
Maximal Fat Metabolism Explained by Lactate-Carbohydrate Model


Abstract
1. Introduction
2. Data, Model, Applications and Influences or Materials and Methods
2.1. Study Design and Participants
2.2. Procedures and Protocol
2.3. Measurements
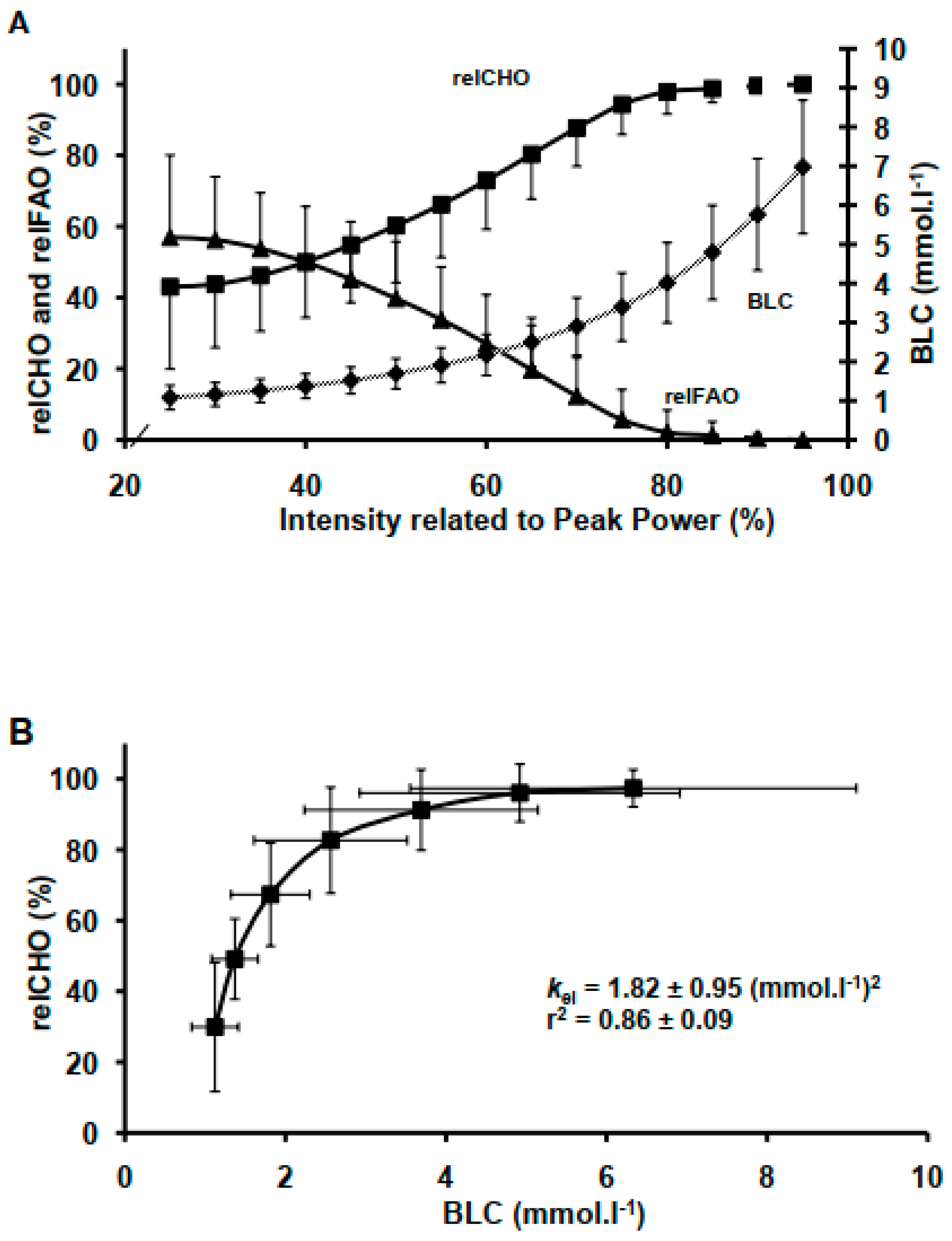
2.4. Data Processing, Analyses and Mathematical Modelling, Analyses and Statistics
2.5. Statistical Analyses
3. Results
3.1. MFO, Fatmax and COP
3.2. BLC, relCHO, relFAO Responses and BLC-relCHO Model
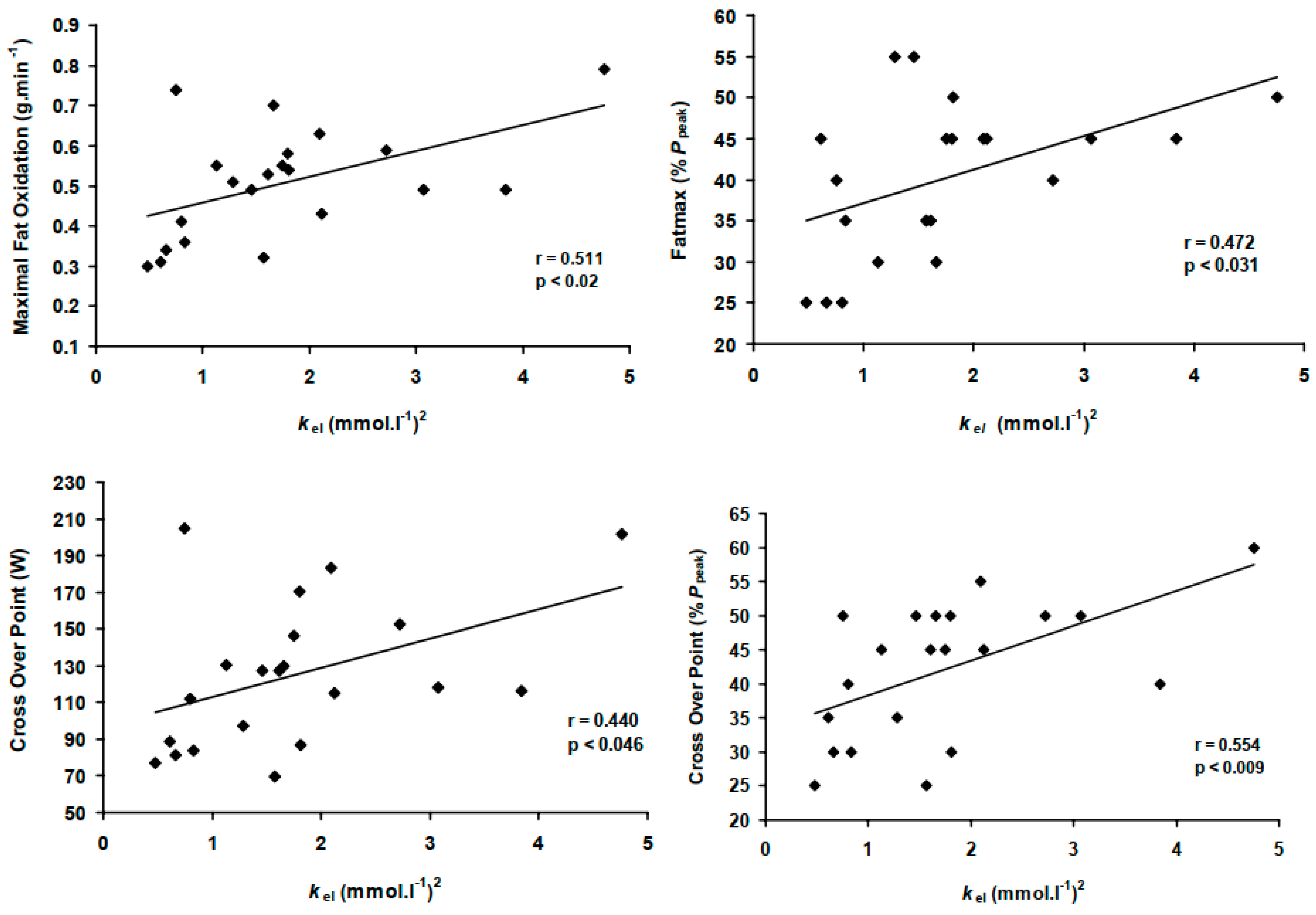
3.3. Bivariate Analysis: kel Correlation with MFO, COP and Associated Exercise Intensities
3.4. The Variability in MFO, Fatmax and COP: Multivariate Analysis
4. Discussion
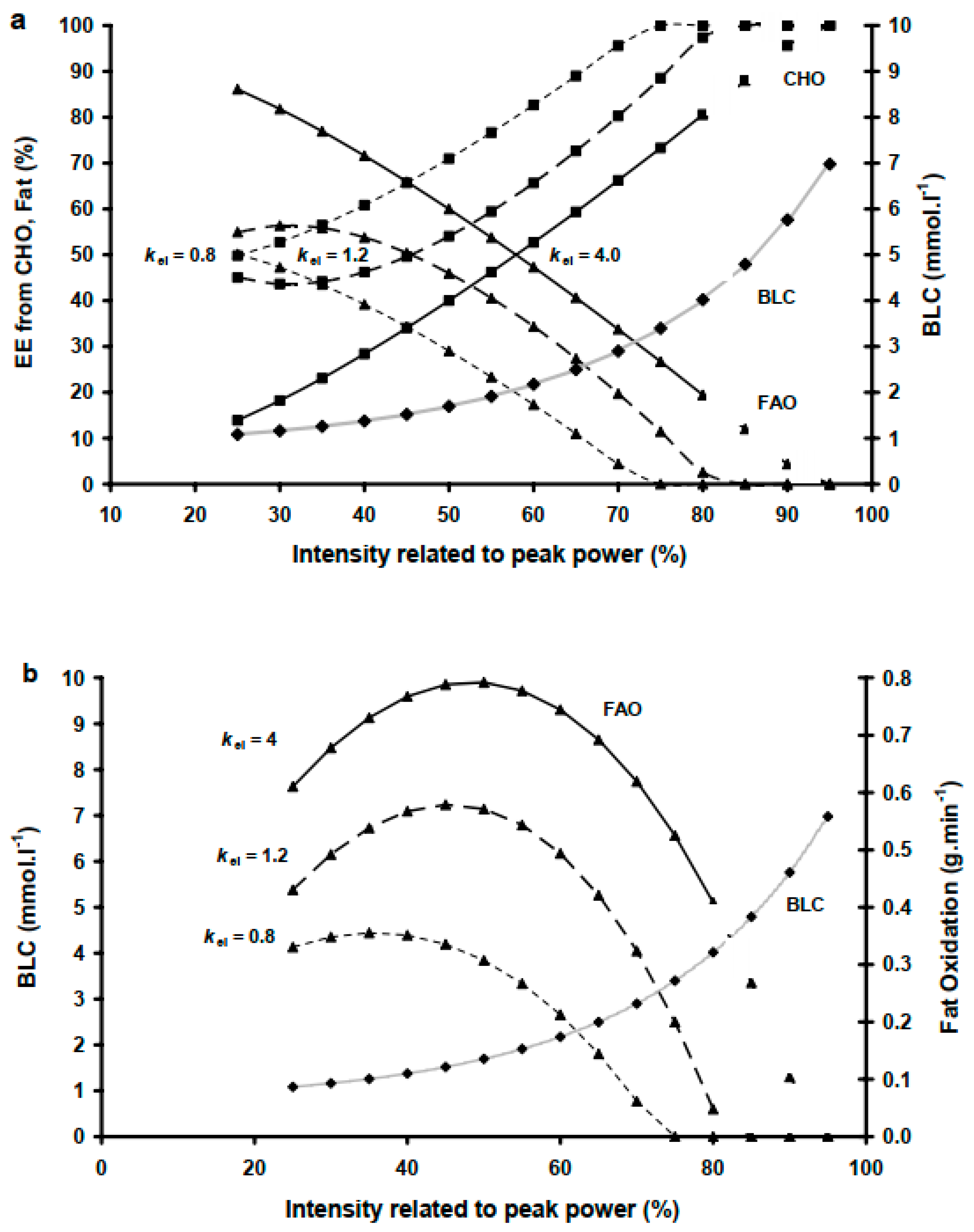
4.1. kel Prediction of MFO, Fatmax and COP
4.2. kel and the Intra-Individual Variability in COP and Fatmax
4.3. Limitations
5. Conclusions and Prospects
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brooks, G.A.; Fahey, T.D.; Baldwin, K.M. Exercise Physiology. Human Bioenergetics and Its Adaptations, 3rd ed.; McGraw Hill: Boston, MA, USA, 2005; pp. 61–82. [Google Scholar]
- Spriet, L.L. New insights into the interaction of carbohydrate and fat metabolism during exercise. Sports Med. 2014, 44 (Suppl. 1), S87–S96. [Google Scholar] [CrossRef] [PubMed]
- Dilworth, L.; Facey, A.; Omoruyi, F. Diabetes Mellitus and Its Metabolic Complications: The Role of Adipose Tissues. Int. J. Mol. Sci. 2021, 22, 7644. [Google Scholar] [CrossRef] [PubMed]
- Jeukendrup, A.; Wallis, G. Measurements of substrate oxidation during exercise by means of gas exchange measurements. Int. J. Sports Med. 2005, 26, S28–S37. [Google Scholar] [CrossRef] [PubMed]
- Frayn, K.N. Calculation of substrate oxidation rates in vivo from gaseous exchange. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1983, 55, 628–634. [Google Scholar] [CrossRef]
- Kuo, C.H.; Harris, M.B.; Jensen, J.; Alkhatib, A.; Ivy, J.L. Editorial: Possible Mechanisms to Explain Abdominal Fat Loss Effect of Exercise Training Other Than Fatty Acid Oxidation. Front. Physiol. 2021, 12, 789463. [Google Scholar] [CrossRef]
- Alkhatib, A. Predictors of exercise performance. In Trends in Human Performance Research; Duncan, M.J., Lyons, M., Eds.; NOVA Science Publisher: New York, NY, USA, 2010; pp. 168–183. [Google Scholar]
- Bergman, B.C.; Brooks, G.A. Respiratory gas-exchange ratios during graded exercise in fed and fasted trained and untrained men. J. Appl. Physiol. 1999, 86, 479–487. [Google Scholar] [CrossRef]
- Helge, J.W.; Fraser, A.M.; Kriketos, A.D.; Jenkins, A.B.; Calvert, G.D.; Ayre, K.J.; Storlien, L.H. Interrelationships between muscle fibre type, substrate oxidation and body fat. Int. J. Obes. Relat. Metab. Disord. 1999, 23, 986–991. [Google Scholar] [CrossRef]
- Venables, M.C.; Achten, J.; Jeukendrup, A.E. Determinants of fat oxidation during exercise in healthy men and women: A cross-sectional study. J. Appl. Physiol. 2005, 98, 160–167. [Google Scholar] [CrossRef]
- Romijn, J.A.; Coyle, E.F.; Sidossis, L.S.; Gastaldelli, A.; Horowitz, J.F.; Endert, E.; Wolfe, R.R. Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am. J. Physiol. 1993, 265 Pt 1, E380–E391. [Google Scholar] [CrossRef]
- Achten, J.; Gleeson, M.; Jeukendrup, A.E. Determination of the exercise intensity that elicits maximal fat oxidation. Med. Sci. Sports Exerc. 2002, 34, 92–97. [Google Scholar] [CrossRef]
- Pérez-Martin, A.; Dumortier, M.; Raynaud, E.; Brun, J.F.; Fédou, C.; Bringer, J.; Mercier, J. Balance of substrate oxidation during submaximal exercise in lean and obese people. Diabetes Metab. 2001, 27 Pt 1, 466–474. [Google Scholar] [PubMed]
- Brun, J.F.; Jean, E.; Ghanassia, E.; Flavier, S.; Mercier, J. Metabolic training: New paradigms of exercise training for metabolic diseases with exercise calorimetry targeting individuals. Ann. Readapt. Med. Phys. 2007, 50, 528–534, 520–7. English, French. [Google Scholar] [CrossRef] [PubMed]
- Nordby, P.; Saltin, B.; Helge, J.W. Whole-body fat oxidation determined by graded exercise and indirect calorimetry: A role for muscle oxidative capacity? Scand. J. Med. Sci. Sports 2006, 16, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A.; Mercier, J. Balance of carbohydrate and lipid utilization during exercise: The “crossover” concept. J. Appl. Physiol. 1994, 76, 2253–2261. [Google Scholar] [CrossRef]
- Hill, A.V.; Long, C.N.H.; Lupton, H. Muscular exercise, lactic acid and the supply and utilisation of oxygen.—Parts VII–VIII. Proc. R. Soc. Lond. 1924, 97, 155–176. [Google Scholar] [CrossRef]
- Achten, J.; Jeukendrup, A.E. Maximal fat oxidation during exercise in trained men. Int. J. Sports Med. 2003, 24, 603–608. [Google Scholar] [CrossRef]
- Achten, J.; Jeukendrup, A.E. Relation between plasma lactate concentration and fat oxidation rates over a wide range of exercise intensities. Int. J. Sports Med. 2004, 25, 32–37. [Google Scholar] [CrossRef]
- Brandou, F.; Dumortier, M.; Garandeau, P.; Mercier, J.; Brun, J.F. Effects of a two-month rehabilitation program on substrate utilization during exercise in obese adolescents. Diabetes Metab. 2003, 29, 20–27. [Google Scholar] [CrossRef]
- Alkhatib, A.; Atcheson, R. Yerba Maté (Ilex paraguariensis) Metabolic, Satiety, and Mood State Effects at Rest and during Prolonged Exercise. Nutrients 2017, 9, 882. [Google Scholar] [CrossRef]
- Meyer, T.; Folz, C.; Rosenberger, F.; Kindermann, W. The reliability of fat. Scand. J. Med. Sci. Sports. 2009, 19, 213–221. [Google Scholar] [CrossRef]
- Michallet, A.S.; Tonini, J.; Regnier, J.; Guinot, M.; Favre-Juvin, A.; Bricout, V.; Halimi, S.; Wuyam, B.; Flore, P. Methodological aspects of crossover and maximum fat-oxidation rate point determination. Diabetes Metab. 2008, 34, 514–523. [Google Scholar] [CrossRef]
- Rynders, C.A.; Angadi, S.S.; Weltman, N.Y.; Gaesser, G.A.; Weltman, A. Oxygen uptake and ratings of perceived exertion at the lactate threshold and maximal fat oxidation rate in untrained adults. Eur. J. Appl. Physiol. 2011, 111, 2063–2068. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bircher, S.; Knechtle, B.; Knecht, H. Is the intensity of the highest fat oxidation at the lactate concentration of 2 mmol L(-1)? A comparison of two different exercise protocols. Eur. J. Clin. Investig. 2005, 35, 491–498. [Google Scholar] [CrossRef] [PubMed]
- González-Haro, C. Maximal fat oxidation rate and cross-over point with respect to lactate thresholds do not have good agreement. Int. J. Sports Med. 2011, 32, 379–385. [Google Scholar] [CrossRef]
- Alkhatib, A. Lactate, Carbohydrate, and Fat Utilisation during Exercise: Interrelationship, and Applications in Exercise Testing; Lambert Academic Publishing: Hamburg, Germany, 2010. [Google Scholar]
- Beneke, R.; Hütler, M.; Leithäuser, R.M. Carbohydrate and fat metabolism related to blood lactate in boys and male adolescents. Eur. J. Appl. Physiol. 2009, 105, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Beneke, R.; Alkhatib, A. High cycling cadence reduces carbohydrate oxidation at given low intensity metabolic rate. Biol. Sport 2015, 32, 27–33. [Google Scholar] [CrossRef]
- Cao, J.; Lei, S.; Zhao, T.; Xie, Y.; Zhou, Z.; Cheng, S.; Wang, X. Changes in Fat Oxidation and Body Composition after Combined Exercise Intervention in Sedentary Obese Chinese Adults. J. Clin. Med. 2022, 11, 1086. [Google Scholar] [CrossRef]
- Péronnet, F.; Massicotte, D. Table of nonprotein respiratory quotient: An update. Can. J. Sport Sci. 1991, 16, 23–29. [Google Scholar]
- Faude, O.; Kindermann, W.; Meyer, T. Lactate threshold concepts: How valid are they? Sports Med. 2009, 39, 469–490. [Google Scholar] [CrossRef]
- Beneke, R. Maximal lactate steady state concentration (MLSS): Experimental and modelling approaches. Eur. J. Appl. Physiol. 2003, 88, 361–369. [Google Scholar] [CrossRef]
- Peric, R.; Nikolovski, Z.; Meucci, M.; Tadger, P.; Ferri Marini, C.; Amaro-Gahete, F.J. A Systematic Review and Meta-Analysis on the Association and Differences between Aerobic Threshold and Point of Optimal Fat Oxidation. Int. J. Environ. Res. Public Health 2022, 19, 6479. [Google Scholar] [CrossRef] [PubMed]
- Billat, V.; Sirvent, P.; Lepretre, P.M.; Koralsztein, J.P. Training effect on performance, substrate balance and blood lactate concentration at maximal lactate steady state in master endurance-runners. Pflug. Arch. 2004, 447, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Mader, A.; Heck, H. A theory of the metabolic origin of “anaerobic threshold”. Int. J. Sports Med. 1986, 7 (Suppl. 1), 45–65. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.C.; Jones, A.M. Oxygen uptake kinetics. Compr. Physiol. 2012, 2, 933–996. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Ppeak (W) | relPpeak (W·kg−1) | (mL·kg−1·min−1) | (mL·min−1) | BLCpeak (mmol·L−1) | RERpeak |
|---|---|---|---|---|---|
| 293.3 ± 39.4 (236.3/410.0) | 4.0 ± 0.6 (2.8/5.2) | 52.7 ± 7.6 (43.2/70.7) | 4505 ± 598 (3378/6213) | 9.4 ± 2.2 (4.9/13.4) | 1.16 ± 0.07 (1.10/1.29) |
| BLC2 (W) | BLC2 | BLC4 (W) | BLC4 |
|---|---|---|---|
| 169.3 ± 43.8 (101.8/307.5) | 62.8 ± 9.7 (41.5/82.6) | 235.2 ± 42.7 (177.2/369) | 82.8 ± 7.8 (72.9/100) |
| Dependent Variables | |||||
|---|---|---|---|---|---|
(mL·kg−1·min−1) | Ppeak (W) | BLC2 (W) | BLC4 (W) | kel (mmol·L−1)2 | |
| MFO (g·min−1) | 0.622 * | 0.570 * | 0.620 * | 0.660 * | 0.510 † |
| Fatmax (%Ppeak) | - | - | - | - | 0.472 † |
| COP (W) | 0.531 † | 0.720 ** | 0.720 ** | 0.760 ** | 0.440 † |
| COP (%Ppeak) | - | - | 0.480 † | 0.470 † | 0.554 * |
| Coefficients | Correlations | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Dependant Variables | Independent Variables | R | R2 | Adjusted R2 | ß | Sig. | Zero Order | Partial | Part |
| MFO (g·min−1) | kel | 0.897 | 0.804 | 0.783 | 0.663 | <0.001 | 0.510 | 0.826 | 0.649 |
| BLC2 | 0.754 | <0.001 | 0.619 | 0.858 | 0.738 | ||||
| Fatmax (%Ppeak) | kel | 0.472 | 0.223 | 0.182 | 0.472 | <0.05 | 0.472 | 0.472 | 0.472 |
| COP (W) | kel | 0.936 | 0.875 | 0.861 | 0.611 | <0.001 | 0.439 | 0.861 | 0.598 |
| BLC2 | 0.844 | <0.001 | 0.719 | 0.919 | 0.826 | ||||
| COP (%Ppeak) | kel | 0.819 | 0.671 | 0.634 | 0.680 | <0.001 | 0.554 | 0.757 | 0.665 |
| BLC2 | 0.616 | <0.001 | 0.478 | 0.725 | 0.603 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alkhatib, A. Maximal Fat Metabolism Explained by Lactate-Carbohydrate Model. Physiologia 2022, 2, 121-131. https://doi.org/10.3390/physiologia2040011
Alkhatib A. Maximal Fat Metabolism Explained by Lactate-Carbohydrate Model. Physiologia. 2022; 2(4):121-131. https://doi.org/10.3390/physiologia2040011
Chicago/Turabian StyleAlkhatib, Ahmad. 2022. "Maximal Fat Metabolism Explained by Lactate-Carbohydrate Model" Physiologia 2, no. 4: 121-131. https://doi.org/10.3390/physiologia2040011
APA StyleAlkhatib, A. (2022). Maximal Fat Metabolism Explained by Lactate-Carbohydrate Model. Physiologia, 2(4), 121-131. https://doi.org/10.3390/physiologia2040011