

MSM Supplementation Is Associated with Reduced Inflammation and Improved Innate Immune Response following In Vitro LPS-Stimulation in Humans after a Bout of Downhill Running


Abstract
:1. Introduction
2. Results
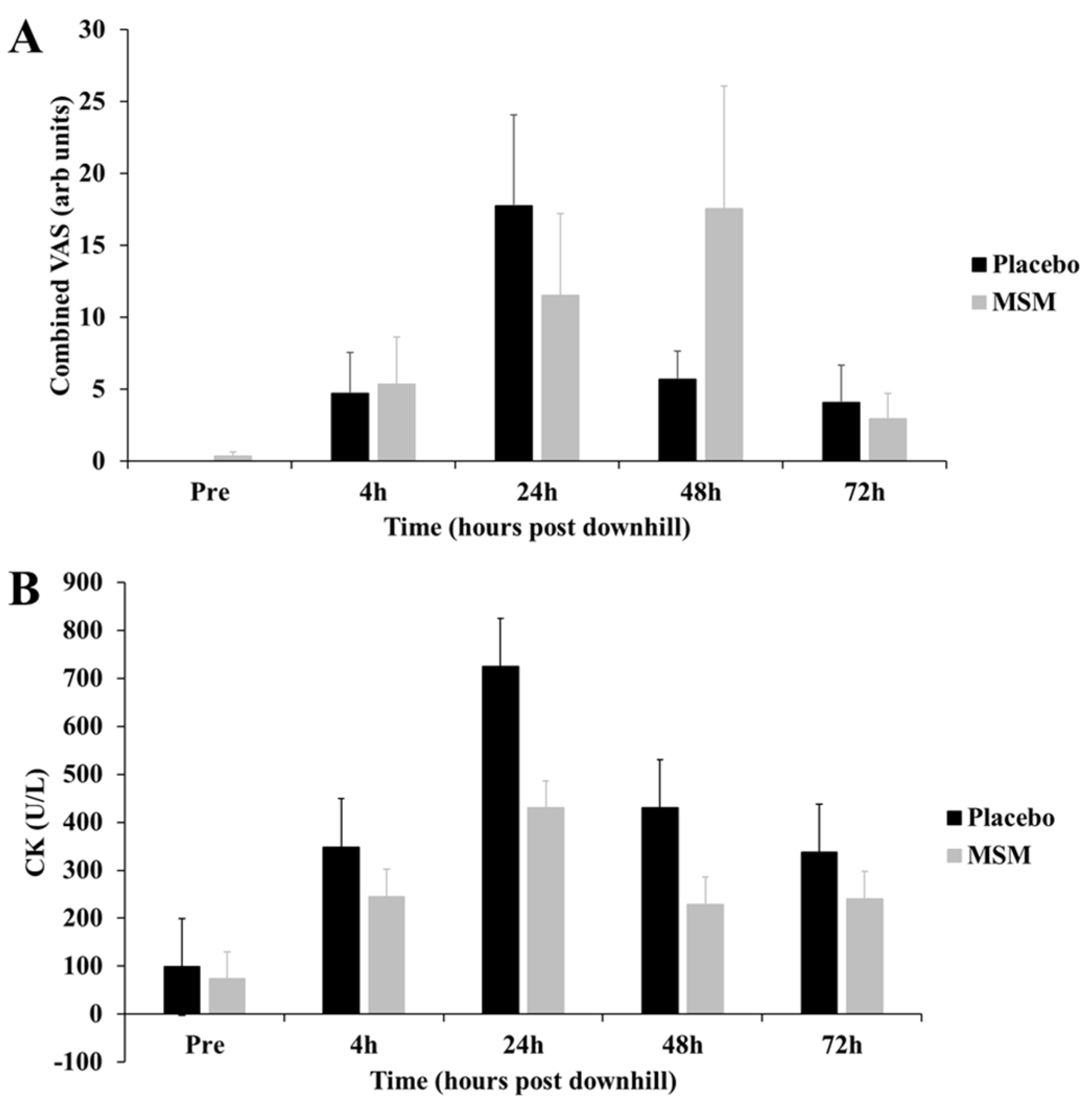
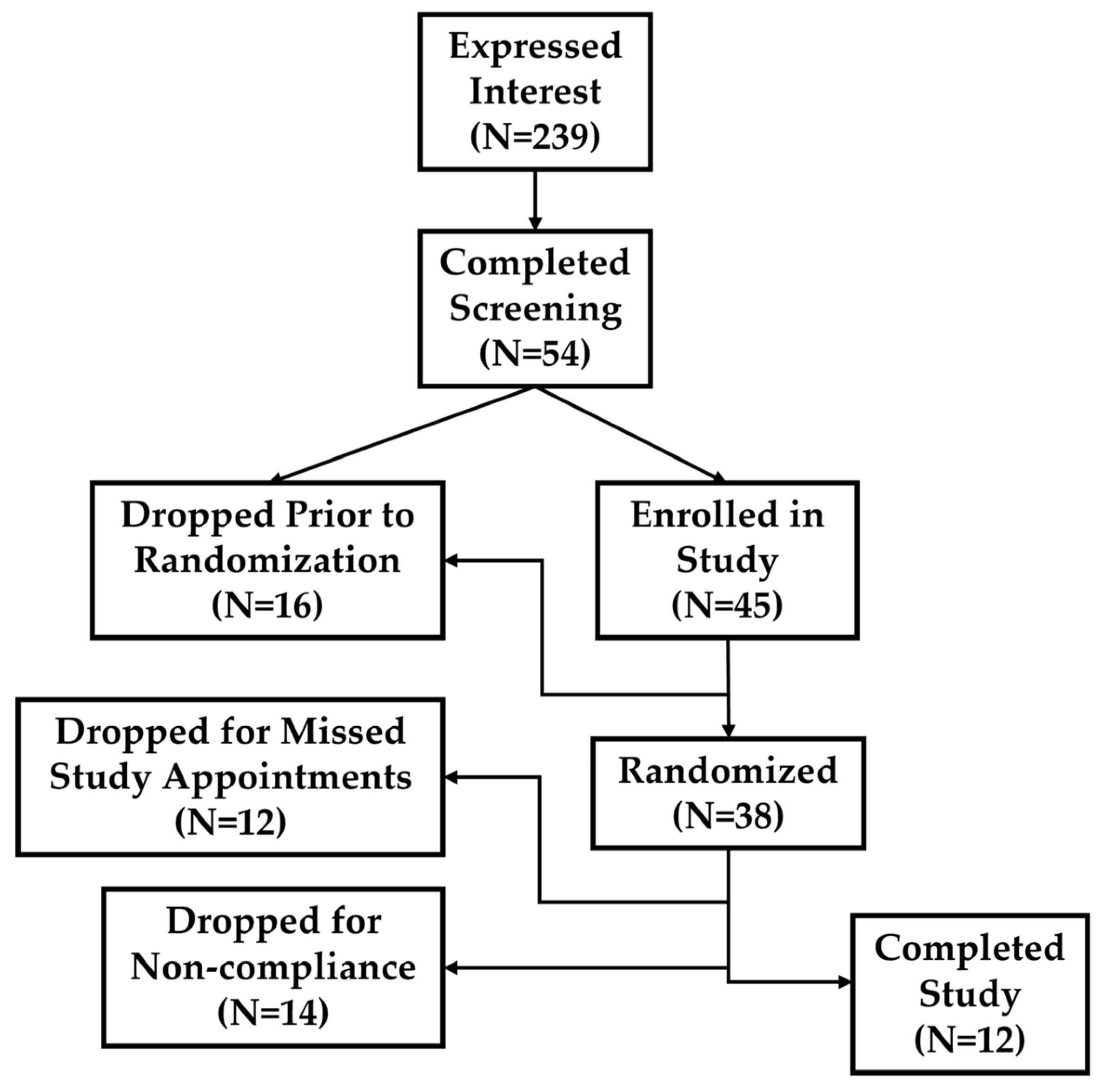
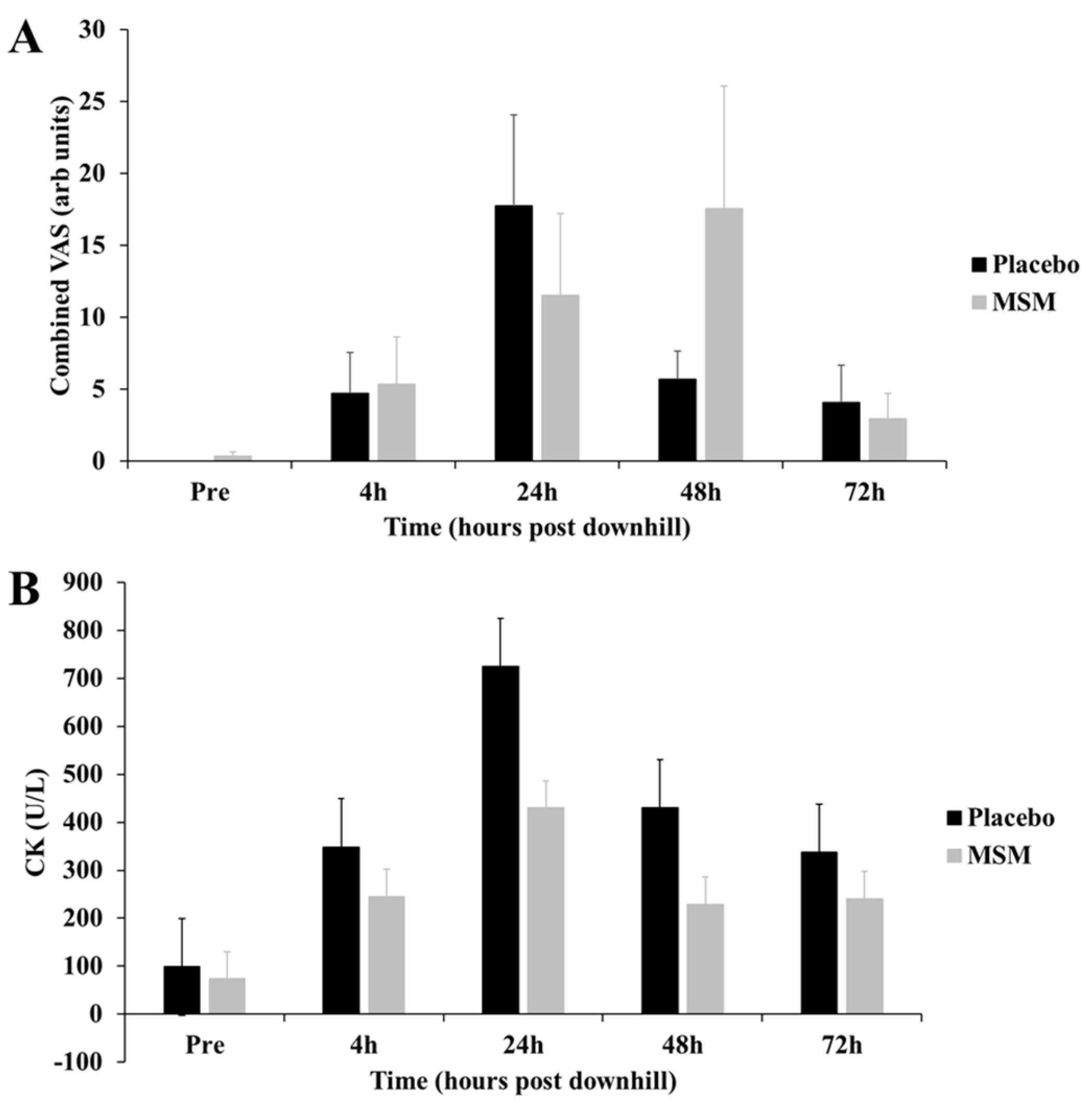
2.1. Exercise-Induced Muscle Damage Response
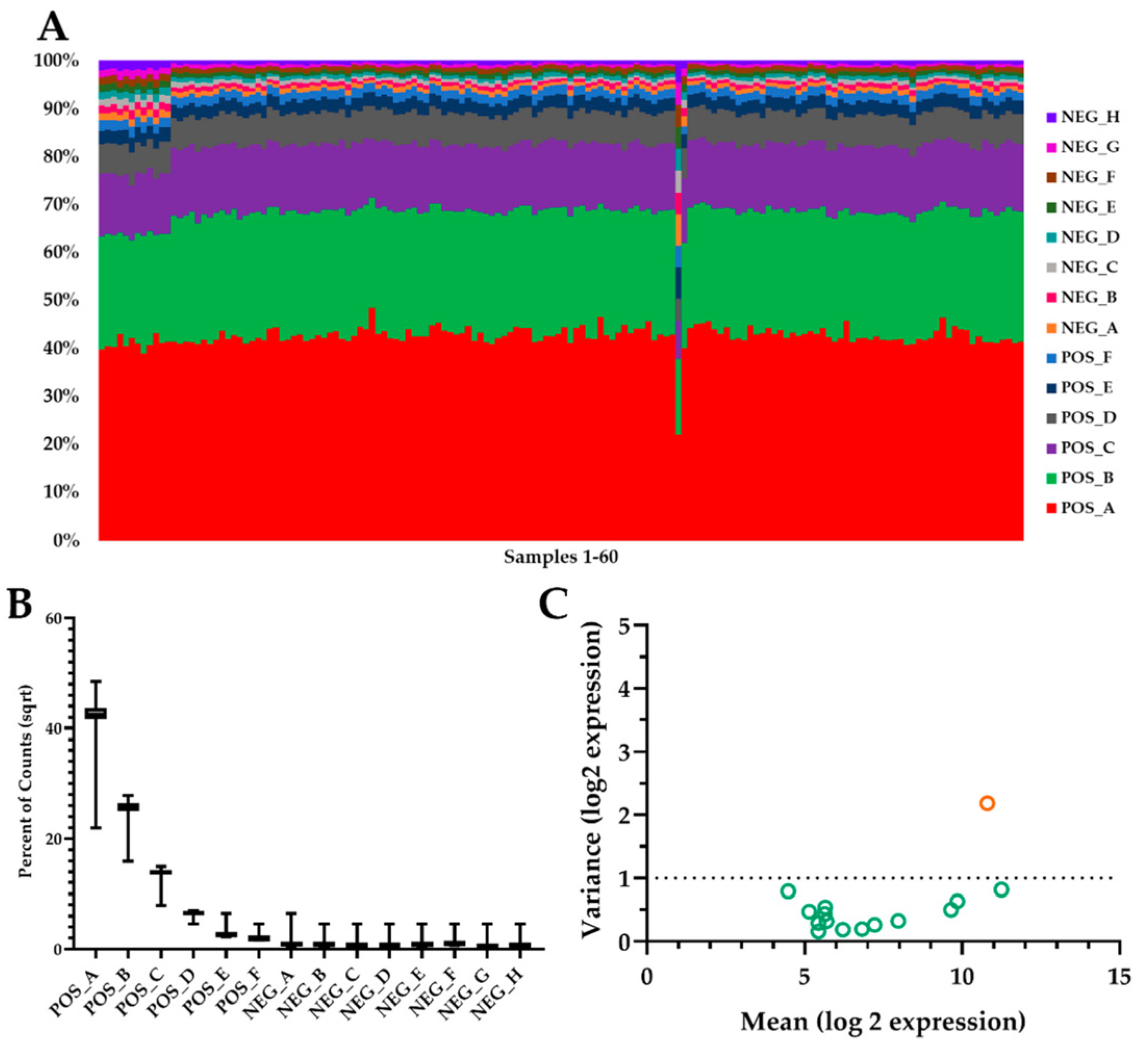
2.2. Global Summary mRNA Response to EIMD
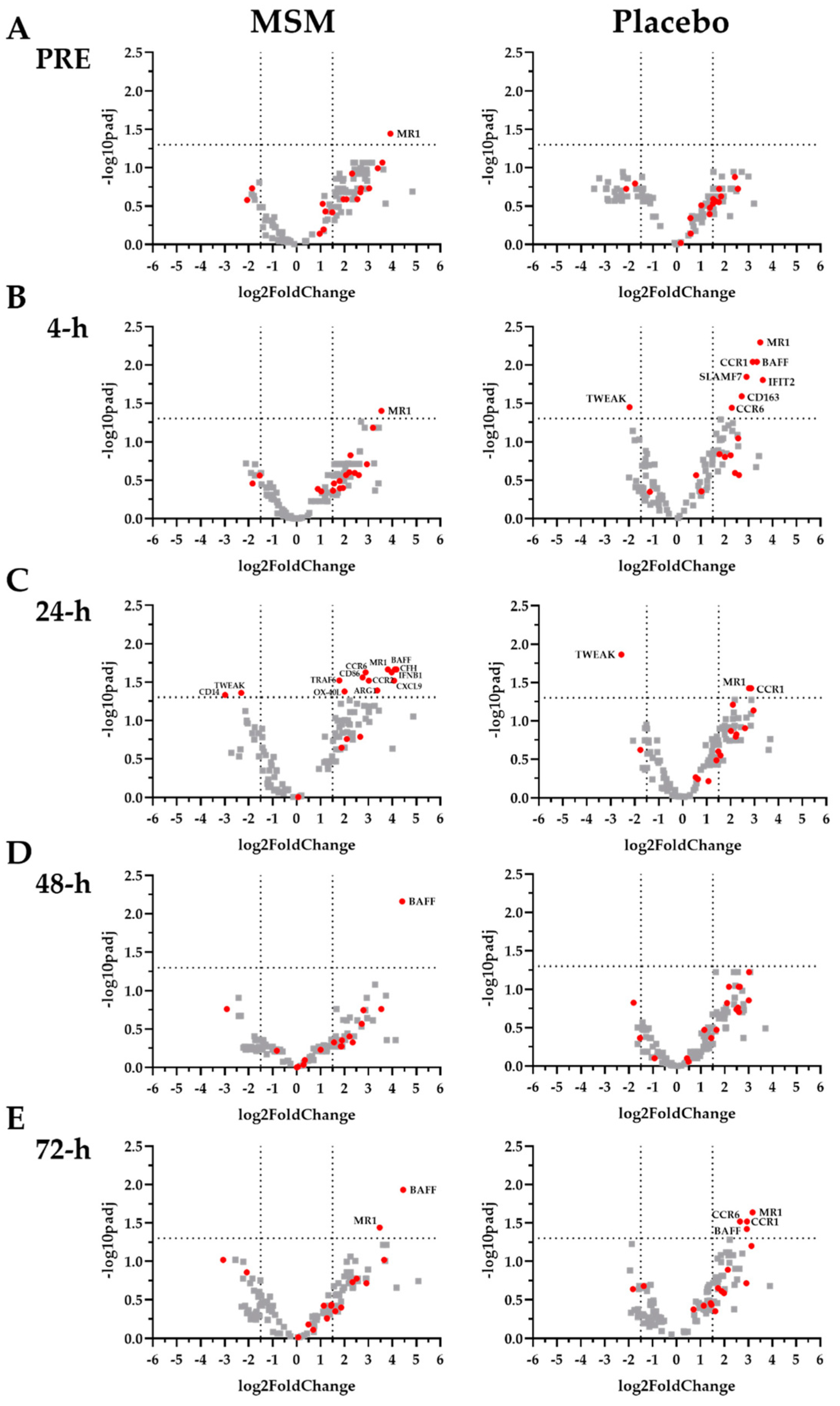
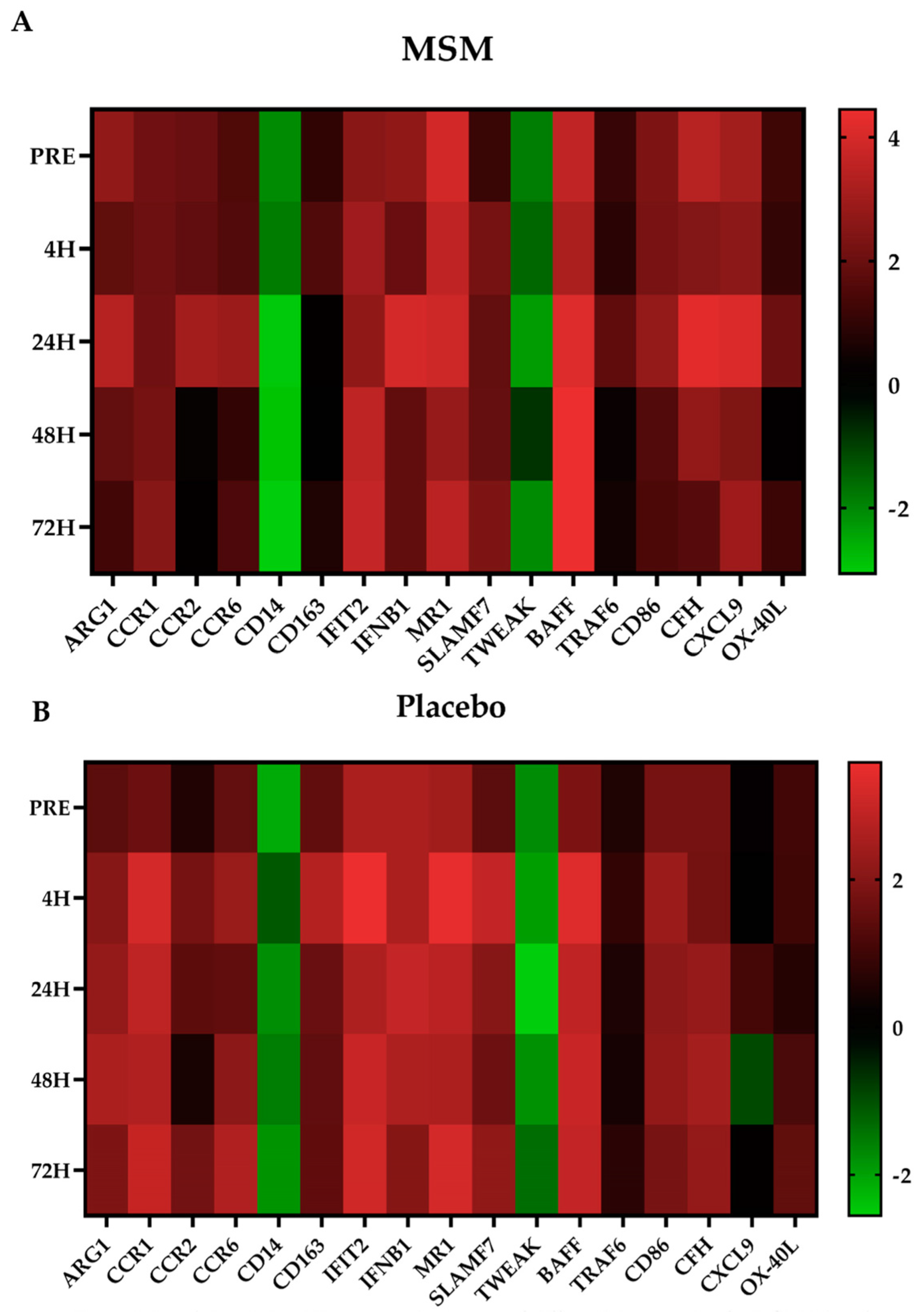
2.3. Inflammation-Associated mRNA
2.4. Viral Immune Response mRNA
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Supplement Conditions
4.3. Statistical Sample Size Analysis
4.4. Blood Sample Collection
4.5. Downhill Running Trial
4.6. Subjective Muscle Soreness
4.7. LPS-Stimulation
4.8. RNA Isolation from Cell Pellets
4.9. Nanostring Analysis
4.10. Statistical Analysis
4.11. Data Interpretation and Presentation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Full Name | Accession ID |
|---|---|---|
| ARG1 | Arginase | NM_000045.2 |
| BAFF | Tumor necrosis factor (ligand) superfamily, member 13b | NM_006573.4 |
| CCR1 | Chemokine (C-C motif) receptor 1 | NM_001295.2 |
| CCR2 | Chemokine (C-C motif) receptor 2 | NM_001123041.2 |
| CCR6 | Chemokine (C-C motif) receptor 6 | NM_031409.2 |
| CD14 | CD14 molecule | NM_000591.2 |
| CD86 | CD86 molecule | NM_175862.3 |
| CD163 | CD163 molecule | NM_004244.4 |
| CFH | Complement factor H | NM_001014975.2 |
| CXCL9 | Chemokine (C-X-C motif) ligand 9 | NM_002416.1 |
| IFIT2 | Interferon-induced protein with tetratricopeptide repeats 2 | NM_001547.4 |
| IFNB1 | Interferon, beta 1, fibroblast | NM_002176.2 |
| MR1 | Major histocompatibility complex, class I-related | NM_001531.2 |
| OX-40L | Tumor necrosis factor (ligand) superfamily, member 4 | NM_003326.2 |
| SLAMF7 | SLAM family member 7 | NM_021181.3 |
| TRAF6 | Tumor necrosis factor receptor-associated factor 6 | NM_145803.1 |
| TWEAK | Tumor necrosis factor (ligand) superfamily, member 12 | NM_003809.2 |
References
- Proske, U.; Morgan, D.L. Muscle damage from eccentric exercise: Mechanism, mechanical signs, adaptation and clinical applications. J. Physiol. 2001, 537, 333–345. [Google Scholar] [CrossRef]
- Bazzucchi, I.; Patrizio, F.; Ceci, R.; Duranti, G.; Sgro, P.; Sabatini, S.; Di Luigi, L.; Sacchetti, M.; Felici, F. The Effects of Quercetin Supplementation on Eccentric Exercise-Induced Muscle Damage. Nutrients 2019, 11, 205. [Google Scholar] [CrossRef] [PubMed]
- Buford, T.W.; Cooke, M.B.; Shelmadine, B.D.; Hudson, G.M.; Redd, L.; Willoughby, D.S. Effects of eccentric treadmill exercise on inflammatory gene expression in human skeletal muscle. Appl. Physiol. Nutr. Metab. 2009, 34, 745–753. [Google Scholar] [CrossRef]
- Park, K.S.; Lee, M.G. Effects of unaccustomed downhill running on muscle damage, oxidative stress, and leukocyte apoptosis. J. Exerc. Nutr. Biochem. 2015, 19, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Rajaee, A.; Barnett, R.; Cheadle, W.G. Pathogen- and Danger-Associated Molecular Patterns and the Cytokine Response in Sepsis. Surg. Infect. 2018, 19, 107–116. [Google Scholar] [CrossRef]
- Imai, Y.; Kuba, K.; Neely, G.G.; Yaghubian-Malhami, R.; Perkmann, T.; van Loo, G.; Ermolaeva, M.; Veldhuizen, R.; Leung, Y.H.; Wang, H.; et al. Identification of oxidative stress and Toll-like receptor 4 signaling as a key pathway of acute lung injury. Cell 2008, 133, 235–249. [Google Scholar] [CrossRef]
- Lucas, K.; Maes, M. Role of the Toll Like receptor (TLR) radical cycle in chronic inflammation: Possible treatments targeting the TLR4 pathway. Mol. Neurobiol. 2013, 48, 190–204. [Google Scholar] [CrossRef]
- Lefebvre, J.S.; Levesque, T.; Picard, S.; Pare, G.; Gravel, A.; Flamand, L.; Borgeat, P. Extra domain A of fibronectin primes leukotriene biosynthesis and stimulates neutrophil migration through activation of Toll-like receptor 4. Arthritis Rheum. 2011, 63, 1527–1533. [Google Scholar] [CrossRef] [PubMed]
- Rani, M.; Sayyadioskoie, S.R.; Galvan, E.M.; Nicholson, S.E.; Schwacha, M.G. Trauma-induced lung injury is associated with infiltration of activated TLR expressing myeloid cells. Cytokine 2021, 141, 155457. [Google Scholar] [CrossRef]
- McFarlin, B.K.; Johnston, C.A.; Tyler, C.; Hutchison, A.T.; Kueht, M.L.; Reeves, R.; Foreyt, J.P. Inflammatory markers are elevated in overweight Mexican-American children. Int. J. Pediatr. Obes. 2007, 2, 235–241. [Google Scholar] [CrossRef]
- Xie, Q.; Shen, W.W.; Zhong, J.; Huang, C.; Zhang, L.; Li, J. Lipopolysaccharide/adenosine triphosphate induces IL-1beta and IL-18 secretion through the NLRP3 inflammasome in RAW264.7 murine macrophage cells. Int. J. Mol. Med. 2014, 34, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Relja, B.; Horstmann, J.P.; Kontradowitz, K.; Jurida, K.; Schaible, A.; Neunaber, C.; Oppermann, E.; Marzi, I. Nlrp1 inflammasome is downregulated in trauma patients. J. Mol. Med. 2015, 93, 1391–1400. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.M.; Lowery, R.P.; Joy, J.M.; Walters, J.A.; Baier, S.M.; Fuller, J.C., Jr.; Stout, J.R.; Norton, L.E.; Sikorski, E.M.; Wilson, S.M.; et al. Beta-Hydroxy-beta-methylbutyrate free acid reduces markers of exercise-induced muscle damage and improves recovery in resistance-trained men. Br. J. Nutr. 2013, 110, 538–544. [Google Scholar] [CrossRef]
- McFarlin, B.K.; Hill, D.W.; Vingren, J.L.; Curtis, J.H.; Tanner, E.A. Dietary Polyphenol and Methylsulfonylmethane Supplementation Improves Immune, DAMP Signaling, and Inflammatory Responses During Recovery From All-Out Running Efforts. Front. Physiol. 2021, 12, 712731. [Google Scholar] [CrossRef] [PubMed]
- McFarlin, B.K.; Venable, A.S.; Henning, A.L.; Sampson, J.N.; Pennel, K.; Vingren, J.L.; Hill, D.W. Reduced inflammatory and muscle damage biomarkers following oral supplementation with bioavailable curcumin. BBA Clin. 2016, 5, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Tanner, E.A.; Gary, M.A.; Davis, A.A.; Michalik, S.; McFarlin, B.K. Alterations in Systemic Inflammatory Response Following a Half-Marathon Race with a Combined Curcumin and Pomegranate Supplement: A Feasibility Study. J. Diet. Suppl. 2021, 18, 461–477. [Google Scholar] [CrossRef]
- Tanner, E.A.; Gary, M.A.; Michalik, S.; Davis, A.A.; McFarlin, B.K. Optimized Curcumin, Pomegranate Extract, and Methylsulfonylmethane Reduce Acute, Systemic Inflammatory Response to a Half-marathon Race. Altern. Ther. Health Med. 2022, 28, 72–81. [Google Scholar] [PubMed]
- Withee, E.D.; Tippens, K.M.; Dehen, R.; Tibbitts, D.; Hanes, D.; Zwickey, H. Effects of Methylsulfonylmethane (MSM) on exercise-induced oxidative stress, muscle damage, and pain following a half-marathon: A double-blind, randomized, placebo-controlled trial. J. Int. Soc. Sports Nutr. 2017, 14, 24. [Google Scholar] [CrossRef]
- Barmaki, S.; Bohlooli, S.; Khoshkhahesh, F.; Nakhostin-Roohi, B. Effect of methylsulfonylmethane supplementation on exercise-Induced muscle damage and total antioxidant capacity. J. Sports Med. Phys. Fit. 2012, 52, 170–174. [Google Scholar]
- Stuempfle, K.J.; Valentino, T.; Hew-Butler, T.; Hecht, F.M.; Hoffman, M.D. Nausea is associated with endotoxemia during a 161-km ultramarathon. J. Sports Sci. 2016, 34, 1662–1668. [Google Scholar] [CrossRef]
- Wallett, A.; McKune, A.; Pyne, D.; Bishop, D.; Girard, O.; Saunders, P.; Periard, J. Repeated-Sprint Exercise in the Heat Increases Indirect Markers of Gastrointestinal Damage in Well-Trained Team-Sport Athletes. Int. J. Sport Nutr. Exerc. Metab. 2022, 32, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Niemela, M.; Niemela, O.; Bloigu, R.; Bloigu, A.; Kangastupa, P.; Juvonen, T. Serum Calprotectin, a Marker of Neutrophil Activation, and Other Mediators of Inflammation in Response to Various Types of Extreme Physical Exertion in Healthy Volunteers. J. Inflamm. Res. 2020, 13, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, S.; Gemma Nasoni, M.; Palma, F.; Citarella, R.; Luchetti, F. Serum changes in sTWEAK and its scavenger receptor sCD163 in ultramarathon athletes running the 24-h race. Cytokine 2021, 137, 155315. [Google Scholar] [CrossRef] [PubMed]
- Gouwy, M.; Struyf, S.; Leutenez, L.; Portner, N.; Sozzani, S.; Van Damme, J. Chemokines and other GPCR ligands synergize in receptor-mediated migration of monocyte-derived immature and mature dendritic cells. Immunobiology 2014, 219, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Demoule, A.; Divangahi, M.; Yahiaoui, L.; Danialou, G.; Gvozdic, D.; Petrof, B.J. Chemokine receptor and ligand upregulation in the diaphragm during endotoxemia and Pseudomonas lung infection. Mediat. Inflamm. 2009, 2009, 860565. [Google Scholar] [CrossRef]
- Siegfried, A.; Berchtold, S.; Manncke, B.; Deuschle, E.; Reber, J.; Ott, T.; Weber, M.; Kalinke, U.; Hofer, M.J.; Hatesuer, B.; et al. IFIT2 is an effector protein of type I IFN-mediated amplification of lipopolysaccharide (LPS)-induced TNF-alpha secretion and LPS-induced endotoxin shock. J. Immunol. 2013, 191, 3913–3921. [Google Scholar] [CrossRef]
- Smiljanovic, B.; Grun, J.R.; Steinbrich-Zollner, M.; Stuhlmuller, B.; Haupl, T.; Burmester, G.R.; Radbruch, A.; Grutzkau, A.; Baumgrass, R. Defining TNF-alpha- and LPS-induced gene signatures in monocytes to unravel the complexity of peripheral blood transcriptomes in health and disease. J. Mol. Med. 2010, 88, 1065–1079. [Google Scholar] [CrossRef]
- Wezel, A.; van der Velden, D.; Maassen, J.M.; Lagraauw, H.M.; de Vries, M.R.; Karper, J.C.; Kuiper, J.; Bot, I.; Quax, P.H. RP105 deficiency attenuates early atherosclerosis via decreased monocyte influx in a CCR2 dependent manner. Atherosclerosis 2015, 238, 132–139. [Google Scholar] [CrossRef]
- Wen, H.; Hogaboam, C.M.; Lukacs, N.W.; Cook, D.N.; Lira, S.A.; Kunkel, S.L. The chemokine receptor CCR6 is an important component of the innate immune response. Eur. J. Immunol. 2007, 37, 2487–2498. [Google Scholar] [CrossRef]
- Schaaf, B.; Luitjens, K.; Goldmann, T.; van Bremen, T.; Sayk, F.; Dodt, C.; Dalhoff, K.; Droemann, D. Mortality in human sepsis is associated with downregulation of Toll-like receptor 2 and CD14 expression on blood monocytes. Diagn. Pathol. 2009, 4, 12. [Google Scholar] [CrossRef]
- Moller, A.S.; Ovstebo, R.; Haug, K.B.; Joo, G.B.; Westvik, A.B.; Kierulf, P. Chemokine production and pattern recognition receptor (PRR) expression in whole blood stimulated with pathogen-associated molecular patterns (PAMPs). Cytokine 2005, 32, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Buechler, C.; Ritter, M.; Orso, E.; Langmann, T.; Klucken, J.; Schmitz, G. Regulation of scavenger receptor CD163 expression in human monocytes and macrophages by pro- and antiinflammatory stimuli. J. Leukoc. Biol. 2000, 67, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Hintz, K.A.; Rassias, A.J.; Wardwell, K.; Moss, M.L.; Morganelli, P.M.; Pioli, P.A.; Givan, A.L.; Wallace, P.K.; Yeager, M.P.; Guyre, P.M. Endotoxin induces rapid metalloproteinase-mediated shedding followed by up-regulation of the monocyte hemoglobin scavenger receptor CD163. J. Leukoc. Biol. 2002, 72, 711–717. [Google Scholar] [CrossRef]
- Zhang, C.; Liao, Y.; Liu, Z.; Zeng, L.; Peng, Z.; Liao, J.; Yang, Z. Mapping the Multi-Organ miRNA-mRNA Regulatory Network in LPS-Mediated Endotoxemic Mice: Exploring the Shared Underlying Key Genes and Mechanisms. Front. Mol. Biosci. 2020, 7, 573327. [Google Scholar] [CrossRef] [PubMed]
- Marshall, A.; Celentano, A.; Cirillo, N.; Mignogna, M.D.; McCullough, M.; Porter, S. Antimicrobial activity and regulation of CXCL9 and CXCL10 in oral keratinocytes. Eur. J. Oral. Sci. 2016, 124, 433–439. [Google Scholar] [CrossRef]
- Karpati, E.; Kremlitzka, M.; Sandor, N.; Hajnal, D.; Schneider, A.E.; Jozsi, M. Complement Factor H Family Proteins Modulate Monocyte and Neutrophil Granulocyte Functions. Front. Immunol. 2021, 12, 660852. [Google Scholar] [CrossRef]
- Olivar, R.; Luque, A.; Cardenas-Brito, S.; Naranjo-Gomez, M.; Blom, A.M.; Borras, F.E.; Rodriguez de Cordoba, S.; Zipfel, P.F.; Aran, J.M. The Complement Inhibitor Factor H Generates an Anti-Inflammatory and Tolerogenic State in Monocyte-Derived Dendritic Cells. J. Immunol. 2016, 196, 4274–4290. [Google Scholar] [CrossRef]
- McFarlin, B.K.; Venable, A.S.; Carpenter, K.C.; Henning, A.L.; Ogenstad, S. Oral Supplementation with Baker’s Yeast Beta Glucan Is Associated with Altered Monocytes, T Cells and Cytokines following a Bout of Strenuous Exercise. Front. Physiol. 2017, 8, 786. [Google Scholar] [CrossRef]
- Simmons, D.P.; Nguyen, H.N.; Gomez-Rivas, E.; Jeong, Y.; Jonsson, A.H.; Chen, A.F.; Lange, J.K.; Dyer, G.S.; Blazar, P.; Earp, B.E.; et al. SLAMF7 engagement superactivates macrophages in acute and chronic inflammation. Sci. Immunol. 2022, 7, eabf2846. [Google Scholar] [CrossRef]
- Dai, W.; Chen, H.; Jiang, J.; Kong, W.; Wang, Y. Silencing MR-1 attenuates inflammatory damage in mice heart induced by AngII. Biochem. Biophys. Res. Commun. 2010, 391, 1573–1578. [Google Scholar] [CrossRef]
- Abbasi, A.; Hauth, M.; Walter, M.; Hudemann, J.; Wank, V.; Niess, A.M.; Northoff, H. Exhaustive exercise modifies different gene expression profiles and pathways in LPS-stimulated and un-stimulated whole blood cultures. Brain Behav. Immun. 2014, 39, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Ao, S.; Zhou, J.; Li, J.; Liang, X.; Yang, X.; Zhang, H.; Liu, B.; Tang, W.; Liu, H.; et al. Methylsulfonylmethane protects against lethal dose MRSA-induced sepsis through promoting M2 macrophage polarization. Mol. Immunol. 2022, 146, 69–77. [Google Scholar] [CrossRef]
- Sheikh, F.; Dickensheets, H.; Gamero, A.M.; Vogel, S.N.; Donnelly, R.P. An essential role for IFN-beta in the induction of IFN-stimulated gene expression by LPS in macrophages. J. Leukoc. Biol. 2014, 96, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Vincent, F.B.; Saulep-Easton, D.; Figgett, W.A.; Fairfax, K.A.; Mackay, F. The BAFF/APRIL system: Emerging functions beyond B cell biology and autoimmunity. Cytokine Growth Factor Rev. 2013, 24, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, A.D.; Vella, A.T.; Croft, M. OX-40: Life beyond the effector T cell stage. Semin. Immunol. 1998, 10, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Gwyer Findlay, E.; Danks, L.; Madden, J.; Cavanagh, M.M.; McNamee, K.; McCann, F.; Snelgrove, R.J.; Shaw, S.; Feldmann, M.; Taylor, P.C.; et al. OX40L blockade is therapeutic in arthritis, despite promoting osteoclastogenesis. Proc. Natl. Acad. Sci. USA 2014, 111, 2289–2294. [Google Scholar] [CrossRef]
- McFarlin, B.K.; Carpenter, K.C.; Davidson, T.; McFarlin, M.A. Baker’s yeast beta glucan supplementation increases salivary IgA and decreases cold/flu symptomatic days after intense exercise. J. Diet. Suppl. 2013, 10, 171–183. [Google Scholar] [CrossRef]
- Mah, E.; Kaden, V.N.; Kelley, K.M.; Liska, D.J. Soluble and Insoluble Yeast beta-Glucan Differentially Affect Upper Respiratory Tract Infection in Marathon Runners: A Double-Blind, Randomized Placebo-Controlled Trial. J. Med. Food 2020, 23, 416–419. [Google Scholar] [CrossRef]
- McFarlin, B.K.; Tanner, E.A.; Hill, D.W.; Vingren, J.L. Prebiotic/probiotic supplementation resulted in reduced visceral fat and mRNA expression associated with adipose tissue inflammation, systemic inflammation, and chronic disease risk. Genes Nutr. 2022, 17, 15. [Google Scholar] [CrossRef]
- da Silva, W.; Machado, A.S.; Lemos, A.L.; de Andrade, C.F.; Priego-Quesada, J.I.; Carpes, F.P. Relationship between exercise-induced muscle soreness, pain thresholds, and skin temperature in men and women. J. Therm. Biol. 2021, 100, 103051. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Baxter, J.; Eischer, C.; Gage, M.; Hunter, S.; Yoon, T. Sex differences in neuromuscular function after repeated eccentric contractions of the knee extensor muscles. Eur. J. Appl. Physiol. 2017, 117, 1119–1130. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, K.C.; Breslin, W.L.; Davidson, T.; Adams, A.; McFarlin, B.K. Baker’s yeast beta-glucan supplementation increases monocytes and cytokines post-exercise: Implications for infection risk? Br. J. Nutr. 2013, 109, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Perkins, J.R.; Dawes, J.M.; McMahon, S.B.; Bennett, D.L.; Orengo, C.; Kohl, M. ReadqPCR and NormqPCR: R packages for the reading, quality checking and normalisation of RT-qPCR quantification cycle (Cq) data. BMC Genom. 2012, 13, 296. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.A. The Benjamini-Hochberg method in the case of discrete test statistics. Int. J. Biostat. 2007, 3, 11. [Google Scholar] [CrossRef] [PubMed]




| Variable | Placebo (N = 7) | Supplement (N = 5) |
|---|---|---|
| Sex (F) | 2 | 4 |
| (M) | 5 | 1 |
| Age (Y) | 22 ± 1 | 20 ± 1 |
| Height (cm) | 177.6 ± 3.8 | 166.9 ± 3.8 |
| Weight (kg) | 72.9 ± 4.9 | 64.8 ± 5.1 |
| BMI (kg/m2) | 33.1 ± 2.2 | 29.4 ± 2.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McFarlin, B.K.; Vingren, J.L.; Hill, D.W.; Bridgeman, E.A. MSM Supplementation Is Associated with Reduced Inflammation and Improved Innate Immune Response following In Vitro LPS-Stimulation in Humans after a Bout of Downhill Running. Muscles 2023, 2, 204-217. https://doi.org/10.3390/muscles2020015
McFarlin BK, Vingren JL, Hill DW, Bridgeman EA. MSM Supplementation Is Associated with Reduced Inflammation and Improved Innate Immune Response following In Vitro LPS-Stimulation in Humans after a Bout of Downhill Running. Muscles. 2023; 2(2):204-217. https://doi.org/10.3390/muscles2020015
Chicago/Turabian StyleMcFarlin, Brian K., Jakob L. Vingren, David W. Hill, and Elizabeth A. Bridgeman. 2023. "MSM Supplementation Is Associated with Reduced Inflammation and Improved Innate Immune Response following In Vitro LPS-Stimulation in Humans after a Bout of Downhill Running" Muscles 2, no. 2: 204-217. https://doi.org/10.3390/muscles2020015
APA StyleMcFarlin, B. K., Vingren, J. L., Hill, D. W., & Bridgeman, E. A. (2023). MSM Supplementation Is Associated with Reduced Inflammation and Improved Innate Immune Response following In Vitro LPS-Stimulation in Humans after a Bout of Downhill Running. Muscles, 2(2), 204-217. https://doi.org/10.3390/muscles2020015