1. Introduction
Nowadays, chronic inflammatory diseases have been clearly recognized as the most critical cause of death in the world, with more than 50% of all deaths being attributable to stroke, cancer, ischemic heart disease, diabetes mellitus, chronic kidney disease, non-alcoholic fatty liver disease, and auto-immune and neurodegenerative diseases [
1]. Therefore, the development of new and alternative preventive and therapeutic strategies against infection, inflammation, and oxidative stress is being explored. Among the multiple medicinal options, bioactive peptides have emerged as one of the most promising.
Bioactive or functional peptides are defined as inactive amino acid sequences within the precursor protein which, once released after microbial fermentation processes, chemical or enzymatic hydrolysis that takes place during food processing or gastrointestinal digestion, can exert different biological activities such as antioxidant, anticancer or antitumor, immunomodulatory, and anti-inflammatory [
2]. Regarding its origin, it should be noted that any food protein source of both vegetable or animal origin can release bioactive peptides. In the case of vegetable proteins, most studies have been focused on cereals and legumes, mainly soybean [
3].
Lunasin is a peptide present in the cotyledon of soybeans that was identified 24 years ago and is encoded by the soybean 2S albumin Gm2S-1 gene [
4]. In recent years, numerous research groups have focused their studies on the evaluation of the biological activities of lunasin, thus far demonstrating chemopreventive, antioxidant, anti-inflammatory, hypocholesterolemic, and modulating properties of the immune, nervous, and cardiovascular systems [
5]. All these properties suggest the potential of lunasin to prevent and/or treat various chronic pathologies associated with oxidative stress, inflammation, and metabolic alterations. With the aim of advancing the knowledge of the biological properties of lunasin, in this work, we will focus on the potential antioxidant and immunomodulatory activity of a lunasin-enriched soybean extract.
2. Materials and Methods
The lunasin peptide was synthesized by DGpeptides Co., Ltd. (Hangzhou, China), with a purity of 95.2%. The albumin-enriched soybean extract (ES) was provided by Reliv International, Inc. (Chesterfield, MO, USA). Lunasin-enriched soybean extract (LES) was obtained from ES by aqueous solvent extraction and its protein concentration was determined by the bicinchoninic acid (BCA) method.
LES was subjected to a simulated digestion process, following the in vitro “INFOGEST” protocol [
6]. The protein profile of LES and its gastric and gastrointestinal digests (GD and GID) was analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) in the Criterion automated equipment (Bio-Rad, Hercules, CA, USA), using 12% Bis-Tris CriterionTM XT Precast Gel polyacrylamide gels (Bio-Rad).
The presence of lunasin in LES and its behavior during the digestive process were studied by Western blot, using synthetic lunasin as a standard (12.5–50 µM). SDS-PAGE was conducted with 16.5% Bis-Tris Criterion™ XT Precast gels (Bio-Rad). Electrophoretic migration was carried out at 60 V for 2 min followed by 100 V for 3 h. Polyvinylidene membranes (PVDF) (Bio-Rad) were used for 30 min transfer using a voltage of 18 V for 30 min. After blocking and washing the membrane, overnight incubation (4 °C) with the primary rabbit antibody against lunasin (Biomedal, Seville, Spain, 1:12,000) followed by mouse IgG-horseradish peroxidase (HRP)-conjugated anti-rabbit IgG secondary antibody incubation (Santa Cruz Biotechnology, Dallas, TX, USA; 1:5000, 1 h, room temperature) was carried out. The membrane was developed with Amersham reagent (Merck, Darmstadt, Germany) for 3 min and images were taken on the Versadoc Imaging System reader (Bio-Rad), using an AF 50 mm f/1 4D photographic objective (Nikon, Tokyo, Japan).
The radical scavenging activity of LES and its digests was determined by the oxygen radical scavenging method (ORAC) and the 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) (ABTS), radical scavenging assay following previously optimized protocols [
7,
8].
Mouse RAW 264.7 macrophages (American Type Culture Collection, Rockville, MD, USA) were grown in Dulbecco’s Modified Eagle’s Medium (DMEM) with high glucose content (Biowest, Riverside, MO, USA) at 37 °C under constant conditions of humidity, 5% CO
2 and 95% air. The evaluation of the effect of LES on cell viability was carried out using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazol (MTT) bromide assay [
9]. The effects on the release of nitric oxide (NO) and the phagocytic activity were measured using the Griess test and the neutral red assay, respectively [
9]. The effect of LES on the cellular production of reactive oxygen species (ROS) was determined using 2′7′-dichlorofluorescein diacetate (DCFH-DA) as a fluorescence probe [
9]. To evaluate the effect of LES on the cellular production of cytokines, ELISA assays were performed following the manufacturer’s instructions (eBioscience, Affymetrix Company, Santa Clara, CA, USA).
The results obtained were analyzed by a one-way ANOVA analysis, followed by a Bonferroni test, using the statistical analysis program GraphPad Prism 7.0 (GraphPad Software, San Diego, CA, USA).
3. Results and Discussion
The protein concentration of ES was 52.7%, higher than that determined for LES (12.6%), corresponding to the soluble proteins in the aqueous solvent used for the extraction. The SDS-PAGE analysis showed that the protein profiles of ES and LES were similar, observing bands with molecular weights between 4 and 138 kDa, although the intensity of the bands was greater for LES, indicating the enrichment of these proteins after the aqueous solvent extraction. Specifically, the presence of the major soybean proteins, β-conglycinin, glycinin and their corresponding subunits, α and α’ of β-conglycinin (82.03 kDa and 76.14 kDa, respectively), and the A1a subunits and glycinin A2 (36.11 kDa) was observed.
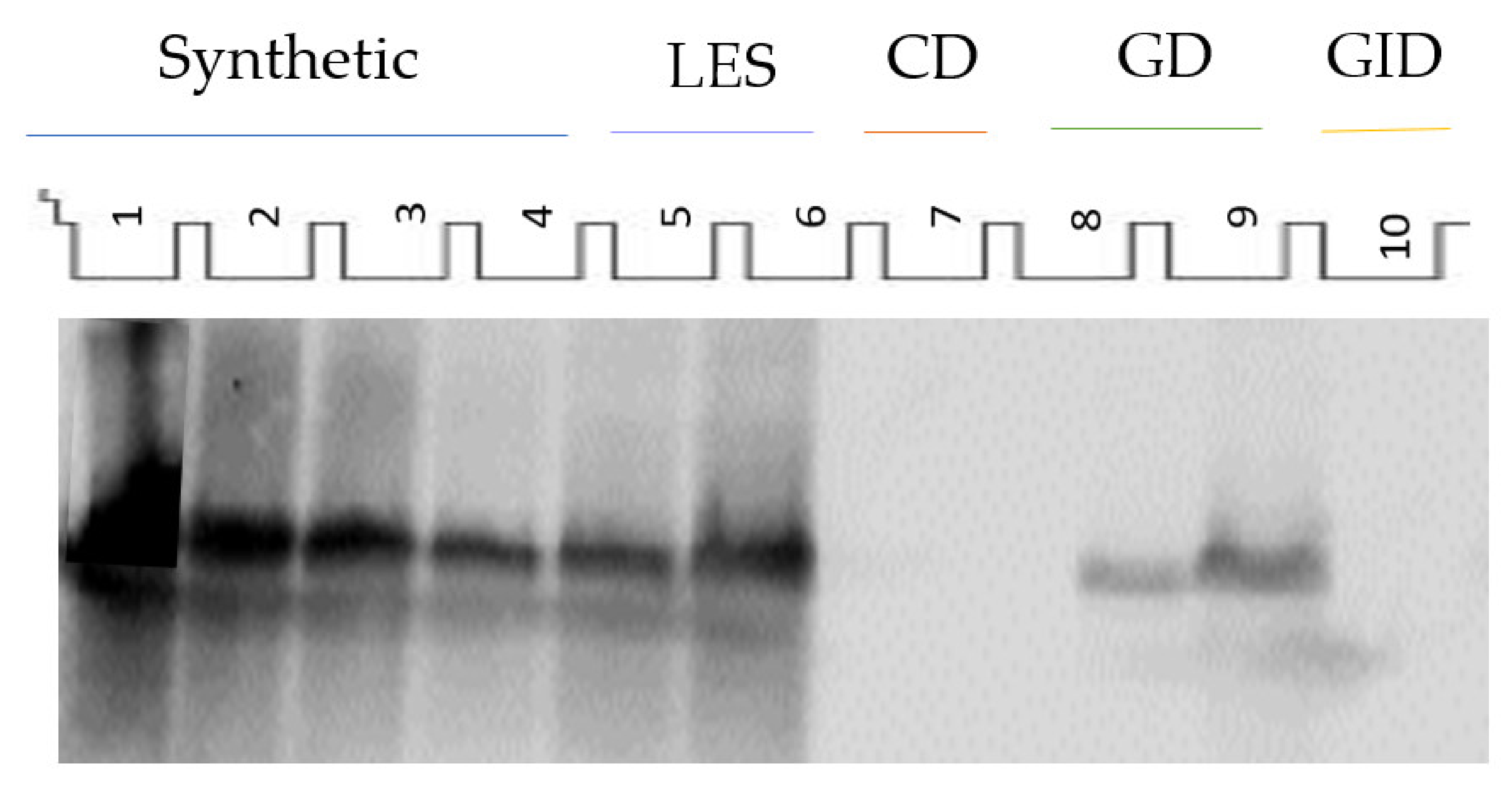
The identification of the lunasin peptide in LES was carried out by Western blot (
Figure 1). Monomeric and dimeric forms of the peptide could be identified. The lunasin concentration obtained for LES was 16.42 mg/g of protein or 2.07 mg/g of extract.
LES was subjected to a simulated gastrointestinal digestion process and its behavior was investigated by SDS-PAGE. After the gastric phase, most of the high molecular weight bands disappeared, releasing lower molecular weight proteins that were visible in the gel. However, some soybean proteins resisted the action of pepsin although they were degraded afterwards during the intestinal phase. Using Western blot, the presence of lunasin was identified in the GD, suggesting the partial resistance of this peptide to the action of pepsin (
Figure 1). Specifically, 60% of the initial lunasin was detected in GD. However, pancreatin degraded lunasin to a greater extent, and only 2.9% of the intact peptide could be visualized on the GID. The naturally occurring inhibitors potentially present in LES (Bowman–Birk inhibitor and/or Kunitz inhibitor [
10]) could be responsible for the greater resistance of lunasin to the digestive process. The intact lunasin remaining at the end of the digestive process might exert its action at a local level or be absorbed, exerting its effects at a systemic level.
The radical scavenging activity of ES, LES, GD, and GID was evaluated by ORAC and ABTS assays. LES showed higher antioxidant activity (0.23 µmol Trolox equivalent (TE)/mg protein in the case of TEAC and 1.67 µmol TE/mg protein in the case of ORAC) than ES (0.11 µmol TE/mg protein for TEAC and 0.50 µmol TE/mg protein for ORAC), indicating that soluble proteins and small peptides extracted with PBS were the main molecules responsible for the antioxidant activity observed. After gastrointestinal digestion, the antioxidant activity increased until reaching TEAC and ORAC values of 0.33 µmol and 2.17 µmol TE/mg protein, respectively, in GID.
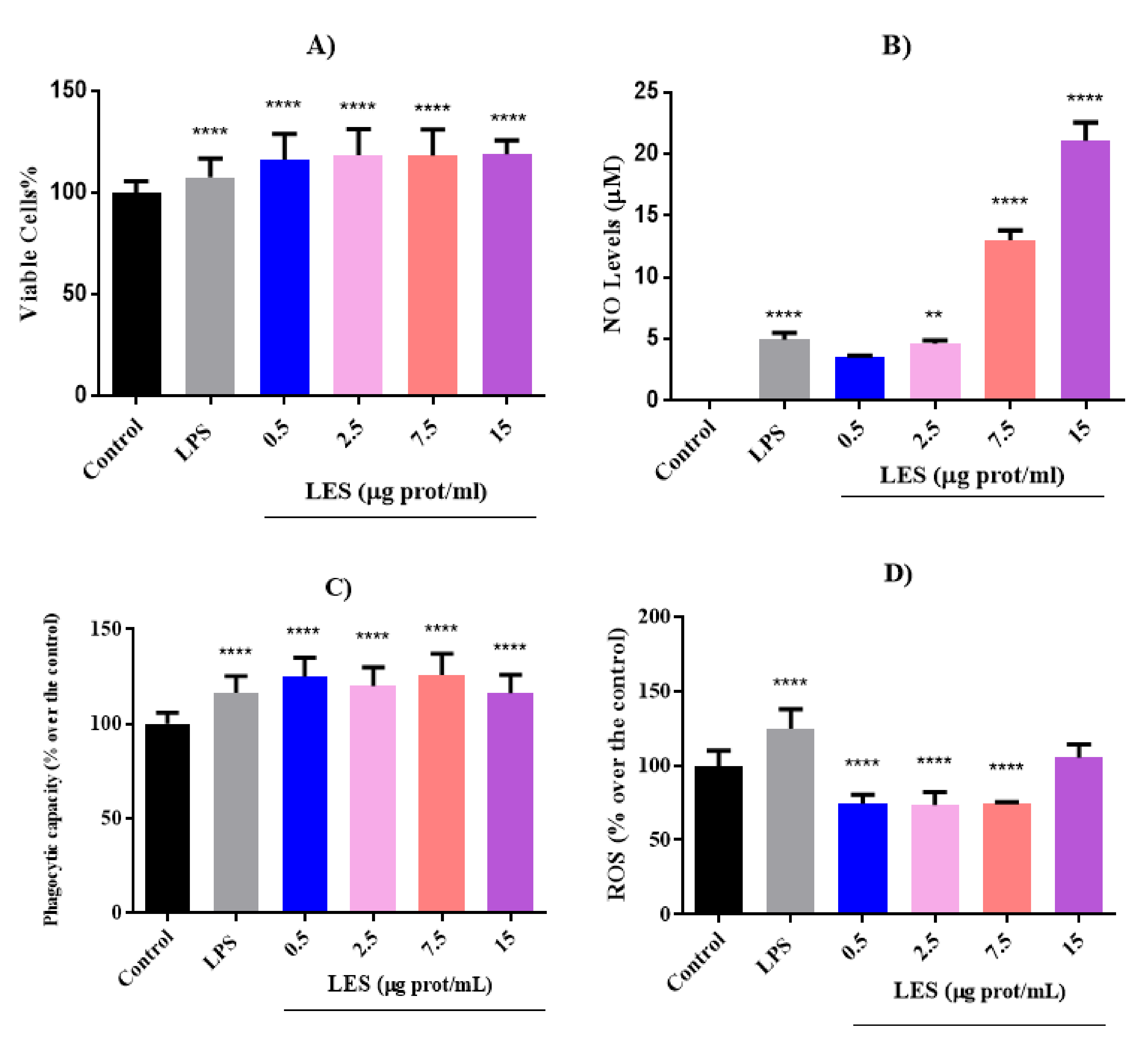
The effects of different doses of LES (ranging from 0.5 to 15 µg protein/mL) were evaluated in macrophages at different treatment times (8, 16, and 24 h). In
Figure 2, the results obtained on cell viability, ROS generation, NO levels, and phagocytic capacity of cells after 8 h treatment are shown.
As shown in
Figure 2A, LES significantly increased the viability of macrophages in a dose-dependent manner (
p < 0.0001) after 8 h of treatment. However, longer incubation times with LES caused a decrease in the percentage of viable cells, being significant for concentrations higher than 7.5 µg/mL at 16 h and 2.5 µg/mL at 24 h. LPS also caused a significant decrease in cell viability after 16 h of treatment, the percentage of viable cells being similar to that determined for the LES concentration of 7.5 µg/mL.
Under basal conditions, NO could not be detected in cell culture (
Figure 2B). However, after stimulation with LPS, NO levels increased significantly in a time-dependent manner, reaching values of 4.96, 17.47, and 34.81 µM at 8, 16, and 24 h, respectively. Similarly, LES also caused a stimulating effect on NO production in a dose- and time-dependent manner. Thus, at the maximum concentration tested of 15 µg/mL, the NO levels detected were 21.08, 43.19, and 63.40 µM at 8, 16, and 24 h of treatment. At 24 h, the stimulating effect of LES at 2.5 µg/mL was similar to that caused by LPS at 10 ng/mL. NO is a molecule produced by various cells, exerting multifunctional properties. It has been shown to act as an inflammation marker, being, in high concentrations, harmful to the body. However, macrophages release this compound as a mediator of their cytotoxic action against microorganisms, favoring their phagocytic function [
11]. Therefore, the results obtained suggest that the immunostimulatory effect of LES in macrophages, by increasing their production of NO, could favor their phagocytic capacity and, on the other hand, exert a vasodilator effect that would help the arrival of other cells of the immune system to the possible source of infection.
Therefore, to evaluate the effect of LES on the phagocytic capacity of macrophages, the neutral red assay was carried out at different treatment times (
Figure 2C). At 8 h, this capacity increased significantly at the four concentrations tested, being similar to the capacity determined after stimulation with LPS (
p < 0.0001). This cellular induction together with the increase in NO levels suggest the immunostimulant action of LES on macrophages, thus favoring the ability of these cells to eliminate pathogens. After increasing the treatment time up to 16 h, the phagocytic capacity was not affected by LPS nor LES, except at the concentration of 2.5 µg/mL of LES, for which a significant increase in phagocytosis was observed. At 24 h, LPS did not modify the phagocytic capacity of the cells. In the case of LES, only the lower concentration caused an increase in the capacity, while the higher concentrations caused a significant decrease in phagocytosis, which could be due to the inhibitory effect of cell viability exerted by the high concentrations of the extract after 24 h incubation with the extract.
As shown in
Figure 2D, 8 h treatment with 100 ng/mL of LPS caused an oxidative effect, resulting in a significant increase in ROS levels, which was also observed after 16 and 24 h. However, LES at concentrations of 0.5, 2.5, and 7.5 µg protein/mL exerted a protective effect against oxidative stress, decreasing ROS levels at the three times tested. This result confirms the antioxidant effect evaluated by the ORAC and ABTS assays. However, the highest concentration caused an oxidizing effect at the three times tested, being significant at 16 and 24 h of treatment. Although the oxidative stress generated could favor a more efficient destruction of microorganisms during the immune response [
11], the increase observed at long incubation times could provoke the observed reduction in macrophage viability.
Finally, LES induced, in a dose-dependent manner, the liberation of IL-6 at the highest concentration assayed. However, these levels were lower than those resulting from the treatment with LPS at 100 and 1000 ng/mL. Moreover, LES exerted a dose-dependent effect on the liberation of IL-10, and in 15 µg protein/mL, the effect was higher than that provoked by the stimulation with LPS.




{kind=link}
{kind=link}

