
QTL Identification of Hull Color for Foxtail Millet [Setaria italica (L.) P. Beauv.] Through Four Phenotype Identification Strategies in a RIL Population

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Phenotypic Determination Method for Hull Color
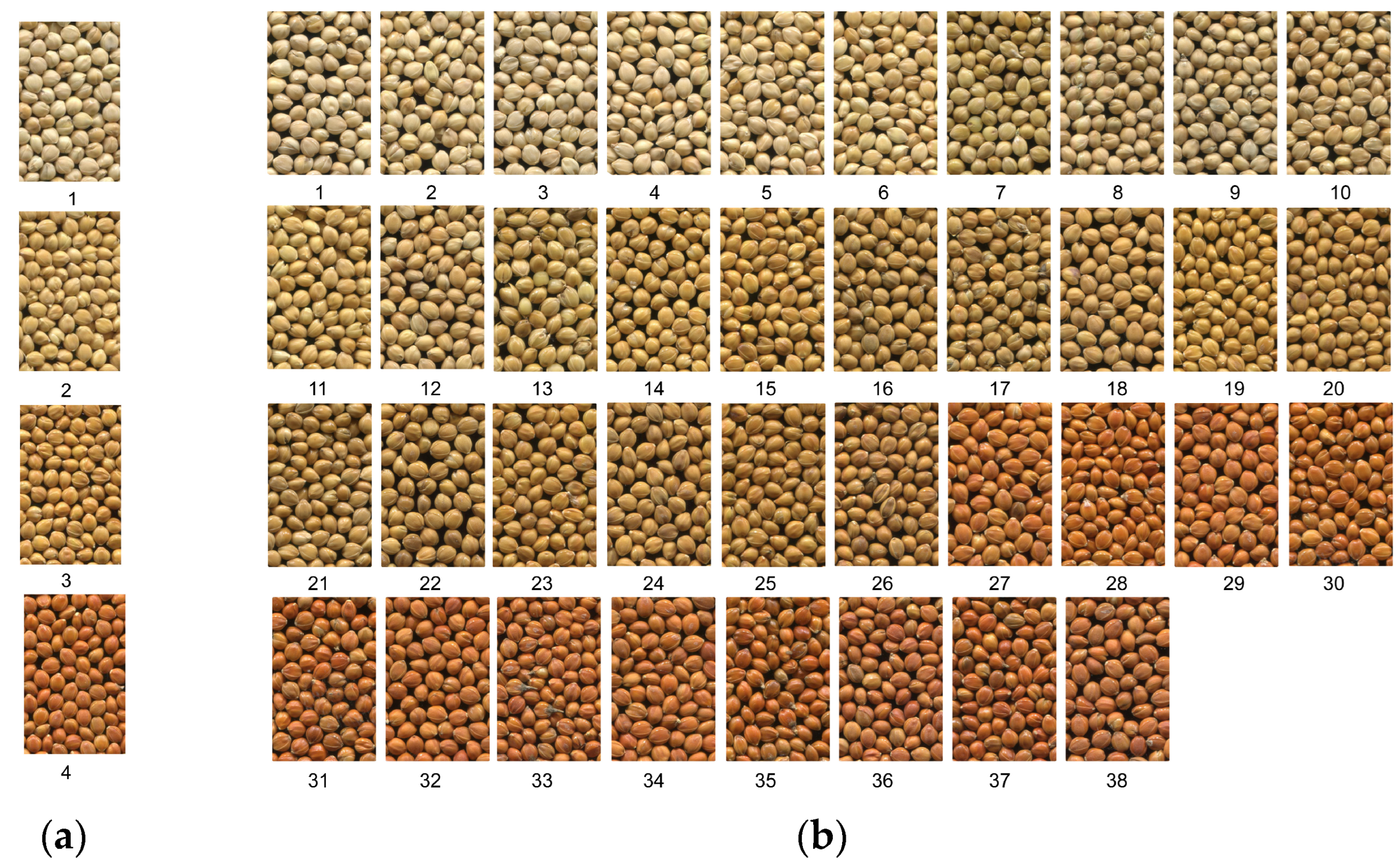
2.2.1. Method I: Visual Grouping Method
2.2.2. Method II: Visual Colorimetric Method
2.2.3. Method III: Lab Determination Method
2.2.4. Method IV: RGB Determination Method
2.2.5. QTL Mapping
2.3. Statistics Analysis
3. Results
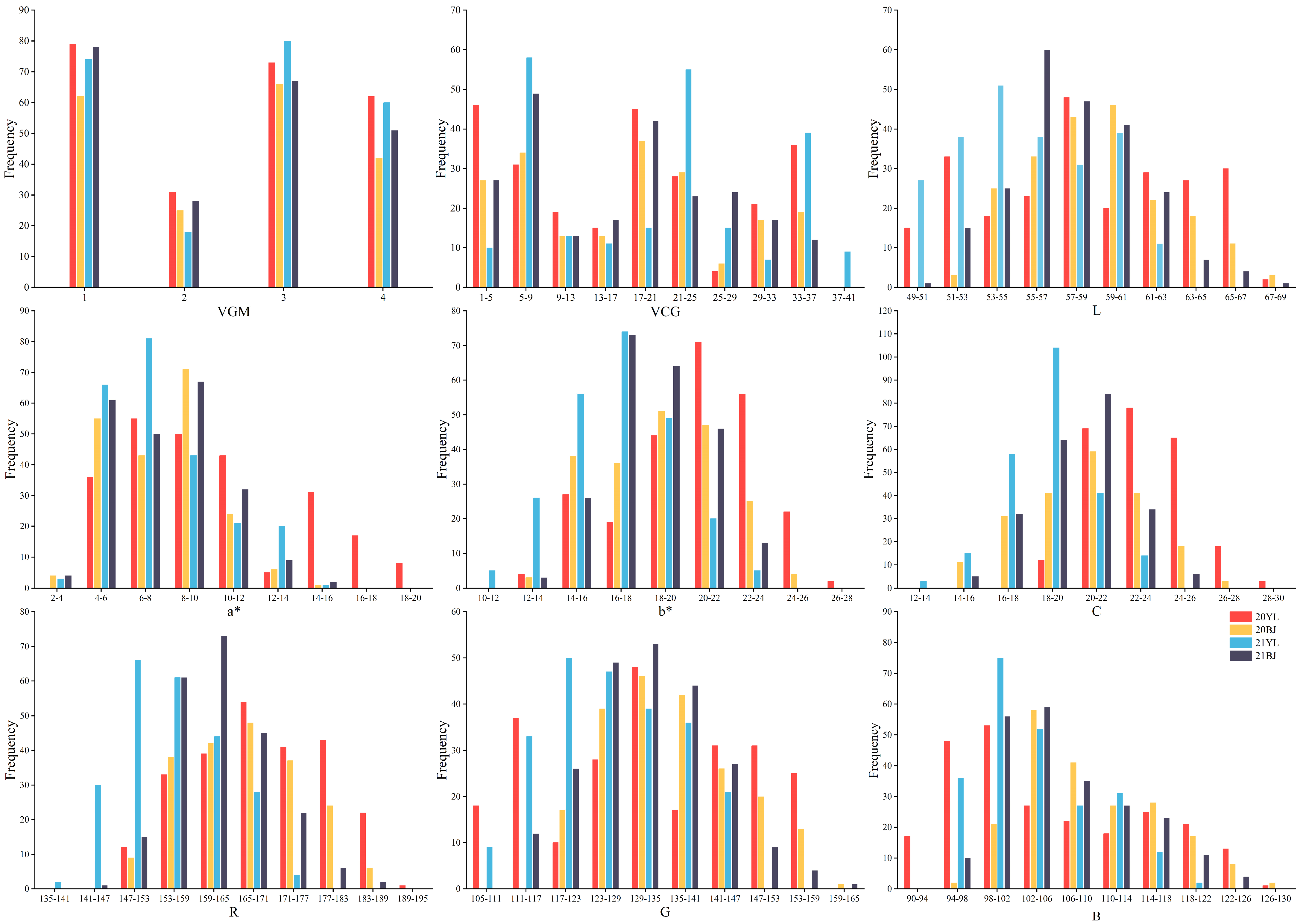
3.1. Phenotypic Variation of Hull Color in RYRIL Population by Four Methods
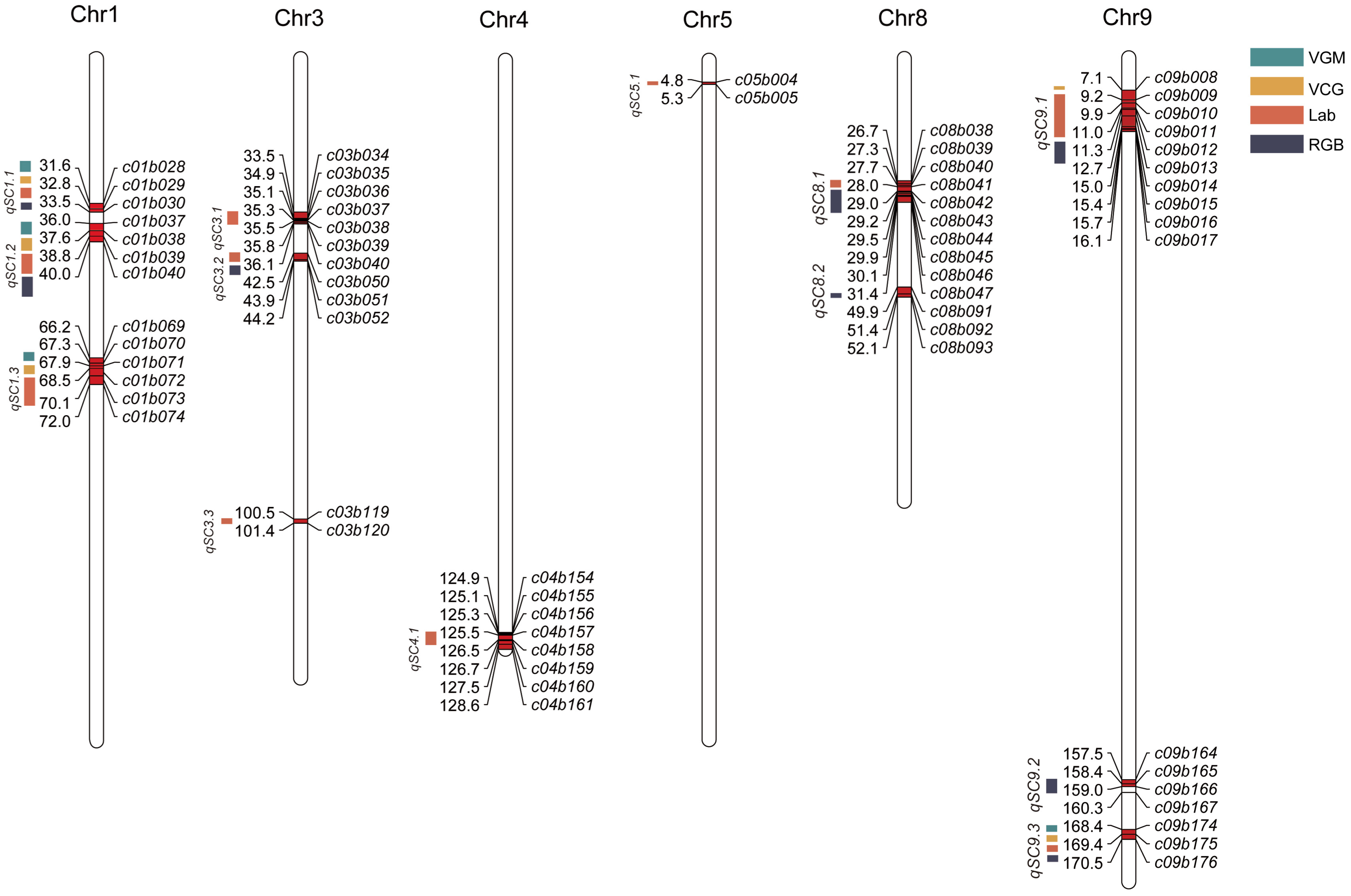
3.2. Identification of QTL Associating with Hull Color in Foxtail Millet
3.3. QTL Analysis with Epistatic Interactions for the Foxtail Millet Hull Color Under Different Detection Methods
3.4. Comparison Analysis of Four Hull Color Detection Methods
4. Discussion
4.1. Foxtail Millet Exhibits a Rich Variation in Grain Hull Color
4.2. Comparison of the Four Phenotype Characterisation Methods
4.3. The Main Effect of QTL with a Stable Environment Is Generally Consistent with Previous Studies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lu, H.; Zhang, J.; Liu, K.; Wu, N.; Li, Y.; Zhou, K.; Ye, M.; Zhang, T.; Zhang, H.; Yang, X.; et al. Earliest domestication of common millet (Panicum miliaceum) in east asia extended to 10,000 years ago. Proc. Natl. Acad. Sci. USA 2009, 106, 7367–7372. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wan, Z.; Perry, L.; Lu, H.; Wang, Q.; Zhao, C.; Li, J.; Xie, F.; Yu, J.; Cui, T.; et al. Early millet use in northern China. Proc. Natl. Acad. Sci. USA 2012, 109, 3726–3730. [Google Scholar] [CrossRef] [PubMed]
- Sachdev, N.; Goomer, S.; Singh, L.R. Foxtail millet: A potential crop to meet future demand scenario for alternative sustainable protein. J. Sci. Food Agric. 2021, 101, 831–842. [Google Scholar] [CrossRef]
- Liang, S.Z. Arabidopsis thaliana Recombinant SERKs and Setaria italica BAK1 Involved in the Regulation of Brassinosteroid Signaling and Cell Death Pathway. Master’s Thesis, Northwest Normal University, Lanzhou, China, 2019. [Google Scholar]
- Gao, Y.; Yuan, Y.; Zhang, X.; Song, H.; Yang, Q.; Yang, P.; Gao, X.; Gao, J.; Feng, B. Conuping BSA-seq and RNA-seq reveal the molecular pathway and genes associated with the plant height of foxtail millet (Setaria Italica). Int. J. Mol. Sci. 2022, 23, 11824. [Google Scholar] [CrossRef]
- Liu, D.; Cui, Y.; He, J.; Li, S.; Li, Q.; Liang, D.; Wang, J.; Shi, X.; Wang, C.; Dong, K.; et al. Genetic diversity and classification of the cytoplasm of chinese elite foxtail millet [Setaria Italica (L.) P. Beauv.] parental lines revealed by chloroplast deoxyribonucleic acid variation. Front. Genet. 2019, 10, 1198. [Google Scholar] [CrossRef]
- Yang, Y.; Jia, G.; Deng, L.; Qin, L.; Chen, E.; Cong, X.; Zou, R.; Wang, H.; Zhang, H.; Liu, B.; et al. Genetic variation of yellow pigment and its components in foxtail millet (Setaria Italica (L.) P. Beauv.) from different eco-regions in China. J. Integr. Agric. 2017, 16, 2459–2469. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, Y.; Zhang, B.; He, L.; Liu, L.; Wang, X.; Li, H.; Han, Y. The relationship between SiLCYB gene and beige formation in the biosynthesis pathway of carotenoids in foxtail millet. Mol. Plant Breed. 2016, 14, 1341–1351. [Google Scholar]
- He, J.H.; Yang, T.Y.; Wu, G.Z. Evaluation on nutritive quality of local varieties for foxtail millet in Gansu Province. Plant Genet. Resour. Sci. 2002, 3, 41–44. [Google Scholar]
- Xia, X.Y.; Shi, Z.G.; Zhang, T.; Chen, Y.; Xiang, J.Y.; Ji, X.M.; Cheng, R.H. The research on bird-disaster degree in different grain colors of foxtail millet lines. J. Hebei Agric. Sci. 2014, 18, 4–6. [Google Scholar]
- Winkel-Shirley, B. Flavonoid biosynthesis a colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar]
- Zhang, S.; Hu, X.; Miao, H.; Chu, Y.; Cui, F.; Yang, W.; Xu, S.; Guo, J.; Fu, C.; Song, X.; et al. Imaged-based phenotyping accelerated QTL mapping and QTL x environment interaction analysis of testa colour in peanut (Arachis hypogaea). Plant Breed. 2021, 140, 884–895. [Google Scholar] [CrossRef]
- Liu, F.; Chen, H.; Yang, L.; You, L.; Ju, J.; Yang, S.; Wang, X.; Liu, Z. QTL mapping and transcriptome analysis reveal candidate genes regulating seed color in brassica napus. Int. J. Mol. Sci. 2023, 24, 9262. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.M. Genetic Mechanism and Gene Regulation Network of Flavonoid Pigmentation in Rice (Oryza sativa L.). Ph.D. Thesis, China Agricultural University, Beijing, China, 2017. [Google Scholar]
- Emiliani, J.; Grotewold, E.; Falcone Ferreyra, M.L.; Casati, P. Flavonols protect Arabidopsis plants against UV-B deleterious effects. Mol. Plant 2013, 6, 1376–1379. [Google Scholar] [CrossRef]
- Nakabayashi, R.; Mori, T.; Saito, K. Alternation of flavonoid accumulation under drought stress in Arabidopsis thaliana. Plant Signal. Behav. 2014, 9, e29518. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, R.; Yonekura-Sakakibara, K.; Urano, K.; Suzuki, M.; Yamada, Y.; Nishizawa, T.; Matsuda, F.; Kojima, M.; Sakakibara, H.; Shinozaki, K.; et al. Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J. 2014, 77, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, T.; Maekawa, M.; Oki, T.; Suda, I.; Iida, S.; Shimada, H.; Takamure, I.; Kadowaki, K. The Rc and Rd genes are involved in proanthocyanidin synthesis in rice pericarp. Plant J. 2007, 49, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gao, J.; Qie, Q.; Yang, Y.; Hou, S.; Wang, X.; Li, X.; Han, Y. Comparative analysis of flavonoid metabolites in foxtail millet (Setaria italica) with different eating quality. Life 2021, 11, 578. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.D. High-Density Genetic Linkage Map Construction and QTL Mapping of Seed Coat Color in Sesame (Sesamum indicum L.). Masters’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2016. [Google Scholar]
- Kuang, Q.; Yu, Y.; Attree, R.; Xu, B. A comparative study on anthocyanin, saponin, and oil profiles of black and red seed coat peanut (Arachis hypogacea) grown in China. Int. J. Food Prop. 2017, 20, 131–140. [Google Scholar] [CrossRef]
- Wan, L.; Li, B.; Pandey, M.K.; Wu, Y.; Lei, Y.; Yan, L.; Dai, X.; Jiang, H.; Zhang, J.; Wei, G.; et al. Transcriptome analysis of a new peanut seed coat mutant for the physiological regulatory mechanism involved in seed coat cracking and pigmentation. Front. Plant Sci. 2016, 7, 1491. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Liu, J.; Cheng, L.; Zhang, Y.; Hou, S.; Sun, Z.; Li, H.; Han, Y. Carotenoid composition and expression of biosynthetic genes in yellow and white foxtail millet [Setaria italica (L.) Beauv]. J. Cereal Sci. 2019, 85, 84–90. [Google Scholar] [CrossRef]
- Xie, H.; Hou, J.; Fu, N.; Wei, M.; Li, Y.; Yu, K.; Song, H.; Li, S.; Liu, J. Identification of qtl related to anther color and hull color by rad sequencing in a ril population of Setaria italica. BMC Genom. 2021, 22, 556. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.M.; Wang, C.Y.; Wang, P.; Zhu, Z.X.; Lu, X.C. QTL analysis of tannin content and color of grain in sorghum. J. Plant Genet. Resour. 2017, 8, 860–866. [Google Scholar]
- Yang, Y.; Zhao, T.; Wang, F.; Liu, L.; Liu, B.; Zhang, K.; Qin, J.; Yang, C.; Qiao, Y. Identification of candidate genes for soybean seed coat-related traits using QTL mapping and GWAS. Front. Plant Sci. 2023, 14, 1190503. [Google Scholar] [CrossRef]
- Ding, P.P. Genetic Analysis and QTL Mapping of Peel Color in Pepper Fruit. Master’s Thesis, Anhui Agricultural University, Anhui, China, 2017. [Google Scholar]
- Yu, B.; Boyle, K.; Zhang, W.; Robinson, S.J.; Higgins, E.; Ehman, L.; Relf-Eckstein, J.-A.; Rakow, G.; Parkin, I.A.P.; Sharpe, A.G.; et al. Multi-trait and multi-environment QTL analysis reveals the impact of seed colour on seed composition traits in Brassica napus. Mol. Breed. 2016, 36, 111. [Google Scholar] [CrossRef]
- Cichy, K.A.; Fernandez, A.; Kilian, A.; Kelly, J.D.; Galeano, C.H.; Shaw, S.; Brick, M.; Hodkinson, D.; Troxtell, E. QTL analysis of canning quality and color retention in black beans (phaseolus vulgaris L.). Mol. Breed. 2014, 33, 139–154. [Google Scholar] [CrossRef]
- Sooriyapathirana, S.S.; Khan, A.; Sebolt, A.M.; Wang, D.; Bushakra, J.M.; Lin-Wang, K.; Allan, A.C.; Gardiner, S.E.; Chagné, D.; Iezzoni, A.F. QTL analysis and candidate gene mapping for skin and flesh color in sweet cherry fruit (Prunus avium L.). Tree Genet. Genomes 2010, 6, 821–832. [Google Scholar] [CrossRef]
- Guo, S.; Chai, S.; Guo, Y.; Shi, X.; Han, F.; Qu, T.; Xing, L.; Yang, Q.; Gao, J.; Gao, X.; et al. Mapping of major qtl and candidate gene analysis for hull colour in foxtail millet (Setaria italica (L.) P. Beauv.). BMC Genom. 2023, 24, 458. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Fang, Z.Y.; Li, X.X.; Li, Q.H.; Cheng, J.Q.; Song, J.P.; Wang, H.P.; Qiu, Y. Inheritance of fruit flesh color in Cucumis sativus (L.). J. Plant Genet. Resour. 2011, 12, 216–222. [Google Scholar]
- Zapotoczny, P.; Majewska, K. A comparative analysis of colour measurements of the seed coat and endosperm of wheat kernels performed by various techniques. Int. J. Food Prop. 2010, 13, 75–89. [Google Scholar] [CrossRef]
- Lv, L.; Jin, L.M.; Lee, M.; Zhang, A.P.; Zhao, G.W. QTL mapping of seed color-related traits in yellow maize. Seed 2019, 38, 37–40+45. [Google Scholar]
- Ahmad, I.S.; Reid, J.F.; Paulsen, M.R.; Sinclair, J.B. Color classifier for symptomatic soybean seeds using image processing. Plant Dis. 1999, 83, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.M.; Devos, K.M.; Liu, C.J.; Wang, R.Q.; Gale, M.D. Construction of RFLP-based maps of foxtail millet, Setaria Italica (L.) P. Beauv. Theor. Appl. Genet. 1998, 96, 31–36. [Google Scholar] [CrossRef]
- Yazdizadeh, M.; Fahmideh, L.; Mohammadi-Nejad, G.; Solouki, M.; Nakhoda, B. Association analysis between agronomic traits and AFLP markers in a wide germplasm of proso millet (Panicum miliaceum L.) under normal and salinity stress conditions. BMC Plant Biol. 2020, 20, 427. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Zhang, L.; Hu, J.; Liu, Y.; Zhou, L.; Ping, W.; Zou, J.; Li, H. Genetic characterization of hull color using BSR-seq and genome re-sequencing approaches in foxtail millet. Front. Plant Sci. 2022, 13, 1019496. [Google Scholar] [CrossRef]
- Jia, X.; Zhang, Z.; Liu, Y.; Zhang, C.; Shi, Y.; Song, Y.; Wang, T.; Li, Y. Development and genetic mapping of SSR markers in foxtail millet [Setaria Italica (L.) P. Beauv.]. Theor. Appl. Genet. 2009, 118, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.Q.; Gao, J.H.; Guan, Z.B.; Mao, L.P. Preliminary study on chromosomal location and linkage relationship of several agronomic trait genes in foxtail millet. Acta Agron. Sin. 2007, 01, 9–14. [Google Scholar]
- Jia, G.; Huang, X.; Zhi, H.; Zhao, Y.; Zhao, Q.; Li, W.; Chai, Y.; Yang, L.; Liu, K.; Lu, H.; et al. A haplotype map of genomic variations and genome-wide association studies of agronomic traits in foxtail millet (Setaria italica). Nat. Genet. 2013, 45, 957–961. [Google Scholar] [CrossRef]
- Wang, J.; Yang, H.; Du, G.; Wang, Z.; Zou, H.; Du, X.; Li, Y.; Peng, J.; Guo, E.; Yong, J.; et al. Mapping of sihc1, which controls hull color, using a high-density genetic map based on restriction site-associated DNA sequencing in foxtail millet [Setaria italica (L.) P. Beauv.]. Mol. Breed. 2017, 37, 128. [Google Scholar] [CrossRef]
- Liu, G.H.; Yu, J.X.; Ren, L.J. Progress of RGB detection methods. J. Anal. Sci. 2020, 36, 591–596. [Google Scholar]
- Wang, C.; Chai, S.; Li, S.; Liu, D.; Han, H.; Wu, Y.; Li, Y.; Ma, Z.; Zhang, L.; Gao, X.; et al. Genetic dissection of foxtail millet bristles using combined QTL mapping and RNA-seq. Theor. Appl. Genet. 2025, 138, 33. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL ICI mapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 2015, 3, 269–283. [Google Scholar] [CrossRef]
- Nyquist, W.E.; Baker, R.J. Estimation of heritability and prediction of selection response in plant populations. Crit. Rev. Plant Sci. 1991, 10, 235–322. [Google Scholar] [CrossRef]
- Zhang, H.; Miao, H.; Wei, L.; Li, C.; Zhao, R.; Wang, C. Genetic analysis and QTL mapping of seed coat color in Sesame (Sesamum Indicum L.). PLoS ONE 2013, 8, e63898. [Google Scholar] [CrossRef] [PubMed]
- Vadivoo, A.S.; Joseph, R.; Ganesan, N.M. Genetic variability and diversity for protein and calcium contents in finger millet (Eleusine coracana (L.) Gaertn) in relation to grain color. Plant Foods Hum. Nutr. 1998, 52, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.Q. Genetic Analysis and QTL Mapping on Agriculturally Important Traits in Foxtail Millet. Master’s Thesis, Northwest A&F University, Xianyang, China, 2022. [Google Scholar]
- Alam, A.; Abdullah, M.; Mishra, R.S. Colour contrast enhancement method by scaling the dc coefficients in CIE-LAB colour space. Int. J. Comput. Appl. 2014, 97, 1–6. [Google Scholar] [CrossRef]
- Philipp, I.; Rath, T. Improving plant discrimination in image processing by use of different color space transformations. Comput. Electron. Agric. 2002, 35, 1–15. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Method | Trait | Environment | Parents | RIL Population | |||||
|---|---|---|---|---|---|---|---|---|---|
| Yugu 18 | Hongjiugu | Means ± SD | Range | Skewness | Kurtosis | Variable Coefficient (%) | |||
| I | VGM | 20YL | 1.00 | 4.00 | 2.48 ± 1.19 | 1.00–4.00 | −0.08 | −1.52 | 47.81 |
| 20BJ | 1.00 | 4.00 | 2.45 ± 1.15 | 1.00–4.00 | −0.08 | −1.46 | 46.92 | ||
| 21YL | 1.00 | 4.00 | 2.54 ± 1.19 | 1.00–4.00 | −0.21 | −1.49 | 46.70 | ||
| 21BJ | 1.00 | 4.00 | 2.40 ± 1.18 | 1.00–4.00 | 0.01 | −1.53 | 49.19 | ||
| II | VCM | 20YL | 6.00 | 32.00 | 17.91 ± 10.93 | 1.00–37.00 | 0.20 | −1.21 | 61.02 |
| 20BJ | 6.00 | 32.00 | 17.70 ± 10.04 | 1.00–37.00 | 0.20 | −1.09 | 56.69 | ||
| 21YL | 6.00 | 32.00 | 20.27 ± 10.97 | 1.00–38.00 | 0.04 | −1.29 | 54.10 | ||
| 21BJ | 6.00 | 32.00 | 17.01 ± 9.72 | 1.00–37.00 | 0.22 | −1.13 | 57.14 | ||
| III | L* | 20YL | 62.60 ** | 53.47 | 58.61 ± 4.97 | 49.46–67.21 | −0.04 | −1.16 | 8.48 |
| 20BJ | 64.21 ** | 54.46 | 59.17 ± 3.43 | 52.93–67.57 | 0.36 | −0.60 | 5.79 | ||
| 21YL | 63.93 ** | 51.09 | 55.42 ± 3.41 | 49.08–62.78 | 0.19 | −0.99 | 6.15 | ||
| 21BJ | 62.32 ** | 52.90 | 57.73 ± 3.28 | 50.58–68.15 | 0.34 | 0.03 | 5.68 | ||
| a* | 20YL | 6.06 | 11.06 ** | 10.00 ± 3.83 | 4.77–18.75 | 0.71 | −0.62 | 38.30 | |
| 20BJ | 5.49 | 7.46 ** | 7.70 ± 2.18 | 2.73–14.44 | 0.29 | −0.30 | 28.37 | ||
| 21YL | 6.63 | 13.83 ** | 7.65 ± 2.43 | 3.04–14.04 | 0.70 | −0.15 | 31.76 | ||
| 21BJ | 5.77 | 8.79 ** | 7.82 ± 2.39 | 2.58–14.42 | 0.34 | −0.50 | 30.54 | ||
| B* | 20YL | 23.44 ** | 17.56 | 20.42 ± 2.92 | 12.50–27.35 | −0.45 | −0.32 | 14.31 | |
| 20BJ | 22.69 ** | 12.79 | 18.91 ± 2.70 | 12.95–25.24 | 0.07 | −0.75 | 14.30 | ||
| 21YL | 25.25 ** | 16.25 | 16.83 ± 2.45 | 10.68–22.66 | 0.06 | −0.34 | 14.57 | ||
| 21BJ | 22.11 * | 14.05 | 18.43 ± 2.15 | 13.65–23.73 | 0.08 | −0.50 | 11.68 | ||
| C* | 20YL | 24.21 ** | 20.77 | 23.15 ± 2.07 | 18.28–29.54 | 0.27 | −0.25 | 8.94 | |
| 20BJ | 23.35 ** | 14.81 | 20.54 ± 2.70 | 14.93–26.36 | −0.03 | −0.70 | 13.13 | ||
| 21YL | 26.11 ** | 21.35 | 18.71 ± 1.93 | 12.96–23.70 | −0.04 | 0.37 | 10.29 | ||
| 21BJ | 22.85 ** | 16.59 | 20.18 ± 2.01 | 14.78–25.15 | −0.02 | −0.14 | 9.97 | ||
| IV | R | 20YL | 176.00 ** | 155.83 | 169.67 ± 9.87 | 150.00–189.33 | −0.03 | −0.96 | 5.82 |
| 20BJ | 179.22 ** | 150.78 | 166.96 ± 8.65 | 147.67–186.67 | 0.07 | −0.77 | 5.18 | ||
| 21YL | 181.33 ** | 153.00 | 155.70 ± 7.49 | 138.00–173.67 | 0.24 | −0.75 | 4.81 | ||
| 21BJ | 174.00 * | 149.33 | 162.98 ± 7.29 | 146.00–188.33 | 0.47 | 0.35 | 4.47 | ||
| G | 20YL | 145.89 ** | 119.67 | 133.38 ± 14.90 | 106.67–158.67 | −0.09 | −1.17 | 11.17 | |
| 20BJ | 150.44 ** | 124.67 | 136.30 ± 9.71 | 118.33–161.33 | 0.39 | −0.58 | 7.13 | ||
| 21YL | 149.00 ** | 111.67 | 126.81 ± 9.83 | 107.00–147.00 | 0.11 | −0.98 | 7.75 | ||
| 21BJ | 145.56 ** | 120.00 | 132.54 ± 9.56 | 111.67–161.67 | 0.27 | −0.15 | 7.21 | ||
| B | 20YL | 109.56 ** | 97.50 | 105.32 ± 9.30 | 91.00–126.33 | 0.52 | −0.98 | 8.83 | |
| 20BJ | 115.11 ** | 108.11 | 109.24 ± 6.76 | 96.00–128.00 | 0.54 | −0.50 | 6.19 | ||
| 21YL | 110.00 ** | 94.00 | 103.55 ± 5.86 | 94.00–118.00 | 0.58 | −0.61 | 5.66 | ||
| 21BJ | 111.22 ** | 102.00 | 106.51 ± 6.50 | 96.00–125.33 | 0.68 | −0.36 | 6.10 | ||
| Trait | Source | DF | Sum of Square | Mean Square | F-Value | p-Value | H2 |
|---|---|---|---|---|---|---|---|
| L* | G | 249.00 | 33,194.66 | 133.31 | 142.71 | <0.001 | 0.92 |
| E | 3.00 | 5669.19 | 1889.73 | 2022.98 | <0.001 | ||
| G × E | 656.00 | 7533.67 | 11.48 | 12.29 | <0.001 | ||
| Error | 1816.00 | 1696.38 | 0.93 | ||||
| a* | G | 249.00 | 17,718.18 | 71.16 | 161.23 | <0.001 | 0.92 |
| E | 3.00 | 2806.67 | 935.56 | 2119.79 | <0.001 | ||
| G × E | 656.00 | 4008.35 | 6.11 | 13.84 | <0.001 | ||
| Error | 1816.00 | 801.48 | 0.44 | ||||
| b* | G | 249.00 | 11,903.01 | 47.80 | 50.60 | <0.001 | 0.83 |
| E | 3.00 | 4713.88 | 1571.29 | 1663.27 | <0.001 | ||
| G × E | 656.00 | 6290.05 | 9.59 | 10.15 | <0.001 | ||
| Error | 1816.00 | 1715.58 | 0.94 | ||||
| C* | G | 249.00 | 7268.45 | 29.19 | 28.11 | <0.001 | 0.76 |
| E | 3.00 | 7360.86 | 2453.62 | 2362.70 | <0.001 | ||
| G × E | 656.00 | 5761.24 | 8.78 | 8.46 | <0.001 | ||
| Error | 1816.00 | 1885.88 | 1.04 | ||||
| R | G | 249.00 | 137,940.86 | 553.98 | 73.80 | <0.001 | 0.87 |
| E | 3.00 | 78,684.20 | 26,228.07 | 3494.23 | <0.001 | ||
| G × E | 656.00 | 54,585.31 | 83.21 | 11.09 | <0.001 | ||
| Error | 1816.00 | 13631.10 | 7.51 | ||||
| G | G | 249.00 | 290,243.59 | 1165.64 | 162.86 | <0.001 | 0.93 |
| E | 3.00 | 32,300.08 | 10766.69 | 1504.27 | <0.001 | ||
| G × E | 656.00 | 60,297.04 | 91.92 | 12.84 | <0.001 | ||
| Error | 1816.00 | 12,997.91 | 7.16 | ||||
| B | G | 249.00 | 118,416.31 | 475.57 | 91.49 | <0.001 | 0.92 |
| E | 3.00 | 11,400.34 | 3800.12 | 731.03 | <0.001 | ||
| G × E | 656.00 | 25,628.89 | 39.07 | 7.52 | <0.001 | ||
| Error | 1816.00 | 9440.11 | 5.20 |
| Methods | Traits | QTLi | Marker Interval | QTLj | Marker Interval | Epistatic Effect (AA) | Epistatic × Environment Interaction Effect | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AA | h2/% | AA-E1 | AA-E2 | AA-E3 | AA-E4 | h2/% | ||||||
| I | HCVGM | qGM1.2 | C01B039–C01B040 | qGM9.1 | C09B174–C09B175 | 0.22 c | 2.34 | 0.00 | 0.00 | 0.00 | 0.00 | 0.03 |
| II | HCVCM | qCM1.2 | C01B039–C01B040 | qCM9.2 | C09B174–C09B175 | 0.82 c | 2.53 | 0.00 | 0.00 | 0.00 | 0.00 | 0.03 |
| HCVCM | qCM1.4 | C01B042–C01B043 | qCM9.2 | C09B174–C09B175 | 1.58 c | 0.57 | 0.00 | 0.00 | 0.00 | 0.00 | 0.10 | |
| III | HCL | qL*2.4 | C02B060–C02B061 | qL*3.7 | C03B022–C03B023 | 0.37 c | 0.60 | 0.00 | 0.00 | 0.00 a | 0.00 | 0.03 |
| HCa | qa*1.2 | C01B039–C01B040 | qa*9.1 | C09B174–C09B175 | 0.31 c | 2.52 | 0.36 c | -0.15 | −0.09 | −0.12 | 0.67 | |
| HCa | qa*1.3 | C01B041–C01B042 | qa*9.2 | C09B172–C09B173 | 0.36 c | 0.42 | 0.00 | 0.00 | 0.00 a | 0.00 | 0.01 | |
| HCb | qb*1.3 | C01B041–C01B042 | qb*9.4 | C09B175–C09B176 | −0.47 c | 0.96 | −0.53 c | 0.10 | 0.32 | 0.10 | 0.70 | |
| HCC | qC*7.2 | C07B139–C07B140 | qC*9.3 | C09B021–C09B022 | 0.33 c | 1.17 | −0.03 | 0.17 | −0.04 | −0.09 | 0.31 | |
| IV | HCR | qR*1.2 | C01B053–C01B054 | qR*9.3 | C09B092–C09B093 | 1.55 c | 0.51 | 0.00 | 0.00 | 0.00 | 0.00 | 0.13 |
| HCR | qR*1.2 | C01B053–C01B054 | qR*9.4 | C09B100–C09B101 | −2.41 c | 0.03 | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | |
| HCG | qG*1.2 | C01B039–C01B040 | qG*9.3 | C09B174–C09B175 | 0.97 c | 0.68 | −0.76 | 0.04 | 0.55 | 0.18 | 0.29 | |
| HCG | qG*2.1 | C02B061–C02B062 | qG*3.2 | C03B024–C03B025 | 1.00 c | 0.45 | 0.00 | 0.00 | 0.00 | 0.00 | 0.09 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Z.; Chai, S.; Wu, Y.; Li, Y.; Han, H.; Song, H.; Gao, J.; Feng, B.; Yang, P. QTL Identification of Hull Color for Foxtail Millet [Setaria italica (L.) P. Beauv.] Through Four Phenotype Identification Strategies in a RIL Population. Seeds 2025, 4, 10. https://doi.org/10.3390/seeds4010010
Ma Z, Chai S, Wu Y, Li Y, Han H, Song H, Gao J, Feng B, Yang P. QTL Identification of Hull Color for Foxtail Millet [Setaria italica (L.) P. Beauv.] Through Four Phenotype Identification Strategies in a RIL Population. Seeds. 2025; 4(1):10. https://doi.org/10.3390/seeds4010010
Chicago/Turabian StyleMa, Zhixiu, Shaohua Chai, Yongjiang Wu, Yujie Li, Huibing Han, Hui Song, Jinfeng Gao, Baili Feng, and Pu Yang. 2025. "QTL Identification of Hull Color for Foxtail Millet [Setaria italica (L.) P. Beauv.] Through Four Phenotype Identification Strategies in a RIL Population" Seeds 4, no. 1: 10. https://doi.org/10.3390/seeds4010010
APA StyleMa, Z., Chai, S., Wu, Y., Li, Y., Han, H., Song, H., Gao, J., Feng, B., & Yang, P. (2025). QTL Identification of Hull Color for Foxtail Millet [Setaria italica (L.) P. Beauv.] Through Four Phenotype Identification Strategies in a RIL Population. Seeds, 4(1), 10. https://doi.org/10.3390/seeds4010010