

Attenuation of a Field Strain of Infectious Laryngotracheitis Virus in Primary Chicken Culture Cells and Adaptation to Secondary Chicken Embryo Fibroblasts


Abstract
1. Introduction
2. Materials and Methods
2.1. Virus Strain
2.2. Primary Chicken Cells and Cell Line
2.3. Serial Passages
2.4. Conventional PCR, Real-Time PCR, and Reverse Transcriptase (RT)-PCR
2.5. Bacteria Sterility of Tissue Culture Passages
2.6. Preparation of Hyperimmune Sera and Indirect Immunofluorescent Antibody Test
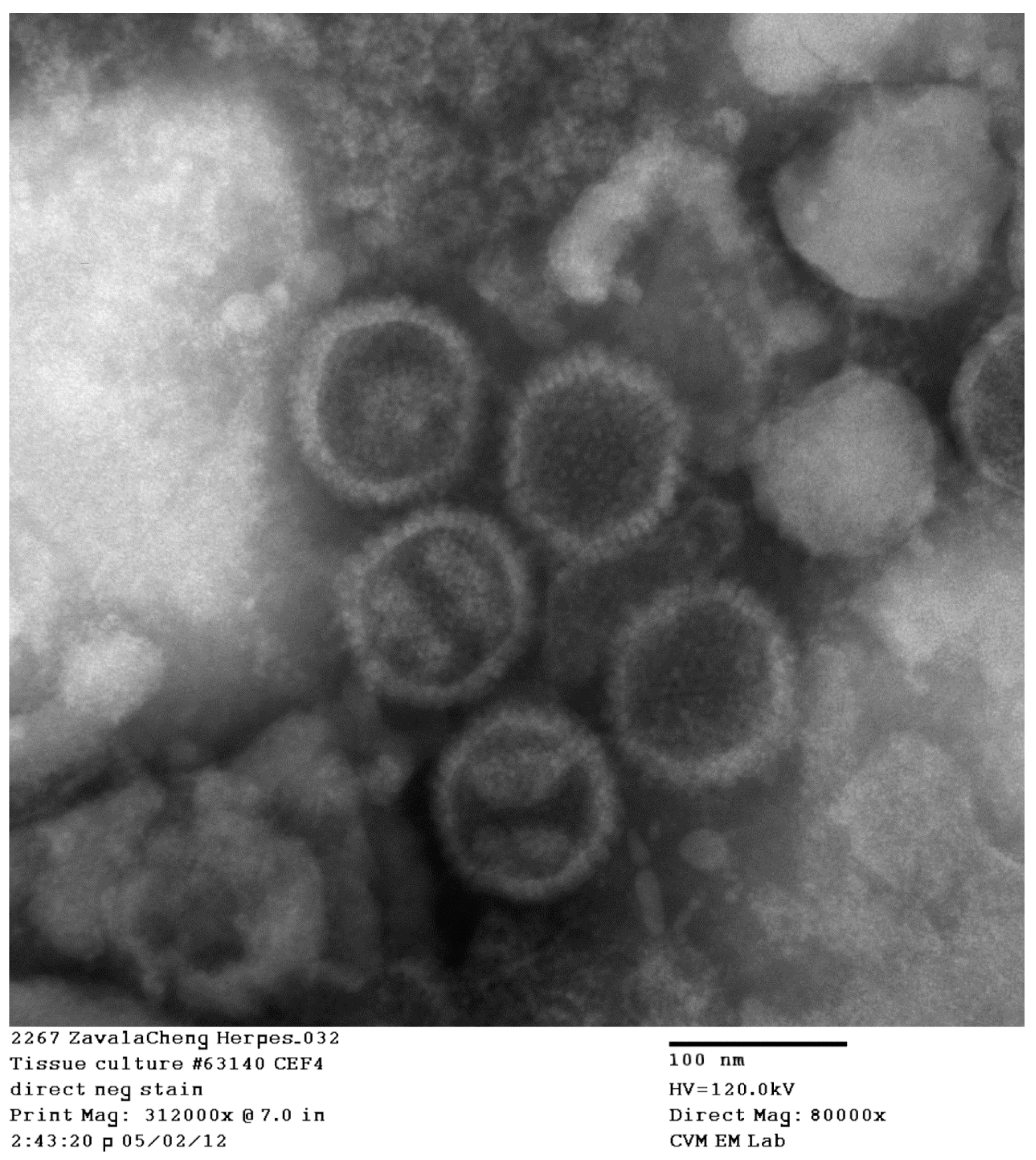
2.7. Electron Microscopy-Negative Stain Technique
2.8. Experimental Design to Evaluate Attenuation of CEL and CEF 63140 Passages
2.9. Statistical Analysis
3. Results
3.1. Serial Passaging in Primary Chicken Cells
3.2. Virus Titration
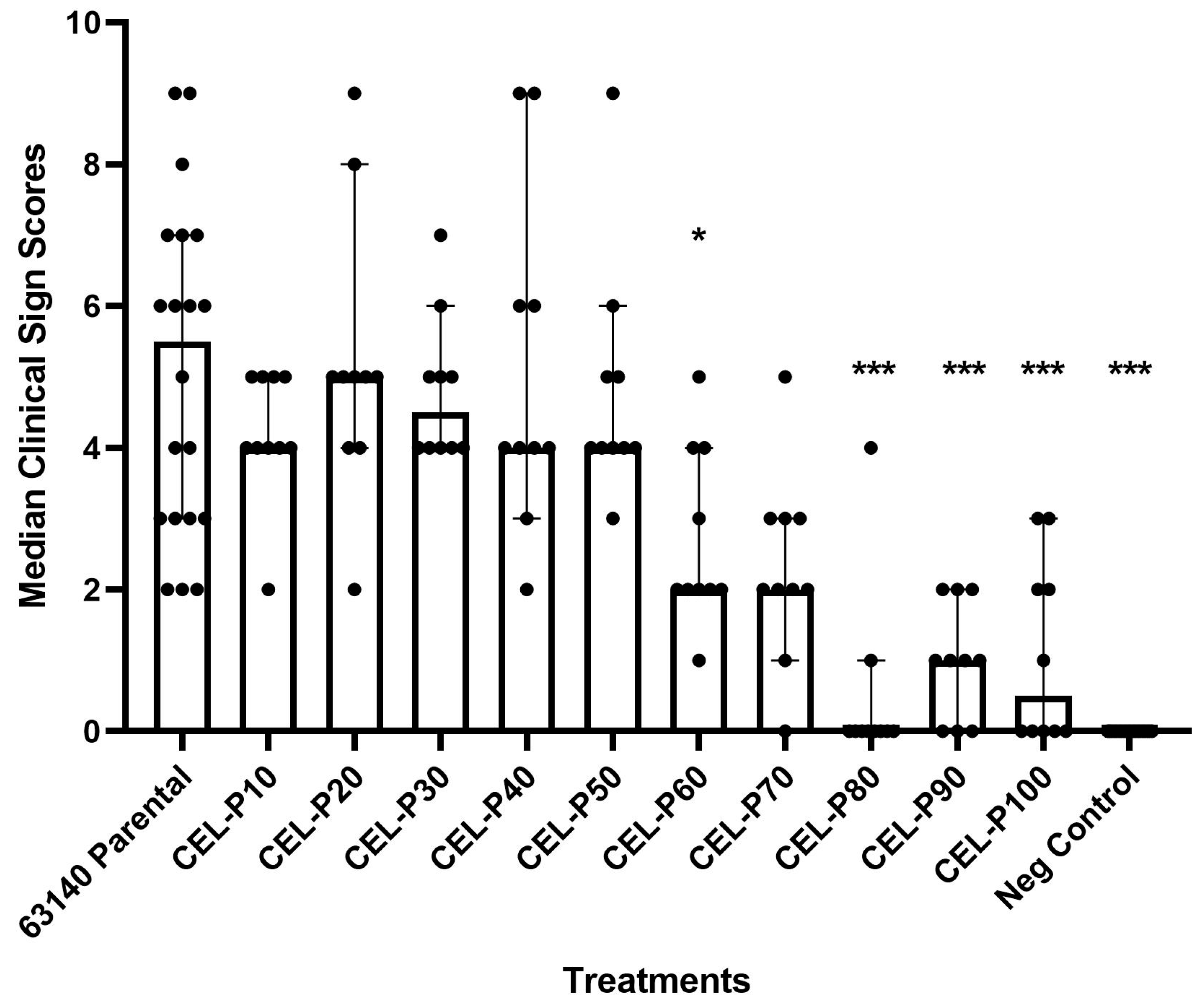
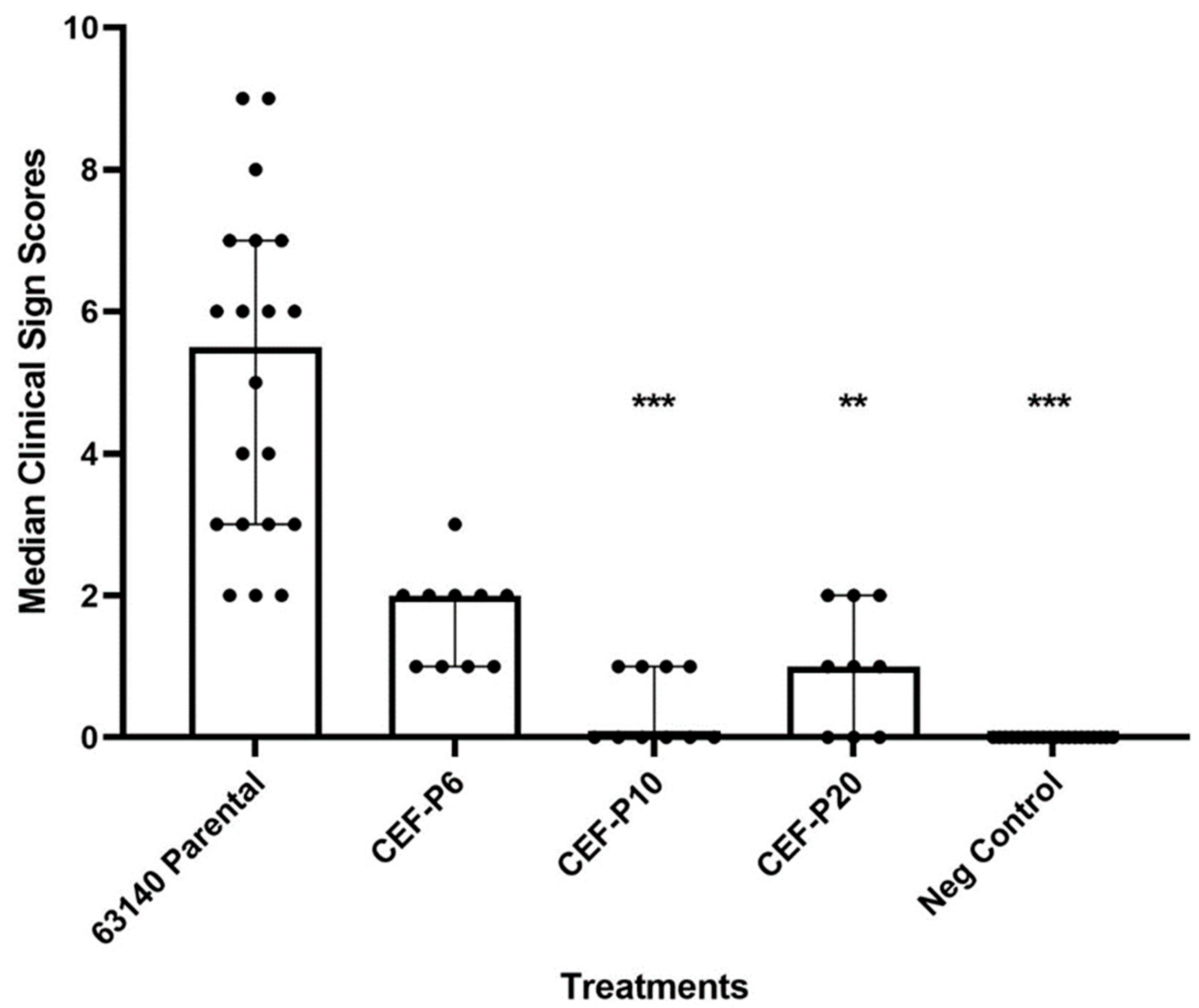
3.3. Attenuation 63140 Passages in CEL and CEF
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garcia, M.; Spatz, S. Infectious Laryngotracheitis. In Diseases of Poultry; Swayne, D.E., Boulianne, M., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., Wit, S., Grimes, T., Johnson, D., Kromm, M., et al., Eds.; Wiley Blackwell: Hoboken, NJ, USA, 2020; pp. 189–209. [Google Scholar]
- Garcia, M.; Zavala, G. Commercial Vaccines and Vaccination Strategies Against Infectious Laryngotracheitis: What We Have Learned and Knowledge Gaps That Remain. Avian Dis. 2019, 63, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Alls, A.A.; Ipson, J.R.; Vaughan, W.D. Studies on an ocular infectious laryngotracheitis vaccine. Avian Dis. 1969, 13, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Gelenczei, E.F.; Marty, E.W. Studies on a Tissue-Culture-Modified Infectious Laryngotracheitis Virus. Avian Dis. 1964, 8, 105–122. [Google Scholar] [CrossRef]
- Hein, R.; Koopman, R.; Garcia, M.; Armour, N.; Dunn, J.R.; Barbosa, T.; Martinez, A. Review of Poultry Recombinant Vector Vaccines. Avian Dis. 2021, 65, 438–452. [Google Scholar] [CrossRef] [PubMed]
- Coppo, M.J.; Noormohammadi, A.H.; Browning, G.F.; Devlin, J.M. Challenges and recent advancements in infectious laryngotracheitis virus vaccines. Avian Pathol. 2013, 42, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Palomino-Tapia, V.A.; Zavala, G.; Cheng, S.; Garcia, M. Long-term protection against a virulent field isolate of infectious laryngotracheitis virus induced by inactivated, recombinant, and modified live virus vaccines in commercial layers. Avian Pathol. 2019, 48, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Vagnozzi, A.; Zavala, G.; Riblet, S.M.; Mundt, A.; Garcia, M. Protection induced by commercially available live-attenuated and recombinant viral vector vaccines against infectious laryngotracheitis virus in broiler chickens. Avian Pathol. 2012, 41, 21–31. [Google Scholar] [CrossRef]
- Parra, S.H.S.; Nuñez, L.F.N.; Ferreira, A.J.P. Epidemiology of Avian Infectious Laryngotracheitis with Special Focus to South America: An update. Braz. J. Poult. Sci. 2016, 18, 551–562. [Google Scholar] [CrossRef]
- Thilakarathne, D.S.; Coppo, M.J.C.; Hartley, C.A.; Diaz-Mendez, A.; Quinteros, J.A.; Fakhri, O.; Vaz, P.K.; Devlin, J.M. Attenuated infectious laryngotracheitis virus vaccines differ in their capacity to establish latency in the trigeminal ganglia of specific pathogen free chickens following eye drop inoculation. PLoS ONE 2019, 14, e0213866. [Google Scholar] [CrossRef]
- Perez Contreras, A.; van der Meer, F.; Checkley, S.; Joseph, T.; King, R.; Ravi, M.; Peters, D.; Fonseca, K.; Gagnon, C.A.; Provost, C.; et al. Analysis of Whole-Genome Sequences of Infectious laryngotracheitis Virus Isolates from Poultry Flocks in Canada: Evidence of Recombination. Viruses 2020, 12, 1302. [Google Scholar] [CrossRef]
- Perez-Contreras, A.; Barboza-Solis, C.; Najimudeen, S.M.; Checkley, S.; Meer, F.V.; Joseph, T.; King, R.; Ravi, M.; Peters, D.; Fonseca, K.; et al. Pathogenic and Transmission Potential of Wildtype and Chicken Embryo Origin (CEO) Vaccine Revertant Infectious Laryngotracheitis Virus. Viruses 2021, 13, 541. [Google Scholar] [CrossRef] [PubMed]
- Salinas-Saavedra, M.; Rock, A.Q.; Martindale, M.Q. Germ layer-specific regulation of cell polarity and adhesion gives insight into the evolution of mesoderm. eLife 2018, 7, e36740. [Google Scholar] [CrossRef] [PubMed]
- Morizono, K.; Chen, I.S. Receptors and tropisms of envelope viruses. Curr. Opin. Virol. 2011, 1, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Nomaguchi, M.; Fujita, M.; Miyazaki, Y.; Adachi, A. Viral Tropism. Front. Microbiol. 2012, 3, 281. [Google Scholar] [CrossRef] [PubMed]
- Barrass, S.V.; Butcher, S.J. Advances in high-throughput methods for the identification of virus receptors. Med. Microbiol. Immunol. 2020, 209, 309–323. [Google Scholar] [CrossRef] [PubMed]
- McFadden, G.; Mohamed, M.R.; Rahman, M.M.; Bartee, E. Cytokine determinants of viral tropism. Nat. Rev. Immunol. 2009, 9, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Koda, Y.; Yamada, S.; Ichihara, T.; Ichihara, T. Experiment on Infectious Laryngotracheitis (ILT) Virus in Tissue Culture Modification and Some Properties of ILT Virus. J. Jpn. Vet. Med. Assoc. 1972, 25, 668–671. [Google Scholar] [CrossRef][Green Version]
- Kawaguchi, T.; Nomura, K.; Hirayama, Y.; Kitagawa, T. Establishment and Characterization of a Chicken Hepatocellular-Carcinoma Cell-Line, LMH. Cancer Res. 1987, 47, 4460–4464. [Google Scholar]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty Per Cent Endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Lei, C.; Yang, J.; Hu, J.; Sun, X. On the Calculation of TCID(50) for Quantitation of Virus Infectivity. Virol. Sin. 2021, 36, 141–144. [Google Scholar] [CrossRef]
- Shapouri, M.R.S.; Kane, M.; Letarte, M.; Bergeron, J.; Arella, M.; Silim, A. Cloning, Sequencing and Expression of the S1 Gene of Avian Reovirus. J. Gen. Virol. 1995, 76, 1515–1520. [Google Scholar] [CrossRef] [PubMed]
- Raue, R.; Hess, M. Hexon based PCRs combined with restriction enzyme analysis for rapid detection and differentiation of fowl adenoviruses and egg drop syndrome virus. J. Virol. Methods 1998, 73, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Kang, S.H.; Bae, Y.J.; Hong, J.T.; Chung, Y.B.; Lee, C.K.; Song, S. PCR-based detection of Mycoplasma species. J. Microbiol. 2006, 44, 42–49. [Google Scholar] [PubMed]
- Williams, S.M.; American Association of Avian Pathologists. A Laboratory Manual for the Isolation, Identification and Characterization of Avian Pathogens, 6th ed.; American Association of Avian Pathologists: Jacksonville, FL, USA, 2016; p. xii. 400p. [Google Scholar]
- Hayat, M.A.; Miller, S.E. Negative Staining; McGraw-Hill Pub. Co.: New York, NY, USA, 1990; 253p. [Google Scholar]
- Hughes, C.S.; Jones, R.C. Comparison of cultural methods for primary isolation of infectious laryngotracheitis virus from field material. Avian Pathol. 1988, 17, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Portz, C.; Lopez de Almeida, L.; Júnior, A.B.; Reck, H.; Franco, A.C.; Canal, C.W. Comparision of different cell cultures for replication of infectious laryngotracheitis virus from chickens. Acta Sci. Vet. 2008, 36, 101–105. [Google Scholar] [CrossRef]
- Schnitzlein, W.M.; Radzevicius, J.; Tripathy, D.N. Propagation of infectious laryngotracheitis virus in an avian liver cell line. Avian Dis. 1994, 38, 211–217. [Google Scholar] [CrossRef]
- Schnitzlein, W.M.; Tripathy, D.N. Replication of infectious laryngotracheitis virus in a quail cell line, QT-35. Avian Dis. 1995, 39, 528–531. [Google Scholar] [CrossRef]
- Pulsford, M.F. The growth of three strains of infectious laryngotracheitis virus of fowls in tissue culture. Aust. J. Exp. Biol. Med. Sci. 1960, 38, 153–162. [Google Scholar] [CrossRef]
- Izuchi, T.; Hasegawa, A.; Miyamoto, T. Studies on a live virus vaccine against infectious laryngotracheitis of chickens. I. Biological properties of attenuated strain C7. Avian Dis. 1983, 27, 918–926. [Google Scholar] [CrossRef]
- Oldoni, I.; Garcia, M. Characterization of infectious laryngotracheitis virus isolates from the US by polymerase chain reaction and restriction fragment length polymorphism of multiple genome regions. Avian Pathol. 2007, 36, 167–176. [Google Scholar] [CrossRef]
- Spatz, S.J.; Volkening, J.D.; Keeler, C.L.; Kutish, G.F.; Riblet, S.M.; Boettger, C.M.; Clark, K.F.; Zsak, L.; Afonso, C.L.; Mundt, E.S.; et al. Comparative full genome analysis of four infectious laryngotracheitis virus (Gallid herpesvirus-1) virulent isolates from the United States. Virus Genes 2012, 44, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Katz, S.L.; Enders, J.F.; Holloway, A. The development and evaluation of an attenuated measles virus vaccine. Am. J. Public Health Nations Health 1962, 52 (Suppl. S2), 5–10. [Google Scholar] [CrossRef] [PubMed]
- Jordan, I.; Sandig, V. Matrix and backstage: Cellular substrates for viral vaccines. Viruses 2014, 6, 1672–1700. [Google Scholar] [CrossRef] [PubMed]
- Gelenczei, E.F.; Marty, E.W. Strain Stability and Immunologic Characteristics of a Tissue-Culture-Modified Infectious Laryngotracheitis Virus. Avian Dis. 1965, 14, 44–56. [Google Scholar] [CrossRef]
- Alcantara, D.; Rodrigues, M.N.; Franciolli, A.L.; Da Fonseca, E.T.; Silva, F.M.; Carvalho, R.C.; Fratini, P.; Sarmento, C.A.; Ferreira, A.J.; Miglino, M.A. Embryonic development of endoderm in chicken (Gallus gallus domesticus). Microsc. Res. Tech. 2013, 76, 803–810. [Google Scholar] [CrossRef]
- Kiecker, C.; Bates, T.; Bell, E. Molecular specification of germ layers in vertebrate embryos. Cell. Mol. Life Sci. 2016, 73, 923–947. [Google Scholar] [CrossRef]
- Sharma, J.M.; Witter, R.L. Embryo Vaccination against Marek’s Disease with Serotypes 1, 2 and 3 Vaccines Administered Singly or in Combination. Avian Dis. 1983, 27, 453–463. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PASSAGE | CEK | CEL | CEFy (Started from CEL-P52) |
|---|---|---|---|
| 06 | ND b | ND b | log10 4.38 (log103.95–4.65) a |
| 10 | log10 6.50 (log106.50–6.50) d | log10 6.38 (log105.95–6.65) c | log10 5.50 (log105.50–5.50) b |
| 20 | log10 4.83 (log104.47–5.32) b | log10 6.63 (log106.35–7.05) c | log10 4.50 (log104.50–4.50) a |
| 30 | log10 3.17 (log102.67–3.52) a | log10 5.83 (log105.47–6.33) bc | ND z |
| 40 | log10 6.63 (log106.34–7.05) d | log10 6.50 (log106.50–6.50) c | ND z |
| 50 | log10 5.83 (log105.47–6.33) cd | log10 5.63 (log105.35–6.05) b | ND z |
| 60 | log10 5.17 (log104.67–5.53) bc | log10 5.68 (log105.15–6.26) b | ND z |
| 70 | ND z | log10 5.38 (log104.95–5.65) ab | ND z |
| 80 | ND z | log10 5.63 (log105.35–6.05) bc | ND z |
| 90 | ND z | log10 6.63 (log106.35–7.05) c | ND z |
| 100 | ND z | log10 6.50 (log106.50–6.50) c | ND z |
| 95% CI | log10 5.36 (log10 4.01–6.70) | log10 6.08 (log10 5.73–6.43) | log10 4.79 (log10 3.27–6.32) |
| Coefficient of Variation | 8.07% | 23.98% | 12.83% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palomino-Tapia, V.A.; Zavala, G.; Cheng, S.; Garcia, M. Attenuation of a Field Strain of Infectious Laryngotracheitis Virus in Primary Chicken Culture Cells and Adaptation to Secondary Chicken Embryo Fibroblasts. Poultry 2023, 2, 516-530. https://doi.org/10.3390/poultry2040038
Palomino-Tapia VA, Zavala G, Cheng S, Garcia M. Attenuation of a Field Strain of Infectious Laryngotracheitis Virus in Primary Chicken Culture Cells and Adaptation to Secondary Chicken Embryo Fibroblasts. Poultry. 2023; 2(4):516-530. https://doi.org/10.3390/poultry2040038
Chicago/Turabian StylePalomino-Tapia, Victor A., Guillermo Zavala, Sunny Cheng, and Maricarmen Garcia. 2023. "Attenuation of a Field Strain of Infectious Laryngotracheitis Virus in Primary Chicken Culture Cells and Adaptation to Secondary Chicken Embryo Fibroblasts" Poultry 2, no. 4: 516-530. https://doi.org/10.3390/poultry2040038
APA StylePalomino-Tapia, V. A., Zavala, G., Cheng, S., & Garcia, M. (2023). Attenuation of a Field Strain of Infectious Laryngotracheitis Virus in Primary Chicken Culture Cells and Adaptation to Secondary Chicken Embryo Fibroblasts. Poultry, 2(4), 516-530. https://doi.org/10.3390/poultry2040038