M. Can Caenorhabditis elegans Serve as a Reliable Model for Drug and Nutraceutical Discovery? Applied Biosciences. 2025; 4(2):23. https://doi.org/10.3390/applbiosci4020023
Chicago/Turabian StyleDeji-Oloruntoba, Opeyemi. O., Taiwo. O. Elufioye, Stephen Adeniyi Adefegha, and Miran Jang. 2025. "Can Caenorhabditis elegans Serve as a Reliable Model for Drug and Nutraceutical Discovery?" Applied Biosciences 4, no. 2: 23. https://doi.org/10.3390/applbiosci4020023
APA StyleDeji-Oloruntoba, O. O., Elufioye, T. O., Adefegha, S. A., & Jang, M. (2025). Can Caenorhabditis elegans Serve as a Reliable Model for Drug and Nutraceutical Discovery? Applied Biosciences, 4(2), 23. https://doi.org/10.3390/applbiosci4020023