
Genetic Parameters in Mesocotyl Elongation and Principal Components for Corn in High Valleys, Mexico

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
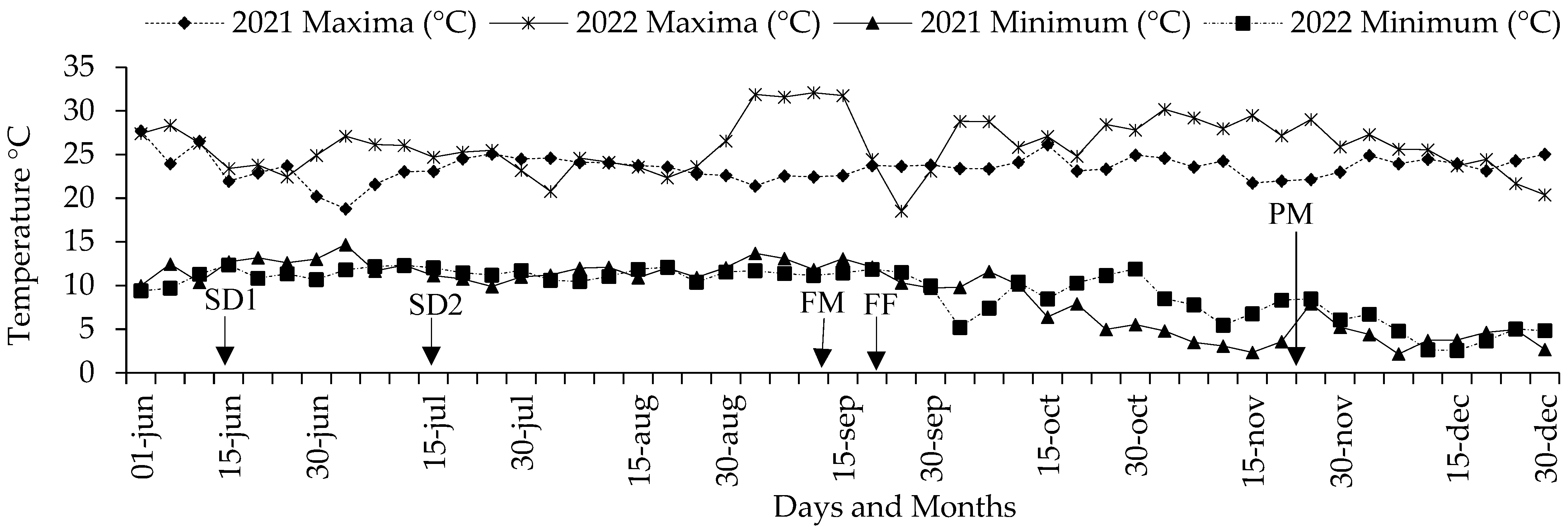
2.1. Localization of the Experiments
2.2. Genetic Material and Conditions of Experiments
2.3. Variables and Statistical Analysis
3. Results
3.1. Variability in the GCA and SCA
3.2. GCA of the Parents
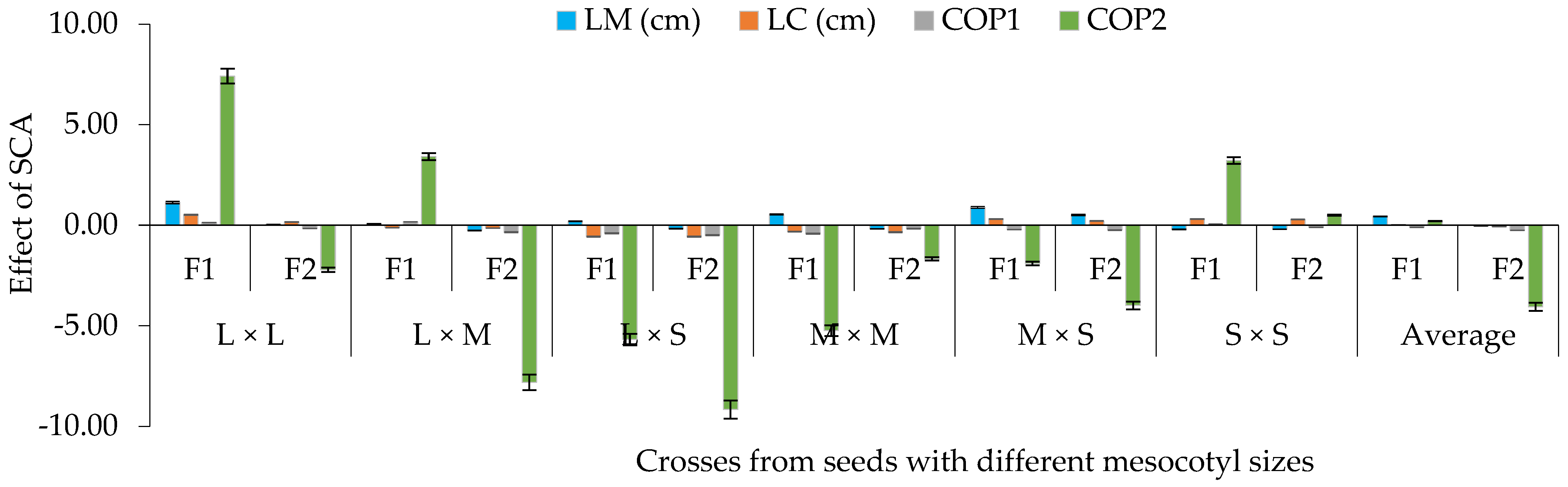
3.3. SCA for Crosses
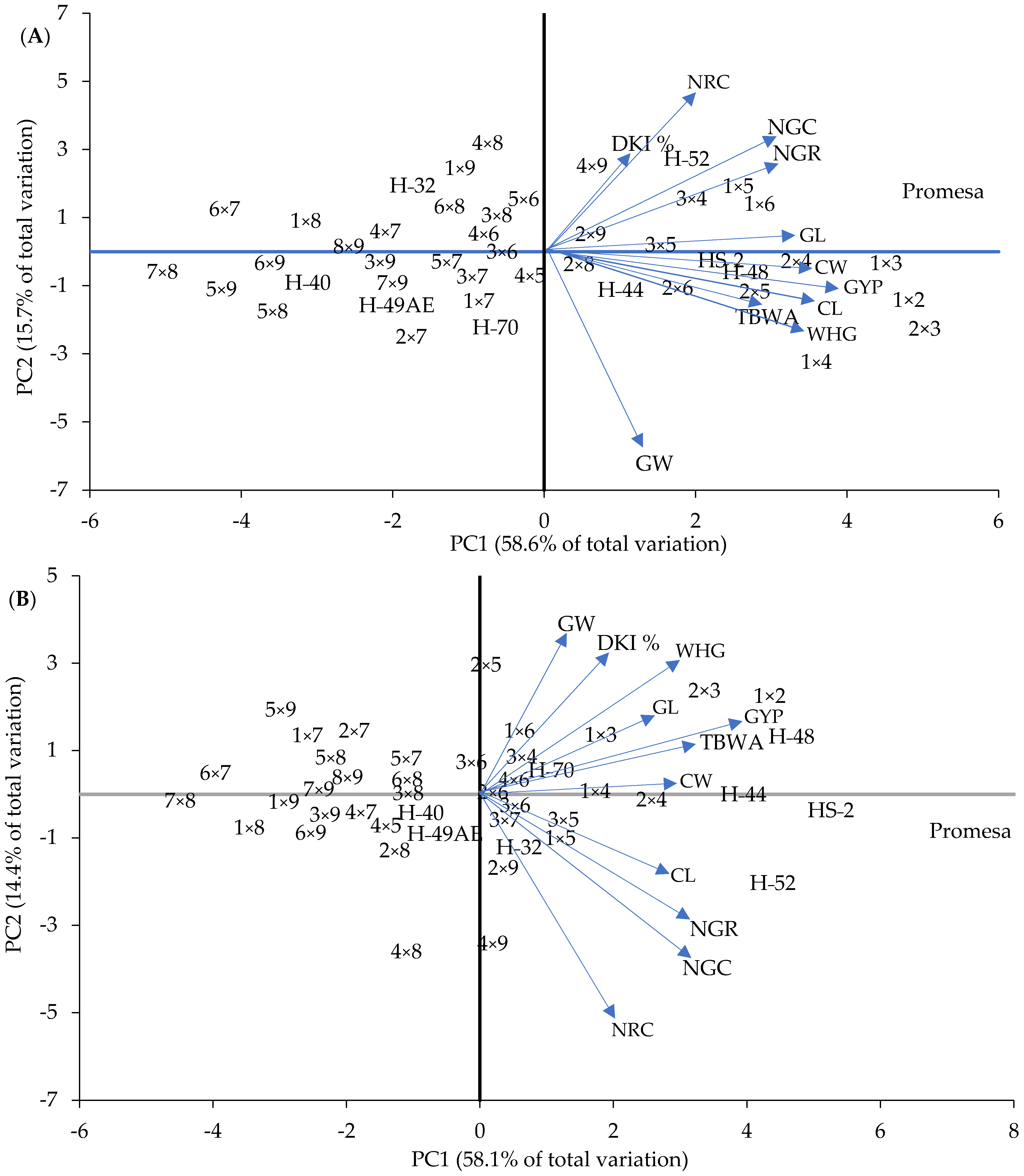
3.4. Interaction between Crosses, Yield and Length of Mesocotyl
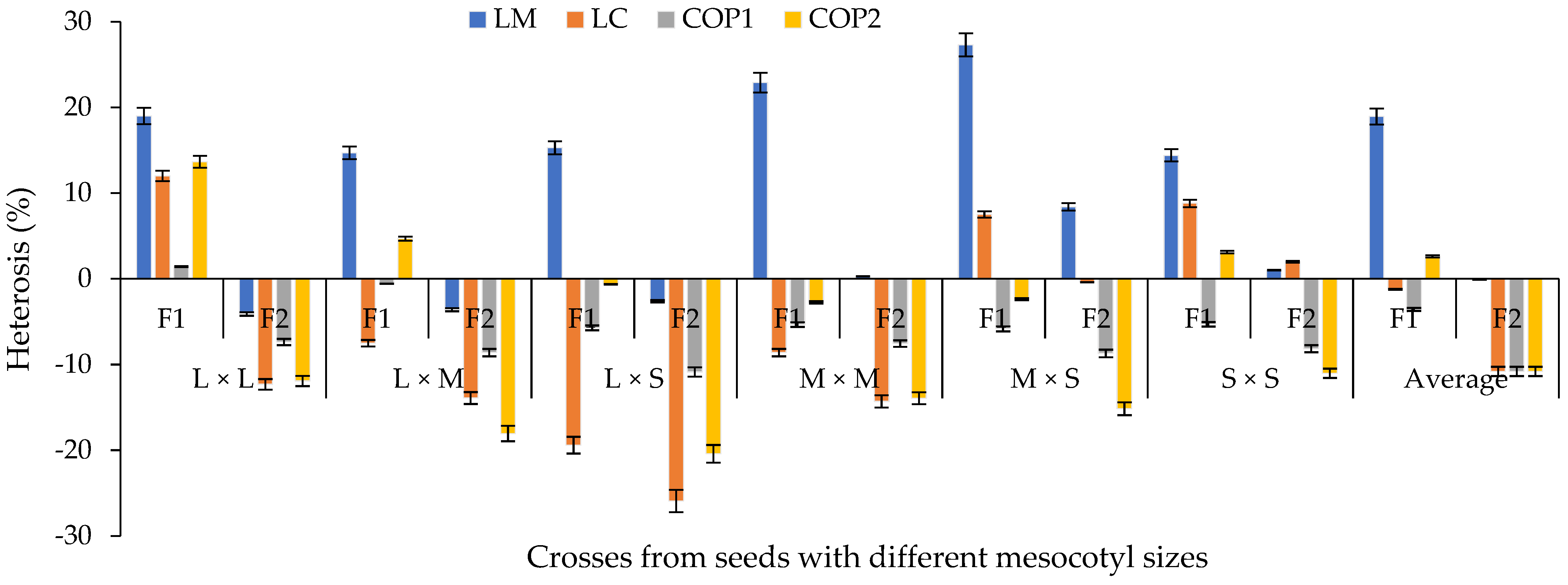
3.5. Heterosis and Inbreeding Depression (ID)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jameel, A.A.; Hassan, H.B. Genetic analysis by using partial diallel crossing of maize in high plant densities (Estimation GCA, SCA and Some Genetic Parameters). Earth Environ. Sci. 2021, 910, 012135. [Google Scholar] [CrossRef]
- Begna, T. Combining ability and heterosis in plant improvement. Open J. Plant Sci. 2021, 6, 108–117. [Google Scholar] [CrossRef]
- Rodríguez, P.G.; Treviño, R.J.E.; Ojeda, Z.M.C.; Cervantes, O.F.; Ávila, P.M.A.; Gámez, V.A.J. Genetic parameters and combining ability of corn lines for grain. Rev. Mex. Agric. Sci. 2020, 11, 1867–1878. [Google Scholar] [CrossRef]
- Wu, X.; Liu, Y.; Zhang, Y.; Gu, R. Advances in research on the mechanism of heterosis in plants. Front. Plant Sci. 2021, 12, 745726. [Google Scholar] [CrossRef] [PubMed]
- Samayoa, L.F.; Olukolu, A.B.; Jian, C.Y.; Chen, Q.; Stetter, M.G.; York, A.M.; Sánchez, G.J.J.; Glaubitz, J.C.; Bradbury, P.J.; Romay, M.C.; et al. Domestication reshaped the genetic basis of inbreeding depression in a maize landrace compared to its wild relative, teosinte. PLoS Genet. 2021, 17, e1009797. [Google Scholar] [CrossRef] [PubMed]
- Hasan, R.S.; Husain, M.A. Estimation combining ability and some genetic parameters for maize (Zea mays L.) and its componenets using line x tester. J. Duhok Univ. 2022, 25, 229–237. [Google Scholar] [CrossRef]
- Yu, K.; Wang, H.; Liu, X.; Xu, C.; Li, Z.; Xu, X.; Liu, J.; Wang, Z.; Xu, Y. Large-scale analysis of combining ability and heterosis for development of hybrid maize breeding strategies using diverse germplasm resources. Front. Plant Sci. 2020, 11, 660. [Google Scholar] [CrossRef]
- Griffing, B. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci. 1956, 9, 463–493. Available online: https://www.publish.csiro.au/bi/pdf/bi9560463 (accessed on 28 August 2022). [CrossRef]
- Mohamed, M.H.R.; Mohamed, D.Z.; Rizk, A.S.H. Combining abilities and genetic parameters for grain yield and some agronomic traits in maize (Zea mays L.). Beni-Suef Univ. J. Basic Appl. Sci. 2022, 11, 108. [Google Scholar] [CrossRef]
- Souza, N.I.L.; Torre, S.F.A.; Santos, R.U.; Contreras, I.S.R.; Alberto, C.S.; Dutra, M.Z. Combining ability and heterotic pattern in relation to F1 performance of tropical and temperate-adapted sweet corn lines. Bragantia 2022, 81, e3422. [Google Scholar] [CrossRef]
- Yuan, Z.; Xiaoqiang, Z.; Wenli, L.; Dan, Z.; Wenqi, Z. Heterosis and genetic effects analysis of deep-seeding traits in maize under different sowing environments. J. Nucl. Agric. Sci. 2021, 35, 556–566. [Google Scholar] [CrossRef]
- Yue, H.; Gauch, G.H.; Wei, J.; Xie, J.; Chen, S.; Peng, H.; Bu, J.; Jiang, X. Genotype by environment interaction analysis for grain yield and yield components of summer maize hybrids across the huanghuaihai region in China. Agriculture 2022, 12, 602. [Google Scholar] [CrossRef]
- Sáenz, R.M.N.; Cassab, G.I. Primary root and mesocotyl elongation in maize seedlings: Two organs with antagonistic growth below the soil surface. Plants 2021, 10, 1274. [Google Scholar] [CrossRef] [PubMed]
- Benavente, E.; Giménez, E. Modern approaches for the genetic improvement of rice, wheat and maize for abiotic constraints-related traits: A comparative overview. Agriculture 2021, 11, 376. [Google Scholar] [CrossRef]
- Allier, A.; Teyssedre, S.; Lehermeier, C.; Moreau, L.; Charcosset, A. Optimized breeding strategies to harness genetic resources with different performance levels. BMC Genom. 2020, 21, 349. [Google Scholar] [CrossRef] [PubMed]
- Villalobos, G.A.; Benítez, R.I.; Castillo, G.F.; Mendoza, C.M.D.C.; Espinosa, C.A. Assessment of elongation of the mesocotyl-coleoptile and biomass in parents and crosses of corn seedlings of the high Valleys of Mexico. Seeds 2023, 2, 449–473. [Google Scholar] [CrossRef]
- Lu, Y.H.; Xu, Y.L.; Xiao, J.Z.; Bao, C.S.; Cheng, L.; Deng, F.Z.; Huai, J.T.; Chun, H.L.; Yong, X.L.; Yun, S.S.; et al. Genome-wide identification and comparative analysis of drought related genes in roots of two maize inbred lines with contrasting drought tolerance by RNA sequencing. J. Int. Agric. 2020, 8, 449–464. [Google Scholar] [CrossRef]
- Yan, W.; Frégeau, R.J. Genotype by yield*trait (GYT) biplot: A novel approach for genotype selection based on multiple traits. Sci. Rep. 2018, 8, 8242. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, X.; Fu, J.; Wang, H.; Wang, J.; Huang, H.; Zhang, A. Enhancing genetic gain through genomic selection: From Livestock to Plants. Plant Commun. 2020, 1, 100005. [Google Scholar] [CrossRef]
- Miedaner, T.; Juroszek, T. Global warming and increasing maize cultivation demand comprehensive efforts in disease and insect resistance breeding in north-western Europe. Plants Pathol. 2021, 70, 1032–1046. [Google Scholar] [CrossRef]
- Villalobos, L.M.A.; Arroyo, B.A.; Quintero, J.A.; Iturriaga, G. Biotechnological advances to improve abiotic stress tolerance in crops. Int. J. Mol. Sci. 2022, 23, 12053. [Google Scholar] [CrossRef] [PubMed]
- SAS. The SAS System Release for Windows 9.0; SAS Institute Inc.: Cary, NC, USA, 2002. [Google Scholar]
- Yan, W. Crop variety trials: Data management and analysis. Crop Sci. 2014, 54, 2910–2911. [Google Scholar] [CrossRef]
- Labroo, M.R.; Studer, A.J.; Rutkoski, J.E. Heterosis and hybrid crop breeding: A multidisciplinary review. Front. Genet. 2021, 12, 643761. [Google Scholar] [CrossRef] [PubMed]
- López, L.C.; Tadeo, M.R.; García, J.Z.J.; Espinosa, A.C.; Mejía, A.J.C. General and specific combinatorial aptitude of varietal hybrids of low inbreeding yellow corn. Rev. Mex. Agric. Sci. 2021, 12, 699–711. [Google Scholar] [CrossRef]
- Baker, R.J. Issues in diallel analysis. Crop Sci. 1978, 18, 533–536. [Google Scholar] [CrossRef]
- Benowicz, A.; Stoehr, M.; Hamann, A.; Yanchuk, D.A. Estimation of the F2 generation segregation variance and relationships among growth, frost damage, and bud break in coastal Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) wide-crosses. Ann. For. Sci. 2020, 77, 28. [Google Scholar] [CrossRef]
- Nasir, M.S.M.; Nagy, J.J. Evaluation of plant characteristics related to grain yield of FAO410 and FAO340 hybrids using regression models. Cereal Res. Commun. 2021, 49, 161–169. [Google Scholar] [CrossRef]
- Aman, J.; Bantte, K.; Alamerew, S.; Berhe, S.D. Correlation and path coefficient analysis of yield and yield components of quality protein maize (Zea mays L.) hybrids at jimma, western Ethiopia. Int. J. Agron. 2020, 2020, 9651537. [Google Scholar] [CrossRef]
- Noori, R.M. Yield and yield components of maize and soil physical properties as affected by tillage practices and organic mulching. Saudi J. Biol. Sci. 2021, 28, 7152–7159. [Google Scholar] [CrossRef]
- Díaz, C.P.; Hidalgo, M.E.; Mendoza, P.M.; Cieza, R.I.; Jara, C.T.W.; Valdés, R.O.A. New thilinear hybrid of hard yellow corn for the Peruvian tropic. Agron. Meso. 2023, 34, 51177. [Google Scholar] [CrossRef]
- Gutiérrez, H.G.F.; Arellano, V.J.L.; Ceja, T.L.F.; García, R.E.; Quiroz, F.F.R. Heterosis in the germination process and seed characteristics of the maize hybrid (Zea mays L.) HAZUL 10E. Agro Product. 2023, 16, 119–125. [Google Scholar] [CrossRef]
- Adebiyi, O.O.; Gbemavo, G.K.; Adewole, A.K. Genetic variability for mesocotyl length in maize. Biol. Life Sci. Forum 2021, 4, 26. [Google Scholar] [CrossRef]
- Yi, Q.; Liu, Y.; Hou, X.; Zhang, X.; Li, H.; Zhang, J.; Liu, H.; Hu, Y.; Yu, G.; Li, Y.; et al. Genetic dissection of yield-related traits and mid-parent heterosis for those traits in maize (Zea mays L.). BMC Plant Biol. 2019, 19, 392. [Google Scholar] [CrossRef] [PubMed]
- Mariano, C.L.; Neves, S.P.; Valéria, C.N.; Crispim, F.A.J.; Mendes, R.M.P.; Santos, M.P.G. Inbreeding depression and genetic variability of populations for green maize production. Rev. Ceres Vicosa 2022, 69, 709–719. [Google Scholar] [CrossRef]
- Kashiani, P.; Saleh, G.; Abdullah, N.A.P.; Abdullah, S.N. Variation and Genetic Studies on Selected Sweet Corn Inbred Lines. Asian J. Crop Sci. 2010, 2, 78–84. [Google Scholar] [CrossRef]
- Kuki, M.C.; Scapim, C.A.; Barth, P.R.J.; Torre, F.A.S.; Contreras, S.R.I.; Bengosi, B.F.A. Inbreeding depression and average genetic components in green corn genotypes. Rural Sci. 2017, 47, e20160024. [Google Scholar] [CrossRef]
- Días, M.M.F.S.; Bart, P.R.J.; Aparecida, D.S.T.; Ary, R.D.; Akira, M.R.; Gibim, E.T.; Lago, G.G.D. Partial diallel and genetic divergence analyses in maize inbred lines. Acta Scientiarum. Agriculture 2021, 43, e53540. [Google Scholar] [CrossRef]
- Nardino, M.; Barros, W.; Olivato, T.; Damião, C.C.; E Silva, F.F.; DE Pelegrín, A.J.; DE Souza, V.Q.; Carvalho, I.R.; Szareski, V.J.; DE Oliveira, A.C.; et al. Multivariate diallel analysis by factor analysis for establish mega-traits. An. Acab. Bras. Sci. 2020, 92, e20180874. [Google Scholar] [CrossRef]
- Muthoni, J.; Shimelis, H. Mating designs commonly used in plant breeding: A review. AJCS 2020, 12, 1855–1869. [Google Scholar] [CrossRef]
- Matongera, N.; Ndhlela, T.; Van, B.A.; Kamutando, C.N.; Labuschagne, M. Combining ability and testcross performance of multi-nutrient maize under stress and non-stress environments. Front. Plant Sci. 2023, 14, 1070302. [Google Scholar] [CrossRef]
- Inocente, G.; Domingos, G.D.; Araújo, P.M.; Ruas, P.M. Heritability and combined parental information to define the number of crosses in circulant diallels. Crop Breed. Appl. Biotechnol. 2021, 2, e37472125. [Google Scholar] [CrossRef]
- Fan, X.M.; Zhang, Y.D.; Yao, W.H.; Bi, Y.Q.; Liu, L.; Chen, H.M.; Kang, M.S. Reciprocal diallel crosses impact combining ability, variance estimation, and heterotic group classification. Crop Sci. 2014, 54, 89–97. [Google Scholar] [CrossRef]
- Lessa, A.G.M.; Fonseca, R.; Damião, C.C.; Carneiro, J.J.M. Estimation of variances of the effects of incomplete diallels using a matrix approach. Genet. Mole. Biol. 2004, 27, 409–417. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Z.; Xu, Y.; Li, P.; Zhang, X.; Xu, C. Using genomic data to improve the estimation of general combining ability based on sparse partial diallel cross designs in maize. Crop J. 2020, 8, 819–829. [Google Scholar] [CrossRef]
- Xavier, S.L.F.; Pestana, J.K.; Sekiya, A.; Krause, D.M.; Moreira, P.R.M.; Ferreira, J.M. Partial diallel and potential of super sweet corn inbred lines bt2 to obtain hybrids. Hort. Bras. 2019, 37, 278–284. [Google Scholar] [CrossRef]
- Cervantes, O.F.; Hernández, E.J.; Rangel, L.J.A.; Andrio, E.E.; Mendoza, E.M.; Rodríguez, P.G.; Guevara, A.L.P. General and specific combinatorial aptitude in seed quality of s3 corn lines. Revest Mex. Phytotech. 2016, 39, 259–268. [Google Scholar] [CrossRef]
- Bhusal, T.N.; Lal, M.G. Heterosis, Combining ability and their inter-relationship for morphological and quality traits in yellow maize (Zea mays L.) single-crosses across environments. Agrivita J. Agric. Sci. 2020, 42, 174–190. [Google Scholar] [CrossRef]
- Ramos, G.P.H.; Pereira, A.C.; Colombari, M.F.J.; Pereira, P.T.; Nakano, H.R.P.; Santos, P.M.G. Diallel analysis: Choosing parents to introduce new variability in a recurrent selection population. Agriculture 2022, 13, 1320. [Google Scholar] [CrossRef]
- Barrera, G.L.A.; Legaria, P.S.J.; Ortega, R.P. Genetic diversity in populations of mexican maize races. Revest Mex. Phytotech. 2020, 43, 121–125. [Google Scholar] [CrossRef]
- Oliveira, S.T.; Oliveira, F.T.; Amaral, A.J.T.; Almeida, J.F.E.; Barboza, R.B.; Mendonça, F.M.S.; Teixeira, A.J.F.; Henrique, K.S.; Jário, L.V.; Nicácio, V.F.; et al. Additive and non-additive effects on the control of key agronomic traits in popcorn lines under contrasting phosphorus conditions. Plants 2022, 11, 2216. [Google Scholar] [CrossRef]
- Tafolla, R.M.A.; Rincón, F.S.; Ruiz, A.T.N.; Sánchez, J.R.F.; Martínez, M.R.J.; Benavides, A.M. Genetic and environmental effects of prolificacy in relation to maize yield. Agrociencia 2021, 55, 507–521. [Google Scholar] [CrossRef]
- Nemergut, T.K.; Thomison, R.P.; Carter, R.P.; Lindsey, J.A. Planting depth affects corn emergence, growth and development, and yield. Agron. J. 2021, 113, 3351–3360. [Google Scholar] [CrossRef]
- Pierre, F.; Rodríguez, Y.I.; Colbert, W.R.; Rosas, C.J. Agronomic behavior of native and improved corn varieties in a low fertility soil. Ceiba 2003, 56, 16–30. [Google Scholar] [CrossRef]
- Ortez, O.A.; McMechan, A.J.; Hoegemeyer, T.; Ciampitti, I.A.; Nielsen, R.L.; Thomison, P.R.; Abendroth, L.J.; Elmore, R.W. Conditions potentially affecting corn ear formation, yield, and abnormal ears: A review. Crop For. Turf. Mgmt. 2022, 8, e20173. [Google Scholar] [CrossRef]
- Wang, L.; Yu, X.; Gao, J.; Ma, D.; Guo, H.; Hu, S. Patterns of influence of meteorological elements on maize grain weight and nutritional quality. Agriculture 2023, 13, 424. [Google Scholar] [CrossRef]
- Stewart, S.; Kitchen, N.; Yost, M.; Stephen, C.L.; Carter, P. Planting depth and within-field soil variability impacts on corn stand establishment and yield. Agrosystems Geosci. Environ. 2021, 4, e20186. [Google Scholar] [CrossRef]
- Pérez, D.C.F.J.; Córdova, T.L.; Santacruz, V.A.; Castillo, G.F.; Cárdenas, S.E.; Delgado, A.A. Relationship between Initial Vigor, Yield and Its Components in Chalqueño Maize Populations. Tech. Agric. Mex. 2007, 33, 5–16. Available online: http://www.redalyc.org/articulo.oa?id=60833101 (accessed on 13 September 2022).
- Liu, H.; Zhang, L.; Wang, J.; Li, C.; Zeng, X.; Xie, S.; Zhang, Y.; Liu, S.; Hu, S.; Wang, J.; et al. Quantitative trait locus analysis for deep-sowing germination ability in the maize IBM syn10 DH population. Front. Plant Sci. 2017, 8, 813. [Google Scholar] [CrossRef]
- Bousselot, J.M.; Muenchrath, D.; Knapp, D.A.; Reeder, D.J. Emergence and seedling characteristics of maize native to the southwestern USA. Am. J. Llant Sci. 2017, 8, 1304–1318. [Google Scholar] [CrossRef]
- Santiago, L.N.; García, Z.J.G.; Santiago, L.U.; Esquivel, E.G. Yield of varieties of pozolero corn of the ‘elotes occidentales’ race evaluated in High Valleys of Mexico. Revest Mex. Agric. Sci. 2023, 14, 96–104. [Google Scholar] [CrossRef]
- Zhao, X.; Niu, Y.; Hossain, Z.; Zhao, B.; Bai, X.; Mao, T. New insights into light spectral quality inhibits the plasticity elongation of maize mesocotyl and coleoptile during seed germination. Front. Plant Sci. 2023, 14, 1152399. [Google Scholar] [CrossRef]
- Reed, R.C.; Bradford, J.F.; Khanday, I. Seed germination and vigor: Ensuring crop sustainability in a changing climate. Heredity 2022, 128, 450–459. [Google Scholar] [CrossRef]
- Castro, A.M.L.; Sánchez, H.S.B.; Ruelas, R.I.J.; Romero, S.F.C.; Buelna, S.T.; Almada, G.R.V. Morpho-physiological characteristics of corn (Zea mays L.) affected by drought during its vegetative stage. Agro Product. 2021, 14, 71–77. [Google Scholar] [CrossRef]
- Chen, C.X.; Ping, Z.; Yuan, W.Y.; Ning, L.; Bei, T.J.; Xi, L.W.; Pu, W.; Shou, H.R. Grain yield and grain moisture associations with leaf, stem and root characteristics in maize. J. Int. Agric. 2022, 21, 1941–1951. [Google Scholar] [CrossRef]
- Hemada, S.S.H.; Yassien, H.E.; Abd, I.N.E.L.; Haridy, M.H. Combining ability and genetic parameters of some white maize (Zea mays L.) inbred lines using diallel analysis. Arch. Agric. Sci. J. 2020, 3, 82–100. [Google Scholar] [CrossRef]
- Holá, D.; Kočová, M.; Rothová, O.; Wilhelmová, N.; Benešová, M. Recovery of maize (Zea mays L.) inbreds and hybrids from chilling stress of various duration: Photosynthesis and antioxidant enzymes. J. Plant Physiol. 2007, 164, 868–877. [Google Scholar] [CrossRef]
- Sowinski, P.; Rudzinska, L.A.; Adamczyk, J.; Kubica, I.; Fronk, J. Recovery of maize seedling growth, development and photosynthetic efficiency after initial growth at low temperature. J. Plant Physiol. 2005, 162, 67–80. [Google Scholar] [CrossRef]
- Markelz, H.N.; Costich, E.D.; Brutnell, P.T. Photomorphogenic responses in maize seedling development. Plant Physiol. 2003, 133, 1578–1591. [Google Scholar] [CrossRef]
- Cargnelutti, F.A.; Toebe, M. Sample size for principal component analysis in corn. Pesqui. Agropecuária Bras. 2021, 56, e02510. [Google Scholar] [CrossRef]
- Tucker, S.L.; Dohleman, F.G.; Grapov, D.; Flagel, L.; Yang, S.; Wegener, K.M.; Kosola, K.; Swarup, S.; Rapp, R.A.; Bedair, M.; et al. Evaluating maize phenotypic variance, heritability, and yield relationships at multiple biological scales across agronomically relevant environments. Plant Cell Eviron. 2019, 43, 880–902. [Google Scholar] [CrossRef]
- Thapa, M.B.; Acharya, S.; Gyawali, B.; Timilsena, K.; Upadhayaya, J.; Shrestha, J. Genetic variability and trait association in maize (Zea mays L.) varieties for growth and yield traits. Heliyon 2021, 7, e07939. [Google Scholar] [CrossRef]
- Hideyo, N.T.; Bengosi, A.B.F.; Kuki, M.C.; Scapim, A.C.; Barth, J.P.R.; Teixeira, A.; Júnior, A. Testers in supersweet corn lines. Rural Sci. 2020, 50, e20190447. [Google Scholar] [CrossRef]
- González, D.M.R.; Navarro, G.H.; Ortega, P.R.; Flores, S.D.; González, S.V. Peasant strategies for the use and conservation of native corn in Juchitepec, state in Mexico. Agro Product. 2022, 15, 129–143. [Google Scholar] [CrossRef]
- Merrick, L.; Beavis, W.; Edwards, J.; Lübberstedt, T.; Campbell, A.; Muenchrath, D.; Fei, S. Inbreeding and heterosis. In Crop Genetics, 1st ed.; Suza, W.P., Lamkey, K.R., Eds.; Iowa State University Digital Press: Ames, IA, USA, 2023; pp. 177–199. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Degrees of Freedom | GN | LM | LC | TBWS | CL | CW | GL | GW |
|---|---|---|---|---|---|---|---|---|---|
| Repetition | 3 | F1 | 9.83 * | 2.39 ns | 321.4 * | 1.58 ns | 0.25 ns | 1.3 ns | 0.67 ns |
| F2 | 0.95 ns | 0.07 ns | 151.6 ** | 0.88 ns | 0.10 ns | 0.8 ns | 0.65 * | ||
| Crosses | 44 | F1 | 24.79 ** | 3.48 ** | 983.8 ** | 9.79 ** | 0.53 ** | 4.1 ** | 1.25 ** |
| F2 | 12.72 ** | 3.04 ** | 512.4 ** | 7.48 ** | 0.39 ** | 3.7 ** | 0.95 ** | ||
| GCA | 8 | F1 | 78.96 ** | 12.11 ** | 2292.8 ** | 30.21 ** | 1.86 ** | 13.7 ** | 2.01 ** |
| F2 | 50.43 ** | 9.93 ** | 932.8 ** | 12.13 ** | 0.99 ** | 10.3 ** | 1.53 ** | ||
| SCA | 36 | F1 | 12.75 ** | 1.57 ** | 692.9 ** | 5.25 ** | 0.24 ns | 1.9 ** | 1.08 ** |
| F2 | 4.34 ** | 1.51 ** | 419.0 ** | 6.45 ** | 0.25 * | 2.3 ** | 0.82 ** | ||
| Error | 132 | F1 | 3.10 | 0.90 | 118.2 | 1.94 | 0.17 | 1.0 | 0.30 |
| F2 | 0.44 | 0.05 | 18.4 | 2.22 | 0.13 | 0.7 | 0.21 | ||
| Total | 179 | ||||||||
| C.V.% | F1 | 13.7 | 23.1 | 12.9 | 11.3 | 10.2 | 9.2 | 7.4 | |
| F2 | 6.0 | 6.0 | 6.4 | 13.0 | 9.4 | 7.9 | 6.5 | ||
| GCA/SCA | F1 | 0.93 | 0.94 | 0.87 | 0.92 | 0.94 | 0.94 | 0.79 | |
| F2 | 0.96 | 0.93 | 0.82 | 0.79 | 0.89 | 0.90 | 0.79 | ||
| Source of Variation | Degrees of Freedom | GN | NRC | NGR | NGC | GYP | WHG | DKI | TBWA |
|---|---|---|---|---|---|---|---|---|---|
| Repetition | 3 | F1 | 4.22 * | 23.3 ns | 6828.8 ns | 224.7 ns | 28.9 ns | 2.6 ns | 1074.4 ns |
| F2 | 0.24 ns | 19.8 ns | 6545.6 ns | 131.4 ns | 12.9 ns | 6.5 ns | 898.8 ns | ||
| Crosses | 44 | F1 | 7.57 ** | 83.4 ** | 28,187.3 ** | 2548.2 ** | 198.1 ** | 24.2 ** | 11,916.7 ** |
| F2 | 5.70 ** | 64.3 ** | 25,586.6 ** | 1454.6 ** | 99.1 ** | 30.8 ** | 5705.7 ** | ||
| GCA | 8 | F1 | 10.62 ** | 258.1 ** | 72,071.9 ** | 11,163.0 ** | 835.5 ** | 43.6 ** | 49,394.6 ** |
| F2 | 2.98 ns | 134.7 ** | 42,369.2 ** | 5362.9 ** | 393.1 ** | 52.7 ** | 21,562.6 ** | ||
| SCA | 36 | F1 | 6.89 ** | 44.6 ** | 18,435.2 ** | 633.8 ** | 56.4 ** | 19.9 * | 3588.2 ** |
| F2 | 6.30 ** | 48.69 ** | 21,857.2 ** | 586.1 ** | 33.8 ** | 25.9 ** | 2181.9 ** | ||
| Error | 132 | F1 | 1.71 | 20.1 | 4916.7 | 318.7 | 17.8 | 10.7 | 879.5 |
| F2 | 1.95 | 12.4 | 3469.8 | 143.7 | 9.8 | 11.6 | 538.0 | ||
| Total | 179 | ||||||||
| C.V. % | F1 | 9.0 | 21.8 | 22.5 | 22.7 | 18.4 | 3.9 | 15.0 | |
| F2 | 10.0 | 19.7 | 22.7 | 20.6 | 15.6 | 4.1 | 14.7 | ||
| GCA/SCA | F1 | 0.76 | 0.92 | 0.89 | 0.97 | 0.87 | 0.97 | 0.81 | |
| F2 | 0.49 | 0.85 | 0.79 | 0.95 | 0.95 | 0.96 | 0.80 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villalobos-González, A.; Benítez-Riquelme, I.; Castillo-González, F.; Mendoza-Castillo, M.d.C.; Espinosa-Calderón, A. Genetic Parameters in Mesocotyl Elongation and Principal Components for Corn in High Valleys, Mexico. Seeds 2024, 3, 149-168. https://doi.org/10.3390/seeds3010012
Villalobos-González A, Benítez-Riquelme I, Castillo-González F, Mendoza-Castillo MdC, Espinosa-Calderón A. Genetic Parameters in Mesocotyl Elongation and Principal Components for Corn in High Valleys, Mexico. Seeds. 2024; 3(1):149-168. https://doi.org/10.3390/seeds3010012
Chicago/Turabian StyleVillalobos-González, Antonio, Ignacio Benítez-Riquelme, Fernando Castillo-González, Ma. del Carmen Mendoza-Castillo, and Alejandro Espinosa-Calderón. 2024. "Genetic Parameters in Mesocotyl Elongation and Principal Components for Corn in High Valleys, Mexico" Seeds 3, no. 1: 149-168. https://doi.org/10.3390/seeds3010012
APA StyleVillalobos-González, A., Benítez-Riquelme, I., Castillo-González, F., Mendoza-Castillo, M. d. C., & Espinosa-Calderón, A. (2024). Genetic Parameters in Mesocotyl Elongation and Principal Components for Corn in High Valleys, Mexico. Seeds, 3(1), 149-168. https://doi.org/10.3390/seeds3010012