Abstract
Macroalgae and microalgae are two sources of natural ingredients that are of interest for industrial sectors because of their multiple potential applications. Algal extracts are known to be rich in valuable compounds such as fatty acids, minerals, and secondary metabolites, among others. Bioactivities disclosed for these biocompounds include their antifungal capacity via the growth inhibition of foodborne pathogens and spoilage microorganisms such as Aspergillus sp., Fusarium sp., or Penicillium sp. Apart from their antifungal activity, algal extracts can minimize the toxicity of mycotoxins via two molecular mechanisms: their antioxidant and their chelating properties. Antioxidants may reduce the toxic effect of mycotoxins whereas algae bioadsorption, due to their rich composition in macromolecules (polysaccharides or proteins), may have the capacity to bind mycotoxins, prevent their metabolism, and facilitate their systemic release. These three mechanisms involved in the antifungal activity of micro- and macroalgal extracts will be critically assessed along this review in order to disclose their potential application as key tools in the food industry and public health. The use of algae extracts as antifungals and detoxifiers may underline their use of natural additives with no side effects associated that may represent an alternative to extend foodstuffs′ shelf-life and prevent the occurrence of mycotoxins, especially aflatoxin B1, aflatoxin M1, and ochratoxin A. Therefore, this review is aimed to evaluate the multiple benefits that algal extracts may provide to reinforce food safety, reduce food disposal, and prompt the utilization of underused biomass such as algae.
1. Introduction
Marine macroalgae and microalgae have served as nutritional or medical ingredients for thousands of years [1]. Nowadays, science is trying to fully understand their chemical composition to optimize all their associated properties since it is well established that algae biomass is a rich and underexploited source of bioactive compounds [2]. The photosynthetic nature of both macroalgae and microalgae have attracted the attention of industrial sectors. On one hand, they can be easily harvested or cultured. On the other hand, their photosynthetic capacity points to their biochemical richness. Indeed, algae matrixes are well known to be rich in valuable compounds such as proteins, polysaccharides, fatty acids, minerals, vitamins, and secondary metabolites, such as phenolic compounds or pigments, among others [3]. This rich chemical profile is associated with a wide range of bioactivities such as antioxidant, anti-inflammatory, anti-cancer, or antifungal, among others (Figure 1) [4]. Their antifungal capacity may inhibit the growth of foodborne pathogens and spoilage microorganisms such as the mycotoxin producers Aspergillus sp., Fusarium sp., or Penicillium sp. The growth inhibition of these fungal species may have two synergic benefits: avoid food spoilage and prevent toxin exposure to mycotoxins. Among them, aflatoxins and ochratoxin A are included as some of the most relevant mycotoxins regarding food safety and regulation frameworks [5]. Apart from the antifungal capacity of algae biomolecules, they may provide two additional benefits to reduce the impact of food already contaminated with mycotoxins. On one hand they may minimize the toxicological effects of the mycotoxins via their antioxidant, anti-inflammatory, and anti-cancer properties. On the other hand, their rich content in minerals and macromolecules, such as polysaccharides or globular proteins, point them as potential detoxifier agents.
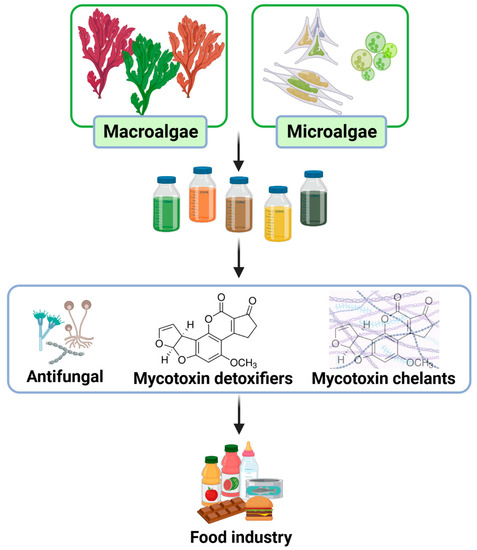
Figure 1.
Process of recovering extracts from algae as natural ingredients to incorporate into food matrixes and prevent the negative impact of mycotoxins in food industry.
Therefore, this review is aimed to present the three pathways involved in the antifungal and detoxifying capacities of algae biomolecules regarding three specific mycotoxins (aflatoxins B1, M1, and ochratoxin A) and three specific genera of fungus (Aspergillus sp., Fusarium sp., and Penicillium sp.).
2. Antifungal Activity
Different algae species have shown antifungal activity against Aspergillus sp., Fusarium sp., or Penicillium sp. (Table 1), although the inhibition growth degree is highly dependent on several factors. Among the most relevant parameters involved in the variability of the antifungal response are the fungal and algae species selected, the algae concentration and application mode, or the solvent used for the biocompounds′ extraction. Indeed, this vast variability of published results strongly hinders their comparison.

Table 1.
Summary of antifungal activities of different macroalgae and microalgae species.
Five brown algae were assessed as inhibitors of the mycelial growth of Aspergillus flavus. The dried biomass of Sargassum despiense, Turbinaria decurrens, Dilophus ligulatus, Cystoseira myrica, and Padina pavonica was added into the culture medium at six different concentrations and were tested (10, 30, 50, 100, 150, and 200 g/mL). The most effective species were T. decurrense, D. ligulatus, and P. pavonia. Indeed, the use of the essential oils extracted from these three algae (0.2 mg/mL) fully inhibited the growth of Aspergillus flavus and also the formation of aflatoxin. Their unsaponifiable matter (0.2 mg/mL) showed a 6-day delay of the mycelial growth, and even though it did not totally inhibit it, when it was further extended, it depressed the aflatoxin formation at rates higher than 70%. Finally, even though the inhibition capacity of fatty acids (0.2 mg/mL) was less aggressive with rates between 25 and 37%, the reduction in aflatoxin formation reached values higher than 85% [11]. Later, published works showed results using at least one of these previously tested species. Although, the results are based on the utilization of algae extracts instead of the addition of the algae powder into the culture medium. For instance, the aqueous extracts of Halimeda opuntia, Jania rubens, and T. decurrens was prepared at a concentration of 100 mg/mL. All three extracts showed effective growth suppression against Aspergillus parasiticus and A. flavus with percentages of inhibition that oscillated around 76 to 78%. In addition, all three extracts showed very relevant detoxifying properties displaying the total or nearly total (>90%) elimination of aflatoxins [8]. Another work used two same algae species but different organic solvents and an extract concentration. Authors used petroleum ether for extracting biomolecules from H. opuntia and T. decurrens and ethyl acetate for extracting Padina pavonica. All three extracts were finally suspended in dimethyl sulfoxide (DMSO) at 50 mg/mL, although different concentrations were applied for each assay. The H. opuntia extract at 3 mg/mL was the most promising one since it retarded the most for the fungal growth and showed inhibition areas of 16 mm for A. flavus and 22 mm for A. niger, while the respective inhibition zones that P. pavonica displayed were 9 and 7 mm, and those of T. decurrens were 10 mm for both fungi [9]. Other effective antifungal algae species is Sargassum muticum. Its methanolic extract at 300 mg/mL showed inhibition growth of four pathogenic fungi: Fusarium moniliforme (30 mm), Pythium ultimum (26 mm), A. flavus (24 mm), and Macrophomina phaseolina (23 mm) [12].
Apart from macroalgae, microalgae were also demonstrated to be effective antifungal agents, such as the biocompounds present in Nannochloropsis sp. and Spirulina sp. Free phenolic acids, with the chlorogenic acid as the most abundant representative, displayed an efficient reduction in mycelial growth of three Fusarium species: F. graminearum, F. meridionale, and F. asiaticum. The growth of Fusarium complex fungal pathogens was effectively reduced when using free phenolic acids of Spirulina sp. at a rate of 0.5 cm/day (EC50 of 34 μg/mL), and at a rate of 0.8 cm/day for Nannochloropsis sp. (EC50 of 50 μg/mL) [13]. Similarly, a methanolic extract of Spirulina sp. in a 1:5 (w:v) ratio was tested as an antifungal agent against Penicillium verrucosum. The phenolic content, mainly represented by gallic and caffeic acids, inhibited the growth of P. verrucosum mycelia from the first day and reached its maximal value (20%) at the ninth day of incubation. Even though this result was not too high, it was higher than the one reached via calcium propionate (16%). Indeed, Spirulina phenolic extract was more effective than this chemical preservative since it did not inhibit P. verrucosum growth until the fifth incubation day. In addition, phenolic compounds from Spirulina maintained the ochratoxin concentration under 0.5 ng/g for five days and reduced its presence in 56% and 29% at the seventh and ninth day against the control, respectively [14].
3. Detoxifying Bioactivities
The restoration capacity of algae biocompounds has been demonstrated in vivo against different mycotoxins (Table 1). Along this section, we present an example of the assessment of the restoration of three different animal models when applying diverse algae extracts as co-treatments of aflatoxins B1, M1, or ochratoxin A. Despite this vast variability in the experimental approaches, results are mainly coincident in regard to the restoration parameters (Table 1).
Ulva prolifera extracts were obtained via an aqueous enzymatic treatment and were analytically determined to have a minimal content of 48% of polysaccharides with a MW around 5000 Da. The most abundant representatives include rhamnose, glucuronic acid, glucose, galactose, and xylose. The co-administration of 2500 mg/kg of U. prolifera polysaccharides and 100 μg/kg of aflatoxin B1 to male broilers improved the reduction in weight that the bursa of Fabricius had suffered in animals only treated with aflatoxin B1 and restored the apoptotic cells found at this immune organ. Regarding the molecular pathways, the administration of U. prolifera polysaccharides reversed the disequilibrium caused by aflatoxin B1 both at the Nrf2 and the mitochondrial apoptotic signaling pathways. Therefore, U. prolifera polysaccharides upregulated the activities and expression of antioxidant enzymes (superoxide dismutase, catalase, or glutathione peroxidase) and the associated proteins (Nrf2, HO-1, p38MAPK, and Bcl-2) while downregulating the expression of proteins associated with apoptosis (caspase-3 and Bax) [6].
The aqueous extract obtained from Amphora cofeaeformis was evaluated as antifungal but also by its capacity to revert the toxic effect of aflatoxin M1. A. cofeaeformis extracts displayed antifungal capacity against all tested fungi, but it was stronger against Fusarium culmorum and F. graminearum than against Aspergillus ochraceus and Aspergillus flavus. In addition, in this work, male albino rats treated with aflatoxin M1 (80 μg/kg) and A. cofeaeformis extracts (20 mg/kg) reverted many of the negative effects exerted by aflatoxin M1. For instance, the use of A. cofeaeformis extracts ameliorated the toxic effects that aflatoxin M1 had on feed intake, body weight, and relative liver weight. In a similar way, the liver and kidney in animals treated with aflatoxin M1 showed strong alterations along the specific indicators (aspartate and alanine transaminases, urea, and creatinine). Animals co-treated with A. cofeaeformis extracts did not recover the normality in these indicator levels, but they were deeply amortiguated. In addition, as it was described for broilers treated with U. prolifera, rats treated with A. cofeaeformis showed an amelioration on the aflatoxin M1-affected levels of glutathione transferase, catalase, and super oxidase dismutase, and a decrease in the levels of malondialdehyde. Finally, at the histopathological level, the aflatoxin M1 tissue affections (hepatocytes-necrosis foci and congested blood sinusoids) were nearly reversed in animals co-treated with A. cofeaeformis extracts [15].
The efficiency of the pigment astaxanthin obtained from Haematococcus pluvialis was tested against the ochratoxin A-induced renal oxidative stress and its mechanism of action. The animal model used, C57BL/J mice, was treated with 100 mg/kg/day of astaxanthin, and, 2 h later, with ochratoxin A at 5 mg/kg/day. The reduction in the body weight induced by ochratoxin A in mice was restored in the group treated with both ochratoxin A and astaxanthin. This group also showed a reduction in the levels of the specific renal markers, serum uric acid, and blood urea nitrogen that ochratoxin A had previously raised. Along the histological results, the tissular anatomy was restored in the group treated with ochratoxin A and astaxanthin with no glomerular swelling, no renal cystic space visible, no cellular degeneration in the tubular epithelium, and decreased necrosis. Similarly, at the cellular level, the disorder that ochratoxin A caused in the mitochondrial cristae and nucleus were not detected when astaxanthin was also administered. In terms of apoptotic markers, the levels augmented by ochratoxin A were significantly reduced in the group additionally treated with astaxanthin. In this sense, and as observed in the other animal models, the levels of the antioxidant enzymes (superoxide dismutase, catalase, or glutathione peroxidase) were reduced while malondialdehyde was increased due to the administration of ochratoxin A. However, the additional administration of astaxanthin reversed these responses. Finally, and as previously pointed out, the binomial treatment of the mycotoxin with an antioxidant booster normalized the effects that the unique treatment with ochratoxin A had altered. Indeed, astaxanthin triggered the upregulation of the mRNA levels of the NRF2 signaling pathway and downregulated those of KEAP1 [16].
4. Physical Detoxification
Algae are a well-known source of minerals and minerals are well-known adsorbent agents. Among the most studied marine organisms, diatoms (Diatomeae) stand out due to their unique properties. These marine unicellular microalgae have silicon dioxide (SiO2) shells that create cavities and canals that provide them with high porosity. Even though diatoms have a low cation-exchange capacity of 42.75 mEq/100, their small particle size (<12 μm for 95% of total mass) confer them with high adsorption rates. Based on these features, they have been widely applied as natural adsorbent agents with detoxifying capabilities against mycotoxins. For instance, when diatoms were used as adsorbent agents for aflatoxins, they showed an index of 95% independently of the pH used. However, for ochratoxin A, this index was reduced to 67%, and it just adsorbed the toxins when tested under acid conditions (pH 3) [18]. Although, diatoms have been further tested as adsorbent agents while other species are being prepared to be tested. That is the case of Eucheuma cottonii, one of the main industrial sources of κ-carrageenan. Therefore, their remaining residues, which are mainly composed of insoluble dietary fiber, are considered to be reused for different applications (Table 1). In this work, residues of E. cottonii were finely pulverized at different particle sizes (from ~45 µm to ~420 µm) and tested as adsorbent agents against aflatoxin B1. Adsorption rates fluctuated between 62% and 88% depending on the particle size. The maximal adsorption was obtained with ~150 µm E. cottonii, and so further assessments were performed using different concentrations of the algae and toxins, incubation times, temperatures, and pH conditions. The best adsorption rate for 2 μg/mL of aflatoxin B1 was obtained with E. cottonii of a ~150 µm particle size at a concentration of 20 mg/mL, which was incubated for 15 min at a pH of 7 and a temperature of 25 °C [10]. Similarly, the seaweed Lithothamnium calcareum was demonstrated to have an adsorption capacity for aflatoxin B1 (Table 1). In this work, two approaches were followed. Firstly, when the toxin concentration was fixed (1 μg/mL), a linear pattern was observed. Higher algal concentrations triggered higher adsorption rates independently of the pH (2 mg/mL of L. calcareum absorbed 77% of aflatoxin B1). Secondly, the concentration of aflatoxin B1 was determined based on the amount of seaweed (μg toxin per mg L. calcareum). In this scenario, the lowest seaweed content of 0.5 mg/mL provided the maximal adsorption with a value of 0.78 μg/mg at pH 6 and a value of 0.62 μg/mg at pH 3 [7]. Finally, a recent work has suggested a synergic solution to remove Chlorella sp. from water by using the filamentous of Aspergillus niger as coagulants (Table 1). The adhesion of algal and fungi cells was induced via electrostatic forces that lead to a flocculation rate of 70–80%. This removal of microalgae from water bodies may represent an alternative and double solution to prevent eutrophic issues, but also to flocculate mycotoxic fungi from drinking waters via simple precipitation [17].
In addition, algae are well-known sources of macromolecules such as proteins or polysaccharides. Although β-D-glucans had been suggested to interact and bind mycotoxins, those from a yeast origin have mainly tested, whereas no algae results have been found so far [19]. However, other molecules obtained from algae have been pointed out as potential binders of mycotoxins. Their main role as binders was the reduction in the mycotoxins’ bioavailability and bioaccessibility, and so they hinder their systemic absorption. In a previously cited work, an extract of A. cofeaeformis that contained at least nine phenolic compounds with catechin and p-coumaric as the major representatives was used to fortify milk. Authors suggest that the A. cofeaeformis-fortified milk may trap aflatoxins, which would limit the toxin′s bioavailability (Table 1) [15]. Another report pointed to chlorophyll and chlorophyllin as two natural inhibitors of the intake of aflatoxin B1. Indeed, their administration provides chemoprotective effects in the rat liver and colon exposed to aflatoxin B1. One of the mechanisms described to be involved is the physical blockage that chlorophyllin induced by forming tight complexes with aflatoxin B1. This scaffold was suggested to reduce the bioavailability and to minimize the tumor incidence [20].
5. Conclusions
Nowadays, customers ask for a stronger presence of natural additives and preservatives in commercial products due to the side effects associated with chemical molecules. The food industry is continuously seeking for innovative and natural compounds that may serve as alternative food preservatives and offer a better safety profile. In this context, macroalgae and microalgae represent two underused, sustainable, and natural matrixes to obtain a huge variability of molecules with already demonstrated bioactivities. Among them, the antimicrobial capacity has been repeatedly tested, although their specific utilization as antifungals for combatting mycotoxins has not been much exploited. Nevertheless, algae have been proven to represent a real approach to prevent the growth of Aspergillus sp., Fusarium sp., and Penicillium sp. and minimize the synthesis of aflatoxins and ochratoxin A. In addition, the inclusion of algae extract as part of animal feed has been observed to provide strong protection against the negative effects of the mycotoxins. These results seem to be reachable independently of the mycotoxin involved and the animal model tested. Finally, the application of algae as complexing agents that reduce the bioavailability and bioaccessibility was found to have positive effects both in in vitro and in vivo assays. Therefore, based on the findings, the application of algae as food or feed preservatives or additives seems to provide multiple benefits to reinforce food safety and reduce food disposal.
Author Contributions
Conceptualization, M.F.-C., P.O. and J.E.; methodology, M.F.-C. and J.E.; software, M.F.-C. and A.G.P.; formal analysis, M.F.-C.; investigation, M.F.-C., J.E., P.G.-O., A.P.-V. and P.D.; resources, M.A.P. and J.S.-G.; writing—original draft preparation, M.F.-C., J.E., P.G.-O., A.P.-V. and P.D.; writing—review and editing, M.F.-C., P.O., L.C. and H.C.; visualization, M.F.-C.; supervision, M.F.-C., M.A.P. and J.S.-G.; funding acquisition, J.S.-G. All authors have read and agreed to the published version of the manuscript.
Funding
The research leading to these results was supported by MICINN supporting the Ramón y Cajal grant for M.A. Prieto (RYC-2017-22891) and the Juan de la Cierva Incorporación for Hui Cao (IJC2020-04605-5-I); by Xunta de Galicia for the program EXCELENCIA-ED431F 2020/12, the post-doctoral grants of M. Fraga-Corral (ED481B-2019/096) and L. Cassani (ED481B-2021/152), and the pre-doctoral grants of A. González Pereira (ED481A-2019/0228) and P. Garcia-Oliveira (ED481A-2019/295). Authors are grateful to AlgaMar enterprise (www.algamar.com) and to Bio Based Industries Joint Undertaking (JU) under grant agreement No 888003 UP4HEALTH Project (H2020-BBI-JTI-2019). The JU receives support from the European Union’s Horizon 2020 research and innovation program and the Bio Based Industries Consortium. The project SYSTEMIC Knowledge hub on Nutrition and Food Security has received funding from national research funding parties in Belgium (FWO), France (INRA), Germany (BLE), Italy (MIPAAF), Latvia (IZM), Norway (RCN), Portugal (FCT), and Spain (AEI) in a joint action of JPI HDHL, JPI-OCEANS, and FACCE-JPI launched in 2019 under the ERA-NET ERA-HDHL (n° 696295).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dillehay, T.D.; Ramirez, C.; Pino, M.; Collins, M.B.; Rossen, J.; Pino-Navarro, J.D. Monte Verde: Seaweed, Food, Medicine, and the Peopling of South America. Science 2008, 320, 784–786. [Google Scholar] [CrossRef]
- Herrero, M.; del Pilar Sánchez-Camargo, A.; Cifuentes, A.; Ibáñez, E. Plants, Seaweeds, Microalgae and Food by-Products as Natural Sources of Functional Ingredients Obtained Using Pressurized Liquid Extraction and Supercritical Fluid Extraction. TrAC-Trends Anal. Chem. 2015, 71, 26–38. [Google Scholar] [CrossRef]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as Nutritional and Functional Food Sources: Revisiting Our Understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Lopez, C.; Pereira, A.G.; Lourenço-Lopes, C.; Garcia-Oliveira, P.; Cassani, L.; Fraga-Corral, M.; Prieto, M.A.; Simal-Gandara, J. Main Bioactive Phenolic Compounds in Marine Algae and Their Mechanisms of Action Supporting Potential Health Benefits. Food Chem. 2021, 341, 128262. [Google Scholar] [CrossRef] [PubMed]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide Contamination of Food-Crops with Mycotoxins: Validity of the Widely Cited ‘FAO Estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Balasubramanian, B.; Zhao, Z.-H.; Liu, W.-C. Marine Algal Polysaccharides Alleviate Aflatoxin B1-Induced Bursa of Fabricius Injury by Regulating Redox and Apoptotic Signaling Pathway in Broilers. Poult. Sci. 2021, 100, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Perali, C.; Magnoli, A.P.; Aronovich, M.; Rosa, C.A.D.R.; Cavaglieri, L.R. Lithothamnium Calcareum (Pallas) Areschoug Seaweed Adsorbs Aflatoxin B1 in Vitro and Improves Broiler Chicken’s Performance. Mycotoxin Res. 2020, 36, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Farghl, A.A.M.; El-Sheekh, M.M.; El-Shahir, A.A. Seaweed Extracts as Biological Control of Aflatoxins Produced by Aspergillus Parasiticus and Aspergillus Flavus. Egypt. J. Biol. Pest Control 2023, 33, 50. [Google Scholar] [CrossRef]
- Ahmed, A.E.; AL-Kahtani, M.M.; El-Diasty, E.M.; Ahmed, A.S.; Saber, H.; Abbas, A.M.; Diab, H.M.; Alshehri, M.A.; Elmansi, A.A.; Hussein, M.A. Diversity of Toxigenic Molds and Mycotoxins Isolated from Dairy Products: Antifungal Activity of Egyptian Marine Algae on Aspergillus and Candida Species. J. Pure Appl. Microbiol. 2020, 14, 215–232. [Google Scholar] [CrossRef]
- Guo, J.; Shi, F.; Sun, M.; Ma, F.; Li, Y. Antioxidant and Aflatoxin B1 Adsorption Properties of Eucheuma Cottonii Insoluble Dietary Fiber. Food Biosci. 2022, 50, 102043. [Google Scholar] [CrossRef]
- Mabrouk, S.S.; El-Shayeb, N.M.A.; El-Refai, A.H.; Sallam, L.A.R.; Hamdy, A.A. Inhibitory Activities of Some Marine Algae on Aflatoxin Accumulation. Appl. Microbiol. Biotechnol. 1985, 22, 152–155. [Google Scholar] [CrossRef]
- Nofal, A.; Azzazy, M.; Ayyad, S.; Abdelsalm, E.; Abousekken, M.S.; Tammam, O. Evaluation of the Brown Alga, Sargassum Muticum Extract as an Antimicrobial and Feeding Additives. Braz. J. Biol. 2024, 84, e259721. [Google Scholar] [CrossRef] [PubMed]
- Scaglioni, P.T.; Pagnussatt, F.A.; Lemos, A.C.; Nicolli, C.P.; Del Ponte, E.M.; Badiale-Furlong, E. Nannochloropsis Sp. and Spirulina Sp. as a Source of Antifungal Compounds to Mitigate Contamination by Fusarium Graminearum Species Complex. Curr. Microbiol. 2019, 76, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Christ-Ribeiro, A.; Graça, C.S.; Kupski, L.; Badiale-Furlong, E.; de Souza-Soares, L.A. Cytotoxicity, Antifungal and Anti Mycotoxins Effects of Phenolic Compounds from Fermented Rice Bran and Spirulina Sp. Process Biochem. 2019, 80, 190–196. [Google Scholar] [CrossRef]
- Badr, A.N.; Abdel-Salam, A.M.; Zaghloul, A.H.; Farrag, A.R.H. Fortified Milk-Beverage with Amphora Algae and Its Functionality for Aflatoxin Inactivation in Rats. J. Food Meas. Charact. 2023, 17, 2340–2352. [Google Scholar] [CrossRef]
- Li, L.; Chen, Y.; Jiao, D.; Yang, S.; Li, L.; Li, P. Protective Effect of Astaxanthin on Ochratoxin A-Induced Kidney Injury to Mice by Regulating Oxidative Stress-Related NRF2/KEAP1 Pathway. Molecules 2020, 25, 1386. [Google Scholar] [CrossRef] [PubMed]
- Takáčová, A.; Bajuszová, M.; Šimonovičová, A.; Šutý, Š.; Nosalj, S. Biocoagulation of Dried Algae Chlorella Sp. and Pellets of Aspergillus Niger in Decontamination Process of Wastewater, as a Presumed Source of Biofuel. J. Fungi 2022, 8, 1282. [Google Scholar] [CrossRef] [PubMed]
- Bočarov-Stančić, A.; Adamović, M.; Salma, N.; Bodroža-Solarov, M.; Vučković, J.; Pantić, V. In Vitro Efficacy of Mycotoxins Adsorption by Natural Mineral Adsorbents. Biotechnol. Anim. Husb. 2011, 27, 1241–1251. [Google Scholar] [CrossRef]
- Yiannikouris, A.; André, G.; Poughon, L.; François, J.; Dussap, C.-G.; Jeminet, G.; Bertin, G.; Jouany, J.-P. Chemical and Conformational Study of the Interactions Involved in Mycotoxin Complexation with β-d-Glucans. Biomacromolecules 2006, 7, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Simonich, M.T.; Egner, P.A.; Roebuck, B.D.; Orner, G.A.; Jubert, C.; Pereira, C.; Groopman, J.D.; Kensler, T.W.; Dashwood, R.H.; Williams, D.E.; et al. Natural Chlorophyll Inhibits Aflatoxin B 1 -Induced Multi-Organ Carcinogenesis in the Rat. Carcinogenesis 2007, 28, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).