

Effects of Varying Concentrations of Eimeria Challenge on the Intestinal Integrity of Broiler Chickens


 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Aspects
2.2. Challenge
2.3. Experimental Design
2.4. Oocyst Count (OPG) and Lesion Score
2.5. Real-Time PCR Analysis
2.6. Macroscopic Score
2.7. Histological Analysis
2.8. Histomorphometric Analysis
2.9. Intestinal Permeability
2.10. Mucosal Sample Analysis
2.11. Statistics
3. Results
3.1. Oocyst Count (OPG) and qPCR
3.2. Mucosa Parameters
3.3. Intestinal Permeability
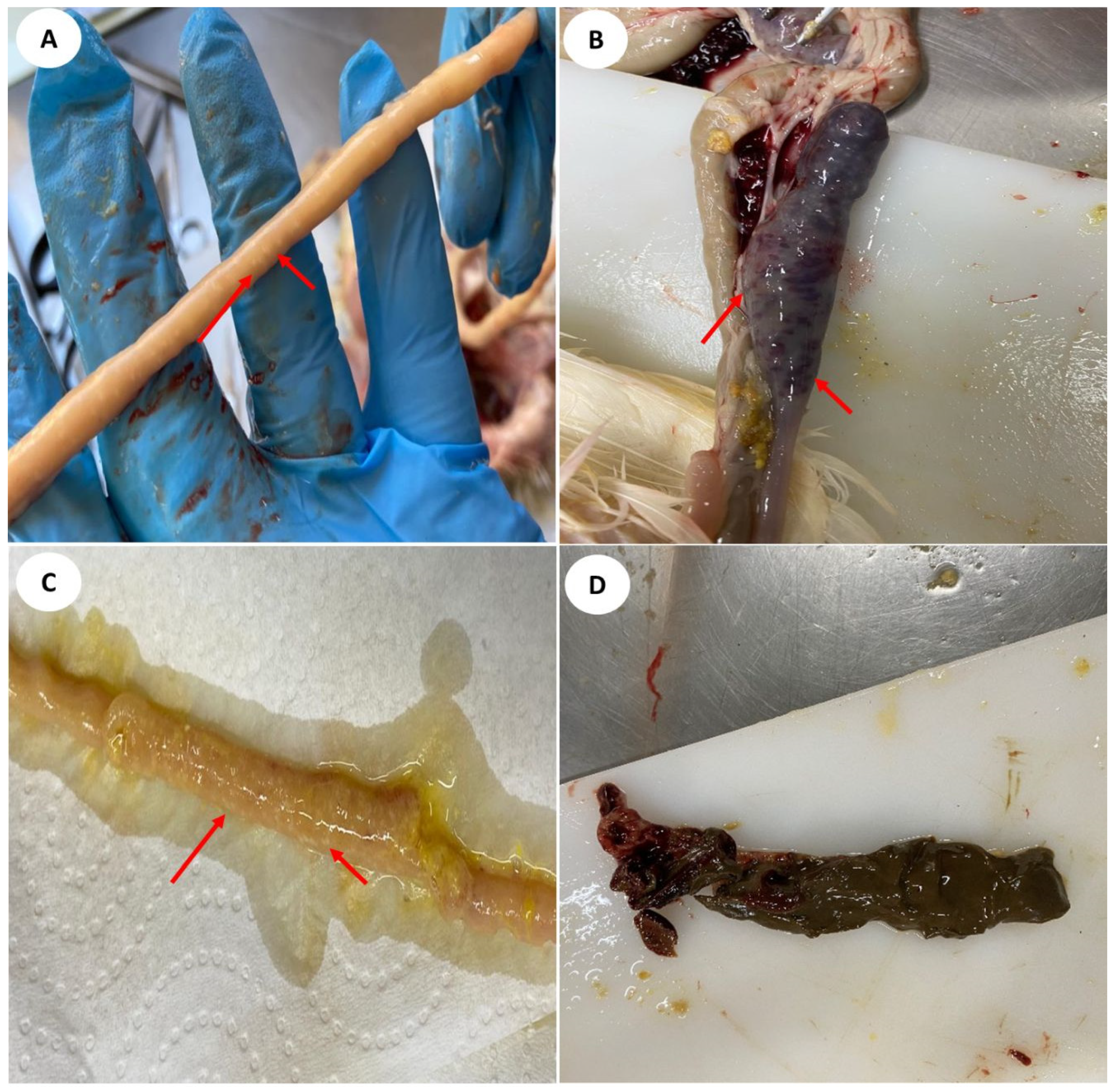
3.4. Assessment of Macroscopic and Histopathological Lesions
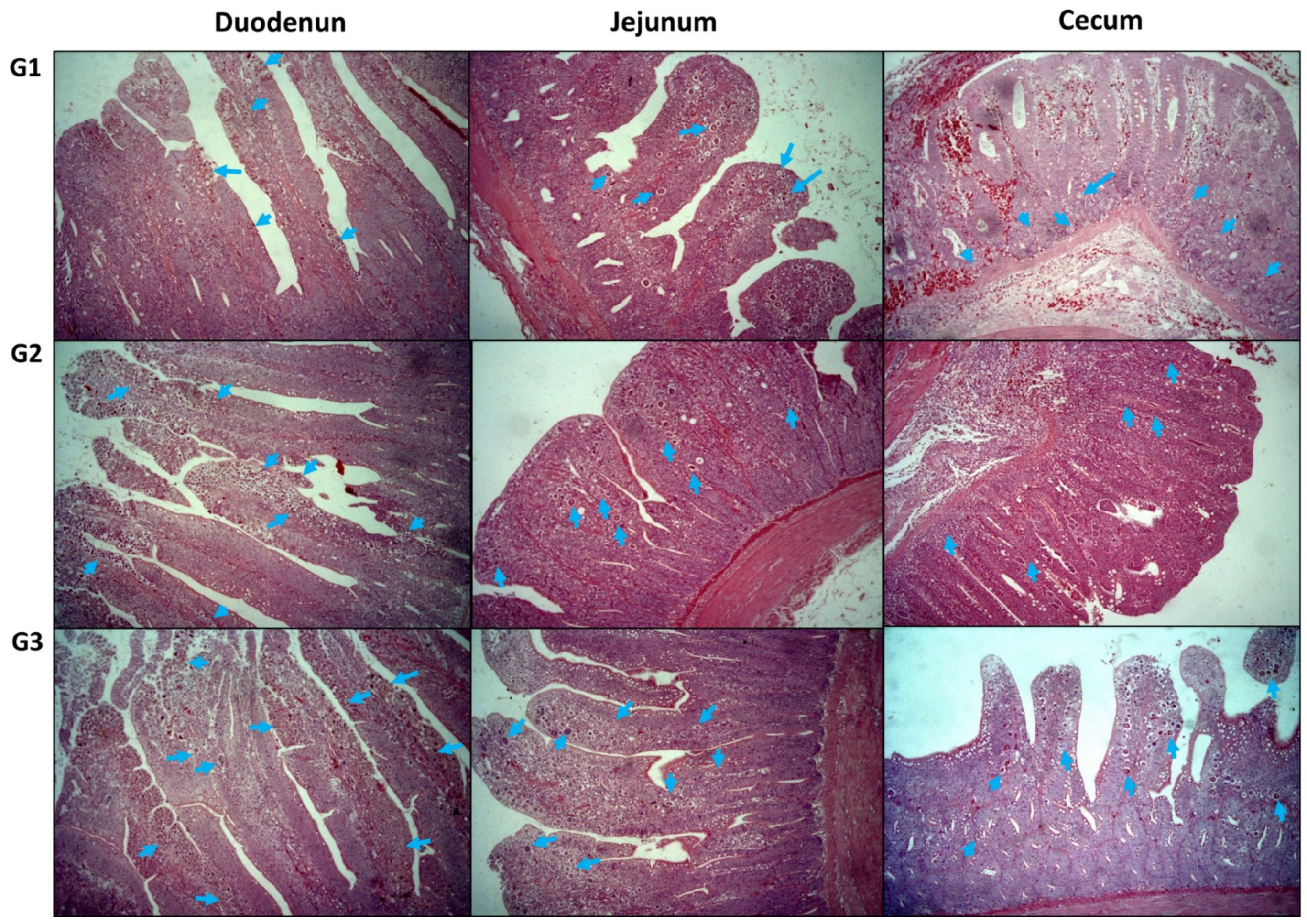
3.5. Histomorphometric Measurements of the Duodenum, Jejunum, and Cecum
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shivaramaiah, C.; Barta, J.R.; Hernandez-Velasco, X.; Tellez, G.; Hargis, B.M. Coccidiosis: Recent advancements in the immunobiology of Eimeria species, preventive measures, and the importance of vaccination as a control tool against these Apicomplexan parasites. Vet. Med. 2014, 5, 23–34. [Google Scholar] [CrossRef]
- Muller, J.; Hemphill, A. In vitro culture systems for the study of apicomplexan parasites in farm animals. Int. J. Parasitol. 2013, 43, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.H.; Lillehoj, H.S.; Lillehoj, E.P. Intestinal immune responses to coccidiosis. Dev. Comp. Immunol. 2000, 24, 303–324. [Google Scholar] [CrossRef] [PubMed]
- Collier, C.T.; Hofacre, C.L.; Payne, A.M.; Anderson, D.B.; Kaiser, P.; Mackie, R.I.; Gaskins, H.R. Coccidia-induced mucogenesis promotes the onset of necrotic enteritis by supporting Clostridium perfringens growth. Vet. Immunol. Immunopathol. 2008, 122, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Chapman, H.D.; Jeffers, T.K. Vaccination of chickens against coccidiosis ameliorates drug resistance in commercial poultry production. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 214–217. [Google Scholar] [CrossRef] [PubMed]
- Bortoluzzi, C.; Paras, K.L.; Applegate, T.J.; Verocai, G.G. Comparison between McMaster and Mini-FLOTAC methods for the enumeration of Eimeria maxima oocysts in poultry excreta. Vet. Parasitol. 2018, 254, 21–25. [Google Scholar] [CrossRef]
- Haug, A.; Gjevre, A.G.; Skjerve, E.; Kaldhusdal, M. A survey of the economic impact of subclinical Eimeria infections in broiler chickens in Norway. Avian Pathol. 2008, 37, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Adedokun, S.A.; Adeola, O. The response in jejunal and ileal nutrient and energy digestibility and the expression of markers of intestinal inflammation in broiler chickens to coccidial vaccine challenge and phytase supplementation. Can. J. Anim. Sci. 2017, 97, 258–267. [Google Scholar] [CrossRef]
- Parker, J.; Oviedo-Rondon, E.O.; Clack, B.A.; Clemente-Hernandez, S.; Osborne, J.; Remus, J.C.; Kettunen, H.; Makivuokko, H.; Pierson, E.M. Enzymes as feed additive to aid in responses against Eimeria species in coccidia-vaccinated broilers fed corn-soybean meal diets with different protein levels. Poult. Sci. 2007, 86, 643–653. [Google Scholar] [CrossRef]
- Rochell, S.J.; Parsons, C.M.; Dilger, R.N. Effects of Eimeria acervulina infection severity on growth performance, apparent ileal amino acid digestibility, and plasma concentrations of amino acids, carotenoids, and alpha 1-acid glycoprotein in broilers. Poult. Sci. 2016, 95, 1573–1581. [Google Scholar] [CrossRef]
- Teng, P.Y.; Yadav, S.; Castro, F.L.S.; Tompkins, Y.H.; Fuller, A.L.; Kim, W.K. Graded Eimeria challenge linearly regulated growth performance, dynamic change of gastrointestinal permeability, apparent ileal digestibility, intestinal morphology, and tight junctions of broiler chickens. Poult. Sci. 2020, 99, 4203–4216. [Google Scholar] [CrossRef]
- Haug, A.; Williams, R.B.; Larsen, S. Counting coccidial oocysts in chicken faeces: A comparative study of a standard McMaster technique and a new rapid method. Vet. Parasitol. 2006, 136, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Conway, D.P.; Mckenzie, M.E.; Dayton, A.D. Relationship of Coccidial Lesion Scores and Weight-Gain in Infections of Eimeria acervulina, E. maxima and E. tenella in Broilers. Avian Pathol. 1990, 19, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Baron, L.F.; Fonseca, F.N.D.; Maciag, S.S.; Bellaver, F.A.V.; Ibeli, A.M.G.; Mores, M.A.Z.; Almeida, G.F.D.; Guterres, S.S.; Bastos, A.P.A.; Paese, K. Toltrazuril-Loaded Polymeric Nanocapsules as a Promising Approach for the Preventive Control of Coccidiosis in Poultry. Pharmaceutics 2022, 14, 392. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.; Reid, W.M. Anticoccidial drugs: Lesion scoring techniques in battery and floor-pen experiments with chickens. Exp. Parasitol. 1970, 28, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, M.A.; Brown, J.; Bounous, D.I. Use of microscopic lesion scores, gross lesion scores and oocyst count scores to detect in chickens. Avian Pathol. 1998, 27, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Vicuña, E.A.; Kuttappan, V.A.; Tellez, G.; Hernandez-Velasco, X.; Seeber-Galarza, R.; Latorre, J.D.; Faulkner, O.B.; Wolfenden, A.D.; Hargis, B.M.; Bielke, L.R. Dose titration of FITC-D for optimal measurement of enteric inflammation in broiler chicks. Poult. Sci. 2015, 94, 1353–1359. [Google Scholar] [CrossRef]
- Baxter, M.F.A.; Merino-Guzman, R.; Latorre, J.D.; Mahaffey, B.D.; Yang, Y.C.; Teague, K.D.; Graham, L.E.; Wolfenden, A.D.; Hernandez-Velasco, X.; Bielke, L.R.; et al. Optimizing Fluorescein Isothiocyanate Dextran Measurement as a Biomarker in a 24-h Feed Restriction Model to Induce Gut Permeability in Broiler Chickens. Front. Vet. Sci. 2017, 4, 56. [Google Scholar] [CrossRef]
- Jenkins, M.C.; Dubey, J.P.; Miska, K.; Fetterer, R. Differences in fecundity of Eimeria maxima strains exhibiting different levels of pathogenicity in its avian host. Vet. Parasitol. 2017, 236, 1–6. [Google Scholar] [CrossRef]
- Williams, R.B. Quantification of the crowding effect during infections with the seven Eimeria species of the domesticated fowl: Its importance for experimental designs and the production of oocyst stocks. Int. J. Parasitol. 2001, 31, 1056–1069. [Google Scholar] [CrossRef]
- Teng, P.Y.; Choi, J.; Yadav, S.; Tompkins, Y.H.; Kim, W.K. Effects of low-crude protein diets supplemented with arginine, glutamine, threonine, and methionine on regulating nutrient absorption, intestinal health, and growth performance of Eimeria-infected chickens. Poult. Sci. 2021, 100, 101427. [Google Scholar] [CrossRef] [PubMed]
- Teng, P.Y.; Choi, J.; Tompkins, Y.; Lillehoj, H.; Kim, W. Impacts of increasing challenge with Eimeria maxima on the growth performance and gene expression of biomarkers associated with intestinal integrity and nutrient transporters. Vet. Res. 2021, 52, 81. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, M.; Allen, P.; Wilkins, G.; Klopp, S.; Miska, K. Eimeria praecox infection ameliorates effects of Eimeria maxima infection in chickens. Vet. Parasitol. 2008, 155, 10–14. [Google Scholar] [CrossRef]
- Conway, D.P.; Sasai, K.; Gaafar, S.M.; Smothers, C.D. Effects of Different Levels of Oocyst Inocula of Eimeria acervulina, E. tenella, and E. maxima on Plasma Constituents, Packed Cell-Volume, Lesion Scores, and Performance in Chickens. Avian Dis. 1993, 37, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Velasco, X.; Chapman, H.D.; Owens, C.M.; Kuttappan, V.A.; Fuente-Martínez, B.; Menconi, A.; Latorre, J.D.; Kallapura, G.; Bielke, L.R.; Rathinam, T.; et al. Absorption and deposition of xanthophylls in broilers challenged with three dosages of oocysts. Brit Poult. Sci. 2014, 55, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.; Vahl, H.A.; Veldman, A. Interaction between nutrition and Eimeria acervulina infection in broiler chickens: Development of an experimental infection model. Brit J. Nutr. 1996, 75, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Assis, R.C.L.; Luns, F.D.; Beletti, M.E.; Assis, R.L.; Nasser, N.M.; Faria, E.S.M.; Cury, M.C. Histomorphometry and macroscopic intestinal lesions in broilers infected with Eimeria acervulina. Vet. Parasitol. 2010, 168, 185–189. [Google Scholar] [CrossRef]
- Sugiharto, S.; Yudiarti, T.; Isroli, I.; Widiastuti, E.; Kusumanti, E. Dietary Supplementation of Probiotics in Poultry Exposed to Heat Stress—A Review. Ann. Anim. Sci. 2017, 17, 591–604. [Google Scholar] [CrossRef]
- Kaetzel, C.S. The polymeric immunoglobulin receptor: Bridging innate and adaptive immune responses at mucosal surfaces. Immunol. Rev. 2005, 206, 83–99. [Google Scholar] [CrossRef]
- Jeurissen, S.H.M.; Boonstra-Blom, A.G.; Al-Garib, S.O.; Hartog, L.; Koch, G. Defence mechanisms against viral infection in poultry: A review. Vet. Quart. 2000, 22, 204–208. [Google Scholar] [CrossRef]
- Mcdougald, L.R. Intestinal protozoa important to poultry. Poult. Sci. 1998, 77, 1156–1158. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Jenkins, M.C. Re-evaluation of the life cycle of Eimeria maxima Tyzzer, 1929 in chickens (Gallus domesticus). Parasitology 2018, 145, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Bedrnik, P.; Jurkovic, P. An antigen from Eimeria tenella merozoites for the indirect fluorescence antibody test. Folia Parasitol. 1979, 26, 169–172. [Google Scholar]
- Bedrnik, P.; Jurkovic, P. Comparison of 2 Different Antigens of Eimeria tenella for Indirect Immunofluorescent Test (Ifat). J. Protozool. 1977, 24, A48–A49. [Google Scholar]
- Augustine, P.C. Cell: Sporozoite interactions and invasion by apicomplexan parasites of the genus Eimeria. Int. J. Parasitol. 2001, 31, 1–8. [Google Scholar] [CrossRef]
- Shirley, M.W.; Smith, A.L.; Tomley, F.M. The biology of avian with an emphasis on their control by vaccination. Adv. Parasit. 2005, 60, 285–330. [Google Scholar] [CrossRef]
- Belote, B.L.; Soares, I.; Tujimoto-Silva, A.; Sanches, A.W.D.; Kraieski, A.L.; Santin, E. Applying I see inside histological methodology to evaluate gut health in broilers challenged with Eimeria. Vet. Parasitol. 2019, 276S, 100004. [Google Scholar] [CrossRef]
- Klasing, K.C. Nutrition and the immune system. Br. Poult. Sci. 2007, 48, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Kogut, M.H.; Klasing, K. An immunologist’s perspective on nutrition, immunity, and infectious diseases: Introduction and overview. J. Appl. Poult. Res. 2009, 18, 103–110. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatment | E. acervulina (No. Oocysts) | E. maxima (No. Oocysts) | E. tenella (No. Oocysts) |
|---|---|---|---|
| Unchallenged (U) | zero | zero | zero |
| Medium-low challenge (G1) | 31,250 | 20,000 | 6250 |
| Medium-high challenge (G2) | 62,500 | 40,000 | 12,500 |
| High challenge (G3) | 125,000 | 80,000 | 25,000 |
| Lesion Score | Macroscopic Finding Observed |
|---|---|
| 0 | No lesion |
| 1 | Wall of the small intestine roughly thinner, easily torn under slight tension, but no rough evidence of necrosis of the mucosa or other abnormalities |
| 2 | One or more necrotic focuses in areas of the mucosa measuring approximately 1 to 5 mm in total thickness |
| 3 | Necrosis greater than 5 mm and consisting of orange/brown necrotic detritus |
| 4 | Large confluent necrotic areas over the total thickness of the mucosa in the small intestine, affecting 25% or more of the small intestine |
| Oocysts g−1 | qPCR Status | Mean Amount in qPCR | ||
|---|---|---|---|---|
| Negative control (U) | Before the challenge | 0 | undetectable | 0 |
| 1 dpi | 0 | undetectable | 0 | |
| 3 dpi | 0 | undetectable | 0 | |
| 5 dpi | 0 | undetectable | 0 | |
| Medium-low challenge (G1) | Before the challenge | 0 | undetectable | 0 |
| 1 dpi | 2.0 × 103 | detectable | 3167.545 | |
| 3 dpi | 2.9 × 104 | detectable | 25,569.347 | |
| 5 dpi | 5.7 × 105 | detectable | 250,501.568 | |
| Medium-high challenge (G2) | Before the challenge | 0 | undetectable | 0 |
| 1 dpi | 3.0 × 103 | detectable | 8665.856 | |
| 3 dpi | 3.6 × 104 | detectable | 31,751.037 | |
| 5 dpi | 7.4 × 105 | detectable | 250,059.875 | |
| High challenge (G3) | Before the challenge | 0 | undetectable | 0 |
| 1 dpi | 9.6 × 104 | detectable | 41,723.154 | |
| 3 dpi | 6.7 × 103 | detectable | 8375.960 | |
| 5 dpi | 3.4 × 106 | detectable | 417,284.204 |
| Group | ||||
|---|---|---|---|---|
| Variable | U | G1 | G2 | G3 |
| pIgR (ng/mg protein) | 12.04 ± 1.20 a | 23.13 ± 1.23 b | 39.45 ± 1.98 c | 36.42 ± 2.31 c |
| IgA (ng/mg protein) | 9.72 ± 0.69 a | 20.49 ± 0.65 b | 18.66 ± 0.64 b | 18.37 ± 0.84 b |
| Group | Unchallenged (U) | Medium-Low Challenge (G1) | Medium-High Challenge (G2) | High Challenge (G3) | Pr > χ2 |
|---|---|---|---|---|---|
| Macroscopic Score | |||||
| Duodenum | 0 ± 0 a | 3.37 ± 0.52 b | 3.25 ± 0.71 b | 3.13 ± 0.64 b | 0.0017 |
| Jejunum | 0 ± 0 a | 1.00 ± 0.75 b | 1.25 ± 0.25 b | 1.37 ± 0.38 b | 0.0091 |
| Cecum | 0 ± 0 a | 2.00 ± 0.93 b | 3.12 ± 0.99 b.c | 3.63 ± 0.52 c | 0.0372 |
| Microscopic Score | |||||
| Duodenum | 0 ± 0 a | 6.13 ± 2.85 b | 7.38 ± 0.52 b | 7.00 ± 1.19 b | 0.0032 |
| Jejunum | 0 ± 0 a | 6.87 ± 2.80 b | 8.00 ± 0.00 b | 6.75 ± 1.40 b | 0.0020 |
| Cecum | 0 ± 0 a | 3.75 ± 2.24 b | 5.12 ± 2.47 b | 6.25 ± 1.16 b | 0.0082 |
| Group | ||||
|---|---|---|---|---|
| Variable | U | G1 | G2 | G3 |
| Duodenum | ||||
| VH, µm | 852 ± 88.64 a | 586.5 ± 178.07 b | 576.9 ± 214.64 b | 636.6 ± 207.68 b |
| CD, µm | 145.4 ± 8.52 | 168.2 ± 21.36 | 173.2 ± 67.72 | 157.5 ± 20.20 |
| MT, µm | 124.3 ± 30.68 a | 211.2 ± 43.73 b | 252.2 ± 66.30 b | 274.5 ± 65.71 b |
| V/C | 5.8 ± 0.69 a | 3.5 ± 0.88 b | 3.5 ± 1.36 b | 4.1 ± 1.36 b |
| GC | 126.8 ± 26.66 a | 56.29 ± 8.48 b | 34.13 ± 9.48 c | 34.89 ± 8.95 c |
| Jejunum | ||||
| VH, µm | 739.7 ± 39.78 a | 389.9 ± 164.42 b | 440.1 ± 124.42 b | 429.4 ± 140.44 b |
| CD, µm | 124.1 ± 11.49 a | 163.4 ± 36.43 b | 152.7 ± 49.42 b | 146.1 ± 22.82 a.b |
| MT, µm | 110.1 ± 10.33 a | 227.8 ± 89.52 b | 198.3 ± 40.52 b | 235.2 ± 21.31 b |
| V/C | 6.0 ± 0.65 a | 2.3 ± 0.64 b | 2.9 ± 1.00 b | 3.1 ± 1.43 b |
| GC | 135.87 ± 22.00 a | 45.94 ± 12.00 b | 31.05 ± 7.50 b | 30.2 ± 4.30 b |
| Cecum | ||||
| VH, µm | 583.5 ± 52.58 a | 256.5 ± 125.70 b | 183.0 ± 42.78 b | 182.0 ± 43.39 b |
| CD, µm | 109.8 ± 9.33 a | 145.4 ± 23.04 b | 137.8 ± 17.88 b | 136.0 ± 20.62 b |
| MT, µm | 101.8 ± 18.81 | 131.3 ± 46.95 | 122.2 ± 23.04 | 102.2 ± 19.99 |
| V/C | 5.3 ± 0.63 a | 1.7 ± 0.70 b | 1.3 ± 0.31 b | 1.3 ± 0.32 b |
| GC | 124.5 ± 13.86 a | 10.3 ± 3.50 b | 6.5 ± 2.76 c | 7.0 ± 2.86 b.c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza, G.C.d.; Esteves, G.F.; Volpato, F.A.; Miotto, R.; Mores, M.A.Z.; Ibelli, A.M.G.; Bastos, A.P. Effects of Varying Concentrations of Eimeria Challenge on the Intestinal Integrity of Broiler Chickens. Poultry 2024, 3, 1-14. https://doi.org/10.3390/poultry3010001
Souza GCd, Esteves GF, Volpato FA, Miotto R, Mores MAZ, Ibelli AMG, Bastos AP. Effects of Varying Concentrations of Eimeria Challenge on the Intestinal Integrity of Broiler Chickens. Poultry. 2024; 3(1):1-14. https://doi.org/10.3390/poultry3010001
Chicago/Turabian StyleSouza, Giovana Camargo de, Giovanna Fernandes Esteves, Franciana Aparecida Volpato, Rovian Miotto, Marcos Antônio Zanella Mores, Adriana Mércia Guaratini Ibelli, and Ana Paula Bastos. 2024. "Effects of Varying Concentrations of Eimeria Challenge on the Intestinal Integrity of Broiler Chickens" Poultry 3, no. 1: 1-14. https://doi.org/10.3390/poultry3010001
APA StyleSouza, G. C. d., Esteves, G. F., Volpato, F. A., Miotto, R., Mores, M. A. Z., Ibelli, A. M. G., & Bastos, A. P. (2024). Effects of Varying Concentrations of Eimeria Challenge on the Intestinal Integrity of Broiler Chickens. Poultry, 3(1), 1-14. https://doi.org/10.3390/poultry3010001