
Karrikins Reduce the Hypocotyl Length of Rapeseed (Brassica napus napus L.) under Continuous Red Light †



{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Plant Material
2.2. Hypocotyl Length Measurement
2.3. Statistical Analysis
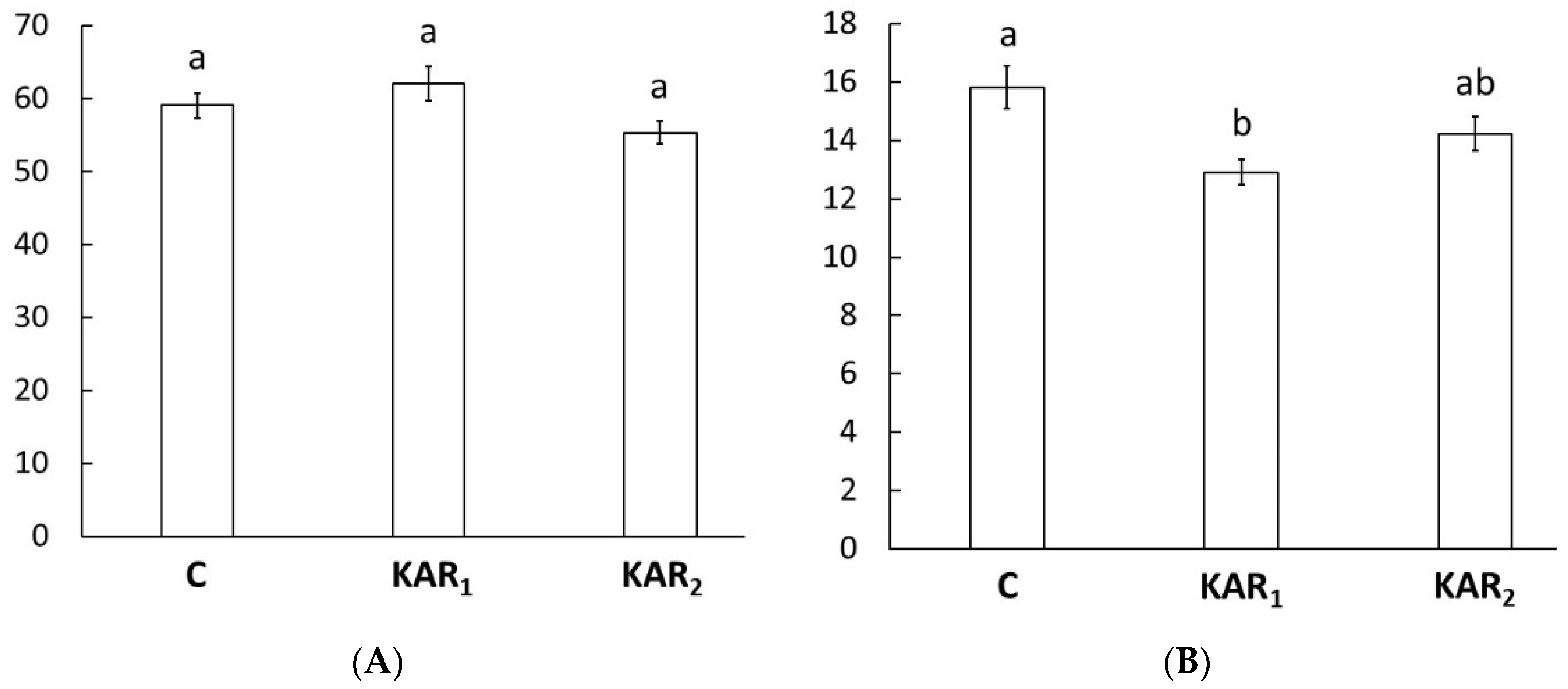
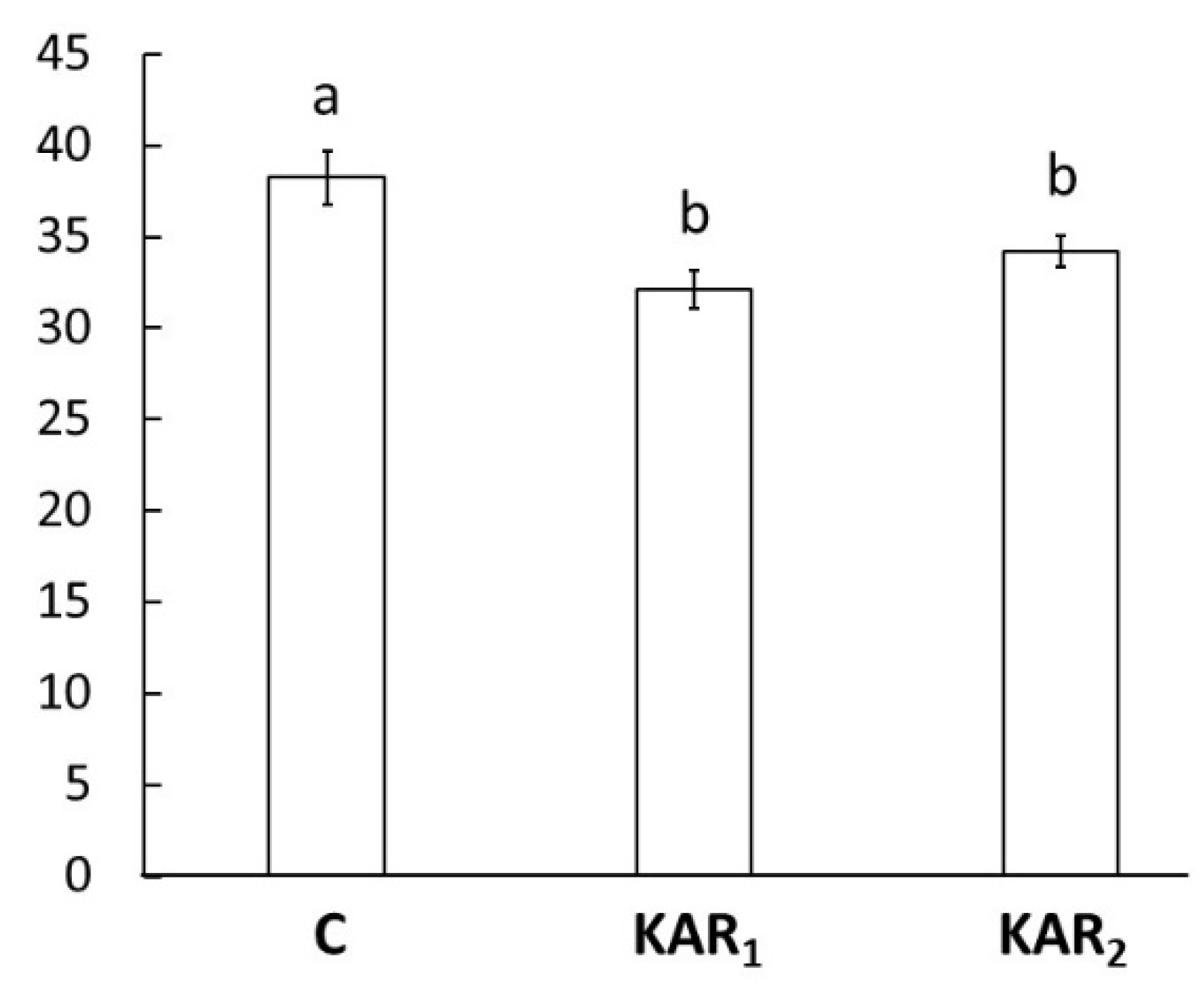
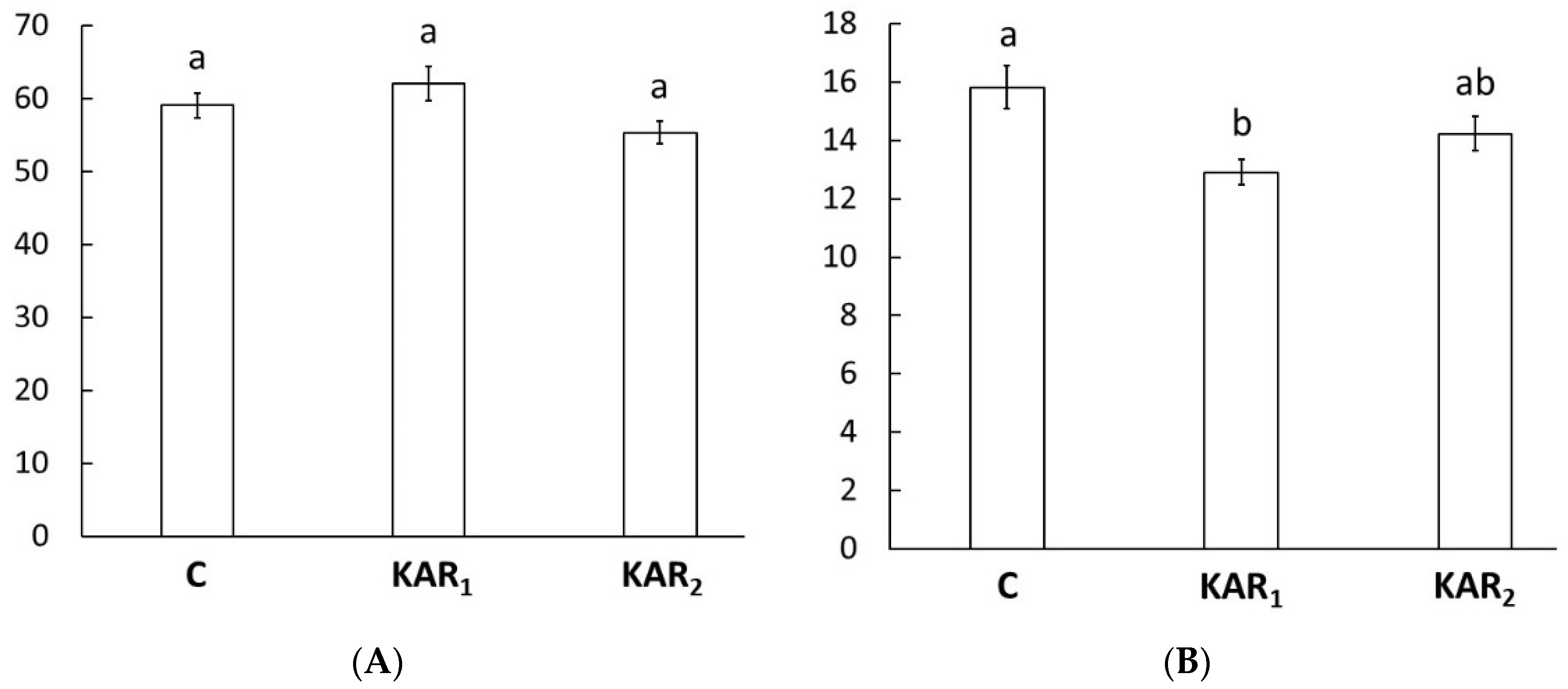
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nelson, D.C.; Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W.; Smith, S.M. Regulation of Seed Germination and Seedling Growth by Chemical Signals from Burning Vegetation. Ann. Rev. Plant Biol. 2012, 63, 107–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flematti, G.; Dixon, K.; Smith, S.M. What are karrikins and how were they ‘discovered’ by plants? BMC Biol. 2015, 13, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W.; Trengove, R.D. A Compound from Smoke That Promotes Seed Germination. Science 2004, 305, 977. [Google Scholar] [CrossRef] [PubMed]
- Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W.; Trengove, R.D. Identification of Alkyl Substituted 2H-Furo[2,3-c]pyran-2-ones as Germination Stimulants Present in Smoke. J. Agric. Food Chem. 2009, 57, 9475–9480. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.C.; Risenborough, J.-A.; Flematti, G.R.; Stevens, J.; Ghisalberti, E.L.; Dixon, K.W.; Smith, S.M. Karrikins Discovered in Smoke Trigger Arabidopsis Seed Germination by a Mechanism Requiring Gibberellic Acid Synthesis and Light. Plant Physiol. 2009, 149, 863–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, D.C.; Flematti, G.R.; Riseborough, J.A.; Ghisalberti, E.L.; Dixon, K.W.; Smith, S.M. Karrikins enhance light responses during germination and seedling development in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 7095–7100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.N.; Khan, Z.; Luo, T.; Liu, J.; Rizwan, M.; Zhang, J.; Xu, Z.; Wu, H.; Hu, L. Seed priming with gibberellic acid and melatonin in rapeseed: Consequences for improving yield and seed quality under drought and non-stress conditions. Ind. Crops Prod. 2020, 156, 112850. [Google Scholar] [CrossRef]
- FAO. Oilcrops, Oils and Meals; FAO: Rome, Italy, 2018; Available online: http://www.fao.org/fileadmin/templates/est/COMM_MARKETS_MONITORING/Oilcrops/Documents/Food_outlook_oilseeds/FO_Oilcrops.pdf (accessed on 25 October 2020).
- Wang, L.; Xu, Q.; Yu, H.; Ma, H.; Li, X.; Yang, J.; Chu, J.; Xie, Q.; Wang, Y.; Smith, S.M.; et al. Strigolactone and Karrikin Signaling Pathways Elicit Ubiquitination and Proteolysis of SMXL2 to Regulate Hypocotyl Elongation in Arabidopsis thaliana. Plant Cell 2020, 32, 2251–2270. [Google Scholar] [CrossRef] [PubMed]
- Van Staden, J.; Sparg, S.G.; Kulkarni, M.G.; Light, M.E. Post-germination effects of the smoke-derived compound3-methyl-2H-furo[2,3-c]pyran-2-one, and its potential as a preconditioning agent. Field Crops Res. 2006, 98, 98–105. [Google Scholar] [CrossRef]
- Folta, K.M.; Spalding, E.P. Unexpected roles for cryptochrome 2 and phototropin revealed by high-resolution analysis of blue light-mediated hypocotyl growth inhibition. Plant J. 2001, 26, 471–478. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antala, M.; Brestic, M. Karrikins Reduce the Hypocotyl Length of Rapeseed (Brassica napus napus L.) under Continuous Red Light. Biol. Life Sci. Forum 2021, 4, 19. https://doi.org/10.3390/IECPS2020-08777
Antala M, Brestic M. Karrikins Reduce the Hypocotyl Length of Rapeseed (Brassica napus napus L.) under Continuous Red Light. Biology and Life Sciences Forum. 2021; 4(1):19. https://doi.org/10.3390/IECPS2020-08777
Chicago/Turabian StyleAntala, Michal, and Marian Brestic. 2021. "Karrikins Reduce the Hypocotyl Length of Rapeseed (Brassica napus napus L.) under Continuous Red Light" Biology and Life Sciences Forum 4, no. 1: 19. https://doi.org/10.3390/IECPS2020-08777
APA StyleAntala, M., & Brestic, M. (2021). Karrikins Reduce the Hypocotyl Length of Rapeseed (Brassica napus napus L.) under Continuous Red Light. Biology and Life Sciences Forum, 4(1), 19. https://doi.org/10.3390/IECPS2020-08777