


Phenolic Compound Production Increased In Vitro Regenerated Cichorium intybus L.


Abstract
1. Introduction
2. Material and Methods
2.1. Plant Sample
2.2. Plant Tissue Culture Studies
2.3. Extraction and Analysis of Phenolic Composition and Polysaccharide (Inulin) of Cichorium intybus L.
- Roots in vivo
- Leaves in vivo
- Stems in vivo
- Flowers in vivo
- Leaves in vitro
- Callus in vitro
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of Variance |
| BAP | Benzyl Amino Purine |
| HPLC | High-Pressure Liquid Chromatography |
| IAA | Indole 3 -Acetic Acid |
| IBA | Indole Butyric Acid |
| LC | Liquid Chromatography |
| LS | Linsmaier & Skoog |
| MS media | Murashige-Skoog media |
| NAA | Naphthalene Acetic acid |
| WO227 | White medium |
| G0209 | Gamborg B5 medium |
| PGR | Plant Growth Regulators |
| UV | Ultraviolet |
References
- Wang, Q.; Cui, J. Perspectives and utilization technologies of chicory (Cichorium intybus L.): A review. Afr. J. Biotechnol. 2011, 10, 1966–1977. [Google Scholar] [CrossRef]
- Liang, X.Y.; Zhang, X.Q.; Bai, S.Q.; Huang, L.K.; Luo, X.M.; Ji, Y.; Jiang, L.F. Genetic diversity and relationship of chicory (Cichorium intybus L.) using sequence-related amplified polymorphism markers. Genet. Mol. Res. 2014, 13, 7736–7746. [Google Scholar] [CrossRef] [PubMed]
- Puhlmann, M.L.; de Vos, W.M. Back to the roots: Revisiting the use of the fiber-rich Cichorium intybus L. Taproots. Adv. Nutr. 2020, 11, 878–889. [Google Scholar] [CrossRef] [PubMed]
- Abas, Y.A. Regeneration of Cichorium intybus L. in Tissue Vulture and Secondary Metabolite Analysis. Master’s Thesis, Van Yuzuncu Yil University, Van, Turkey, May 2018. [Google Scholar]
- Al-Snafi, A.E. Medical importance of Cichorium intybus—A review. IOSR J. Pharmasy 2016, 6, 41–56. [Google Scholar]
- Nandagopal, S.; Kumari, B.D.R. Adenine sulphate induced high frequency shoot organogenesis in callus and in vitro flowering of Cichorium intybus L. cv. Focus-a potent medicinal plant. Acta Agric. Slov. 2006, 87, 415–425. [Google Scholar]
- Kumari, B.D.R.; Ranjitha Kumari, B.D.; Nandagopal, S. Phytochemical and Antibacterial Studies of Chicory (Cichorium intybus L.)—A Multipurpose Medicinal Plant. Adv. Biol. Res. 2007, 1, 17–21. [Google Scholar]
- Maroufi, A.; van Bockstaele, E.; de Loose, M. Differential expression of fructan 1-exohydrolase genes involved in inulin biodegradation in chicory (Cichorium intybus) cultivars. Aust. J. Crop Sci. 2012, 6, 1362–1368. [Google Scholar]
- Doliński, R.; Olek, A. Micropropagation of wild chicory (Cichorium intybus L. var. silvestre Bisch.) from leaf explants. Acta Sci. Pol. Cultus 2013, 12, 33–44. [Google Scholar]
- Zahid, A. Transformation and Agrotechnology Strategies for Improved Levels of Secondary Metabolite in Cichorium intybus L.; Jamia Hamdard University: Jamia Hambdar, India, 2012. [Google Scholar]
- Meena, A.; Renu, S. Phytochemical screening of sterols extracted from Cichorium intybus in vivo and in vitro. World J. Pharm. Pharm. Sci. 2014, 3, 1349–1360. [Google Scholar]
- Starmans, D.A.J.; Nijhuis, H.H. Extraction of secondary metabolites from plant material: A review. Trends Food Sci. Technol. 1996, 7, 191–197. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Ibrahim, M.; El-Bahr, M.; Rady, M. In-vitro adventitious root production of Cichorium endivia L. and antioxidants, total phenolic, and total flavonoids assessments. Egypt. Pharm. J. 2019, 18, 216. [Google Scholar] [CrossRef]
- Dreschke, G.; Papirio, S.; Sisinni, D.M.G.; Lens, P.N.L.; Esposito, G. Effect of feed glucose and acetic acid on continuous biohydrogen production by Thermotoga neapolitana. Bioresour. Technol. 2019, 273, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, R.; Irvin, L.M.; Vaidya, B.N.; Dhekney, S.A.; Joshee, N. Optimizing plant regeneration and genetic transformation of Paulownia elongata. Biocatal. Agric. Biotechnol. 2021, 33, 101970. [Google Scholar] [CrossRef]
- Barreiro, K.; Dwivedi, O.P.; Valkonen, S.; Groop, P.H.; Tuomi, T.; Holthofer, H.; Rannikko, A.; Yliperttula, M.; Siljander, P.; Laitinen, S.; et al. Urinary extracellular vesicles: Assessment of pre-analytical variables and development of a quality control with focus on transcriptomic biomarker research. J. Extracell. Vesicles 2021, 10, e12158. [Google Scholar] [CrossRef]
- Eray, N.; Dalar, A.; Turker, M. The effects of abiotic stressors and signal molecules on phenolic composition and antioxidant activities of in vitro regenerated Hypericum perforatum (St. John’s Wort). South African J. Bot. 2020, 133, 253–263. [Google Scholar] [CrossRef]
- Gharari, Z.; Hanachi, P.; Sadeghinia, H.; Walker, T.R. Cichorium intybus bio-callus synthesized silver nanoparticles: A promising antioxidant, antibacterial and anticancer compound. Int. J. Pharm. 2022, 625, 122062. [Google Scholar] [CrossRef]
- Othman, M.; Helmi, L.M.; Hosni, A.M. Effect of growth regulator NAA and IBA applications on total phenolic and flavonoid compounds extracted from in vitro produced callus of chicory plant (Cichorium intybus L.). Arab. Univ. J. Agric. Sci. 2019, 27, 1929–1936. [Google Scholar] [CrossRef]
- Farooq, I.; Qadri, Z.A.; Rather, Z.A.; Nazki, I.T.; Banday, N.; Rafiq, S.; Masoodi, K.Z.; Noureldeen, A.; Mansoor, S. Optimization of an improved, efficient and rapid in vitro micropropagation protocol for Petunia hybrida Vilm. Cv. “Bravo.” Saudi J. Biol. Sci. 2021, 28, 3701–3709. [Google Scholar] [CrossRef]
- Lee, N.W.; Wang, H.Y.; Du, C.L.; Yuan, T.H.; Chen, C.Y.; Yu, C.J.; Chan, C.C. Air-polluted environmental heavy metal exposure increase lung cancer incidence and mortality: A population-based longitudinal cohort study. Sci. Total Environ. 2022, 810, 152186. [Google Scholar] [CrossRef]
- Chavda, J.; Dwsai, B.; Hha, S.; Tandel, M.; Patel, D. Effect of PGR on clonal propagation of tion of madhunashini (Gymnema Sylvestre R. BR) through rooted cutting. An. Int. Q. J. Life Sci. 2015, 10, 1645–1648. [Google Scholar]
- Gâteblé, G.; Pastor, M. Ontogenic stage, auxin type and concentration influence rooting of Oxera sulfurea stem cuttings. In Proceedings of the I International Symposium on the Labiatae: Advances in Production, Biotechnology and Utilisation 723, Sanremo, Italy, 22–25 February 2006; pp. 269–272. [Google Scholar]
- Dalar, A.; Konczak, I. Cichorium intybus from Eastern Anatolia: Phenolic composition, antioxidant and enzyme inhibitory activities. Ind. Crops Prod. 2014, 60, 79–85. [Google Scholar] [CrossRef]
- Innocenti, M.; Gallori, S.; Giaccherini, C.; Ieri, F.; Vincieri, F.F.; Mulinacci, N. Evaluation of the Phenolic Content in the Aerial Parts of Different Varieties of Cichorium intybus L. J. Agric. Food Chem. 2005, 53, 6497–6502. [Google Scholar] [CrossRef] [PubMed]
- Vanzo, A.; Cecotti, R.; Vrhovsek, U.; Torres, A.M.; Mattivi, F.; Passamonti, S. The Fate of trans-Caftaric Acid Administered into the Rat Stomach. J. Agric. Food Chem. 2007, 55, 1604–1611. [Google Scholar] [CrossRef] [PubMed]
- Weisshaar, B.; Jenkins, G.I. Phenylpropanoid biosynthesis and its regulation. Curr. Opin. Plant Biol. 1998, 1, 251–257. [Google Scholar] [CrossRef]
- Dixon’, R.A.; Paiva, N.L. Stress-lnduced Phenylpropanoid Metabolism. Plant Cell. 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Lee, J.; Scagel, C.F. Chicoric acid: Chemistry, distribution, and production. Front. Chem. 2013, 1. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Type of Media | Type of PGRs and Concentration | Type of Explant | Type of Regeneration |
|---|---|---|---|
| B5 | BAP 0.14 mg L−1 IAA 0.16 mg L−1 | Leaf | Callus + shoot |
| MS | NAA 0.17 mg L−1 IBA 0.17 mg L−1 | Leaf | Callus + shoot |
| MS | IAA 0.16 mg L−1 BAP 0.14 mg L−1 | Leaf | Callus |
| MS | IBA 0.16 mg L−1 NAA 0.02 mg L−1 GA3 0.06 mg L−1 | Leaf | Callus + shoot |
| MS | IBA 0.16 mg L−1 BAP 0.024 mg L−1 | Leaf | Callus + root |
| B5 | IBA 0.055 mgL−1 IAA 10.17 mg L−1 | Leaf | Callus + shoot |
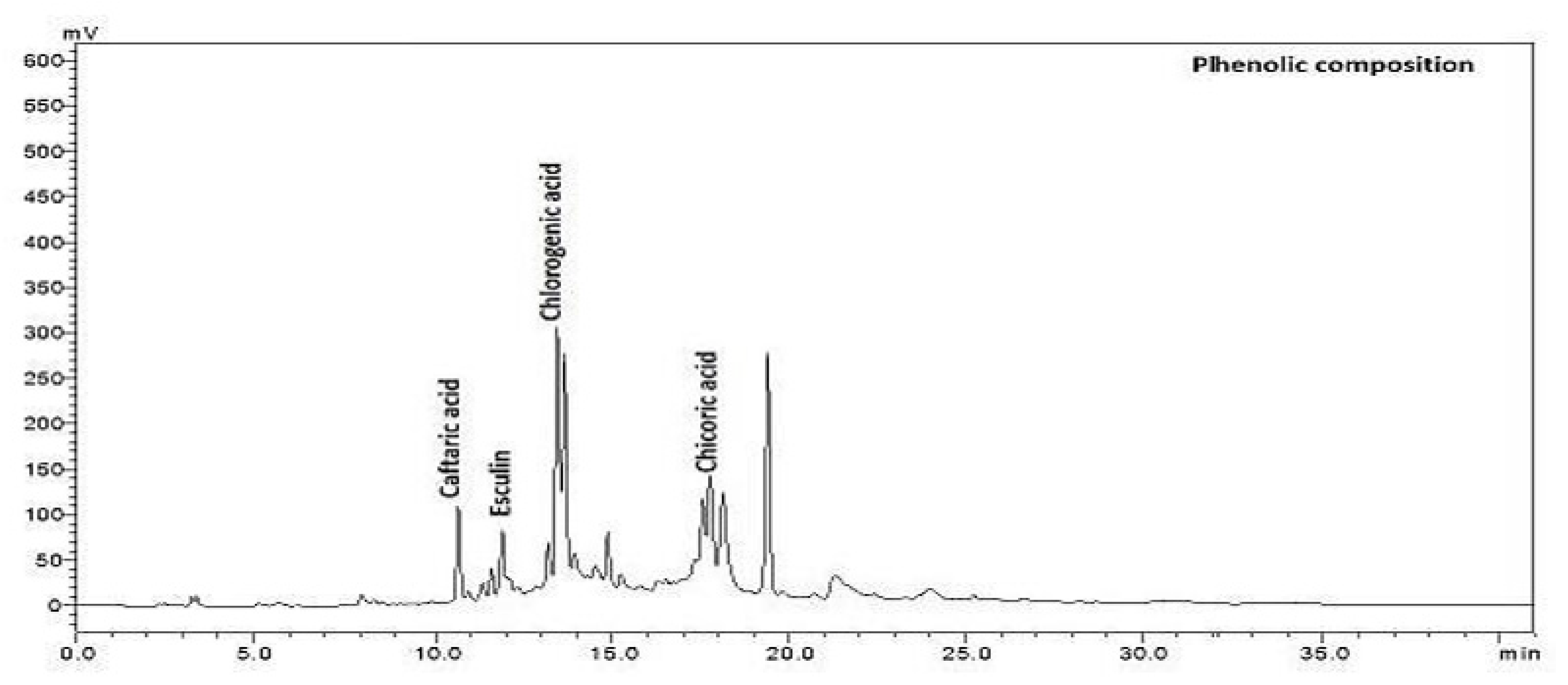
| Sample ID | Concentration of Phenolic Compounds (mg/g) | |||
|---|---|---|---|---|
| Chicoric Acid | Caftaric Acid | Esculin | Chlorogenic Acid | |
| Root in vivo | 0.2 ± 0.01 f | 0.2 ± 0.001 c | 12.1 ± 0.1 c | 6.3 ± 0.1 d |
| Stems in vivo | 5.4 ± 0.15 b | 0.4 ± 0.005 b | 4.6 ± 0.1 d | 3.9 ± 0.08 f |
| Leaf in vivo | 2.0 ± 0.01 c | 0.5 ± 0.005 b | 24.2 ± 0.1 b | 10.4 ± 0.2 c |
| Flower in vivo | 17.3 ± 1.2 a | 0.2 ± 0.002 c | 65.2 ± 0.1 a | 12.2 ± 0.1 b |
| Leaf in vitro | 1.1 ± 0.005 d | 6.1 ± 0.4 a | 2.3 ± 0.002 e | 15.4 ± 0.1 a |
| Callus in vitro | 0.3 ± 0.003 e | 0.4 ± 0.004 b | 0.7 ± 0.01 f | 5.6 ± 0.1 e |
| Sample | The Concentration of Inulin (mg/g Dry Weight) |
|---|---|
| Root in vivo | 37.9 ± 0.5 a |
| Stems in vivo | 16.1 ± 1.2 d |
| Leaves in vivo | 33.4 ± 0.5 b |
| Flower in vivo | 17.9 ± 1.6 d |
| Leaves in vitro | 28.8 ± 0.5 c |
| Callus in vitro | 14.3 ± 0.5 e |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abas, Y.A.; Eroğlu, A.; Dalar, A.; Türker, M.; Ozdemir, F.A.; Sołowski, G. Phenolic Compound Production Increased In Vitro Regenerated Cichorium intybus L. Appl. Biosci. 2023, 2, 84-93. https://doi.org/10.3390/applbiosci2010008
Abas YA, Eroğlu A, Dalar A, Türker M, Ozdemir FA, Sołowski G. Phenolic Compound Production Increased In Vitro Regenerated Cichorium intybus L. Applied Biosciences. 2023; 2(1):84-93. https://doi.org/10.3390/applbiosci2010008
Chicago/Turabian StyleAbas, Yousif Abdullah, Ayten Eroğlu, Abdullah Dalar, Musa Türker, Fethi Ahmet Ozdemir, and Gaweł Sołowski. 2023. "Phenolic Compound Production Increased In Vitro Regenerated Cichorium intybus L." Applied Biosciences 2, no. 1: 84-93. https://doi.org/10.3390/applbiosci2010008
APA StyleAbas, Y. A., Eroğlu, A., Dalar, A., Türker, M., Ozdemir, F. A., & Sołowski, G. (2023). Phenolic Compound Production Increased In Vitro Regenerated Cichorium intybus L. Applied Biosciences, 2(1), 84-93. https://doi.org/10.3390/applbiosci2010008