
Improving Plant Performance Through Microbiome Manipulation: The Potential Role of Current Bioengineering Approaches

 ,
,  ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Plant–Microbes Interactions and Their Significance
3. Functional Aspect of Plant Microbiomes
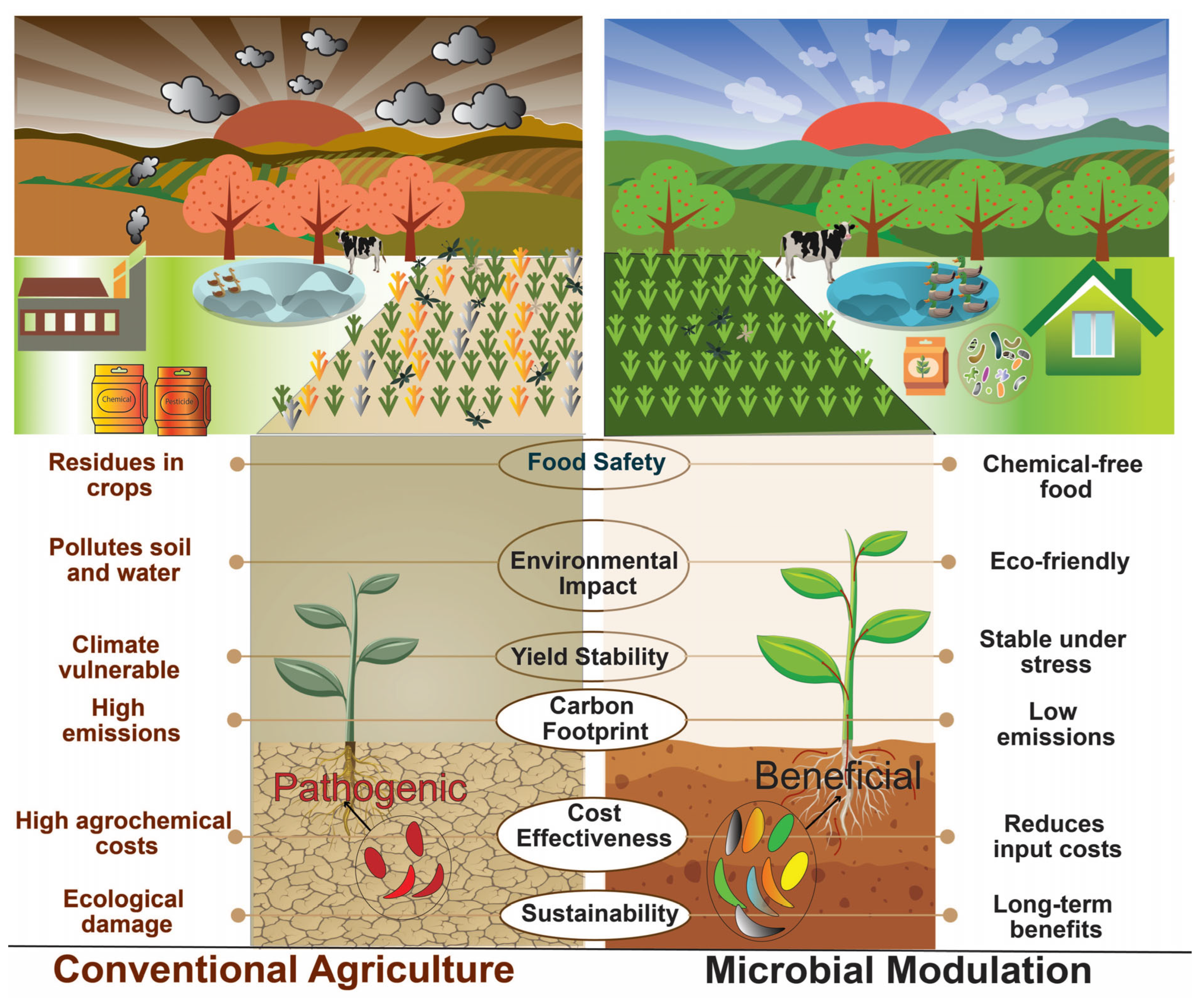
4. Dysbiosis in Plant Microbiomes
5. Approaches for Microbial Manipulation
5.1. Crop Rotation
5.2. Organic Fertilization
5.3. Tillage Intensity
5.4. Reconstructing the Existing Microbiome by Microbial Inoculation
6. Engineering Approach to Improve Plant Performance
6.1. Rhizosphere Engineering
6.2. Microbiome Engineering
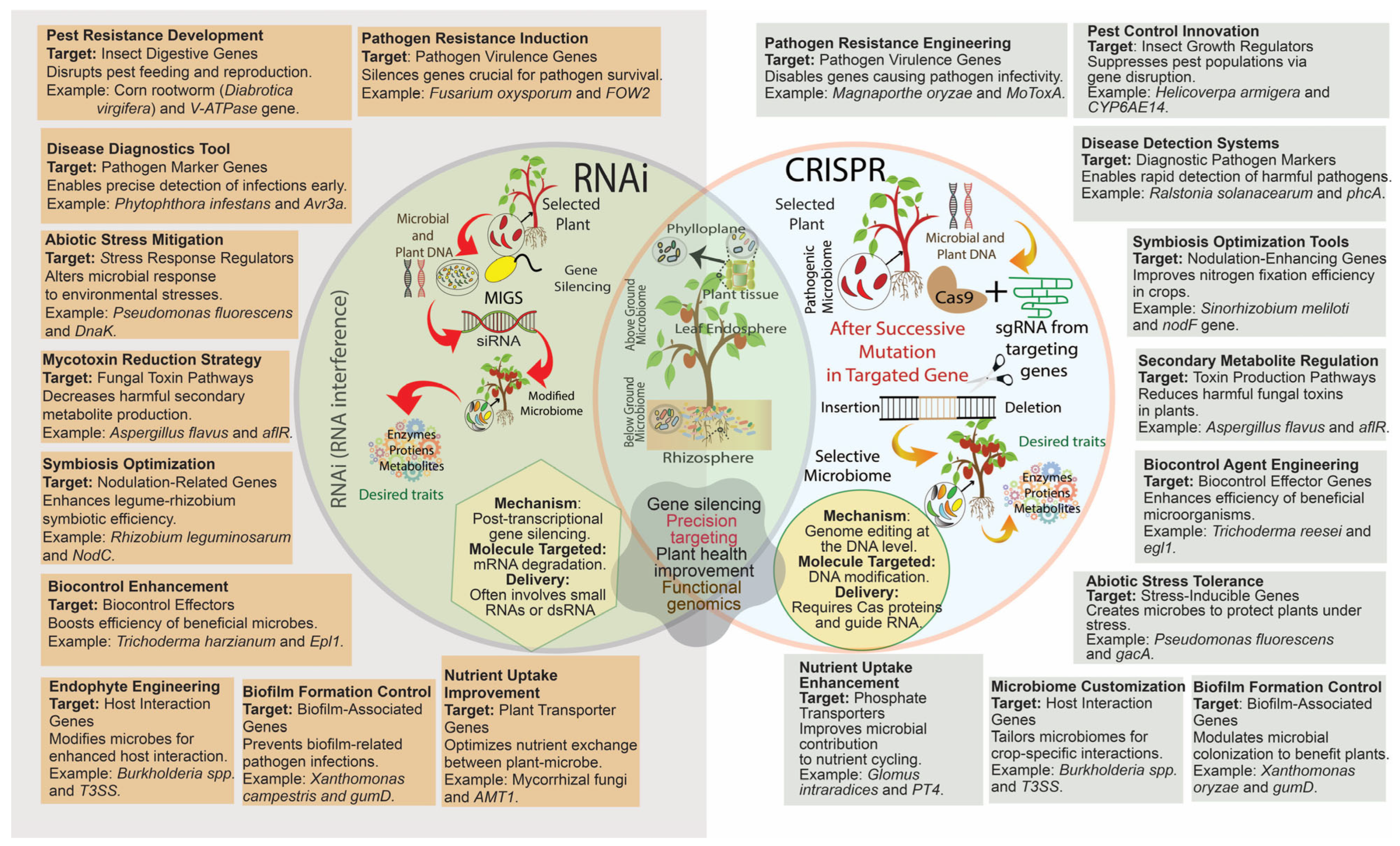
6.2.1. CRISPER/CAS
6.2.2. RNA Interference (RNAi)
7. Future Prospects and Recent Developments
7.1. Role of Exudate Metabolites
7.2. Use of Soil Probiotics
7.3. Optimization of Inoculant Size and Growth Conditions Under In Situ Conditions
7.4. Role of NGS, Metagenomics of Uncultured Microbiome, and Use of Genetic Tools
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arif, I.; Batool, M.; Schenk, P.M. Plant Microbiome Engineering: Expected Benefits for Improved Crop Growth and Resilience. Trends Biotechnol. 2020, 38, 1385–1396. [Google Scholar] [CrossRef] [PubMed]
- Afridi, M.S.; Ali, S.; Salam, A.; César Terra, W.; Hafeez, A.; Sumaira; Ali, B.; AlTami, M.S.; Ameen, F.; Ercisli, S.; et al. Plant Microbiome Engineering: Hopes or Hypes. Biology 2022, 11, 1782. [Google Scholar] [CrossRef] [PubMed]
- Mesny, F.; Hacquard, S.; Thomma, B.P. Co-Evolution within the Plant Holobiont Drives Host Performance. EMBO Rep. 2023, 24, e57455. [Google Scholar] [CrossRef] [PubMed]
- Lyu, D.; Msimbira, L.A.; Nazari, M.; Antar, M.; Pagé, A.; Shah, A.; Monjezi, N.; Zajonc, J.; Tanney, C.A.; Backer, R.; et al. The Coevolution of Plants and Microbes Underpins Sustainable Agriculture. Microorganisms 2021, 9, 1036. [Google Scholar] [CrossRef]
- Glick, B.R.; Gamalero, E. Recent Developments in the Study of Plant Microbiomes. Microorganisms 2021, 9, 1533. [Google Scholar] [CrossRef]
- Compant, S.; Cassan, F.; Kostić, T.; Johnson, L.; Brader, G.; Trognitz, F.; Sessitsch, A. Harnessing the Plant Microbiome for Sustainable Crop Production. Nat. Rev. Microbiol. 2024, 23, 9–23. [Google Scholar] [CrossRef]
- Solomon, W.; Janda, T.; Molnár, Z. Unveiling the significance of rhizosphere: Implications for plant growth, stress response, and sustainable agriculture. Plant Physiol. Biochem. 2024, 206, 108290. [Google Scholar] [CrossRef]
- Kaul, S.; Choudhary, M.; Gupta, S.; Dhar, M.K. Engineering Host Microbiome for Crop Improvement and Sustainable Agriculture. Front. Microbiol. 2021, 12, 635917. [Google Scholar] [CrossRef]
- Solanki, M.K.; Joshi, N.C.; Singh, P.K.; Singh, S.K.; Santoyo, G.; de Azevedo, L.C.B.; Kumar, A. From Concept to Reality: Transforming Agriculture through Innovative Rhizosphere Engineering for Plant Health and Productivity. Microbiol. Res. 2023, 279, 127553. [Google Scholar] [CrossRef]
- Wang, Z.; Solanki, M.K.; Kumar, A.; Solanki, A.C.; Pang, F.; Ba, Z.X.; Niu, J.Q.; Ren, Z.X. Promoting Plant Resilience Against Stress by Engineering Root Microenvironment with Streptomyces Inoculants. Microbiol. Res. 2023, 277, 127509. [Google Scholar] [CrossRef]
- Lau, S.E.; Teo, W.F.A.; Teoh, E.Y.; Tan, B.C. Microbiome Engineering and Plant Biostimulants for Sustainable Crop Improvement and Mitigation of Biotic and Abiotic Stresses. Discov. Food 2022, 2, 9. [Google Scholar] [CrossRef]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial Interactions within the Plant Holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Nadarajah, K.; Abdul Rahman, N.S.N. Plant–Microbe Interaction: Aboveground to Belowground, from the Good to the Bad. Int. J. Mol. Sci. 2021, 22, 10388. [Google Scholar] [CrossRef] [PubMed]
- Mommer, L.; Hinsinger, P.; Prigent-Combaret, C.; Visser, E.J. Advances in the Rhizosphere: Stretching the Interface of Life. Plant Soil 2016, 407, 1–8. [Google Scholar] [CrossRef]
- Galindo-Castañeda, T.; Hartmann, M.; Lynch, J.P. Location: Root Architecture Structures Rhizosphere Microbial Associations. J. Exp. Bot. 2024, 75, 594–604. [Google Scholar] [CrossRef]
- Hartman, K.; van der Heijden, M.G.; Roussely-Provent, V.; Walser, J.C.; Schlaeppi, K. Deciphering Composition and Function of the Root Microbiome of a Legume Plant. Microbiome 2017, 5, 2. [Google Scholar] [CrossRef]
- Yuan, T.; Ren, W.; Wang, Z.; Fry, E.L.; Tang, S.; Yin, J.; Zhang, J.; Jia, Z. How Does the Pattern of Root Metabolites Regulating Beneficial Microorganisms Change with Different Grazing Pressures? Front. Plant Sci. 2023, 14, 1180576. [Google Scholar] [CrossRef]
- Kumar, C.; Esposito, A.; Bertani, I.; Musonerimana, S.; Midekssa, M.J.; Tesfaye, K.; Derr, D.C.; Donaldson, L.; Piazza, S.; Bez, C.; et al. Sorghum rhizosphere bacteriome studies and generation of multistrain beneficial bacterial consortia. Microbiol. Res. 2025, 292, 128036. [Google Scholar] [CrossRef]
- Chai, Y.N.; Schachtman, D.P. Root Exudates Impact Plant Performance under Abiotic Stress. Trends Plant Sci. 2022, 27, 80–91. [Google Scholar] [CrossRef]
- Brummell, D.A.; Dal Cin, V.; Crisosto, C.H.; Labavitch, J.M. Cell Wall Metabolism During Maturation, Ripening and Senescence of Peach Fruit. J. Exp. Bot. 2004, 55, 2029–2039. [Google Scholar] [CrossRef]
- Fierer, N. Embracing the Unknown: Disentangling the Complexities of the Soil Microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zuo, S.; Xu, L.; Zou, Y.; Song, W. Study on Diversity of Endophytic Bacterial Communities in Seeds of Hybrid Maize and Their Parental Lines. Arch. Microbiol. 2012, 194, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; Hardoim, C.C.; van Overbeek, L.S.; van Elsas, J.D. Dynamics of Seed-Borne Rice Endophytes on Early Plant Growth Stages. PLoS ONE 2012, 7, e30438. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Yan, Y.; Li, X.; Liu, Y.; Lu, X. Rhizosphere Soil Microbial Community and Its Response to Different Utilization Patterns in the Semi-Arid Alpine Grassland of Northern Tibet. Front. Microbiol. 2022, 13, 931795. [Google Scholar] [CrossRef]
- Nan, J.; Chao, L.; Ma, X.; Xu, D.; Mo, L.; Zhang, X.; Zhao, X.; Bao, Y. Microbial Diversity in the Rhizosphere Soils of Three Stipa Species from the Eastern Inner Mongolian Grasslands. Glob. Ecol. Conserv. 2020, 22, e00992. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Y.; Zhang, P.; Trivedi, P.; Riera, N.; Wang, Y.; Liu, X.; Fan, G.; Tang, J.; Coletta-Filho, H.D.; et al. The Structure and Function of the Global Citrus Rhizosphere Microbiome. Nat. Commun. 2018, 9, 4894. [Google Scholar] [CrossRef]
- Liu, H.; Carvalhais, L.C.; Crawford, M.; Singh, E.; Dennis, P.G.; Pieterse, C.M.; Schenk, P.M. Inner Plant Values: Diversity, Colonization and Benefits from Endophytic Bacteria. Front. Microbiol. 2017, 8, 2552. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Ver Loren van Themaat, E.; Schulze-Lefert, P. Structure and Functions of the Bacterial Microbiota of Plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef]
- Sessitsch, A.; Hardoim, P.; Döring, J.; Weilharter, A.; Krause, A.; Woyke, T.; Mitter, B.; Hauberg-Lotte, L.; Friedrich, F.; Rahalkar, M.; et al. Functional Characteristics of an Endophyte Community Colonizing Rice Roots as Revealed by Metagenomic Analysis. Mol. Plant Microbe Interact. 2012, 25, 28–36. [Google Scholar] [CrossRef]
- Singh, R.; Pandey, K.D.; Singh, M.; Singh, S.K.; Hashem, A.; Al-Arjani, A.F.; Abd Allah, E.F.; Singh, P.K.; Kumar, A. Isolation and Characterization of Endophytes Bacterial Strains of Momordica charantia L. and Their Possible Approach in Stress Management. Microorganisms 2022, 10, 290. [Google Scholar] [CrossRef]
- Goulart, M.C.; Cueva-Yesquén, L.G.; Hidalgo Martinez, K.J.; Attili-Angelis, D.; Fantinatti-Garboggini, F. Comparison of Specific Endophytic Bacterial Communities in Different Developmental Stages of Passiflora incarnata Using Culture-Dependent and Culture-Independent Analysis. Microbiol. Open 2019, 8, e896. [Google Scholar] [CrossRef] [PubMed]
- Vincze, É.B.; Becze, A.; Laslo, É.; Mara, G. Beneficial Soil Microbiomes and Their Potential Role in Plant Growth and Soil Fertility. Agriculture 2024, 14, 152. [Google Scholar] [CrossRef]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Agunbiade, V.F.; Babalola, O.O. Drought Stress Amelioration Attributes of Plant-Associated Microbiome on Agricultural Plants. Bioinform. Biol. Insights 2024, 18, 11779322241233442. [Google Scholar] [CrossRef]
- Mejia, G.; Jara-Servin, A.; Romero-Chora, L.; Hernandez-Alvarez, C.; Peimbert, M.; Cruz-Ortega, R.; Alcaraz, L.D. Rhizosphere Microbiome Influence on Tomato Growth under Low-Nutrient Settings. bioRxiv 2024. [Google Scholar] [CrossRef]
- Mukherjee, A.; Singh, B.N.; Kaur, S.; Sharma, M.; de Araújo, A.S.F.; de Araujo Pereira, A.P.; Morya, R.; Puopolo, G.; Moënne-Loccoz, Y.; Melo, V.M.M.; et al. Unearthing the Power of Microbes as Plant Microbiome for Sustainable Agriculture. Microbiol. Res. 2024, 286, 127780. [Google Scholar] [CrossRef]
- Leontidou, K.; Genitsaris, S.; Papadopoulou, A.; Kamou, N.; Bosmali, I.; Matsi, T.; Madesis, P.; Vokou, D.; Karamanoli, K.; Mellidou, I. Plant Growth Promoting Rhizobacteria Isolated from Halophytes and Drought-Tolerant Plants: Genomic Characterisation and Exploration of Phyto-Beneficial Traits. Sci. Rep. 2020, 10, 14857. [Google Scholar] [CrossRef]
- Yang, J.; Kloepper, J.W.; Ryu, C.M. Rhizosphere Bacteria Help Plants Tolerate Abiotic Stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef]
- Ning, Z.; Lin, K.; Gao, M.; Han, X.; Guan, Q.; Ji, X.; Yu, S.; Lu, L. Mitigation of Salt Stress in Rice by the Halotolerant Plant Growth-Promoting Bacterium Enterobacter asburiae D2. J. Xenobiotics 2024, 14, 333–349. [Google Scholar] [CrossRef]
- Khumairah, F.H.; Setiawati, M.R.; Fitriatin, B.N.; Simarmata, T.; Alfaraj, S.; Ansari, M.J.; Enshasy, H.A.E.; Sayyed, R.Z.; Najafi, S. Halotolerant Plant Growth-Promoting Rhizobacteria Isolated from Saline Soil Improve Nitrogen Fixation and Alleviate Salt Stress in Rice Plants. Front. Microbiol. 2022, 13, 905210. [Google Scholar] [CrossRef]
- Sarker, P.K.; Karmoker, D.; Shohan, M.U.S.; Saha, A.K.; Rima, F.S.; Begum, R.A.; Islam, M.R.; Seraj, Z.I. Effects of Multiple Halotolerant Rhizobacteria on the Tolerance, Growth, and Yield of Rice Plants under Salt Stress. Folia Microbiol. 2023, 68, 55–72. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Kan, D.; Tian, M.; Ruan, Y. Phosphate-Solubilizing Bacteria Reshaped the Rhizosphere Microbiome and Metabolic Profile of Wheat to Inhibit Cd Absorption. Environ. Exp. Bot. 2024, 226, 105929. [Google Scholar] [CrossRef]
- Kour, D.; Sharma, B.; Negi, R.; Kumar, S.; Kaur, S.; Kaur, T.; Khan, S.S.; Kour, H.; Ramniwas, S.; Rustegi, S.; et al. Microbial Amelioration of Heavy Metal Toxicity in Plants for Agro-Environmental Sustainability. Water Air Soil Pollut. 2024, 235, 431. [Google Scholar] [CrossRef]
- Samaras, A.; Roumeliotis, E.; Ntasiou, P.; Karaoglanidis, G. Bacillus subtilis MBI600 Promotes Growth of Tomato Plants and Induces Systemic Resistance Contributing to the Control of Soilborne Pathogens. Plants 2021, 10, 1113. [Google Scholar] [CrossRef] [PubMed]
- Kamath, A.; Shukla, A.; Patel, D. Quorum Sensing and Quorum Quenching: Two Sides of the Same Coin. Physiol. Mol. Plant Pathol. 2023, 123, 101927. [Google Scholar] [CrossRef]
- Shreshtha, A.; Schikora, A. AHL-Priming for Enhanced Resistance as a Tool in Sustainable Agriculture. FEMS Microbiol. Ecol. 2020, 96, fiaa226. [Google Scholar]
- Vesuna, A.P.; Nerurkar, A.S. Biocontrol Impact of AHL-Degrading Actinobacteria on Quorum Sensing-Regulated Virulence of Phytopathogen Pectobacterium carotovorum subsp. carotovorum BR1. Plant Soil 2020, 453, 371–388. [Google Scholar] [CrossRef]
- Ketehouli, T.; Pasche, J.; Buttrós, V.H.; Goss, E.M.; Martins, S.J. The Underground World of Plant Disease: Rhizosphere Dysbiosis Reduces Above-Ground Plant Resistance to Bacterial Leaf Spot and Alters Plant Transcriptome. Environ. Microbiol. 2024, 26, e16676. [Google Scholar] [CrossRef]
- Wahab, A.; Muhammad, M.; Munir, A.; Abdi, G.; Zaman, W.; Ayaz, A.; Khizar, C.; Reddy, S.P.P. Role of Arbuscular Mycorrhizal fungi in Regulating Growth, Enhancing Productivity, and Potentially Influencing Ecosystems under Abiotic and Biotic Stresses. Plants 2023, 12, 3102. [Google Scholar] [CrossRef]
- Revillini, D.; David, A.S.; Reyes, A.L.; Knecht, L.D.; Vigo, C.; Allen, P.; Searcy, C.A.; Afkhami, M.E. Allelopathy-Selected Microbiomes Mitigate Chemical Inhibition of Plant Performance. New Phytol. 2023, 240, 2007–2019. [Google Scholar] [CrossRef]
- Yang, L.; Wang, L.; Chu, J.; Zhao, H.; Zhao, J.; Zang, H.; Yang, Y.; Zeng, Z. Improving Soil Quality and Wheat Yield through Diversified Crop Rotations in the North China Plain. Soil Tillage Res. 2024, 244, 106231. [Google Scholar] [CrossRef]
- Gan, Y.; Hamel, C.; O’Donovan, J.T.; Cutforth, H.; Zentner, R.P.; Campbell, C.A.; Niu, Y.; Poppy, L. Diversifying Crop Rotations with Pulses Enhances System Productivity. Sci. Rep. 2015, 5, 14625. [Google Scholar] [CrossRef] [PubMed]
- Venter, Z.S.; Jacobs, K.; Hawkins, H.J. The Impact of Crop Rotation on Soil Microbial Diversity: A Meta-Analysis. Pedobiologia 2016, 59, 215–223. [Google Scholar] [CrossRef]
- Shu, X.; He, J.; Zhou, Z.; Xia, L.; Hu, Y.; Zhang, Y.; Zhang, Y.; Luo, Y.; Chu, H.; Liu, W.; et al. Organic Amendments Enhance Soil Microbial Diversity, Microbial Functionality and Crop Yields: A Meta-Analysis. Sci. Total Environ. 2022, 829, 154627. [Google Scholar] [CrossRef]
- Purohit, H.J.; Pandit, P.; Pal, R.; Warke, R.; Warke, G.M. Soil Microbiome: An Intrinsic Driver for Climate Smart Agriculture. J. Agric. Food Res. 2024, 18, 101433. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial Diversity Drives Multifunctionality in Terrestrial Ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef]
- Sun, X.L.; Zhao, J.; You, Y.M.; Sun, O.J. Soil Microbial Responses to Forest Floor Litter Manipulation and Nitrogen Addition in a Mixed-Wood Forest of Northern China. Sci. Rep. 2016, 6, 19536. [Google Scholar] [CrossRef]
- Rodríguez-Berbel, N.; Soria, R.; Ortega, R.; Lucas-Borja, M.E.; Miralles, I. Benefits of Applying Organic Amendments from Recycled Wastes for Fungal Community Growth in Restored Soils of a Limestone Quarry in a Semiarid Environment. Sci. Total Environ. 2022, 806, 151226. [Google Scholar] [CrossRef]
- Xu, S.; Yuan, M.; Chapman, S.J.; Zheng, N.; Yao, H.; Kuzyakov, Y. Bio-Converted Organic Wastes Shape Microbiota in Maize Rhizosphere: Localization and Identification in Enzyme Hotspots. Soil Biol. Biochem. 2023, 184, 109105. [Google Scholar] [CrossRef]
- Shi, C.H.; Wang, X.Q.; Jiang, S.; Zhang, L.Q.; Luo, J. Revealing the Role of the Rhizosphere Microbiota in Reproductive Growth for Fruit Productivity when Inorganic Fertilizer is Partially Replaced by Organic Fertilizer in Pear Orchard Fields. Microb. Biotechnol. 2023, 16, 1373–1392. [Google Scholar] [CrossRef]
- Wang, L.; Wang, H.; Liu, M.; Xu, J.; Bian, H.; Chen, T.; You, E.; Deng, C.; Wei, Y.; Yang, T.; et al. Effects of Different Fertilization Conditions and Different Geographical Locations on the Diversity and Composition of the Rhizosphere Microbiota of Qingke (Hordeum vulgare L.) Plants in Different Growth Stages. Front. Microbiol. 2023, 14, 1094034. [Google Scholar]
- Ayangbenro, A.S.; Chukwuneme, C.F.; Ayilara, M.S.; Kutu, F.R.; Khantsi, M.; Adeleke, B.S.; Glick, B.R.; Babalola, O.O. Harnessing the Rhizosphere Soil Microbiome of Organically Amended Soil for Plant Productivity. Agronomy 2022, 12, 3179. [Google Scholar] [CrossRef]
- Lahbouki, S.; Hashem, A.; Kumar, A.; Abd_Allah, E.F.; Meddich, A. Integration of Horse Manure Vermicompost Doses and Arbuscular Mycorrhizal Fungi to Improve Fruit Quality and Soil Fertility in Tomato Field Facing Drought Stress. Plants 2024, 13, 1449. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Fan, W.; Xu, G.; Cao, Y.; Zhao, X.; Hao, S.; Deng, B.; Ren, S.; Hu, S. Bio-Organic Fertilizers Improve Dendrocalamus farinosus Growth by Remolding the Soil Microbiome and Metabolome. Front. Microbiol. 2023, 14, 1117355. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, T.; Mei, X.; Wang, N.; Li, X.; Yang, Q.; Dong, C.; Jiang, G.; Lin, J.; Xu, Y.; et al. Bio-Organic Fertilizer Promotes Pear Yield by Shaping the Rhizosphere Microbiome Composition and Functions. Microbiol. Spectr. 2022, 10, e03572-22. [Google Scholar] [CrossRef]
- Govaerts, B.; Mezzalama, M.; Sayre, K.D.; Crossa, J.; Lichter, K.; Troch, V.; Vanherck, K.; De Corte, P.; Deckers, J. Long-Term Consequences of Tillage, Residue Management, and Crop Rotation on Selected Soil Micro-Flora Groups in the Subtropical Highlands. Appl. Soil Ecol. 2008, 38, 197–210. [Google Scholar] [CrossRef]
- Shanmugam, G.S.; Buehring, N.W.; Prevost, J.D.; Kingery, W.L. Soil Bacterial Community Diversity and Composition as Affected by Tillage Intensity Treatments in Corn-Soybean Production Systems. Microbiol. Res. 2021, 12, 157–172. [Google Scholar] [CrossRef]
- Dewi, R.K.; Huang, Q.; Hashimi, R.; Komatsuzaki, M. Enhancing agroecosystem sustainability: Integrative soil health strategies in regenerative organic soybean production on Andosol in Japan. Geoderma Reg. 2025, 40, e00910. [Google Scholar] [CrossRef]
- Hu, X.; Liu, J.; Liang, A.; Gu, H.; Liu, Z.; Jin, J.; Wang, G. Soil metagenomics reveals reduced tillage improves soil functional profiles of carbon, nitrogen, and phosphorus cycling in bulk and rhizosphere soils. Agric. Ecosyst. Environ. 2025, 379, 109371. [Google Scholar] [CrossRef]
- Yang, D.; Liu, Y.; Wang, Y.; Gao, F.; Zhao, J.; Li, Y.; Li, X. Effects of soil tillage, management practices, and mulching film application on soil health and peanut yield in a continuous cropping system. Front. Microbiol. 2020, 11, 570924. [Google Scholar] [CrossRef]
- Sessitsch, A.; Wakelin, S.; Schloter, M.; Maguin, E.; Cernava, T.; Champomier-Verges, M.C.; Charles, T.C.; Cotter, P.D.; Ferrocino, I.; Kriaa, A.; et al. Microbiome Interconnectedness Throughout Environments with Major Consequences for Healthy People and a Healthy Planet. Microbiol. Mol. Biol. Rev. 2023, 87, e00212-22. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, M.; Ballard, R.A.; Wright, D. Soil Microbial Inoculants for Sustainable Agriculture: Limitations and Opportunities. Soil Use Manag. 2022, 38, 1340–1369. [Google Scholar] [CrossRef]
- Beltran-Medina, I.; Romero-Perdomo, F.; Molano-Chavez Gutiérrez, A.Y.; Silva, A.M.; Estrada-Bonilla, G. Inoculation of phosphate-solubilizing bacteria improves soil phosphorus mobilization and maize productivity. Nutr. Cycl. Agroecosyst. 2023, 126, 21–34. [Google Scholar] [CrossRef]
- Nunes, P.S.; Junior, G.V.L.; Mascarin, G.M.; Guimarães, R.A.; Medeiros, F.H.; Arthurs, S.; Bettiol, W. Microbial Consortium of Biological Products: Do They Have a Future? Biol. Control 2024, 188, 105439. [Google Scholar] [CrossRef]
- Cao, Z.; Yan, W.; Ding, M.; Yuan, Y. Construction of Microbial Consortia for Microbial Degradation of Complex Compounds. Front. Bioeng. Biotechnol. 2022, 10, 1051233. [Google Scholar] [CrossRef]
- Wang, Y.; Dall’Agnol, R.F.; Bertani, I.; Bez, C.; Venturi, V. Identification of Synthetic Consortia from a Set of Plant-Beneficial Bacteria. Microb. Biotechnol. 2024, 17, e14330. [Google Scholar] [CrossRef]
- Manjunath, M.; Khokhar, A.; Chary, G.R.; Singh, M.; Yadav, S.K.; Gopinath, K.A.; Jyothilakshmi, N.; Srinivas, K.; Prabhakar, M.; Singh, V.K. Microbial Consortia Enhance the Yield of Maize under Sub-Humid Rainfed Production System of India. Front. Sustain. Food Syst. 2023, 7, 1108492. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Babalola, O.O. Bacterial Consortium for Improved Maize (Zea mays L.) Production. Microorganisms 2019, 7, 519. [Google Scholar] [CrossRef]
- Yin, C.; Hagerty, C.H.; Paulitz, T.C. Synthetic Microbial Consortia Derived from Rhizosphere Soil Protect Wheat Against a Soilborne Fungal Pathogen. Front. Microbiol. 2022, 13, 908981. [Google Scholar] [CrossRef]
- Dessaux, Y.; Grandclément, C.; Faure, D. Engineering the Rhizosphere. Trends Plant Sci. 2016, 21, 266–278. [Google Scholar] [CrossRef]
- Pantigoso, H.A.; Newberger, D.; Vivanco, J.M. The Rhizosphere Microbiome: Plant-Microbial Interactions for Resource Acquisition. J. Appl. Microbiol. 2022, 133, 2864–2876. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P. Harnessing root architecture to address global challenges. Plant J. 2022, 109, 415–431. [Google Scholar] [CrossRef] [PubMed]
- Zhong, V.; Archibald, B.N.; Brophy, J.A.N. Transcriptional and post-transcriptional controls for tuning gene expression in plants. Curr. Opin. Plant Biol. 2023, 71, 102315. [Google Scholar] [CrossRef] [PubMed]
- Yaschenko, A.E.; Fenech, M.; Mazzoni-Putman, S.; Alonso, J.M.; Stepanova, A.N. Deciphering the molecular basis of tissue-specific gene expression in plants: Can synthetic biology help? Curr. Opin. Plant Biol. 2022, 68, 102241. [Google Scholar] [CrossRef]
- Lee, J.Y.; Colinas, J.; Wang, J.Y.; Mace, D.; Ohler, U.; Benfey, P.N. Transcriptional and posttranscriptional regulation of transcription factor expression in Arabidopsis roots. Proc. Natl. Acad. Sci. USA 2006, 103, 6055–6060. [Google Scholar] [CrossRef]
- Postma, J.A.; Dathe, A.; Lynch, J.P. The optimal lateral root branching density for maize depends on nitrogen and phosphorus availability. Plant Physiol. 2014, 166, 590–602. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Shen, Q.; Huang, X.-F.; Vivanco, J.M.; Reardon, K.F.; Zhang, R. Rhizosphere interactions: Root exudates, microbes, and microbial communities. Botany 2014, 92, 267–275. [Google Scholar]
- Cao, Y.; Zhang, Z.; Ling, N.; Yuan, Y.; Zheng, X.; Shen, B.; Shen, Q. Bacillus subtilis SQR 9 can control Fusarium wilt in cucumber by colonizing plant roots. Biol. Fertil. Soils 2011, 47, 495–506. [Google Scholar] [CrossRef]
- Kumar, A.; Dubey, A. Rhizosphere Microbiome: Engineering Bacterial Competitiveness for Enhancing Crop Production. J. Adv. Res. 2020, 24, 337–352. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The Rhizosphere Microbiome: Significance of Plant Beneficial, Plant Pathogenic, and Human Pathogenic Microorganisms. FEMS Microbiol. Rev. 2019, 37, 634–663. [Google Scholar] [CrossRef] [PubMed]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Zhuang, J.; Zhou, C.; Li, H.; Liu, C.; Liu, C.; Liu, W. CRISPR/Cas9-Mediated Genome Editing of the Filamentous Fungus Neurospora crassa. Methods Mol. Biol. 2019, 1620, 145–159. [Google Scholar]
- Bano, S.; Wu, X.; Zhang, X. Towards Sustainable Agriculture: Rhizosphere Microbiome Engineering. Appl. Microbiol. Biotechnol. 2021, 105, 7141–7160. [Google Scholar] [CrossRef]
- Phour, M.; Sindhu, S.S. Mitigating Abiotic Stress: Microbiome Engineering for Improving Agricultural Production and Environmental Sustainability. Planta 2022, 256, 85. [Google Scholar] [CrossRef]
- Albright, M.B.; Louca, S.; Winkler, D.E.; Feeser, K.L.; Haig, S.J.; Whiteson, K.L.; Emerson, J.B.; Dunbar, J. Solutions in Microbiome Engineering: Prioritizing Barriers to Organism Establishment. ISME J. 2022, 16, 331–338. [Google Scholar] [CrossRef]
- Mahmud, K.; Missaoui, A.; Lee, K.; Ghimire, B.; Presley, H.W.; Makaju, S. Rhizosphere Microbiome Manipulation for Sustainable Crop Production. Curr. Plant Biol. 2021, 27, 100210. [Google Scholar] [CrossRef]
- Ke, J.; Wang, B.; Yoshikuni, Y. Microbiome engineering: Synthetic biology of plant-associated microbiomes in sustainable agriculture. Trends Biotechnol. 2021, 39, 244–261. [Google Scholar] [CrossRef]
- Chesneau, G.; Herpell, J.; Garrido-Oter, R.; Hacquard, S. From synthetic communities to synthetic ecosystems: Exploring causalities in plant–microbe–environment interactions. New Phytol. 2025, 245, 496–502. [Google Scholar] [CrossRef]
- Shelake, R.M.; Pramanik, D.; Kim, J.Y. Exploration of plant-microbe interactions for sustainable agriculture in CRISPR era. Microorganisms 2019, 7, 269. [Google Scholar] [CrossRef]
- Prabhukarthikeyan, S.R.; Parameswaran, C.; Keerthana, U.; Teli, B.; Jagannadham, P.T.; Cayalvizhi, B.; Panneerselvam, P.; Senapati, A.; Nagendran, K.; Kumari, S.; et al. Understanding the plant-microbe interactions in CRISPR/Cas9 era: Indeed a sprinting start in marathon. Curr. Genom. 2020, 21, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K.; Barrangou, R. Applications of CRISPR technologies across the food supply chain. Annu. Rev. Food Sci. Technol. 2019, 10, 133–150. [Google Scholar] [CrossRef]
- Hendel, A.; Bak, R.O.; Clark, J.T.; Kennedy, A.B.; Ryan, D.E.; Roy, S.; Steinfeld, I.; Lunstad, B.D.; Kaiser, R.J.; Wilkens, A.B.; et al. Chemically modified guide RNAs enhance CRISPR-Cas genome editing in human primary cells. Nat. Biotechnol. 2015, 33, 985–989. [Google Scholar] [CrossRef] [PubMed]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Giraud, T.; Gladieux, P.; Gavrilets, S. Linking the emergence of fungal plant diseases with ecological speciation. Trends Ecol. Evol. 2010, 25, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Davierwala, A.P.; Reddy, A.P.K.; Lagu, M.D.; Ranjekar, P.K.; Gupta, V.S. Marker-assisted selection of bacterial blight resistance genes in rice. Biochem. Genet. 2001, 39, 261–278. [Google Scholar] [CrossRef]
- Muñoz, I.V.; Sarrocco, S.; Malfatti, L.; Baroncelli, R.; Vannacci, G. CRISPR-Cas for fungal genome editing: A new tool for the management of plant diseases. Front. Plant Sci. 2019, 10, 135. [Google Scholar] [CrossRef]
- Glandorf, D.C. Re-evaluation of biosafety questions on genetically modified biocontrol bacteria. Eur. J. Plant Pathol. 2019, 154, 43–51. [Google Scholar] [CrossRef]
- Susan, A.; Yadav, M.K.; Kar, S.; Aravindan, S.; Ngangkham, U.; Raghu, S.; Prabhukarthikeyan, S.R.; Keerthana, U.; Mukherjee, S.C.; Salam, J.L.; et al. Molecular identification of blast resistance genes in rice landraces from northeastern India. Plant Pathol. 2019, 68, 537–546. [Google Scholar] [CrossRef]
- Cao, Y.; Wu, Y.; Zheng, Z.; Song, F. Overexpression of the rice EREBP-like gene OsBIERF3 enhances disease resistance and salt tolerance in transgenic tobacco. Physiol. Mol. Plant Pathol. 2005, 67, 202–211. [Google Scholar] [CrossRef]
- Malnoy, M.; Viola, R.; Jung, M.H.; Koo, O.J.; Kim, S.; Kim, J.S.; Velasco, R.; Nagamangala Kanchiswamy, C. DNA-free genetically edited grapevine and apple protoplasts using CRISPR/Cas9 ribonucleoproteins. Front. Plant Sci. 2016, 7, 1904. [Google Scholar] [CrossRef] [PubMed]
- de Toledo Thomazella, D.P.; Brail, Q.; Dahlbeck, D.; Staskawicz, B. CRISPR-Cas9 mediated mutagenesis of a DMR6 ortholog in tomato confers broad-spectrum disease resistance. bioRxiv 2016. [Google Scholar] [CrossRef]
- Wang, X.; Tu, M.; Wang, D.; Liu, J.; Li, Y.; Li, Z.; Wang, Y.; Wang, X. CRISPR/Cas9-mediated efficient targeted mutagenesis in grape in the first generation. Plant Biotechnol. J. 2018, 16, 844–855. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y. Citrus Canker Requires Targeting of a Susceptibility Gene by Specific TAL Effectors Present in Xanthomonas Citri; University of Florida: Gainesville, FL, USA, 2013. [Google Scholar]
- Peng, A.; Chen, S.; Lei, T.; Xu, L.; He, Y.; Wu, L.; Yao, L.; Zou, X. Engineering canker-resistant plants through CRISPR/Cas9-targeted editing of the susceptibility gene CsLOB1 promoter in citrus. Plant Biotechnol. J. 2017, 15, 1509–1519. [Google Scholar] [CrossRef]
- Prihatna, C.; Barbetti, M.J.; Barker, S.J. A novel tomato fusarium wilt tolerance gene. Front. Microbiol. 2018, 9, 1226. [Google Scholar] [CrossRef]
- Niu, D.; Hamby, R.; Sanchez, J.N.; Cai, Q.; Yan, Q.; Jin, H. RNAs—A New Frontier in Crop Protection. Curr. Opin. Biotechnol. 2021, 70, 204–212. [Google Scholar] [CrossRef]
- Mezzetti, B.; Smagghe, G.; Arpaia, S.; Christiaens, O.; Dietz-Pfeilstetter, A.; Jones, H.; Kostov, K.; Sabbadini, S.; Opsahl-Sorteberg, H.G.; Ventura, V.; et al. RNAi: What is its position in agriculture? J. Pest Sci. 2020, 93, 1125–1130. [Google Scholar] [CrossRef]
- Nowara, D.; Gay, A.; Lacomme, C.; Shaw, J.; Ridout, C.; Douchkov, D.; Hensel, G.; Kumlehn, J.; Schweizer, P. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell 2010, 22, 3130–3141. [Google Scholar] [CrossRef]
- Koch, A.; Biedenkopf, D.; Furch, A.; Weber, L.; Rossbach, O.; Abdellatef, E.; Linicus, L.; Johannsmeier, J.; Jelonek, L.; Goesmann, A.; et al. An RNAi-based control of Fusarium graminearum infections through spraying of long dsRNAs involves a plant passage and is controlled by the fungal silencing machinery. PLoS Pathog. 2016, 12, e1005901. [Google Scholar] [CrossRef]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Zotti, M.; Dos Santos, E.A.; Cagliari, D.; Christiaens, O.; Taning, C.N.T.; Smagghe, G. RNA interference technology in crop protection against arthropod pests, pathogens, and nematodes. Pest Manag. Sci. 2018, 74, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Di Lelio, I.; Barra, E.; Coppola, M.; Corrado, G.; Rao, R.; Caccia, S. Transgenic plants expressing immunosuppressive dsRNA improve entomopathogen efficacy against Spodoptera littoralis larvae. J. Pest Sci. 2022, 95, 1413–1428. [Google Scholar] [CrossRef]
- Mao, J.; Zeng, F. Plant-mediated RNAi of a gap gene enhances tobacco tolerance against the Myzus persicae. Transgenic Res. 2014, 23, 145–152. [Google Scholar] [CrossRef]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef]
- Carthew, R.W.; Sontheimer, E.J. Origins and mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642. [Google Scholar] [CrossRef]
- Quiza, L.; St-Arnaud, M.; Yergeau, E. Harnessing Phytomicrobiome Signaling for Rhizosphere Microbiome Engineering. Front. Plant Sci. 2015, 6, 507. [Google Scholar] [CrossRef]
- Singh, R.K.; Singh, P.; Sharma, A.; Guo, D.J.; Upadhyay, S.K.; Song, Q.Q.; Verma, K.K.; Li, D.P.; Malviya, M.K.; Song, X.P.; et al. Unraveling Nitrogen Fixing Potential of Endophytic Diazotrophs of Different Saccharum Species for Sustainable Sugarcane Growth. Int. J. Mol. Sci. 2022, 23, 6242. [Google Scholar] [CrossRef]
- Jin, J.; Wang, M.; Lu, W.; Zhang, L.; Jiang, Q.; Jin, Y.; Lu, K.; Sun, S.; Cao, Q.; Wang, Y.; et al. Effect of Plants and Their Root Exudate on Bacterial Activities During Rhizobacterium–Plant Remediation of Phenol from Water. Environ. Int. 2019, 127, 114–124. [Google Scholar] [CrossRef]
- Hemarajata, P.; Versalovic, J. Effects of Probiotics on Gut Microbiota: Mechanisms of Intestinal Immunomodulation and Neuromodulation. Ther. Adv. Gastroenterol. 2013, 6, 39–51. [Google Scholar] [CrossRef]
- Mishra, S.; Mishra, A.; Chauhan, P.S.; Mishra, S.K.; Kumari, M.; Niranjan, A.; Nautiyal, C.S. Pseudomonas putida NBRIC19 Dihydrolipoamide Succinyltransferase (SucB) Gene Controls Degradation of Toxic Allelochemicals Produced by Parthenium hysterophorus. J. Appl. Microbiol. 2012, 112, 793–808. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gao, J.; Bai, Z.; Wu, S.; Li, X.; Wang, N.; Du, X.; Fan, H.; Zhuang, G.; Bohu, T.; et al. Unraveling Mechanisms and Impact of Microbial Recruitment on Oilseed Rape (Brassica napus L.) and the Rhizosphere Mediated by Plant Growth-Promoting Rhizobacteria. Microorganisms 2021, 9, 161. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Philp, J.; Wang, Y.; Hu, J.; Wei, Y.; Li, J.; Ryder, M.; Toh, R.; Zhou, Y.; Denton, M.D.; et al. Plant Growth-Promoting Rhizobacteria Burkholderia vietnamiensis B418 Inhibits Root-Knot Nematode on Watermelon by Modifying the Rhizosphere Microbial Community. Sci. Rep. 2022, 12, 8381. [Google Scholar] [CrossRef] [PubMed]
- Epstein, H.E.; Smith, H.A.; Torda, G.; van Oppen, M.J. Microbiome Engineering: Enhancing Climate Resilience in Corals. Front. Ecol. Environ. 2019, 17, 100–108. [Google Scholar] [CrossRef]
- Knief, C. Analysis of Plant-Microbe Interactions in the Era of Next Generation Sequencing Technologies. Front. Plant Sci. 2014, 5, 216. [Google Scholar] [CrossRef]
- Mueller, U.G.; Linksvayer, T.A. Microbiome breeding: Conceptual and practical issues. Trends Microbiol. 2022, 30, 997–1011. [Google Scholar] [CrossRef]
- Prasannath, K.; Peter Arulraj, S.; Shakthivel, K.; Rakulan, T.; Chandranath Karunarathna, S. Revisiting the ecological significance, composition, and functions of plant microbiome: A review. N. Z. J. Crop Hortic. Sci. 2025, 1–33. [Google Scholar] [CrossRef]
- Misu, I.J.; Kayess, M.O.; Siddiqui, M.N.; Gupta, D.R.; Islam, M.N.; Islam, T. Microbiome engineering for sustainable rice production: Strategies for biofertilization, stress tolerance, and climate resilience. Microorganisms 2025, 13, 233. [Google Scholar] [CrossRef]
- Singh, V.; Upadhyay, R.S.; Sarma, B.K.; Singh, H.B. Seed bio-priming with Trichoderma asperellum effectively modulates plant growth promotion in pea. Int. J. Agric. Environ. Biotechnol. 2016, 9, 361–365. [Google Scholar] [CrossRef]
- Rana, A.; Rani, A.; Nayana, K.R.; Deswal, S.; Singh, A.P.; Rana, S.; Chahar, M.; Singh, N.; Dhaka, R.K. Biotic stress alleviation in plants using rhizobacteria: An overview of mechanism of action, antimicrobial compounds production, (nano) formulations and employment methods. Indian J. Microbiol. 2025, 1–27. [Google Scholar] [CrossRef]
- Qiu, Z.; Egidi, E.; Liu, H.; Kaur, S.; Singh, B.K. New Frontiers in Agriculture Productivity: Optimised Microbial Inoculants and In Situ Microbiome Engineering. Biotechnol. Adv. 2019, 37, 107371. [Google Scholar] [CrossRef] [PubMed]
- Giri, V.P.; Pandey, S.; Kumari, M.; Tripathi, A.; Katiyar, R.; White, J.C.; Mishra, A. Hybridization of Chitosan and Biosynthesized Silver Nanoparticles to Enhance Antimicrobial Activity Against Phytopathogens in Tomato (Solanum lycopersicum). ACS Agric. Sci. Technol. 2022, 2, 719–733. [Google Scholar] [CrossRef]
- Tan, C.; Kalhoro, M.T.; Faqir, Y.; Ma, J.; Osei, M.D.; Khaliq, G. Climate-Resilient Microbial Biotechnology: A Perspective on Sustainable Agriculture. Sustainability 2022, 14, 5574. [Google Scholar] [CrossRef]
- Kumari, M.; Pandey, S.; Mishra, S.K.; Giri, V.P.; Agarwal, L.; Dwivedi, S.; Pandey, A.K.; Nautiyal, C.S.; Mishra, A. Omics-Based Mechanistic Insight into the Role of Bioengineered Nanoparticles for Biotic Stress Amelioration by Modulating Plant Metabolic Pathways. Front. Bioeng. Biotechnol. 2020, 8, 242. [Google Scholar] [CrossRef] [PubMed]
- Jatan, R.; Chauhan, P.S.; Lata, C. High-Throughput Sequencing and Expression Analysis Suggest the Involvement of Pseudomonas putida RA-Responsive miRNAs in Growth and Development of Arabidopsis. Int. J. Mol. Sci. 2020, 21, 5468. [Google Scholar] [CrossRef]
- Chauhan, P.S.; Lata, C.; Tiwari, S.; Chauhan, A.S.; Mishra, S.K.; Agrawal, L.; Chakrabarty, D.; Nautiyal, C.S. Transcriptional Alterations Reveal Bacillus amyloliquefaciens-Rice Cooperation Under Salt Stress. Sci. Rep. 2019, 9, 11912. [Google Scholar] [CrossRef]
- Hakim, S.; Nawaz, M.S.; Siddique, M.J.; Hayat, M.; Gulzar, U.; Imran, A. Metagenomics for Rhizosphere Engineering. In Rhizosphere Engineering; Academic Press: Cambridge, MA, USA, 2022; pp. 395–416. [Google Scholar]
- Lawson, C.E.; Harcombe, W.R.; Hatzenpichler, R.; Lindemann, S.R.; Löffler, F.E.; O’Malley, M.A.; García Martín, H.; Pfleger, B.F.; Raskin, L.; Venturelli, O.S.; et al. Common Principles and Best Practices for Engineering Microbiomes. Nat. Rev. Microbiol. 2019, 17, 725–741. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joshi, D.; Kaushik, A.; Kumar, R.; Arya, A.; Santoyo, G.; Singh, V.K.; Kashyap, N.; Solanki, M.K.; Kumari, M.; Bhardwaj, N.; et al. Improving Plant Performance Through Microbiome Manipulation: The Potential Role of Current Bioengineering Approaches. Bacteria 2025, 4, 12. https://doi.org/10.3390/bacteria4010012
Joshi D, Kaushik A, Kumar R, Arya A, Santoyo G, Singh VK, Kashyap N, Solanki MK, Kumari M, Bhardwaj N, et al. Improving Plant Performance Through Microbiome Manipulation: The Potential Role of Current Bioengineering Approaches. Bacteria. 2025; 4(1):12. https://doi.org/10.3390/bacteria4010012
Chicago/Turabian StyleJoshi, Diksha, Amit Kaushik, Reetesh Kumar, Aditi Arya, Gustavo Santoyo, Vipin Kumar Singh, Nikhil Kashyap, Manoj Kumar Solanki, Madhuree Kumari, Nikunaj Bhardwaj, and et al. 2025. "Improving Plant Performance Through Microbiome Manipulation: The Potential Role of Current Bioengineering Approaches" Bacteria 4, no. 1: 12. https://doi.org/10.3390/bacteria4010012
APA StyleJoshi, D., Kaushik, A., Kumar, R., Arya, A., Santoyo, G., Singh, V. K., Kashyap, N., Solanki, M. K., Kumari, M., Bhardwaj, N., & Kumar, A. (2025). Improving Plant Performance Through Microbiome Manipulation: The Potential Role of Current Bioengineering Approaches. Bacteria, 4(1), 12. https://doi.org/10.3390/bacteria4010012