
Case Report: Inflammation-Driven Species-Level Shifts in the Oral Microbiome of Refractory Feline Chronic Gingivostomatitis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
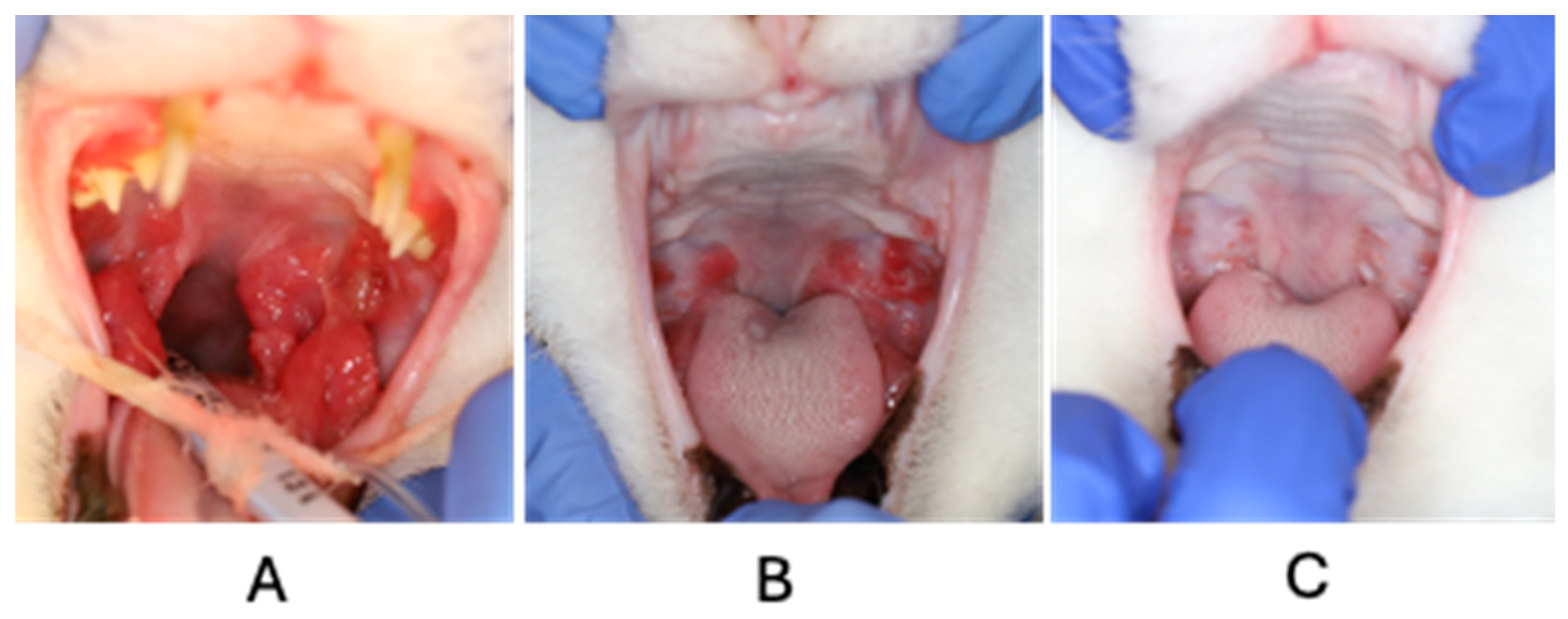
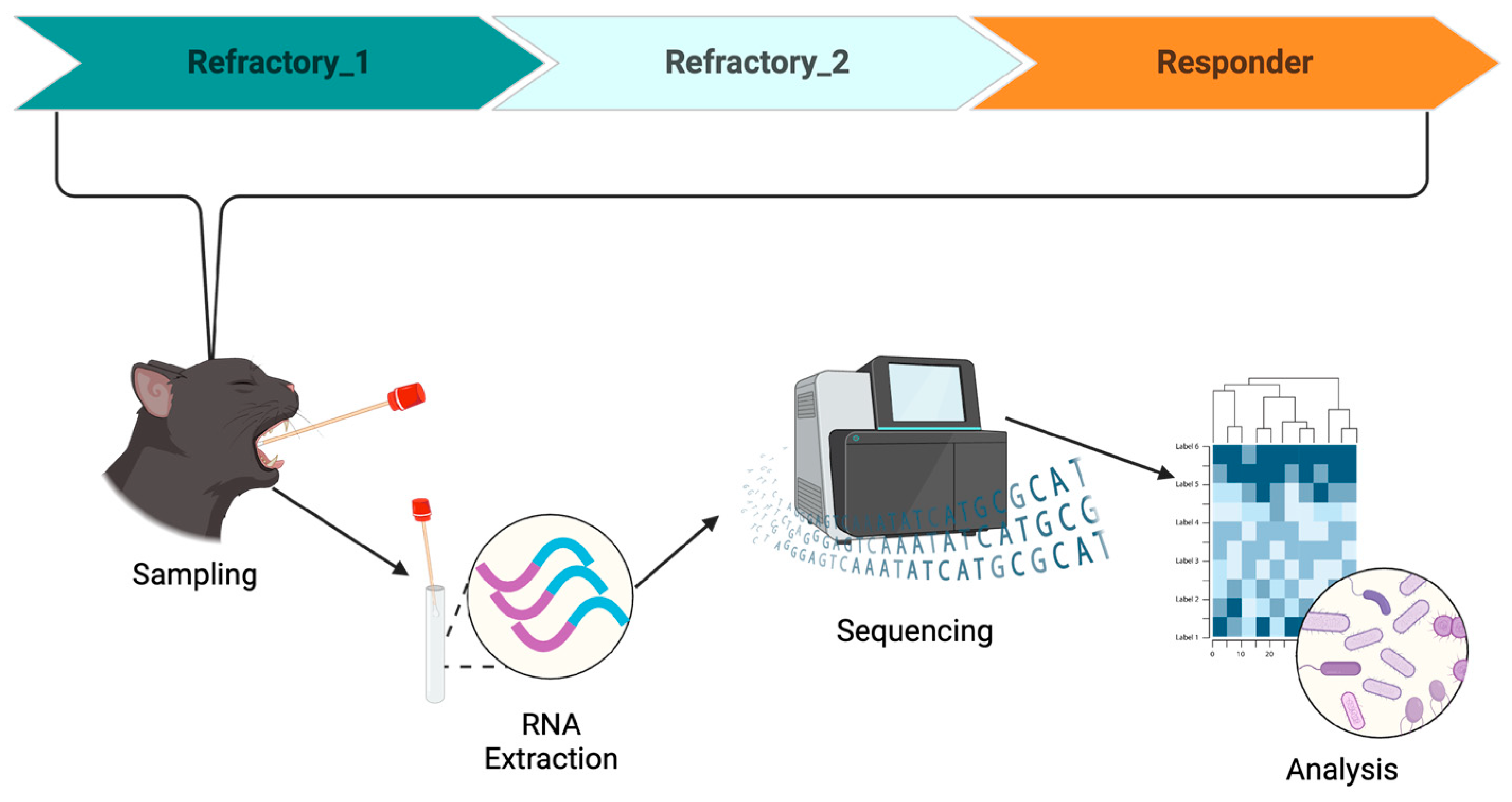
2. Case Description
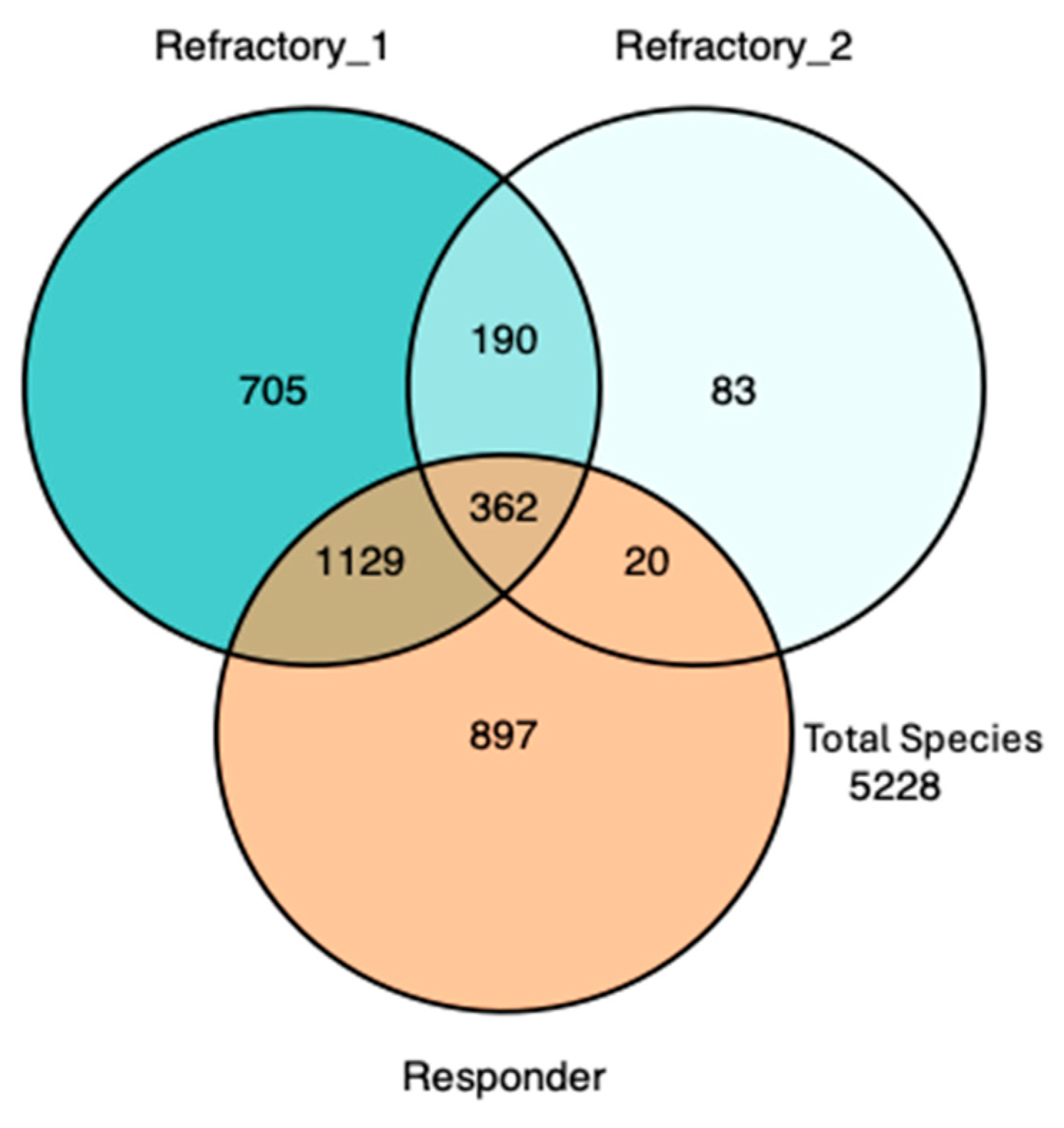
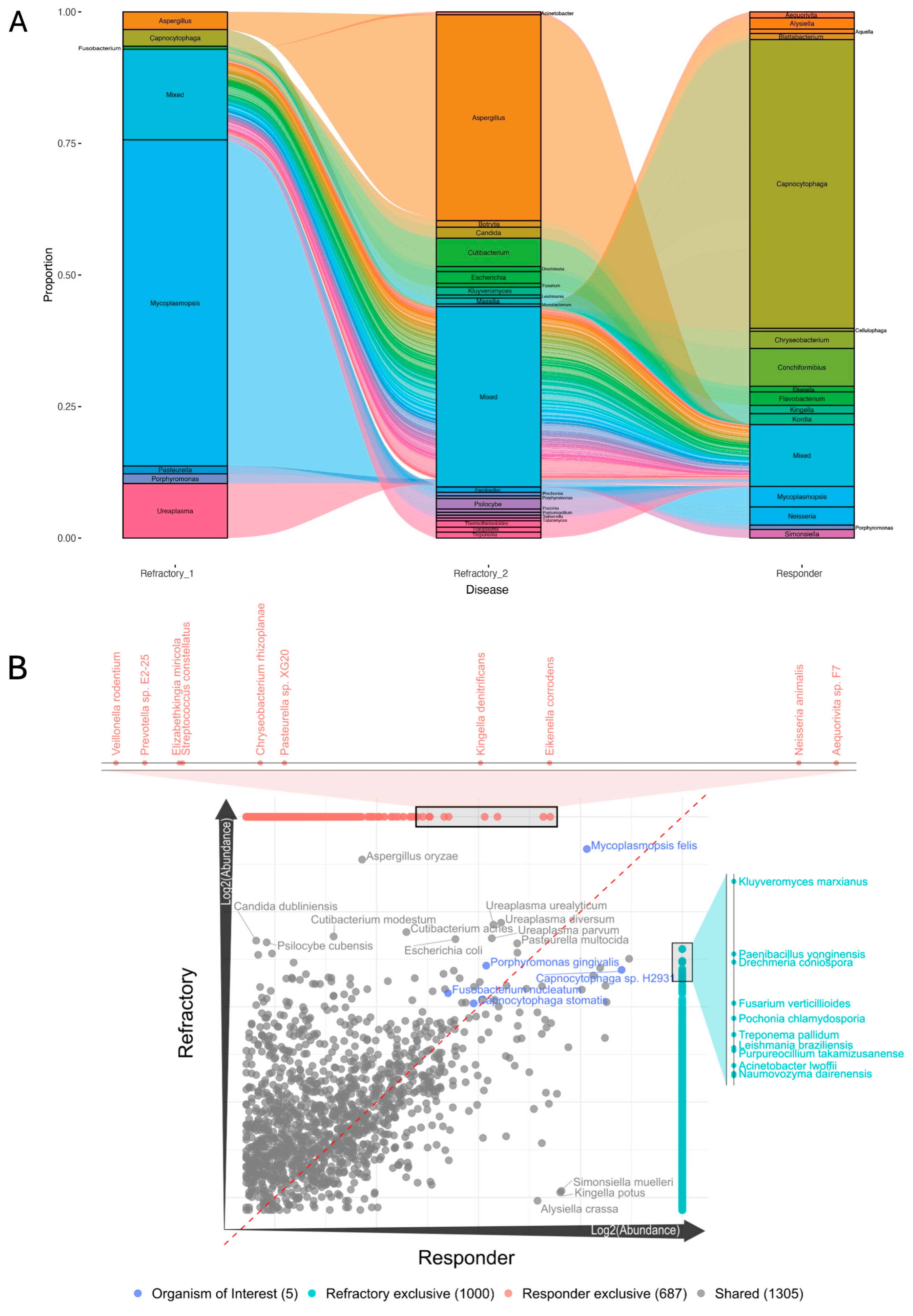
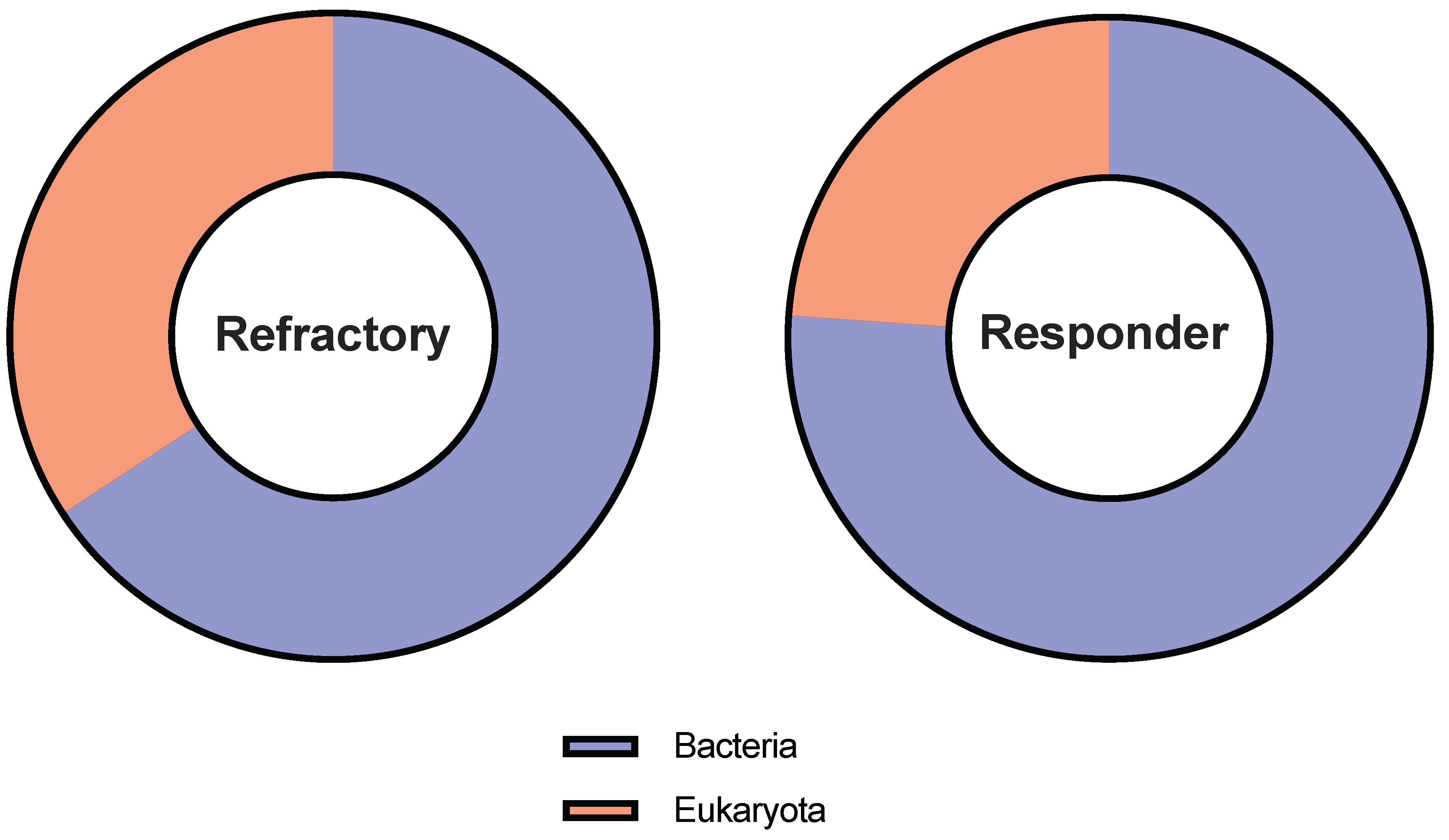
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davis, E.M.; Weese, J.S. Oral Microbiome in Dogs and Cats: Dysbiosis and the Utility of Antimicrobial Therapy in the Treatment of Periodontal Disease. Veter. Clin. N. Am. Small Anim. Pract. 2021, 52, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Krumbeck, J.A.; Reiter, A.M.; Pohl, J.C.; Tang, S.; Kim, Y.J.; Linde, A.; Prem, A.; Melgarejo, T. Characterization of Oral Microbiota in Cats: Novel Insights on the Potential Role of Fungi in Feline Chronic Gingivostomatitis. Pathogens 2021, 10, 904. [Google Scholar] [CrossRef] [PubMed]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: Dynamic communities and host interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef]
- Davis, E.M. Gene Sequence Analyses of the Healthy Oral Microbiome in Humans and Companion Animals. J. Veter. Dent. 2016, 33, 97–107. [Google Scholar] [CrossRef]
- Rodrigues, M.X.; Bicalho, R.C.; Fiani, N.; Lima, S.F.; Peralta, S. The subgingival microbial community of feline periodontitis and gingivostomatitis: Characterization and comparison between diseased and healthy cats. Sci. Rep. 2019, 9, 12340. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.; Croft, J.; O’flynn, C.; Deusch, O.; Colyer, A.; Allsopp, J.; Milella, L.; Davis, I.J. A Pyrosequencing Investigation of Differences in the Feline Subgingival Microbiota in Health, Gingivitis and Mild Periodontitis. PLoS ONE 2015, 10, e0136986. [Google Scholar] [CrossRef]
- Lin, D.; Yang, L.; Wen, L.; Lu, H.; Chen, Q.; Wang, Z. Crosstalk between the oral microbiota, mucosal immunity, and the epithelial barrier regulates oral mucosal disease pathogenesis. Mucosal Immunol. 2021, 14, 1247–1258. [Google Scholar] [CrossRef]
- Akimbekov, N.S.; Digel, I.; Yerezhepov, A.Y.; Shardarbek, R.S.; Wu, X.; Zha, J. Nutritional factors influencing microbiota-mediated colonization resistance of the oral cavity: A literature review. Front. Nutr. 2022, 9, 1029324. [Google Scholar] [CrossRef]
- Soltero-Rivera, M.; Goldschmidt, S.; Arzi, B. Feline chronic gingivostomatitis: Current concepts in clinical management. J. Feline Med. Surg. 2023, 25. [Google Scholar] [CrossRef]
- Soltero-Rivera, M.; Shaw, C.; Arzi, B.; Lommer, M.; Weimer, B.C. Feline Chronic Gingivostomatitis Diagnosis and Treatment through Transcriptomic Insights. Pathogens 2024, 13, 192. [Google Scholar] [CrossRef]
- Soltero-Rivera, M.; Vapniarsky, N.; Rivas, I.L.; Arzi, B. Clinical, radiographic and histopathologic features of early-onset gingivitis and periodontitis in cats (1997–2022). J. Feline Med. Surg. 2023, 25. [Google Scholar] [CrossRef] [PubMed]
- Winer, J.N.; Arzi, B.; Verstraete, F.J.M. Therapeutic Management of Feline Chronic Gingivostomatitis: A Systematic Review of the Literature. Front. Veter. Sci. 2016, 3, 54. [Google Scholar] [CrossRef] [PubMed]
- Bacali, C.; Vulturar, R.; Buduru, S.; Cozma, A.; Fodor, A.; Chiș, A.; Lucaciu, O.; Damian, L.; Moldovan, M.L. Oral Microbiome: Getting to Know and Befriend Neighbors, a Biological Approach. Biomedicines 2022, 10, 671. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, T.; Shiba, T.; Komatsu, K.; Watanabe, T.; Shimogishi, M.; Shibasaki, M.; Koyanagi, T.; Nagai, T.; Katagiri, S.; Takeuchi, Y.; et al. Discrimination of Bacterial Community Structures among Healthy, Gingivitis, and Periodontitis Statuses through Integrated Metatranscriptomic and Network Analyses. mSystems 2021, 6, e0088621. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.G.; Rojas, C.A.; Scarsella, E.; Entrolezo, Z.; Jospin, G.; Hoffman, S.L.; Force, J.; MacLellan, R.H.; Peak, M.; Shope, B.H.; et al. The Oral Microbiome across Oral Sites in Cats with Chronic Gingivostomatitis, Periodontal Disease, and Tooth Resorption Compared with Healthy Cats. Animals 2023, 13, 3544. [Google Scholar] [CrossRef]
- Fried, W.A.; Soltero-Rivera, M.; Ramesh, A.; Lommer, M.J.; Arzi, B.; DeRisi, J.L.; Horst, J.A. Use of unbiased metagenomic and transcriptomic analyses to investigate the association between feline calicivirus and feline chronic gingivostomatitis in domestic cats. Am. J. Veter. Res. 2021, 82, 381–394. [Google Scholar] [CrossRef]
- Peralta, S.; Grenier, J.K.; Webb, S.M.; Miller, A.D.; Miranda, I.C.; Parker, J.S.L. Transcriptomic signatures of feline chronic gingivostomatitis are influenced by upregulated IL6. Sci. Rep. 2023, 13, 13437. [Google Scholar] [CrossRef]
- Hong, B.-Y.; Sobue, T.; Choquette, L.; Dupuy, A.K.; Thompson, A.; Burleson, J.A.; Salner, A.L.; Schauer, P.K.; Joshi, P.; Fox, E.; et al. Chemotherapy-induced oral mucositis is associated with detrimental bacterial dysbiosis. Microbiome 2019, 7, 66. [Google Scholar] [CrossRef]
- Negrini, T.d.C.; Carlos, I.Z.; Duque, C.; Caiaffa, K.S.; Arthur, R.A. Interplay Among the Oral Microbiome, Oral Cavity Conditions, the Host Immune Response, Diabetes Mellitus, and Its Associated-Risk Factors—An Overview. Front. Oral Health 2021, 2, 697428. [Google Scholar] [CrossRef]
- Moreno, C.M.; Boeree, E.; Freitas, C.M.T.; Weber, K.S. Immunomodulatory role of oral microbiota in inflammatory diseases and allergic conditions. Front. Allergy 2023, 4, 1067483. [Google Scholar] [CrossRef]
- Dolieslager, S.M.J.; Lappin, D.F.; Bennett, D.; Graham, L.; Johnston, N.; Riggio, M.P. The influence of oral bacteria on tissue levels of Toll-like receptor and cytokine mRNAs in feline chronic gingivostomatitis and oral health. Veter Immunol. Immunopathol. 2013, 151, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Yama, K.; Nishimoto, Y.; Kumagai, K.; Jo, R.; Harada, M.; Maruyama, Y.; Aita, Y.; Fujii, N.; Inokuchi, T.; Kawamata, R.; et al. Dysbiosis of oral microbiome persists after dental treatment-induced remission of periodontal disease and dental caries. mSystems 2023, 8, e0068323. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, M.; Polizzi, A.; Santonocito, S.; Romano, A.; Lombardi, T.; Isola, G. Impact of Oral Microbiome in Periodontal Health and Periodontitis: A Critical Review on Prevention and Treatment. Int. J. Mol. Sci. 2022, 23, 5142. [Google Scholar] [CrossRef] [PubMed]
- Kong, N.; Ng, W.; Lee, V.; Kelly, L.; Weimer, B. Production and analysis of high molecular weight genomic DNA for NGS pipelines using Agilent DNA extraction kit (p/n 200600). In Agilent Technologies Application Note; Agilent Technologies: Santa Clara, CA, USA, 2013. [Google Scholar]
- Basbas, C.; Garzon, A.; Schlesener, C.; van Heule, M.; Profeta, R.; Weimer, B.C.; Silva-Del-Rio, N.; Byrne, B.A.; Karle, B.; Aly, S.S.; et al. Unveiling the microbiome during post-partum uterine infection: A deep shotgun sequencing approach to characterize the dairy cow uterine microbiome. Anim. Microbiome 2023, 5, 59. [Google Scholar] [CrossRef] [PubMed]
- Garzon, A.; Basbas, C.; Schlesener, C.; Silva-Del-Rio, N.; Karle, B.M.; Lima, F.S.; Weimer, B.C.; Pereira, R.V. WGS of intrauterine E. coli from cows with early postpartum uterine infection reveals a non-uterine specific genotype and virulence factors. mBio 2024, 15, e0102724. [Google Scholar] [CrossRef]
- Beck, K.L.; Haiminen, N.; Chambliss, D.; Edlund, S.; Kunitomi, M.; Huang, B.C.; Kong, N.; Ganesan, B.; Baker, R.; Markwell, P.; et al. Monitoring the microbiome for food safety and quality using deep shotgun sequencing. npj Sci. Food 2021, 5, 3. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/.
- Lu, J.; Breitwieser, F.P.; Thielen, P.; Salzberg, S.L. Bracken: Estimating species abundance in metagenomics data. PeerJ Comput. Sci. 2017, 3, e104. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Alcock, B.P.; Huynh, W.; Chalil, R.; Smith, K.W.; Raphenya, A.R.; A Wlodarski, M.; Edalatmand, A.; Petkau, A.; A Syed, S.; Tsang, K.K.; et al. CARD 2023: Expanded curation, support for machine learning, and resistome prediction at the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2022, 51, D690–D699. [Google Scholar] [CrossRef]
- Yang, J.; Chen, L.; Sun, L.; Yu, J.; Jin, Q. VFDB 2008 release: An enhanced web-based resource for comparative pathogenomics. Nucleic Acids Res. 2007, 36, D539–D542. [Google Scholar] [CrossRef] [PubMed]
- Sturgeon, A.; Pinder, S.L.; Costa, M.C.; Weese, J.S. Characterization of the oral microbiota of healthy cats using next-generation sequencing. Vet. J. 2014, 201, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Sakanaka, A.; Kuboniwa, M.; Shimma, S.; Alghamdi, S.A.; Mayumi, S.; Lamont, R.J.; Fukusaki, E.; Amano, A. Fusobacterium nucleatum Metabolically Integrates Commensals and Pathogens in Oral Biofilms. mSystems 2022, 7, e0017022. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Zhou, F.; Wang, H.; Xu, X.; Xu, S.; Zhang, C.; Zhang, Y.; Lu, M.; Zhang, Y.; Zhou, M.; et al. Systemic antibiotics increase microbiota pathogenicity and oral bone loss. Int. J. Oral Sci. 2023, 15, 4. [Google Scholar] [CrossRef]
- Bartold, P.M.; Van Dyke, T.E. An appraisal of the role of specific bacteria in the initial pathogenesis of periodontitis. J. Clin. Periodontol. 2018, 46, 6–11. [Google Scholar] [CrossRef]
- Van Dyke, T.E.; Bartold, P.M.; Reynolds, E.C. The Nexus Between Periodontal Inflammation and Dysbiosis. Front. Immunol. 2020, 11, 511. [Google Scholar] [CrossRef]
- Blackmore, D.K.; Hill, A.; Jackson, O.F. The incidence of mycoplasma in pet and colony maintained cats. J. Small Anim. Pract. 1971, 12, 207–217. [Google Scholar] [CrossRef]
- Foster, S.; Martin, P.; Braddock, J.; Malik, R. A retrospective analysis of feline bronchoalveolar lavage cytology and microbiology (1995–2000). J. Feline Med. Surg. 2004, 6, 189–198. [Google Scholar] [CrossRef]
- Bemis, D.A. Bordetella and Mycoplasma Respiratory Infections in Dogs and Cats. Veter. Clin. N. Am. Small Anim. Pract. 1992, 22, 1173–1186. [Google Scholar] [CrossRef]
- Hooper, P.T.; Ireland, L.A.; Carter, A. Mycoplasma polyarthritis in a cat with probable severe immune deficiency. Aust. Veter J. 1985, 62, 352. [Google Scholar] [CrossRef]
- Campbell, L.; Fox, J.; Snyder, S. Ocular bacteria and mycoplasma of the clinically normal cat. Feline Pract. 1973, 3, 10–12. [Google Scholar]
- Jin, F.-J.; Hu, S.; Wang, B.-T.; Jin, L. Advances in Genetic Engineering Technology and Its Application in the Industrial Fungus Aspergillus oryzae. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Ledder, R.G.; Madhwani, T.; Sreenivasan, P.K.; De Vizio, W.; McBain, A.J. An in vitro evaluation of hydrolytic enzymes as dental plaque control agents. J. Med. Microbiol. 2009, 58, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, A.M.L.; Belstrøm, D. The role of natural salivary defences in maintaining a healthy oral microbiota. J. Dent. 2019, 80, S3–S12. [Google Scholar] [CrossRef] [PubMed]
- Barachetti, L.; Mortellaro, C.M.; Di Giancamillo, M.; Giudice, C.; Martino, P.; Travetti, O.; Miller, P.E. Bilateral orbital and nasal aspergillosis in a cat. Veter. Ophthalmol. 2009, 12, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Renner, K.; Hill, S.; Grinberg, A.; Weeden, A. Pancreatic candidiasis in a cat. J. Feline Med. Surg. 2021, 7. [Google Scholar] [CrossRef]
- Reagan, K.L.; Dear, J.D.; Kass, P.H.; Sykes, J.E. Risk factors for Candida urinary tract infections in dogs and cats. J. Veter. Intern. Med. 2019, 33, 648–653. [Google Scholar] [CrossRef]
- Hennet, P.R.; Camy, G.A.; McGahie, D.M.; Albouy, M.V. Comparative efficacy of a recombinant feline interferon omega in refractory cases of calicivirus-positive cats with caudal stomatitis: A randomised, multi-centre, controlled, double-blind study in 39 cats. J. Feline Med. Surg. 2011, 13, 577–587. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaw, C.A.; Soltero-Rivera, M.; Profeta, R.; Weimer, B.C. Case Report: Inflammation-Driven Species-Level Shifts in the Oral Microbiome of Refractory Feline Chronic Gingivostomatitis. Bacteria 2025, 4, 1. https://doi.org/10.3390/bacteria4010001
Shaw CA, Soltero-Rivera M, Profeta R, Weimer BC. Case Report: Inflammation-Driven Species-Level Shifts in the Oral Microbiome of Refractory Feline Chronic Gingivostomatitis. Bacteria. 2025; 4(1):1. https://doi.org/10.3390/bacteria4010001
Chicago/Turabian StyleShaw, Claire A., Maria Soltero-Rivera, Rodrigo Profeta, and Bart C. Weimer. 2025. "Case Report: Inflammation-Driven Species-Level Shifts in the Oral Microbiome of Refractory Feline Chronic Gingivostomatitis" Bacteria 4, no. 1: 1. https://doi.org/10.3390/bacteria4010001
APA StyleShaw, C. A., Soltero-Rivera, M., Profeta, R., & Weimer, B. C. (2025). Case Report: Inflammation-Driven Species-Level Shifts in the Oral Microbiome of Refractory Feline Chronic Gingivostomatitis. Bacteria, 4(1), 1. https://doi.org/10.3390/bacteria4010001