
Effect of Land-Use Change on Arbuscular Mycorrhizal Fungi Diversity in an Argentinean Endemic Native Forest †

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. AMF Diversity
2.4. Soil Analysis: Physical and Chemical Properties
2.5. Statistical Analysis
3. Results
3.1. AMF Diversity
3.2. Soil Analysis
3.3. Cluster Analysis
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, S.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Miller, R.M.; Jastrow, J.D. Extraradical hyphal development of vesicular-arbuscular mycorrhizal fungi in a chronosequence of prairie restorations. In Mycorrhizus in Ecosystems; Read, D.J., Lewis, D.H., Fitter, A.H., Alexander, I.J., Eds.; CAB International: Wallingford, UK, 1992; pp. 171–176. [Google Scholar]
- Leake, J.; Johnson, D.; Donnelly, D.; Muckle, G.; Boddy, L.; Read, D. Networks of power and influence: The role of mycorrhizal mycelium in controlling plant communities and agroecosystem functioning. Can. J. Bot. 2004, 82, 1016–1045. [Google Scholar] [CrossRef]
- Oehl, F.; Sieverding, E.; Ineichen, K.; Mäder, P.; Wiemken, A.; Boller, T. Distinct sporulation dynamics of arbuscular mycorrhizal fungal communities from different agroecosystems in long-term microcosms. Agric. Ecosyst. Environ. 2009, 134, 257–268. [Google Scholar] [CrossRef]
- Cabrera, A.L. Regiones Fitogeograficas Argentinas. Enciclopedia Argentina de Agricultura y Jardinería; ACME: Buenos Aires, Argentina, 1976; Volume 2. [Google Scholar]
- Bogino, S.; Roa-Giménez, S.C.; Velasco-Sastre, A.T.; Cangiano, M.L.; Risio-Allione, L.; Rozas, V. Synergetic effects of fire, climate, and management history on Prosopis caldenia recruitment in the Argentinean pampas. J. Arid. Environ. 2015, 117, 59–66. [Google Scholar] [CrossRef]
- SAyDS. Primer Inventario Nacional de Bosques Nativos: Informe Regional Espinal, Segunda Parte; SAyDS: Buenos Aires, Argentina, 2007. [Google Scholar]
- Anderson, D.L.; Del Aguila, J.A.; Bernardón, A.E. Las formaciones vegetales en la provincia de San Luis. Rev. Investig. Agropecu. 1970, 7, 153–183. [Google Scholar]
- Risio, L.; Herrero, C.; Bogino, S.M.; Bravo, F. Aboveground and belowground biomass allocation in native Prosopis caldenia Burkart secondaries woodlands in the semi-arid Argentinean pampas. Biomass Bioenergy 2014, 66, 249–260. [Google Scholar] [CrossRef]
- Gerdemann, J.W.; Nicolson, T.H. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc. 1963, 46, 235–244. [Google Scholar] [CrossRef]
- Walker, C.; Mize, C.W.; Mcnabb, H.S. Populations of endogonaceous fungi at two locations in central Iowa. Can. J. Bot. 1982, 60, 2518–2529. [Google Scholar] [CrossRef]
- Schenck, N.C.; Perez, Y. Manual for the Identification of VA-Mycorrhizal Fungi, 3rd ed.; Synergistic Publications: Gainesville, FL, USA, 1990. [Google Scholar]
- Błaszkowski, J. Glomeromycota; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2012. [Google Scholar]
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U.; Bahram, M.; Döring, M.; Schigel, D.; May, T.; Ryberg, M.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [Google Scholar] [CrossRef] [Green Version]
- Echenique, V.; Pessino, S.; Díaz, M.; Selva, J.P.; Luciani, G.; Zappacosta, D.; Cervigni, G.; Meier, M.; Garbus, I.; Cardone, S.; et al. Aportes de la biotecnología al mejoramiento del pasto llorón (Eragrostis curvula). Rev. Argent. Prod. Anim. 2008, 28, 147–164. [Google Scholar] [CrossRef] [Green Version]
- Azcarate, M.; Baglioni, M.; Brambilla, C.; Brambilla, E.; Fernandez, R.; Noellemeyer, E.; Ostinelli, M.; Perez, M.; Quiroga, A.; Savio, M.; et al. Métodos de análisis e implementación de Calidad en el Laboratorio de Suelos. 2017. Available online: https://inta.gob.ar/sites/default/files/inta_pt_106_kloster.pdf (accessed on 1 December 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Chagnon, P.L.; Bradley, R.L.; Maherali, H.; Klironomos, J.N. A trait-based framework to understand life history of mycorrhizal fungi. Trends Plant Sci. 2013, 18, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Longo, S.; Cofré, N.; Soteras, F.; Grilli, G.; Lugo, M.; Urcelay, C. Taxonomic and Functional Response of Arbuscular Mycorrhizal Fungi to Land Use Change in Central Argentina. In Recent Advances on Mycorrhizal Fungi; Springer: Cham, Switzerland, 2016; pp. 81–90. [Google Scholar]
- Weber, S.E.; Diez, J.M.; Andrews, L.V.; Goulden, M.L.; Aronson, E.L.; Allen, M.F. Responses of arbuscular mycorrhizal fungi to multiple coinciding global change drivers. Fungal Ecol. 2019, 40, 62–71. [Google Scholar] [CrossRef]
- Waldrop, M.P.; Zak, D.R.; Blackwood, C.B.; Curtis, C.D.; Tilman, D. Resource availability controls fungal diversity across a plant diversity gradient. Ecol. Lett. 2006, 9, 1127–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ontivero, R.E.; Voyron, S.; Risio Allione, L.V.; Bianco, P.; Bianciotto, V.; Iriarte, H.J.; Lugo, M.A.; Lumini, E. Impact of land use history on the arbuscular mycorrhizal fungal diversity in arid soils of Argentinean farming fields. FEMS Microbiol. Lett. 2020, 367, 1–11. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
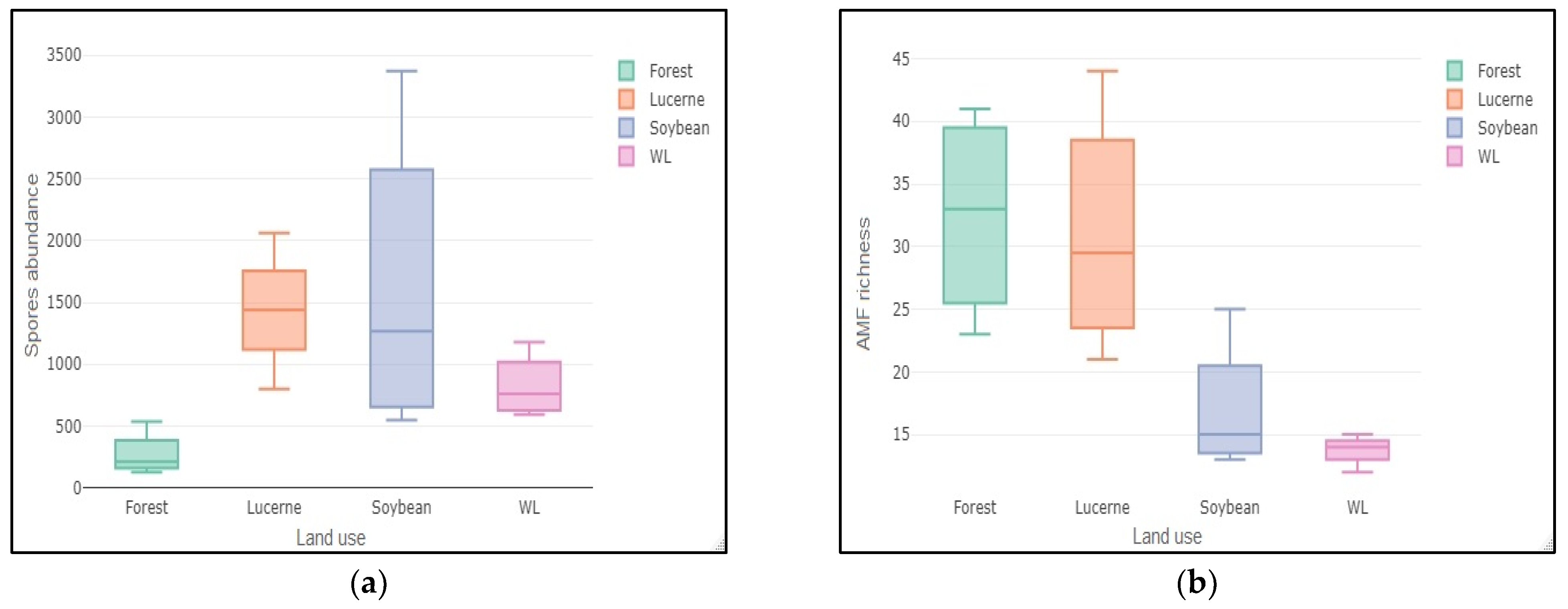
| Variable | Forest | Weeping Lovegrass | Lucerne | Soybean |
|---|---|---|---|---|
| Richness | 32.5 ± 4.21 a | 13.65 ± 0.63 b | 31 ± 4.98 ac | 17 ± 2.74 bc |
| Abundance | 271.5 ± 90.77 a | 822 ± 131.23 ab | 1435.25 ± 257.43 b | 1613.25 ± 644.23 b |
| Variable | Forest | Weeping Lovegrass | Lucerne | Soybean |
|---|---|---|---|---|
| Carbon (%) | 0.026 ± 0.004 a | 0.026 ± 0.003 a | 0.015 ± 0.001 b | 0.017 ± 0.002 ab |
| Phosphorous (g/kg) | 33.05 ± 0.77 a | 25.05 ± 1.27 ab | 23.33 ± 0.54 b | 24.33 ± 1.52 b |
| Nitrogenous (mg/g) | 0.066 ± 0.0020 a | 0.064 ± 0.0031 a | 0.068 ± 0.0024 a | 0.071 ± 0.0034 a |
| Electrical Conductivity (dS/ms) | 0.31 ± 0.02 a | 0.29 ± 0.02 a | 0.26 ± 0.01 a | 0.29 ± 0.01 a |
| Organic Matter (%) | 1.40 ± 0.20 a | 1.05 ± 0.08 a | 0.90 ± 0.05 ab | 0.80 ± 0.03 b |
| pH | 6.42 ± 0.17 a | 6.18 ± 0.08 a | 5.95 ± 0.11 a | 5.91 ± 0.06 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ontivero, R.E.; Risio, L.V.; Iriarte, H.J.; Lugo, M.A. Effect of Land-Use Change on Arbuscular Mycorrhizal Fungi Diversity in an Argentinean Endemic Native Forest. Biol. Life Sci. Forum 2022, 15, 15. https://doi.org/10.3390/IECD2022-12430
Ontivero RE, Risio LV, Iriarte HJ, Lugo MA. Effect of Land-Use Change on Arbuscular Mycorrhizal Fungi Diversity in an Argentinean Endemic Native Forest. Biology and Life Sciences Forum. 2022; 15(1):15. https://doi.org/10.3390/IECD2022-12430
Chicago/Turabian StyleOntivero, Roberto Emanuel, Lucía V. Risio, Hebe J. Iriarte, and Mónica A. Lugo. 2022. "Effect of Land-Use Change on Arbuscular Mycorrhizal Fungi Diversity in an Argentinean Endemic Native Forest" Biology and Life Sciences Forum 15, no. 1: 15. https://doi.org/10.3390/IECD2022-12430
APA StyleOntivero, R. E., Risio, L. V., Iriarte, H. J., & Lugo, M. A. (2022). Effect of Land-Use Change on Arbuscular Mycorrhizal Fungi Diversity in an Argentinean Endemic Native Forest. Biology and Life Sciences Forum, 15(1), 15. https://doi.org/10.3390/IECD2022-12430