
Declines in Brook Trout Abundance Linked to Atmospheric Warming in Maryland, USA


Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Data
2.2. Trend Analysis
2.3. Environmental Analysis
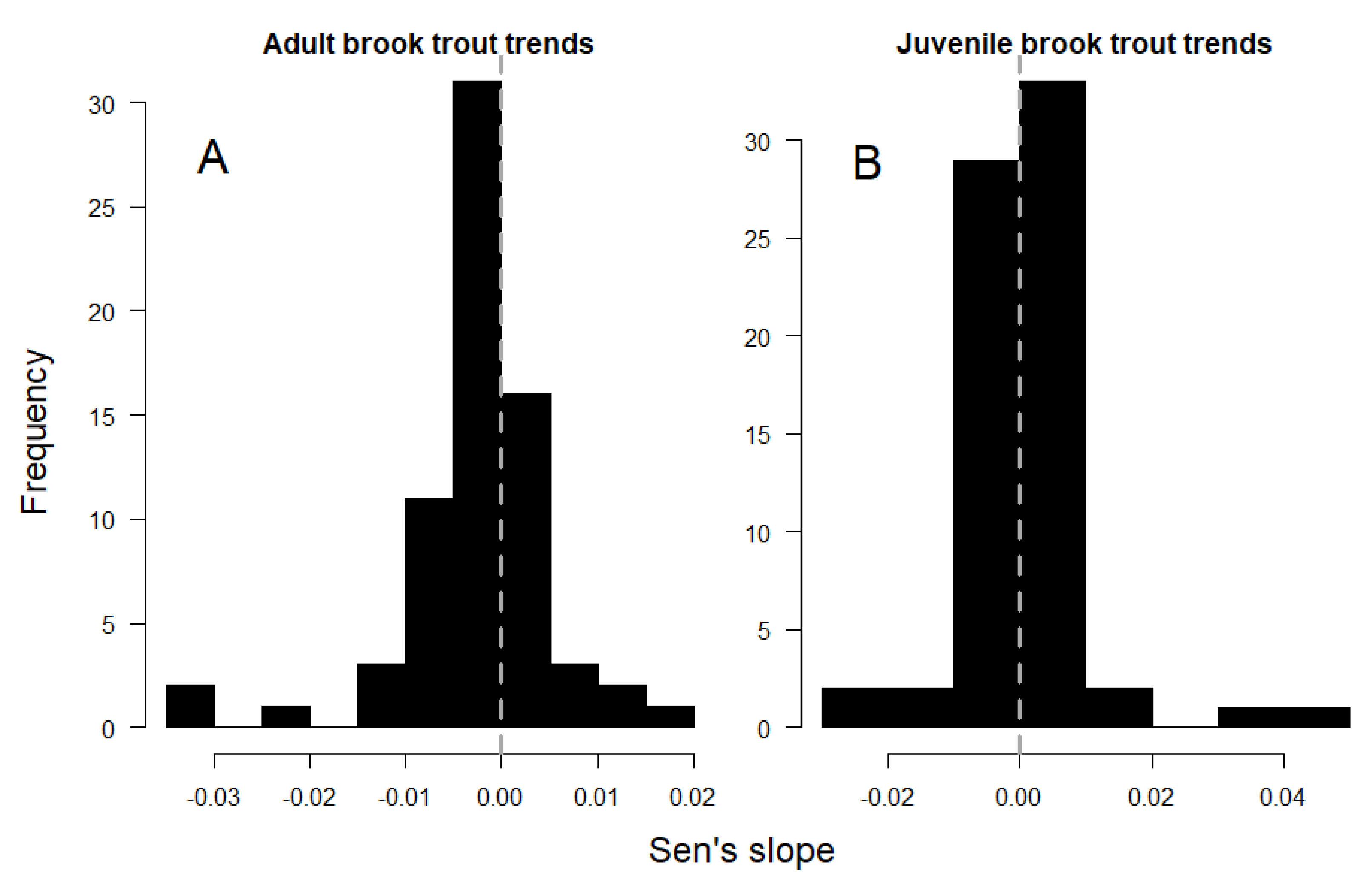
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Code | Latitude | Longitude | Adult Brook Trout Trend | Juvenile Brook Trout Trend |
|---|---|---|---|---|
| ANTI-101-S | 39.65833 | −77.5452 | −0.0023 | −0.0009 |
| ANTM6191 | 39.66278 | −77.5322 | −0.0050 | 0.0010 |
| CSLM0001 | 39.64850 | −79.1386 | 0.0167 | 0.0347 |
| CSLM0002 | 39.63892 | −79.1651 | 0.0140 | 0.0458 |
| CSLM0004 | 39.69296 | −79.1869 | −0.0084 | 0.0007 |
| CSLM0005 | 39.60816 | −79.1945 | −0.0045 | 0.0058 |
| DOBL0025 | 39.65434 | −76.9076 | −0.0053 | −0.0045 |
| LBRR5468 | 39.40120 | −76.9194 | −0.0086 | −0.0048 |
| LGNF6439 | 39.52438 | −76.5368 | 0.0012 | −0.0083 |
| LIBE-102-S | 39.44055 | −76.8641 | 0.0000 | 0.0000 |
| LOCH-120-S | 39.47878 | −76.6818 | −0.0002 | 0.0002 |
| LRRR0008 | 39.47943 | −76.6781 | −0.0029 | 0.0039 |
| LRRR2892 | 39.59946 | −76.7063 | −0.0054 | −0.0093 |
| LRRR6263 | 39.57730 | −76.6777 | −0.0113 | −0.0051 |
| PBRR0003 | 39.69090 | −76.7679 | −0.0347 | −0.0135 |
| PBRR0013 | 39.67948 | −76.7715 | −0.0118 | −0.0229 |
| PBRR0018 | 39.67653 | −76.7708 | 0.0001 | 0.0026 |
| PBRR0048 | 39.70136 | −76.8058 | 0.0068 | −0.0050 |
| PBRR1933 | 39.64534 | −76.7198 | −0.0053 | −0.0004 |
| PBRR3834 | 39.69151 | −76.7777 | −0.0018 | 0.0018 |
| PRLN-626-S | 39.54581 | −78.9055 | −0.0040 | −0.0020 |
| SAVA-204-S | 39.50378 | −79.1556 | −0.0012 | 0.0001 |
| SAVA-225-S | 39.59930 | −79.0668 | −0.0007 | 0.0000 |
| SAVA-276-S | 39.54123 | −79.2134 | −0.0037 | −0.0055 |
| SAVG0002 | 39.48595 | −79.0834 | −0.0007 | −0.0006 |
| SAVG0010 | 39.61981 | −79.1437 | −0.0010 | 0.0040 |
| SAVG0011 | 39.60364 | −79.1206 | −0.0012 | −0.0012 |
| SAVG0012 | 39.59179 | −79.1041 | −0.0109 | −0.0054 |
| SAVG0020 | 39.56443 | −79.1982 | −0.0048 | 0.0024 |
| SAVG0021 | 39.55553 | −79.1549 | 0.0015 | −0.0076 |
| SAVG0022 | 39.55001 | −79.1470 | 0.0023 | 0.0024 |
| SAVG0030 | 39.47330 | −79.1953 | −0.0050 | 0.0015 |
| SAVG0031 | 39.49378 | −79.1685 | −0.0029 | −0.0007 |
| SAVG0040 | 39.58355 | −79.1712 | 0.0020 | 0.0029 |
| SAVG0041 | 39.55862 | −79.1525 | 0.0014 | 0.0027 |
| SAVG0042 | 39.54492 | −79.1414 | −0.0033 | 0.0007 |
| SAVG0050 | 39.53464 | −79.1873 | −0.0087 | 0.0007 |
| SAVG0051 | 39.51245 | −79.1616 | −0.0065 | 0.0006 |
| SAVG0052 | 39.51356 | −79.1562 | 0.0001 | 0.0008 |
| SAVG0060 | 39.63718 | −79.0589 | −0.0003 | 0.0029 |
| SAVG0061 | 39.62556 | −79.0627 | −0.0033 | 0.0033 |
| SAVG0062 | 39.60268 | −79.0712 | 0.0001 | 0.0040 |
| SAVG0070 | 39.61250 | −79.0281 | −0.0072 | −0.0016 |
| SAVG0071 | 39.60046 | −79.0385 | −0.0075 | 0.0017 |
| SAVG0072 | 39.59710 | −79.0541 | −0.0066 | −0.0018 |
| SAVG0080 | 39.59596 | −79.0652 | −0.0002 | −0.0009 |
| SAVG0081 | 39.58618 | −79.0849 | −0.0019 | −0.0003 |
| SAVG0082 | 39.56458 | −79.1084 | −0.0009 | 0.0000 |
| SAVG0084 | 39.59995 | −79.0547 | −0.0004 | 0.0002 |
| SAVG5004 | 39.58635 | −79.0955 | −0.0238 | −0.0064 |
| UMNC0001 | 39.58006 | −77.4320 | −0.0002 | 0.0005 |
| UMNC0002 | 39.48999 | −77.4736 | 0.0000 | −0.0248 |
| UMNC1235 | 39.55769 | −77.4820 | −0.0306 | −0.0160 |
| UMNC1333 | 39.52934 | −77.4720 | 0.0020 | 0.0025 |
| UMNC2652 | 39.58222 | −77.4518 | −0.0009 | 0.0001 |
| UMNC2897 | 39.53516 | −77.4674 | −0.0022 | 0.0032 |
| UMNC2964 | 39.62983 | −77.4439 | 0.0021 | 0.0000 |
| UMNC3372 | 39.53806 | −77.4666 | 0.0038 | 0.0022 |
| UMNC3449 | 39.62125 | −77.4375 | 0.0000 | 0.0000 |
| UMNC5720 | 39.66916 | −77.4790 | 0.0004 | 0.0006 |
| UMNC5722 | 39.66115 | −77.4813 | 0.0000 | 0.0017 |
| UMNC6096 | 39.48436 | −77.4658 | −0.0006 | −0.0001 |
| UMNC6577 | 39.58553 | −77.4363 | −0.0006 | 0.0000 |
| UMNC6836 | 39.53912 | −77.4761 | 0.0055 | 0.0051 |
| UMNC6857 | 39.63344 | −77.4765 | −0.0012 | 0.0000 |
| UMON-119-S | 39.58739 | −77.4893 | −0.0031 | −0.0009 |
| UMON-288-S | 39.60939 | −77.4349 | −0.0041 | −0.0018 |
| YOGH0009 | 39.53182 | −79.4029 | 0.0111 | 0.0105 |
| YOGH0010 | 39.53203 | −79.4040 | 0.0057 | 0.0159 |
| YOUG-432-S | 39.64264 | −79.2798 | 0.0006 | −0.0003 |
References
- Hudy, M.; Thieling, T.M.; Gillespie, N.; Smith, E.P. Distribution, status, and land use characteristics of subwatersheds within the native range of brook trout in the eastern United States. N. Am. J. Fish. Manag. 2008, 28, 1069–1085. [Google Scholar] [CrossRef]
- Stranko, S.A.; Hilderbrand, R.H.; Morgan, R.P.; Staley, M.W.; Becker, A.J.; Roseberry-Lincoln, A.; Perry, E.S.; Jacobson, P.T. Brook trout declines with land cover and temperature changes in Maryland. N. Am. J. Fish. Manag. 2008, 28, 1223–1232. [Google Scholar] [CrossRef]
- Childress, E.S.; Demarest, D.E.; Wofford, J.E.B.; Hitt, N.P.; Letcher, B.H. Strong variation in brook trout trends across geology, elevation, and stream size in Shenandoah National Park. Trans. Am. Fish. Soc. 2024, 153, 250–263. [Google Scholar] [CrossRef]
- McKenna, J.E.; Johnson, J.H. Landscape models of brook trout abundance and distribution in lotic habitat with field validation. N. Am. J. Fish. Manag. 2011, 31, 742–756. [Google Scholar] [CrossRef]
- Wagner, T.; DeWeber, J.T.; Detar, J.; Kristine, D.; Sweka, J.A. Spatial and temporal dynamics in brook trout density: Implications for population monitoring. N. Am. J. Fish. Manag. 2014, 34, 258–269. [Google Scholar] [CrossRef]
- DeWeber, J.T.; Wagner, T. Predicting brook trout occurrence in stream reaches throughout their native range in the eastern United States. Trans. Am. Fish. Soc. 2015, 144, 11–24. [Google Scholar] [CrossRef]
- Kanno, Y.; Letcher, B.H.; Rosner, A.L.; O’Neil, K.P.; Nislow, K.H. Environmental factors affecting brook trout occurrence in headwater stream segments. Trans. Am. Fish. Soc. 2015, 144, 373–382. [Google Scholar] [CrossRef]
- Jin, C.; Zha, T.; Guo, X.; Li, X.; Liu, X.; Jiang, Y.; Guo, Z.; Bourque, C.P. Forest-cover-loss control on year-round river flow dynamics in the upper Saint John River (Wolastoq) basin, Northeastern North America from 2001 to 2019. J. Hydrol. 2023, 623, 129776. [Google Scholar] [CrossRef]
- Swank, W.T.; Vose, J.M.; Elliott, K.J. Long-term hydrologic and water quality responses following commercial clearcutting of mixed hardwoods on a southern Appalachian catchment. For. Ecol. Manag. 2001, 143, 163–178. [Google Scholar] [CrossRef]
- Studinski, J.M.; Hartman, K.J.; Niles, J.M.; Keyser, P. The effects of riparian forest disturbance on stream temperature, sedimentation, and morphology. Hydrobiologia 2012, 686, 107–117. [Google Scholar] [CrossRef]
- Meisner, J.D. Effect of climatic warming on the southern margins of the native range of brook trout, Salvelinus fontinalis. Can. J. Fish. Aquat. Sci. 1990, 47, 1065–1070. [Google Scholar] [CrossRef]
- Bassar, R.D.; Letcher, B.H.; Nislow, K.H.; Whiteley, A.R. Changes in seasonal climate outpace compensatory density-dependence in eastern brook trout. Glob. Chang. Biol. 2016, 22, 577–593. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Letcher, B.L.; Nislow, K.H. Context-specific influence of water temperature on brook trout growth rates in the field. Freshw. Biol. 2010, 55, 2253–2264. [Google Scholar] [CrossRef]
- Al-Chokhachy, R.; Letcher, B.H.; Muhlfeld, C.C.; Dunham, J.; Cline, T.; Hitt, N.P.; Roberts, J.J.; Schmetterling, D. Stream size, temperature and density explain body sizes of freshwater salmonids across a range of climate conditions. Can. J. Fish. Aquat. Sci. 2022, 79, 1729–1744. [Google Scholar] [CrossRef]
- Ries, R.D.; Perry, S.A. Potential effects of global climate warming on brook trout growth and prey consumption in central Appalachian streams, USA. Clim. Res. 1995, 5, 197–206. [Google Scholar] [CrossRef]
- Chadwick, J.G., Jr.; Nislow, K.H.; McCormick, S.D. Thermal onset of cellular and endocrine stress responses correspond to ecological limits in brook trout, an iconic cold-water fish. Conserv. Physio. 2015, 3, cov017. [Google Scholar] [CrossRef]
- Hitt, N.P.; Snook, E.; Massie, D. Brook trout use of thermal refugia and foraging habitat influenced by brown trout. Can. J. Fish. Aquat. Sci. 2017, 74, 406–418. [Google Scholar] [CrossRef]
- McCormick, J.H.; Hokanson, E.F.; Jones, B.R. Effects of temperature on growth and survival of young brook trout, Salvelinus fontinalis. J. Fish. Res. Board Can. 1972, 29, 1107–1112. [Google Scholar] [CrossRef]
- Kaushal, S.S.; Likens, G.E.; Jaworski, N.A.; Pace, M.L.; Sides, A.M.; Seekell, D.; Belt, K.T.; Secor, D.H.; Wingate, R.L. Rising stream and river temperatures in the United States. Front. Ecol. Environ. 2010, 8, 461–466. [Google Scholar] [CrossRef]
- Ding, H.; Elmore, A.J. Spatio-temporal patterns in water surface temperature from Landsat time series data in the Chesapeake Bay, U.S.A. Remote Sens. Environ. 2015, 168, 335–348. [Google Scholar] [CrossRef]
- Rice, K.C.; Jastram, J.D. Rising air and stream-water temperatures in Chesapeake Bay region, USA. Clim. Chang. 2015, 128, 127–138. [Google Scholar] [CrossRef]
- Hilderbrand, R.H.; Kashiwagi, M.T.; Prochaska, A.P. Regional and local scale modeling of stream temperatures and spatio-temporal variation in thermal sensitivities. Environ. Manag. 2014, 54, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Snyder, C.D.; Hitt, N.P.; Young, J.A. Accounting for the influence of groundwater on thermal sensitivity of headwater streams to climate change. Ecol. Appl. 2015, 25, 1397–1419. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, A.M.; Corey, E.; Cunjak, R.A.; Linnansaari, T.; Curry, R.A. Salmonid thermal habitat contraction in a hydrogeologically complex setting. Ecosphere 2021, 12, e03797. [Google Scholar] [CrossRef]
- Fausch, K.D.; White, R.J. Competition between brook trout (Salvelinus fontinalis) and brown trout (Salmo trutta) for positions in a Michigan stream. Can. J. Fish. Aquat. Sci. 1981, 38, 1220–1227. [Google Scholar] [CrossRef]
- Wagner, T.; DeWeber, J.T.; Detar, J.; Sweka, J.A. Landscape-scale evaluation of asymmetric interactions between brown trout and brook trout using two-species occupancy models. Trans. Am. Fish. Soc. 2013, 142, 353–361. [Google Scholar] [CrossRef]
- Waters, T.F. Replacement of brook trout by brown trout over 15 years in a Minnesota stream: Production and abundance. Trans. Am. Fish. Soc. 1983, 112, 137–146. [Google Scholar] [CrossRef]
- Hoxmeier, R.J.H.; Dieterman, D.J. Seasonal movement, growth and survival of brook trout in sympatry with brown trout in Midwestern US streams. Ecol. Freshw. Fish 2013, 22, 530–542. [Google Scholar] [CrossRef]
- McKenna, J.E.; Slattery, M.T.; Clifford, K.M. Broad-scale patterns of brook trout responses to introduced brown trout in New York. N. Am. J. Fish. Manag. 2013, 33, 1221–1235. [Google Scholar] [CrossRef]
- Kirk, M.A.; Rosswog, A.N.; Ressel, K.N.; Wissinger, S.A. Evaluating the trade-offs between invasion and isolation for native brook trout and nonnative brown trout in Pennsylvania streams. Trans. Am. Fish. Soc. 2018, 147, 806–817. [Google Scholar] [CrossRef]
- Hoxmeier, R.J.H.; Dieterman, D.J. Long-term population demographics of native brook trout following manipulative reduction of an invader. Biol. Invas. 2016, 18, 2911–2922. [Google Scholar] [CrossRef]
- Olson, K.W.; Pechacek, K.; Benike, H. Brook Trout population response to Brown Trout removal by electrofishing in a Wisconsin Driftless Area stream. N. Am. J. Fish. Manag. 2024, 44, 735–744. [Google Scholar] [CrossRef]
- Odenkirk, J.S.; Isel, M.W. Trends in biomass and relative weight of brook trout in response to introduction of non-native brown trout in an Appalachian mountain stream. J. Southeast. Assoc. Fish Wild. Agencies 2022, 9, 67–72. [Google Scholar]
- Murdoch, A.; Mantyka-Pringle, C.; Sharma, S. The interactive effects of climate change and land use on boreal stream fish communities. Sci. Total Environ. 2020, 700, 134518. [Google Scholar] [CrossRef] [PubMed]
- Carosi, A.; Lorenzoni, F.; Lorenzoni, M. Synergistic effects of climate change and alien fish invasions in freshwater ecosystems: A review. Fishes 2023, 8, 486. [Google Scholar] [CrossRef]
- Genovese, M.; Cessna, J.; Kilian, J.; Stranko, S. Spatial and Temporal Trends in Biological, Physical, and Chemical Data (1995–2020) Collected at 30 Stream Sites within the Maryland Sentinel Site Network; Maryland Department of Natural Resources: Annapolis, MD, USA, 2023; DNR-12-042823-354.
- Agarwal, S.; Suchithra, A.; Singh, S.P. Analysis and interpretation of rainfall trend using Mann-Kendall’s and Sen’s slope method. Indian J. Ecol. 2021, 48, 453–457. [Google Scholar]
- Frimpong, B.F.; Koranteng, A.; Molkenthin, F. Analysis of temperature variability utilising Mann–Kendall and Sen’s slope estimator tests in the Accra and Kumasi Metropolises in Ghana. Environ. Syst. Res. 2022, 11, 24. [Google Scholar] [CrossRef]
- Mann, H.B. Non-parametric tests against trend. Econometrica 1945, 13, 163–171. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods, 4th ed.; Charles Griffin Publishers: London, UK, 1975. [Google Scholar]
- Sen, P.K. Estimates of the regression coefficient based on Kendall’s tau. J. Am. Stat. Assoc. 1968, 63, 1379–1389. [Google Scholar] [CrossRef]
- Pickers, P.A.; Manning, A.C. Investigating bias in the application of curve fitting programs to atmospheric time series. Atmos. Meas. Tech. 2015, 8, 1469–1489. [Google Scholar] [CrossRef]
- Pohlert, T. Trend: Non-Parametric Trend Tests and Change-Point Detection. R Package Version 1.1.6. 2023. Available online: https://CRAN.R-project.org/package=trend (accessed on 1 December 2023).
- Hitt, N.P.; Floyd, M.; Compton, M.; McDonald, K. Threshold responses of blackside dace (Chrosomus cumberlandensis) and Kentucky arrow darter (Etheostoma spilotum) to stream conductivity. Southeast. Nat. 2016, 15, 41–60. [Google Scholar] [CrossRef]
- Hill, R.A.; Weber, M.H.; Leibowitz, S.G.; Olsen, A.R.; Thornbrugh, D.J. The Stream-Catchment (StreamCat) Dataset: A database of watershed metrics for the conterminous United States. J. Am. Water Res. Assoc. 2016, 52, 120–128. [Google Scholar] [CrossRef]
- Hitt, N.P.; Kessler, K.; Rogers, K.M.; Macmillan, H.; Walsh, H. Assessing Native Fish Restoration Potential in Catoctin Mountain Park; U.S. Geological Survey Open-File Report 2020-1137; U.S. Geological Survey: Reston, VA, USA, 2020.
- Thornton, P.E.; Shrestha, R.; Thornton, M.; Kao, S.C.; Wei, Y.; Wilson, B.E. Gridded daily weather data for North America with comprehensive uncertainty quantification. Sci. Data 2021, 8, 190. [Google Scholar] [CrossRef] [PubMed]
- Hufkens, K.; Basler, D.; Milliman, T.; Melaas, E.K.; Richardson, A.D. An integrated phenology modeling framework in R. Methods Ecol. Evol. 2018, 9, 1276–1285. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org (accessed on 15 January 2023).
- Utz, R.; Hartman, K. Density-dependent individual growth and size dynamics of central Appalachian brook trout (Salvelinus fontinalis). Can. J. Fish. Aquat. Sci. 2009, 66, 1072–1080. [Google Scholar] [CrossRef]
- Hitt, N.P.; Rogers, K.M.; Kessler, K.G.; Briggs, M.A.; Fair, J.H.; Dolloff, C.A. Effects of episodic dewatering on brook trout spatial population structure. Freshw. Biol. 2024, 69, 1027–1041. [Google Scholar] [CrossRef]
- Kanno, Y.; Pregler, K.C.; Hitt, N.P.; Letcher, B.H.; Hocking, D.J.; Wofford, J.E.B. Seasonal temperature and precipitation regulate brook trout young-of-the-year abundance and population dynamics. Freshw. Biol. 2016, 61, 88–99. [Google Scholar] [CrossRef]
- Pregler, K.C.; Hanks, R.D.; Childress, E.S.; Hitt, N.P.; Hocking, D.J.; Letcher, B.H.; Wagner, T.; Kanno, Y. State-space analysis of power to detect regional brook trout population trends over time. Can. J. Fish. Aquat. Sci. 2019, 76, 2145–2155. [Google Scholar] [CrossRef]
- Leung, L.R.; Terando, A.; Joseph, R.; Tselioudis, G.; Bruhwiler, L.M.; Cook, B.; Deser, C.; Hall, A.; Hamlington, B.D.; Hoell, A.; et al. Earth systems processes. In Fifth National Climate Assessment; Crimmins, A.R., Avery, C.W., Easterling, D.R., Kunkel, K.E., Stewart, B.C., Maycock, T.K., Eds.; U.S. Global Change Research Program: Washington, DC, USA, 2023. [Google Scholar]
- Menne, M.J.; Durre, I.; Vose, R.S.; Gleason, B.E.; Houston, T.G. An overview of the Global Historical Climatology Network-Daily Database. J. Atmos. Ocean. Technol. 2012, 29, 897–910. [Google Scholar] [CrossRef]
- Hare, D.K.; Helton, A.M.; Johnson, Z.C.; Lane, J.W.; Briggs, M.A. Continental-scale analysis of shallow and deep groundwater contributions to streams. Nat. Commun. 2021, 12, 1450. [Google Scholar] [CrossRef]
- Johnson, Z.C.; Briggs, M.A.; Snyder, C.D.; Johnson, B.G.; Hitt, N.P. Taking heat (downstream): Simulating groundwater and thermal equilibrium controls on annual paired air-water temperature signal transport in headwater streams. J. Hydrol. 2024, 638, 131391. [Google Scholar] [CrossRef]
- Hitt, N.P.; Rogers, K.M.; Kessler, K.G.; Briggs, M.A.; Fair, J.H. Stabilising effects of karstic groundwater on stream fish communities. Ecol. Freshw. Fish 2023, 32, 538–551. [Google Scholar] [CrossRef]
- Kessler, K.; Rogers, K.M.; Marsh, C.; Hitt, N.P. Karst terrain promotes thermal resiliency in headwater streams. Proc. West Va. Acad. Sci. 2023, 95, 1–8. [Google Scholar] [CrossRef]
- Briggs, M.A.; Goodling, P.; Johnson, Z.C.; Rogers, K.M.; Hitt, N.P.; Fair, J.B.; Snyder, C.D. Bedrock depth influences spatial patterns of summer baseflow, temperature, and flow disconnection for mountainous headwater streams. Hydrol. Earth Syst. Sci. 2022, 26, 3989–4011. [Google Scholar] [CrossRef]
- Hoess, R.; Generali, K.A.; Kuhn, J.; Geist, J. Impact of fish ponds on stream hydrology and temperature regime in the context of freshwater pearl mussel conservation. Water 2022, 14, 2490. [Google Scholar] [CrossRef]
- King, R.S.; Baker, M.E.; Whigham, D.F.; Weller, D.E.; Jordan, T.E.; Kazyak, P.F.; Hurd, M.K. Spatial considerations for linking watershed land cover to ecological indicators in streams. Ecol. Appl. 2005, 15, 137–153. [Google Scholar] [CrossRef]
- Peterson, E.E.; Sheldon, F.; Darnell, R.; Bunn, S.E.; Harch, B.D. A comparison of spatially explicit landscape representation methods and their relationship to stream condition. Freshw. Biol. 2011, 56, 590–610. [Google Scholar] [CrossRef]
- Strange, R.J.; Schreck, C.B.; Golden, J.T. Corticoid stress responses to handling and temperature in salmonids. Trans. Am. Fish. Soc. 2011, 106, 213–218. [Google Scholar] [CrossRef]
- Batiuk, R.; Brownson, K.; Dennison, W.; Ehrhart, M.; Hanson, J.; Hanmer, R.; Landry, B.; Reichert-Nguyen, J.; Soueidan, J.; Tassone, S.; et al. Rising Watershed and Bay Water Temperatures: Ecological Implications and Management Responses—A STAC Workshop; STAC Publication Number 23-001; The Scientific and Technical Advisory Committee: Edgewater, MD, USA, 2023. [Google Scholar]



| Physiographic Region | |||||
|---|---|---|---|---|---|
| Category | Variable | Units | Piedmont (n = 15) | Blue Ridge (n = 18) | Appalachian Plateau (n = 37) |
| Geophysical | Elevation [EL] | m | 218 (61) | 425 (50) | 751 (56) |
| Basin area [BA] | km2 | 6.2 (5.2) | 12.6 (8.0) | 25.2 (26.2) | |
| Baseflow index [BFI] | Index | 61.6 (4.1) | 49.4 (1.9) | 40.8 (0.3) | |
| Mean depth to bedrock [DB] | cm | 148 (3) | 133 (12) | 100 (5) | |
| Land use | Change in agricultural area [AG] | % | −0.36 (0.85) | 0.53 (0.56) | −0.19 (1.08) |
| Change in urban area [UR] | % | 0.52 (0.58) | 0.22 (0.24) | 0.47 (1.24) | |
| Change in forest area [FO] | % | 0.21 (1.06) | −0.55 (0.59) | −0.13 (2.03) | |
| Ponds within 1 km [PO] | Count | 0.93 (1.10) | 0.05 (0.24) | 0.08 (0.36) | |
| Non-native trout | Adult brown trout presence | Proportion of sites | 0.13 | 0.19 | 0.21 |
| Juvenile brown trout presence | Proportion of sites | 0.10 | 0.13 | 0.17 | |
| Air temperature | Mean annual air temperature trend [AT] | Sen’s slope | 0.025 (0.003) | 0.013 (0.003) | 0.024 (0.002) |
| ADU | YOY | EL | BA | BFI | DB | AG | UR | FO | PO | AT | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| ADU | - | 0.45 | 0.05 | 0.13 | −0.18 | −0.09 | 0.09 | 0.10 | −0.02 | −0.25 | −0.31 |
| YOY | <0.01 | - | 0.34 | 0.09 | −0.32 | −0.22 | 0.20 | −0.14 | <0.01 | −0.28 | −0.19 |
| EL | 0.68 | <0.01 | - | 0.44 | −0.83 | −0.72 | 0.01 | −0.11 | 0.06 | −0.34 | 0.12 |
| BA | 0.30 | 0.46 | <0.01 | - | −0.57 | −0.46 | −0.25 | 0.09 | 0.27 | −0.43 | −0.07 |
| BFI | 0.14 | 0.01 | <0.01 | <0.01 | - | 0.87 | 0.19 | −0.05 | −0.19 | 0.46 | 0.01 |
| DB | 0.43 | 0.07 | <0.01 | <0.01 | <0.01 | - | 0.23 | −0.05 | −0.25 | 0.43 | −0.06 |
| AG | 0.44 | 0.09 | 0.96 | 0.04 | 0.12 | 0.05 | - | −0.25 | −0.82 | −0.21 | −0.22 |
| UR | 0.40 | 0.25 | 0.37 | 0.44 | 0.71 | 0.69 | 0.04 | - | <0.01 | 0.20 | 0.11 |
| FO | 0.89 | 0.98 | 0.62 | 0.03 | 0.11 | 0.03 | <0.01 | 1.00 | - | 0.23 | 0.28 |
| PO | 0.04 | 0.02 | <0.01 | <0.01 | <0.01 | <0.01 | 0.08 | 0.10 | 0.05 | - | 0.38 |
| AT | 0.01 | 0.11 | 0.31 | 0.57 | 0.97 | 0.60 | 0.07 | 0.35 | 0.02 | <0.01 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hitt, N.P.; Rogers, K.M.; Kelly, Z.A. Declines in Brook Trout Abundance Linked to Atmospheric Warming in Maryland, USA. Hydrobiology 2024, 3, 310-324. https://doi.org/10.3390/hydrobiology3040019
Hitt NP, Rogers KM, Kelly ZA. Declines in Brook Trout Abundance Linked to Atmospheric Warming in Maryland, USA. Hydrobiology. 2024; 3(4):310-324. https://doi.org/10.3390/hydrobiology3040019
Chicago/Turabian StyleHitt, Nathaniel P., Karli M. Rogers, and Zachary A. Kelly. 2024. "Declines in Brook Trout Abundance Linked to Atmospheric Warming in Maryland, USA" Hydrobiology 3, no. 4: 310-324. https://doi.org/10.3390/hydrobiology3040019
APA StyleHitt, N. P., Rogers, K. M., & Kelly, Z. A. (2024). Declines in Brook Trout Abundance Linked to Atmospheric Warming in Maryland, USA. Hydrobiology, 3(4), 310-324. https://doi.org/10.3390/hydrobiology3040019