Abstract
Sulfated polysaccharides (SPs), especially those with immunostimulant effects on shrimp and fish, are bioactive compounds with potential use in aquaculture. In this study, Pacific white shrimp (Penaeus vannamei) were fed diets containing 0%, 0.5%, 1.0%, 1.5%, or 2.0% crude extracts from the microalga Porphyridium cruentum for 30 days. After dietary supplementation, the shrimp were exposed to Vibrio alginolyticus, allowing the assessment of phenoloxidase activity (PO) and total hemocyte count (THC) pre- and post-challenge, as well as shrimp survival rates. Shrimp that received the 1.0% treatment had greater survival (90%) at 48 h post-challenge, while the 0% and 2.0% treatments had survival rates close to 63% in the same period. Furthermore, at 48 h post-challenge, THC was significantly greater (p < 0.05) in the shrimp that received crude extract supplementation compared to that in shrimp of the control group (0%). PO showed a proportional increase according to crude extract inclusion levels. Such dose-dependency in response to PO increases became significantly greater (p < 0.05) in the 2.0% treatment group compared to that in the 0% treatment group in the pre-challenge period and that in the 0% and 0.5% treatment groups in the post-challenge period. These results showed that crude extracts from P. cruentum could be considered immunostimulants by increasing the resistance of P. vannamei exposed to V. alginolyticus, potentially representing a major breakthrough for marine shrimp farming.
1. Introduction
Farming of the Pacific white shrimp Penaeus vannamei stands out among the major species groups in global aquaculture production, which reached 5.8 million tons in 2020 [1]. However, shrimp farming has changed drastically in recent years [2,3] owing to the spread of diseases caused mainly by white spot syndrome virus (WSSV) [4] and Vibrio spp. [5]. Bacteria from the Vibrionaceae family make up the microbiota of seawater aquatic organisms, including shrimp, yet some species have been reported as opportunistic pathogens in stressful situations [5,6].
Despite the consensus on the lack of true adaptive immunity in invertebrates, some studies have suggested that shrimp possess an alternative adaptive immune system; however, the development of vaccines for commercial use still requires many studies [7]. Currently, trials evaluating the use of such practical products as extracts and feed additives to improve shrimp resistance have been conducted [4,5,8,9]. In addition, cellular and humoral parameters can assist in the identification and quantification of possible immunological responses in shrimp, as demonstrated in some studies [10]. In the present study, total hemocyte count (THC) and phenoloxidase activity (PO) are considered markers of immune response since changes in these parameters can be related to pathogen infection under adverse conditions [6,11,12].
Substances with immunostimulant potential in shrimp, such as β-glucans [13], bacterial products [14], and algal polysaccharides [8], can activate nonspecific defense systems and improve animal survival rates. Indeed, the polysaccharides of marine algae have already demonstrated their ability to boost the resistance of P. vannamei against stress conditions and pathogens [8,9]. In addition to its nutritional properties, microalgal biomass has received significant attention for its antioxidant [15], antibacterial [16], antifungal [17], and antiviral properties [18].
The red microalga Porphyridium cruentum is a potential source of sulphated polysaccharides, phycobilins, polyunsaturated fatty acids, and other bioactive substances, but its use in aquaculture and other biotechnological applications has not been well studied [19]. During the stationary phase of their growth curve, these cultivated microalgae produce a massive quantity of exopolysaccharides (EPSs) composed of different sugars, including xylose, galactose, glucose, mannose, arabinose, and sulfated oligosaccharides, which offer a wide range of applications with biotechnological potential [20,21].
Moreover, P. cruentum polysaccharides act as potent inducers of interleukin-6 cytokines and tumor necrosis factor (TNF-α), with no evidence of antigenic activity or hypersensitivity when administered intraperitoneally in mice [22]. In aquaculture research, it has already been found that EPSs from this microalga exhibited anti-VHSV activity (viral hemorrhagic septicemia virus, a key pathogen in fish farming) [23]. Furthermore, immersion baths with EPSs improved immunological parameters in P. vannamei exposed to V. harveyi, making EPSs promising candidates for use as preventive agents against vibriosis [24].
Therefore, investigations related to sulfated polysaccharides (SPs), especially those with immunostimulating effects in shrimp, are desirable in aquaculture. Thus, in the present study, Pacific white shrimp (P. vannamei) were fed diets containing 0%, 0.5%, 1.0%, 1.5%, or 2.0% sulphated polysaccharides (crude extracts) from the microalga P. cruentum for 30 days. After dietary supplementation, P. vannamei shrimp were exposed to V. alginolyticus, and PO activity, THC, and shrimp survival were evaluated pre- and post-challenge in order to verify possible immunomodulatory effects on shrimp.
2. Materials and Methods
2.1. Crude Extract from the Microalga Porphyridium cruentum
The P. cruentum strain was provided by the Laboratory of Algae Cultivation at the Federal University of Santa Catarina (UFSC), Brazil. It was cultivated in an adapted f/2-enriched seawater medium [25], and cultures were carried out until they reached the stationary phase (7–10 days). The polysaccharide (PS) crude extract of the microalgae P. cruentum was then extracted according to the methodology proposed by Farias et al. [26], whereby crude polysaccharide was extracted using a crude papain solution (30 mg mL−1) in 250 mL of 0.1 M sodium acetate buffer (pH 5.0) plus 5 mM EDTA and 5 mM cysteine from 5 g of biomass. The product was filtered and centrifuged at 7965× g for 20 min at 10 °C, after which the crude PS extract in the supernatant was concentrated by precipitation with 16 mL of 10% cetylpyridinium chloride (CPC), washed (200 mL; 0.05% CPC), and subsequently dissolved in 174 mL of 2 M NaCl:ethanol (100:15 v/v). Shortly afterwards, another precipitation with absolute ethanol for 24 h at 4 °C. The product was washed twice with 80% ethanol (200 mL), washed once with absolute ethanol (200 mL), and then oven-dried for 24 h at 60 °C. Crude extract from the microalga P. cruentum exhibited moisture of 7.92, crude protein of 11.39, ether extract of 0.22, ash content of 52.91, and fiber content of 0.45%.
2.2. Dietary Preparations
Different P. cruentum crude extract concentrations (0.5, 1.0, 1.5, and 2.0%) were added to commercial marine shrimp feed (Guabitech 1.6 mm Guabi®, Campinas, Brazil, moisture 100 g, crude protein 400 g, ether extract 110 g, crude fiber 40 g, and mineral matter 140 g/kg). The feed was separately ground into 5.0 kg lots, and each lot received a different supplement according to the P. cruentum crude extract concentrations specified above. These supplemented lots were mixed one by one for 15 min in a model-Y mixer. Subsequently, the mixture of dried ingredients was placed in a mixer and added to warm water in a sufficient amount until a consistent dough was formed. Finally, the dough was pelleted in an electrical grinder, and the formed pellets were oven-dried with air circulation for 24 h at 60 °C, bagged, and stored at 4 °C (Table 1). The control group, without supplementation, was subjected to the same procedures.

Table 1.
Centesimal composition of the experimental feed after the inclusion of crude extract from the microalgae P. cruentum. Results presented on a dry matter basis. The analyses were conducted at the LABNUTRI—Laboratório de Nutrição de Espécies Aquícolas, Federal University of Santa Catarina.
2.3. Feeding Assay
Two thousand specific pathogen-free (SPF) P. vannamei juveniles with an initial weight of 6.6 ± 0.2 g were randomly distributed in 20 glass fiber tanks (12 m2 bottom, 4 tanks for each treatment, and 100 shrimp tank−1). The 5 experimental diets were randomly assigned to 4 tanks. For 30 days, the shrimp were fed ad libitum at 10 am and 5 pm every day. The photoperiod was 12:12, and the water was renewed 50% daily at 8 am and 2 pm. During the feeding assay, the water quality variables remained at average values of 25.9 ± 0.15 °C, 33.0 ± 0.1 ppt salinity, 6.0 ± 0.3 mg L−1 dissolved oxygen, and 0.06 to 0.3 mg L−1 total ammonia.
After the feeding assay, the animals were randomly sampled (8 pools of 5 animals for each treatment, n = 40) for hemato-immunological parameter evaluation before infection, after which the animals were slaughtered on ice. In general, shrimp fed a diet supplemented with P. cruentum crude extract exhibited greater weight gain after 30 days, at 12.43 ± 0.3 g, 13.50 ± 0.8 g, 13.80 ± 0.5 g, 13.55 ± 0.6 g, and 13.18 ± 0.4 g, corresponding to 0%, 0.5%, 1.0%, 1.5%, and 2.0%, respectively [27].
2.4. Hemolymph Sampling for Serum Preparation
Shrimp hemolymph (n = 40 for each treatment) was sampled from the ventral region of each animal’s first abdominal segment with sterile 1.0 mL syringes attached to 21G needles previously cooled (4 °C) to delay coagulation. Eight pools of five animals were made for each treatment. For serum preparation, the hemolymph was allowed to coagulate for 24 h at 4 °C. The clot was frozen (−20 °C) and then thawed to allow cellular disruption and granule content release. After that, the samples were repeatedly centrifuged at 2000× g for 5 min. The supernatant of the disrupted clot, or cytoplasm-enriched serum, was sampled and frozen at −20 °C for subsequent use [8].
2.5. Hemato-Immunological Parameter Evaluation
THC was determined individually in a Neubauer chamber. A 10 µL hemolymph sample was fixed in 4% formaldehyde in Modified Alsever’s Solution (AMS) (27 mM sodium citrate, 336 mM NaCl, 115 mM glucose, 9 mM EDTA, pH 7.0) at a known dilution. Serum protein concentration was determined according to the Bradford method, using bovine serum albumin (BSA) as a standard. The assay was performed in triplicate.
PO activity was determined colorimetrically through the formation of the pigment coral red DOPA-chrome from the enzymatic oxidation of the substrate L-dihydroxyphenylalanine (L-DOPA, Sigma, São Paulo, Brazil) in the presence of serum PO. Fifty-microliter serum samples previously diluted (10-fold) in Tris-buffered saline (TBS) (50 mM Tris, 336 mM NaCl, 5 mM CaCl2, 10 mM MgCl2, pH 7.4) were incubated in equal volumes of trypsin (1 mg mL−1, Sigma) to induce enzymatic activity for 5 min at 20 °C in wells of a 96-well flat bottom microplate. In the control group, the inductor or serum was replaced with an equivalent volume of TBS.
After incubation, the wells received 50 µL of L-DOPA (3 mg mL−1), and the formation of DOPA-chrome was quantified in a microplate reader at 490 nm after 5, 10, 20, and 30 min. Specific PO enzymatic activity was expressed by the variation in absorbance by minute and by total protein in milligrams. One enzymatic unit corresponds to an increase of 0.001 in absorbance per minute and per milligram of protein at 20 °C [28]. The assay was performed in triplicate.
2.6. Vibrio alginolyticus Experimental Challenge
A strain of the marine bacterium V. alginolyticus, from the Belgian Coordinated Collections of Microorganisms—BCCM, was cultivated in brain heart infusion (BHI; Oxoid, São Paulo, Brazil) liquid medium under continuous mixing (200× g) at 30 °C. After 24 h of growth, the bacterial suspension was centrifuged at 2000× g, and the supernatant was discarded. The pellet was suspended in a sterile saline solution of 1.5% NaCl, and the bacterial concentration was adjusted to 105 CFU mL−1. This bacterial dose was previously standardized at the Marine Shrimp Laboratory (LCM), and it was demonstrated to induce 50% mortality (LD50) in shrimp within a 48 h period.
After 30 days of dietary supplementation, 40 shrimp (13.3 ± 0.3 g) from each treatment from the feeding assay were transferred to 20-L tanks (10 shrimp per tank in quadruplicate) containing treated seawater. The animals received an injection on the first dorsal segment of 25 µL of V. alginolyticus suspension (105 CFU mL−1). The group of 40 shrimp (10 per tank) was the negative control group and was injected with 25 µL of sterile saline solution. The survival rates were evaluated at 24 h and 48 h after injection. Hemolymph samples from all survivors were sampled to determine THC and PO activity.
2.7. Statistical Analysis
THC and PO activity data were log(x+1) transformed for normalization (Lilliefors (Kolmogorov-Smirnov) test) and variance homogenization (Levene’s test). One-way analysis of variance (ANOVA), followed by Duncan’s post hoc test, when necessary, was applied to evaluate the effect of crude P. cruentum extract concentration on the humoral immune response of juvenile shrimp to the immunostimulant. Mortality was recorded as the percentage of dead shrimp in each treatment. Between the 0–24 and 48 h periods in which no mortality was recorded, it was assumed that no dead shrimp were removed that day. At the end of each period, the observed mortalities were subtracted from the initial number of shrimp, and the survival rates were estimated. For all analyses, a significance level of 5% was adopted.
3. Results
3.1. Hemato-Immunological Parameters
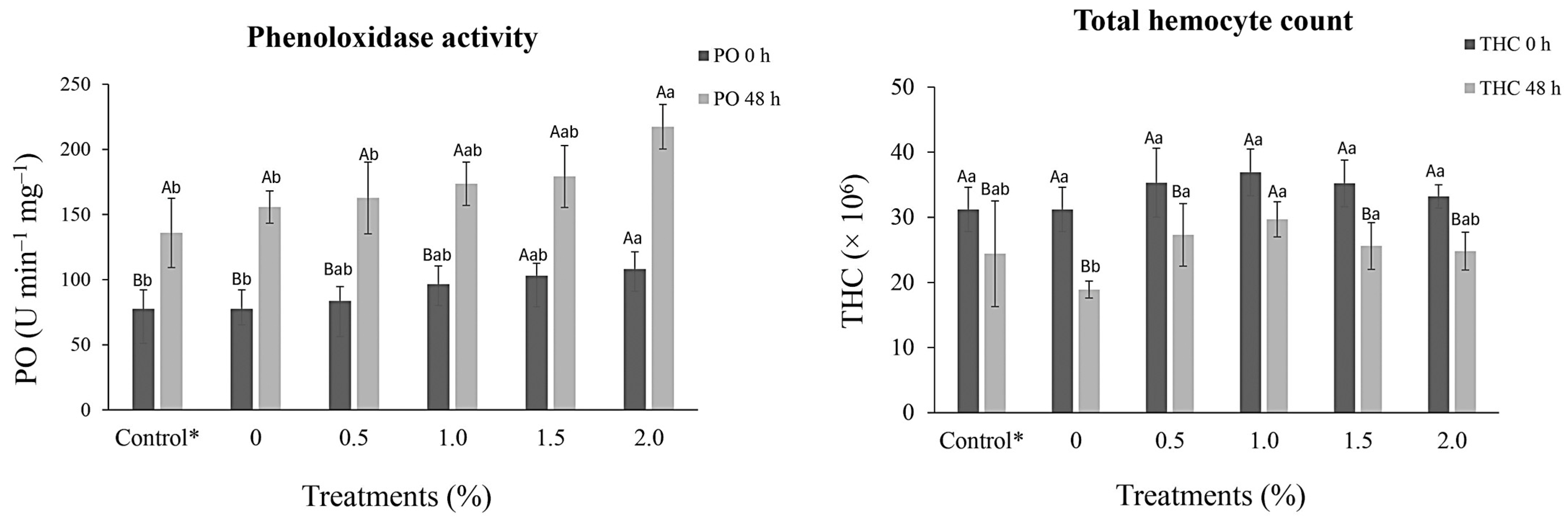
At 30 days following the feeding assay during the pre-challenge period, no significant difference (p > 0.05) was observed in THC levels among treatments. However, an increase in the absolute number of THC was observed, mainly in the 1.0% treatment group compared to that of the control group. After V. alginolyticus challenge, THC decreased in all treatments, but the control (0%) exhibited the most significant decrease at 39.42% (18.9 ± 1.3 × 106 cells mL−1) in comparison to the pre-challenge THC level (31.2 ± 3.4 × 106 cells mL−1) which was significantly different (p < 0.05) from that of the other supplemented treatments (Figure 1).
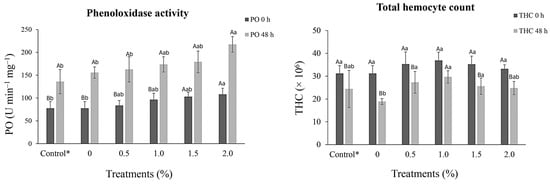
Figure 1.
Immunological responses (mean ± standard deviation) to phenoloxidase activity (PO) and total hemocyte count (THC) of Penaeus vannamei (13.3 ± 0.3 g) before (0 h) and after challenge (48 h) with Vibrio alginolyticus (5.0 × 106 CFU) after 30 days of dietary supplementation with crude extracts from Porphyridium cruentum. (AB) Different capital letters in the same column indicate significant differences (p < 0.05) between the pre- and post-challenge periods. (ab) Different lowercase letters in the same line indicate significant differences (p < 0.05) between treatments. (*) Shrimp injected with a saline solution.
After V. alginolyticus challenge, PO activity values increased in all treatments in proportion to increasing P. cruentum crude extract levels. However, in both the pre- and post-challenge assays, the 2.0% treatment group showed a significant increase (p < 0.05) compared to the PO values reported for the infected or saline-injected control groups (Figure 1).
3.2. Experimental Challenge
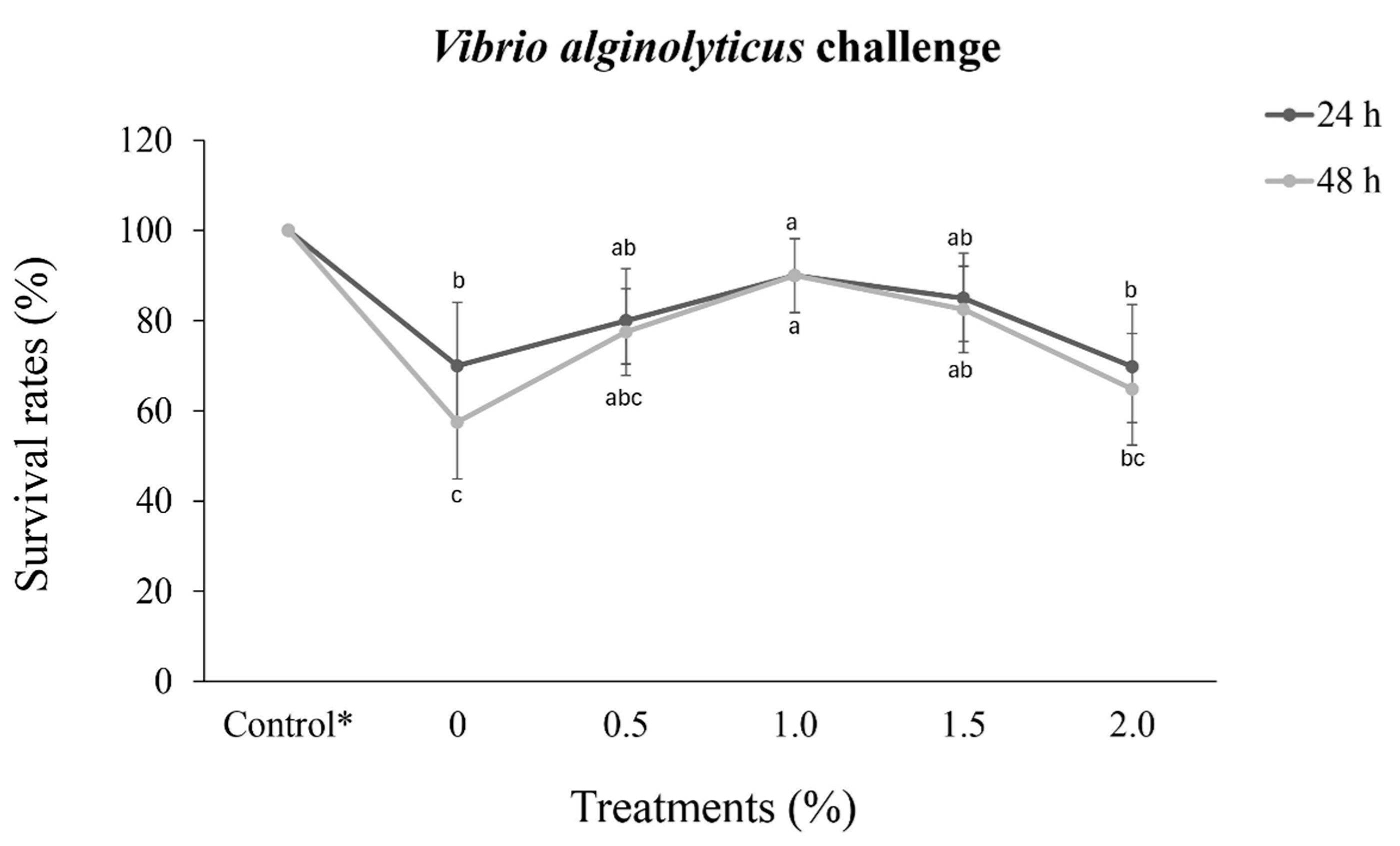
P. vannamei injected only with saline solution presented a 100% survival rate 48 h after challenge. In the other treatments, the survival rate differed significantly (p < 0.05) at both 24 h and 48 h. At 24 h post-challenge, P. vannamei in the 1.0% treatment group showed a significantly greater survival rate (p < 0.05) than that of shrimp in the 2.0% treatment and control groups (0%)
The lowest survival rate at 48 h post-challenge was recorded for the 0% control group (57.5%), and 48 h after challenge, the survival rates remained unchanged in the 1.0% treatment group, further revealing a significant difference in survival rate (p < 0.05) compared to that of either the 2.0% treatment or control groups (0%).
The lowest survival rates occurred in the 0% control group, which presented high mortality in both the 24 h and 48 h post-challenge periods (Figure 2).
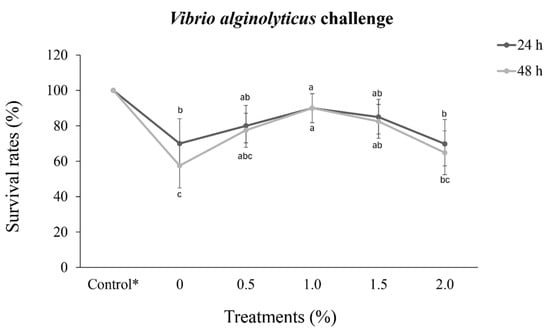
Figure 2.
Survival rates (%) (mean ± standard deviation) of Penaeus vannamei (13.3 ± 0.3 g) challenged with Vibrio alginolyticus (5.0 × 106 CFU) after 30 days of dietary supplementation with crude extracts from Porphyridium cruentum. Different lowercase letters in the same line indicate significant differences (p < 0.05) between treatments. (*) Shrimp fed a diet without supplementation and injected with a saline solution.
4. Discussion
At present, biocompounds that improve marine shrimp immunocompetence have emerged as a sustainable solution against pathogens [2,3,4,8,9,12,18,19,23,29]. Health-promoting additives may contribute to the success of P. vannamei farming [2]. The positive immunostimulant effect of P. cruentum crude extract showed promise since microalgal cultures may be produced using shrimp farming effluents, possibly reducing production costs [30]. Other microalgal genera, such as Arthrospira (Spirulina) [31,32] and Chlorella [33], also exhibited satisfactory results in the immunostimulation of shrimp, boosting phagocytic activity, for example.
Immunocompetent cells or hemocytes are the major sites for the synthesis of immunologically active molecules in shrimp [34] and are directly involved in phagocytosis, nodule and capsule formation, and the production and liberation of cytotoxic and lytic molecules capable of eliminating invasive microorganisms [35]. Huang et al. [36], for instance, reported that the oral use of a sulphated polysaccharide extracted from the brown seaweed Sargassum fusiforme on Fenneropenaeus chinensis infected by V. harveyi resulted in increased resistance to pathogenic bacterial infection. Brown and red seaweed have high levels of sulfated polysaccharides [37], and they have been reported as alternatives for shrimp immunostimulation [8,9,36].
This is the first report on the use of polysaccharides from the red microalga P. cruentum as a feed supplement for P. vannamei. Similar to the results herein reported, the addition of fucoidan from the brown seaweed Sargassum polycystum to the diet of black tiger shrimp significantly increased the survival rate of animals infected with white spot syndrome virus. Moreover, crude fucoidan extract was still able to inhibit the growth of V. harveyi, Staphylococcus aureus, and Escherichia coli [38].
The application of different strategies for the administration of sulfate polysaccharides, such as immersion baths [39], spraying [40], and injection [39,41], has been used. However, oral ingestion via feed may be the least expensive and most practical strategy without damaging its molecular properties. According to Lee et al. [31], the resistance of the shrimp Penaeus merguiensis increased when the shrimp were fed a diet containing the microalga Arthrospira platensis (Spirulina), owing to the activation of hemocyte phagocytic activity against the bacteria V. harveyi, E. coli, S. typhimurium, and B. subtilis. This immunostimulant activity is related to the presence of lipopolysaccharides and peptidoglycans in Spirulina.
After V. alginolyticus infection, the control treatment without P. cruentum crude extract supplementation showed a 39.42% reduction in THC compared to that found in the 1.0% group, suggesting that the supplemented diet could have assisted in maintaining the quantity of circulating hemocytes in the infected shrimp. This result seems to be associated with an inflammatory response from the hemocytes that migrate to the inoculation region, escaping circulation [42]. Furthermore, hemocytes can aggregate in hemocytic nodules, wherein cellular adhesion molecules, such as peroxinectin, act to capture microorganisms in their interior [43] and can be withdrawn from circulation by mechanical processes performed by the gills.
In the present study, the decrease in THC levels in shrimp fed a diet supplemented with P. cruentum crude extract was lower than that in the control group without supplementation. This result may be associated with faster hemocyte repositioning in the circulation by the hematopoietic tissue [42] of shrimp fed the crude extract. Reports in the literature about the effects of sulfated polysaccharide immunostimulants are controversial and difficult to compare, mainly because of the various methodologies used for polysaccharides and their different compositions [44,45].
The relationship between PO activity and hemocyte concentration can be explained by the transport of immunoassayed molecules by these cells [36,46]. In the pre-challenge period, higher THC levels were recorded in the 1.0% supplementation group than those in the control group, but THC did not increase in proportion to the supplementation concentration. This suggests that PO activity could have directly resulted from P. cruentum crude extract, or, more precisely, the negatively charged sulfonate regions that activated the proPO system. This interaction was demonstrated between the glycoproteins present in the viral envelope and sulfated polysaccharides [47].
Notably, judging effective prophylaxis after the first contact with the pathogen is a major obstacle to consolidating the use of immunostimulants in aquaculture [32]. Here, however, it was found that survival after V. alginolyticus challenge was significantly greater in the 1.0% supplementation treatment group, suggesting a baseline from which it could be determined that dietary supplementation was sufficiently effective to increase immunocompetence in P. vannamei, thereby contributing to the increased survival rate.
5. Conclusions
Dietary supplementation with crude extract from the red microalga P. cruentum significantly increased the immunocompetence and survival of the Pacific white shrimp P. vannamei. Considering the results observed in the present study, a concentration of 1.0% is recommended for the dietary inclusion of P. vannamei. However, other immunological parameters, as well as immune protein expression, must be evaluated to confirm these results. In this context, we anticipate further studies to elucidate the mechanisms of action of these polysaccharide crude extracts and their prophylactic efficacy.
Author Contributions
R.Á.O.: conceptualization, methodology, investigation, writing—original draft; R.G.L. methodology, investigation, formal analysis; F.d.N.V.: methodology and investigation; N.C.B.-R.: methodology, investigation, formal analysis; C.Y.B.d.O.: methodology, investigation, writing—original draft; M.A.A.M.B.: methodology and investigation; M.S.O.: formal analysis, writing—original draft, writing—review and editing; D.M.F.: methodology and investigation; R.B.D.: conceptualization, methodology, investigation, funding acquisition, project administration, writing—original draft (equal), writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
The Ministry of Science, Technology and Innovation (MCTI—Brazil) Agreement No. 01245.018395/2023-9) and the Coordination of Superior Level Staff Improvement (CAPES) (Finance Code 001).
Institutional Review Board Statement
All procedures in this research were approved by the Ethics Committee on Animal Use (CEUA) of the Federal University of Santa Catarina—UFSC, Protocol number 8503260819, approve time March in 2022.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data related to this research are available upon prior request.
Acknowledgments
The authors would like to thank the responsible technical personnel and the students of the Marine Shrimp Laboratory (LCM) of Federal University of Santa Catarina (UFSC) for their logistical support in the experimental execution and immunological analysis, the Ministry of Science, Technology and Innovation (MCTI), and the Coordination of Superior Level Staff Improvement.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- FAO—Food and Agriculture Organization of the United Nations. The State of World Fisheries and Aquaculture. In Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Barreto, A.; Peixoto, D.; Fajardo, C.; Pinto, W.; Rocha, R.J.; Conceição, L.E.; Costas, B. Health-promoting additives supplemented in inert microdiets for whiteleg shrimp (Penaeus vannamei) post-larvae: Effects on growth, survival, and health status. Animals 2023, 13, 726. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Abakari, G.; Tan, H.; Wenchang, L.I.U.; Luo, G. Effects of different probiotics (Bacillus subtilis) addition strategies on a culture of Litopenaeus vannamei in biofloc technology (BFT) aquaculture system. Aquaculture 2023, 566, 739216. [Google Scholar] [CrossRef]
- Schleder, D.D.; Peruch, L.G.B.; Poli, M.A.; Ferreira, T.H.; Silva, C.P.; Andreatta, E.R.; Hayashi, L.; Vieira, F.D.N. Effect of brown seaweeds on Pacific white shrimp growth performance, gut morphology, digestive enzymes activity and resistance to white spot virus. Aquaculture 2018, 495, 359–365. [Google Scholar] [CrossRef]
- Filho, L.G.A.D.S.; Diniz, F.M.; Pereira, A.M. Immunostimulants derived from plants and algae to increase resistance of pacific white shrimp (Litopenaeus vannamei) against vibriosis. Stud. Nat. Prod. Chem. 2023, 77, 297–337. [Google Scholar] [CrossRef]
- Le Moullac, G.; Haffner, P. Environmental factors affecting immune responses in Crustacea. Aquaculture 2000, 191, 121–131. [Google Scholar] [CrossRef]
- Amatul-Samahah, M.A.; Omar, W.H.H.W.; Ikhsan, N.F.M.; Azmai, M.N.A.; Zamri-Saad, M.; Ina-Salwany, M.Y. Vaccination trials against vibriosis in shrimp: A review. Aquac. Rep. 2020, 18, 100471. [Google Scholar] [CrossRef]
- Schleder, D.D.; Da Rosa, J.R.; Guimarães, A.M.; Ramlov, F.; Maraschin, M.; Seiffert, W.Q.; Vieira, F.D.N.; Hayashi, L.; Andreatta, E.R. Brown seaweeds as feed additive for white-leg shrimp: Effects on thermal stress resistance, midgut microbiology, and immunology. J. Appl. Phycol. 2017, 29, 2471–2477. [Google Scholar] [CrossRef]
- Cantelli, L.; Goncalves, P.; Guertler, C.; Kayser, M.; Pilotto, M.R.; Barracco, M.A.; Perazzolo, L.M. Dietary supplementation with sulfated polysaccharides from Gracilaria birdiae promotes a delayed immunostimulation in marine shrimp challenged by the white spot syndrome virus. Aquac. Int. 2019, 27, 349–367. [Google Scholar] [CrossRef]
- Kulkarni, A.; Krishnan, S.; Anand, D.; Kokkattunivarthil Uthaman, S.; Otta, S.K.; Karunasagar, I.; Kooloth Valappil, R. Immune responses and immunoprotection in crustaceans with special reference to shrimp. Rev. Aquac. 2021, 13, 431–459. [Google Scholar] [CrossRef]
- Sritunyalucksana, K.; Söderhäll, K. The proPO and clotting system in crustaceans. Aquaculture 2000, 191, 53–69. [Google Scholar] [CrossRef]
- Liu, F.; Shao, G.Y.; Tian, Q.Q.; Cheng, B.X.; Shen, C.; Wang, A.M.; Zhang, J.H.; Tian, H.Y.; Yang, W.P.; Yu, Y.B. Enhanced growth performance, immune responses, immune-related gene expression and disease resistance of red swamp crayfish (Procambarus clarkii) fed dietary glycyrrhizic acid. Aquaculture 2021, 533, 736202. [Google Scholar] [CrossRef]
- Ringø, E.; Song, S.K. Application of dietary supplements (synbiotics and probiotics in combination with plant products and β-glucans) in aquaculture. Aquac. Nutr. 2016, 22, 4–24. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, Y.; Zhang, Q.; Zhang, X.; Yuan, J.; Huang, H.; Xiang, J.; Li, F. A novel candidate gene associated with body weight in the Pacific white shrimp Litopenaeus vannamei. Front. Genet. 2019, 10, 434956. [Google Scholar] [CrossRef] [PubMed]
- Feller, R.; Matos, Â.P.; Mazzutti, S.; Moecke, E.H.; Tres, M.V.; Derner, R.B.; Oliveira, J.V.; Junior, A.F. Polyunsaturated ω-3 and ω-6 fatty acids, total carotenoids and antioxidant activity of three marine microalgae extracts obtained by supercritical CO2 and subcritical n-butane. J. Supercrit. Fluids 2018, 133, 437–443. [Google Scholar] [CrossRef]
- Dantas, D.M.D.M.; Oliveira, C.Y.B.; Costa, R.M.P.B.; Carneiro-da-Cunha, M.D.G.; Gálvez, A.O.; Bezerra, R.D.S. Evaluation of antioxidant and antibacterial capacity of green microalgae Scenedesmus subspicatus. Food Sci. Technol. Int. 2019, 25, 318–326. [Google Scholar] [CrossRef]
- Najdenski, H.M.; Gigova, L.G.; Iliev, I.I.; Pilarski, P.S.; Lukavský, J.; Tsvetkova, I.V.; Ninova, M.S.; Kussovski, V.K. Antibacterial and antifungal activities of selected microalgae and cyanobacteria. Int. J. Food Sci. Technol. 2013, 48, 1533–1540. [Google Scholar] [CrossRef]
- Charoonnart, P.; Worakajit, N.; Zedler, J.A.; Meetam, M.; Robinson, C.; Saksmerprome, V. Generation of microalga Chlamydomonas reinhardtii expressing shrimp antiviral dsRNA without supplementation of antibiotics. Sci. Rep. 2019, 9, 3164. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ji, L.; Shi, Q.; Wu, H.; Fan, J. Advances in the production of bioactive substances from marine unicellular microalgae Porphyridium spp. Bioresour. Technol. 2019, 292, 122048. [Google Scholar] [CrossRef]
- Arad, S.M.; Levy-Ontman, O. Red microalgal cell-wall polysaccharides: Biotechnological aspects. Curr. Opin. Biotechnol. 2010, 21, 358–364. [Google Scholar] [CrossRef]
- Martínez, J.M.; Delso, C.; Álvarez, I.; Raso, J. Pulsed electric field permeabilization and extraction of phycoerythrin from Porphyridium cruentum. Algal Res. 2019, 37, 51–56. [Google Scholar] [CrossRef]
- Casas-Arrojo, V.; Decara, J.; Arrojo-Agudo, M.D.L.; Pérez-Manríquez, C.; Abdala-Díaz, R.T. Immunomodulatory, antioxidant activity and cytotoxic effect of sulfated polysaccharides from Porphyridium cruentum. (sf Gray) Nägeli. Biomolecules 2021, 11, 488. [Google Scholar] [CrossRef] [PubMed]
- Parra-Riofrio, G.; Moreno, P.; García-Rosado, E.; Alonso, M.C.; Uribe-Tapia, E.; Abdala-Diaz, R.T.; Bejar, J. Tetraselmis suecica and Porphyridium cruentum exopolysaccharides show anti-VHSV activity on RTG-2 cells. Aquac. Int. 2023, 31, 3145–3157. [Google Scholar] [CrossRef]
- Risjani, Y.; Mutmainnah, N.; Manurung, P.; Wulan, S.N.; Yunianta. Exopolysaccharide from Porphyridium cruentum (purpureum) is not toxic and stimulates immune response against vibriosis: The assessment using zebrafish and white shrimp Litopenaeus vannamei. Mar. Drugs 2021, 19, 133. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.R.; Ryther, J.H. Studies of marine planktonic diatoms: I. Cyclotella nana Hustedt, and Detonula confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Farias, W.R.; Valente, A.P.; Pereira, M.S.; Mourão, P.A. Structure and anticoagulant activity of sulfated galactans: Isolation of a unique sulfated galactan from the red algae Botryocladia occidentalis and comparison of its anticoagulant action with that of sulfated galactans from invertebrates. J. Biol. Chem. 2000, 275, 29299–29307. [Google Scholar] [CrossRef] [PubMed]
- Ozório, R.Á.; Lopes, R.G.; Góes, B.; Silva, A.C.P.; Derner, R.B.; Fracalossi, D.M. Growth and enzymatic profile of the pacific white shrimp fed with Porphyridium cruentum extract. Bol. Inst. Pesca 2015, 41, 123–131. [Google Scholar]
- Söderhäll, K.; Häll, L. Lipopolysaccharide-induced activation of prophenoloxidase activating system in crayfish haemocyte lysate. Biochim. Biophys. Acta (BBA) Gen. Subj. 1984, 797, 99–104. [Google Scholar] [CrossRef]
- Wang, X.W.; Wang, J.X. Pattern recognition receptors acting in innate immune system of shrimp against pathogen infections. Fish Shellfish Immunol. 2013, 34, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Magnotti, C.; Lopes, R.; Derner, R.; Vinatea, L. Using residual water from a marine shrimp farming BFT system. Part I: Nutrient removal and marine microalgae biomass production. Aquac. Res. 2016, 47, 2435–2443. [Google Scholar] [CrossRef]
- Lee, Y.K.; Chew, P.F.; Soh, B.S.; Tham, L.Y. Enhancing phagocytic activity of hemocytes and disease resistance in the prawn Penaeus merguiensis by feeding Spirulina platensis. J. Appl. Phycol. 2003, 15, 279–287. [Google Scholar] [CrossRef]
- Pilotto, M.R.; Milanez, S.; Moreira, R.T.; Rosa, R.D.; Perazzolo, L.M. Potential immunomodulatory and protective effects of the Arthrospira-based dietary supplement on shrimp intestinal immune defenses. Fish Shellfish Immunol. 2019, 88, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Pakravan, S.; Akbarzadeh, A.; Sajjadi, M.M.; Hajimoradloo, A.; Noori, F. Chlorella vulgaris meal improved growth performance, digestive enzyme activities, fatty acid composition and tolerance of hypoxia and ammonia stress in juvenile Pacific white shrimp Litopenaeus vannamei. Aquac. Nutr. 2018, 24, 594–604. [Google Scholar] [CrossRef]
- Gross, P.S.; Bartlett, T.C.; Browdy, C.L.; Chapman, R.W.; Warr, G.W. Immune gene discovery by expressed sequence tag analysis of hemocytes and hepatopancreas in the Pacific White Shrimp, Litopenaeus vannamei, and the Atlantic White Shrimp, L. Setiferus. Dev. Comp. Immunol. 2001, 25, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Millar, D.A.; Ratcliffe, N.A. Invertebrates. In Immunology: A Comparative Approach; Turner, R.J., Ed.; Wiley: Chichester, UK, 1994; pp. 29–68. [Google Scholar]
- Huang, X.; Zhou, H.; Zhang, H. The effect of Sargassum fusiforme polysaccharide extracts on vibriosis resistance and immune activity of the shrimp, Fenneropenaeus chinensis. Fish Shellfish Immunol. 2006, 20, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Suganya, A.M.; Sanjivkumar, M.; Chandran, M.N.; Palavesam, A.; Immanuel, G. Pharmacological importance of sulphated polysaccharide carrageenan from red seaweed Kappaphycus alvarezii in comparison with commercial carrageenan. Biomed. Pharmacother. 2016, 84, 1300–1312. [Google Scholar] [CrossRef] [PubMed]
- Chotigeat, W.; Tongsupa, S.; Supamataya, K.; Phongdara, A. Effect of fucoidan on disease resistance of black tiger shrimp. Aquaculture 2004, 233, 23–30. [Google Scholar] [CrossRef]
- Yeh, S.T.; Lee, C.S.; Chen, J.C. Administration of hot-water extract of brown seaweed Sargassum duplicatum via immersion and injection enhances the immune resistance of white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2006, 20, 332–345. [Google Scholar] [CrossRef] [PubMed]
- Abbas, E.M.; Al-Souti, A.S.; Sharawy, Z.Z.; El-Haroun, E.; Ashour, M. Impact of dietary administration of seaweed polysaccharide on growth, microbial abundance, and growth and immune-related genes expression of the pacific whiteleg shrimp (Litopenaeus vannamei). Life 2023, 13, 344. [Google Scholar] [CrossRef]
- Yuvaraj, N.; Arul, V. Immunomodulatory effects of seagrass Halophila ovalis polysaccharide mixed feed in adult black tiger shrimp Penaeus monodon and its protective efficacy against white spot syndrome virus infection. Iran. J. Fish. Sci. 2017, 16, 993–1007. Available online: http://hdl.handle.net/1834/12261 (accessed on 20 January 2024).
- Van de Braak, C.B.T.; Botterblom, M.H.A.; Huisman, E.A.; Rombout, J.H.W.M.; Van der Knaap, W.P.W. Preliminary study on haemocyte response to white spot syndrome virus infection in black tiger shrimp Penaeus monodon. Dis. Aquat. Org. 2002, 51, 149–155. [Google Scholar] [CrossRef]
- Jiravanichpaisal, P.; Lee, B.L.; Söderhäll, K. Cell-mediated immunity in arthropods: Hematopoiesis, coagulation, melanization and opsonization. Immunobiology 2006, 211, 213–236. [Google Scholar] [CrossRef] [PubMed]
- Bakky, M.A.H.; Tran, N.T.; Zhang, Y.; Li, S. Utilization of marine macroalgae-derived sulphated polysaccharides as dynamic nutraceutical components in the feed of aquatic animals: A review. Aquac. Res. 2022, 53, 5787–5808. [Google Scholar] [CrossRef]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important determinants for fucoidan bioactivity: A critical review of structure-function relations and extraction methods for fucose-containing sulfated polysaccharides from brown seaweeds. Mar. Drugs 2011, 9, 2106–2130. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.W.; Keyser, P.; Sritunyalucksana, K.; Söderhäll, K. Crustacean haemocytes and haematopoiesis. Aquaculture 2000, 191, 45–52. [Google Scholar] [CrossRef]
- Damonte, E.B.; Matulewicz, M.C.; Cerezo, A.S. Sulfated seaweed polysaccharides as antiviral agents. Curr. Med. Chem. 2004, 11, 2399–2419. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).