
Morpho- and Chemotyping of Holopelagic Sargassum Species Causing Massive Strandings in the Caribbean Region

 , , ,
, , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Morphological Analysis
2.2. Chemical Investigation
2.2.1. Sampling
2.2.2. Chemicals
2.2.3. Pigment Extraction and Analysis Using High Pressure Liquid Chromatography
2.2.4. Terpenes Extraction and Analysis Using Thin Layer Chromatography
2.2.5. Fatty Acid Extraction and Gas-Chromatography Analysis
2.2.6. Phenolic Compounds: Extraction, Purification, and Assay
2.2.7. Nuclear Magnetic Resonance (NMR) Analyses
2.3. Statistical Analyses
3. Results
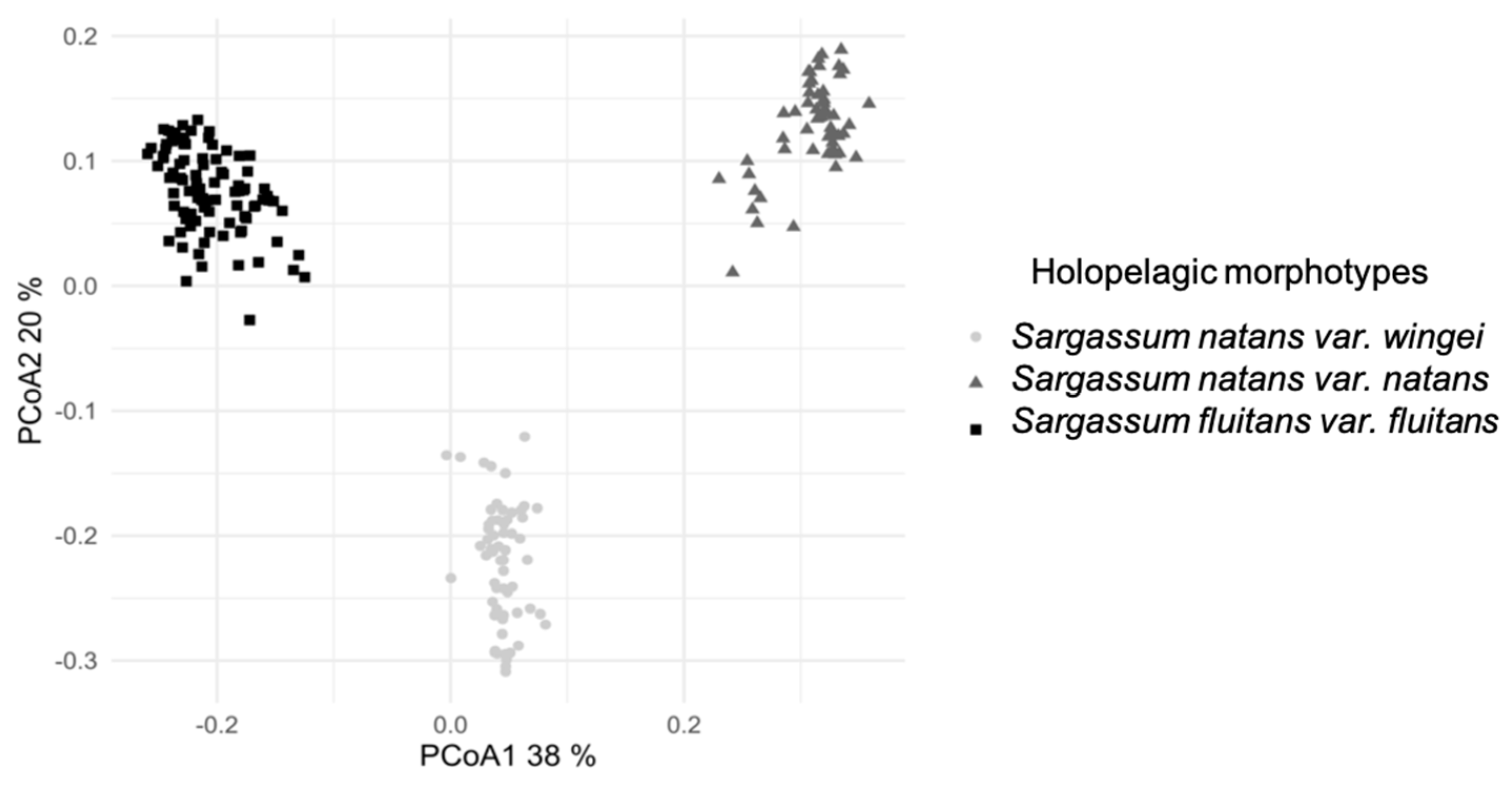
3.1. Morphological Identification of the Three Morphotypes
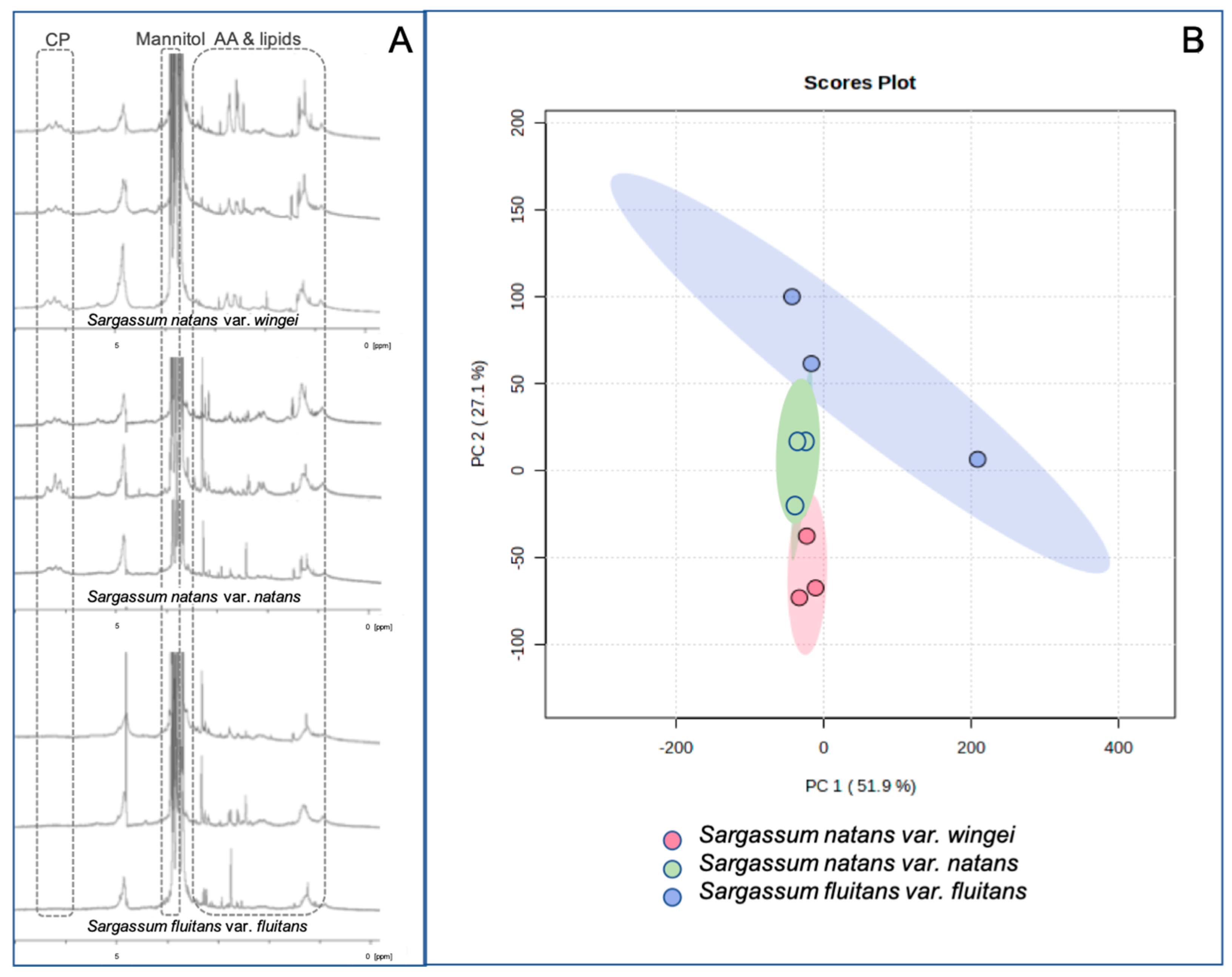
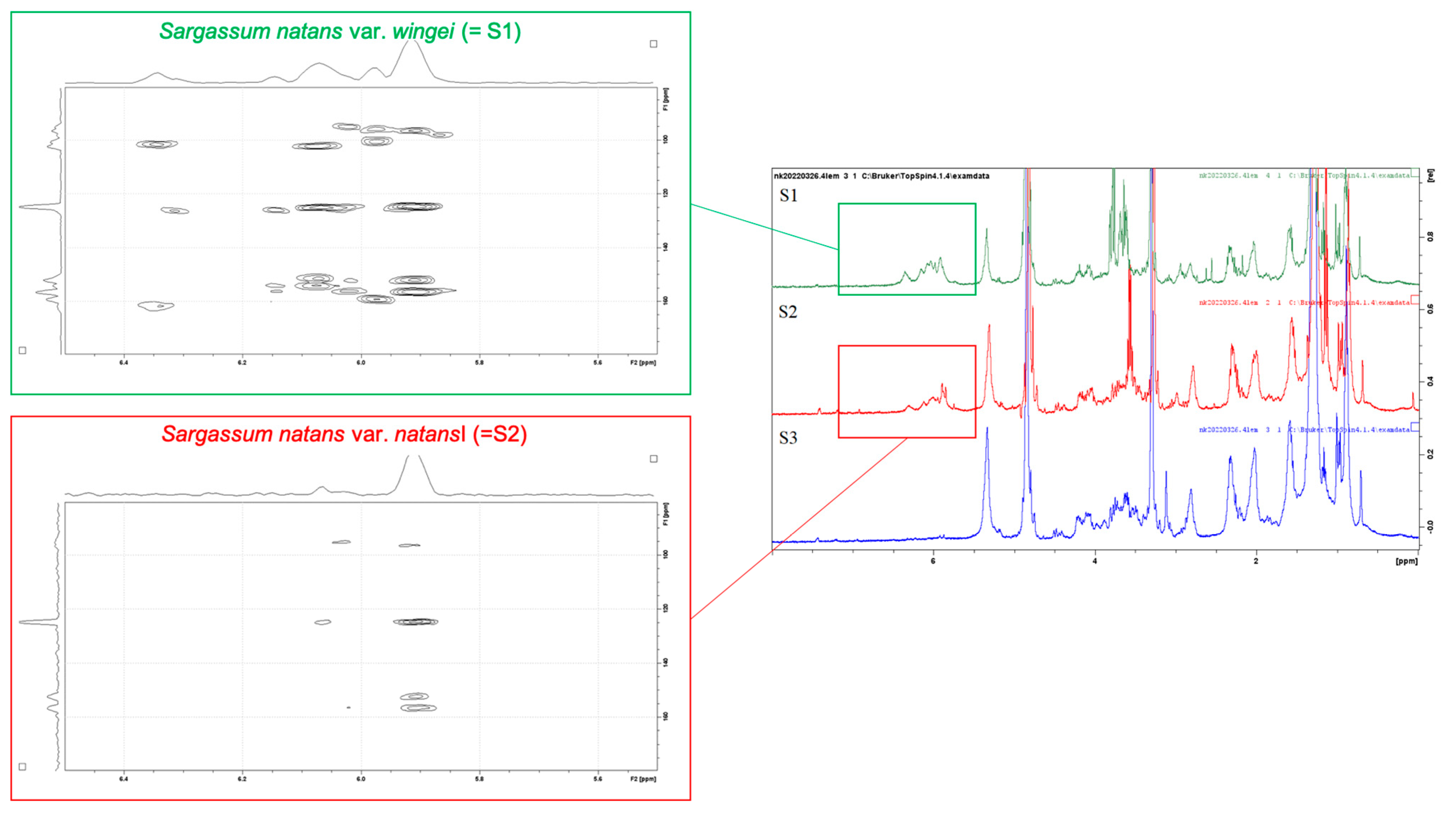
3.2. HR-MAS NMR Fingerprinting Profiles
3.3. Chemical Investigation
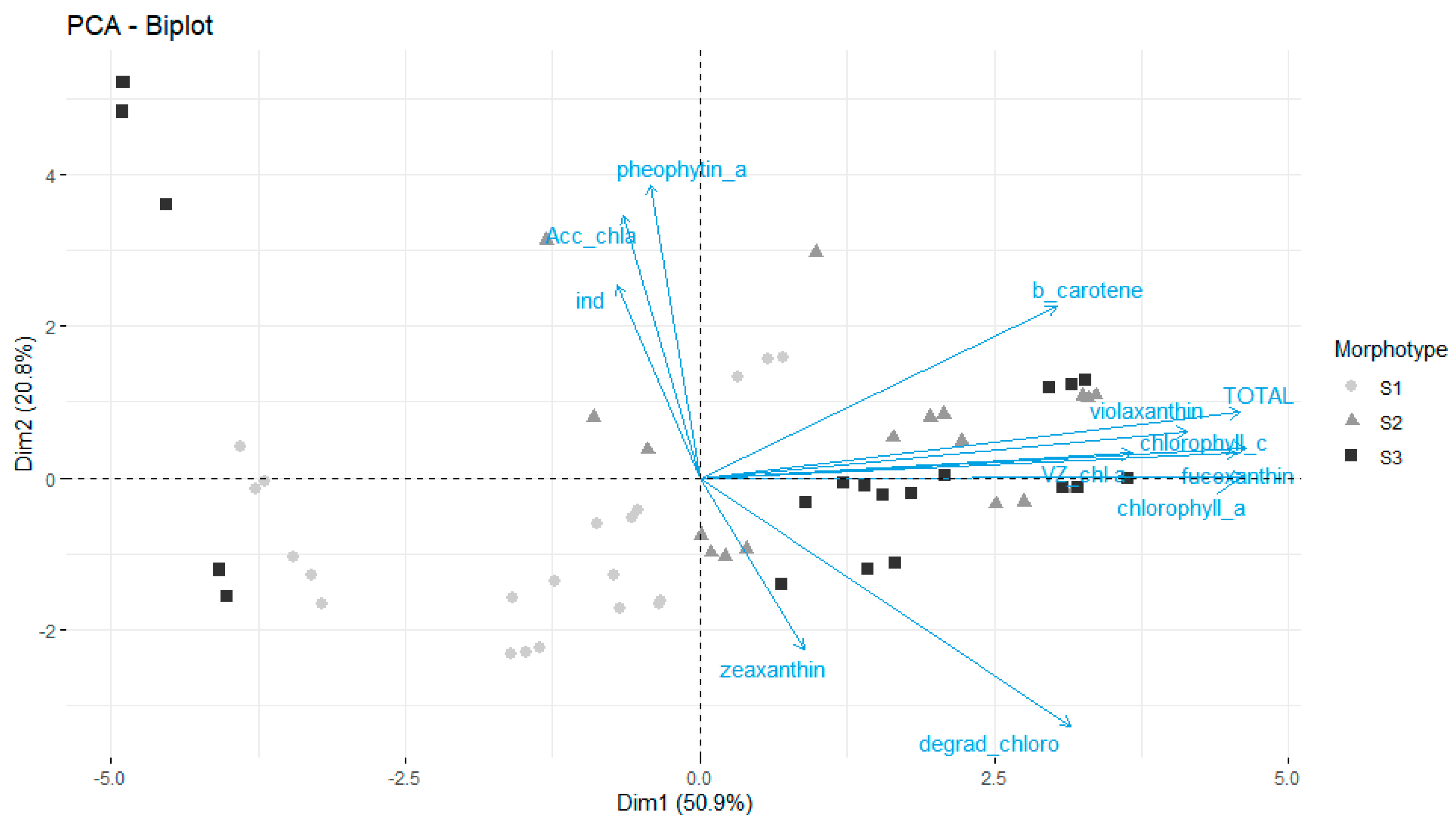
3.3.1. Pigment Level and Composition
3.3.2. Terpene Profiles
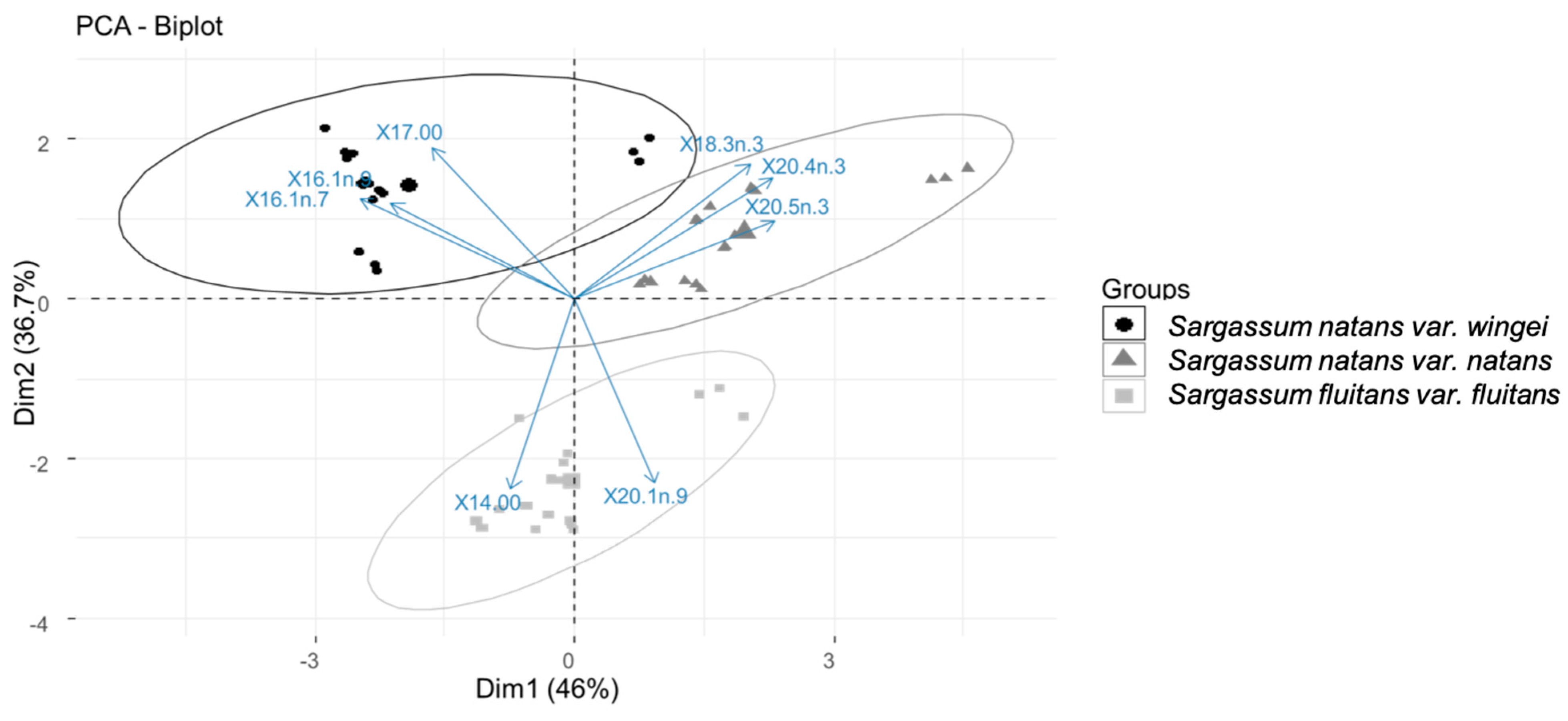
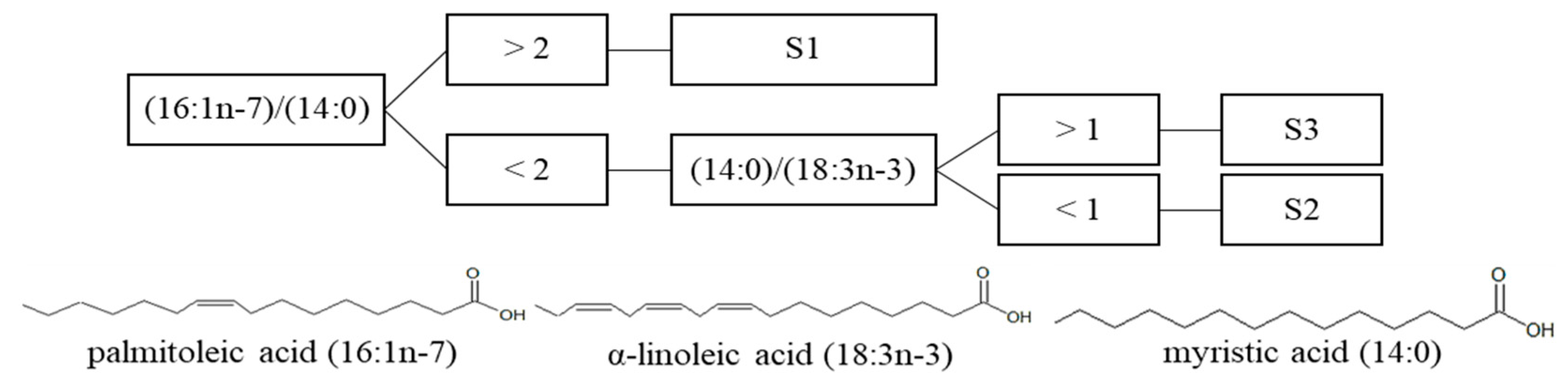
3.3.3. Free Fatty Acid Proportions
3.3.4. Total Phenolic Content and Structure of Phlorotannins
4. Discussion
4.1. Morphological Discrimination of Morphotypes
4.2. Chemical Discrimination of Morphotypes
4.2.1. HR-MAS Fingerprinting
4.2.2. Apolar/Lipidic Compounds: Pigments, Terpenes, and Fatty Acids
4.2.3. Phenolic Compounds
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Szechy, M.T.M.; Guedes, P.M.; Baeta-Neves, M.H.; Oliveira, E.N. Verification of Sargassum natans (Linnaeus) Gaillon (Heterokontophyta: Phaeophyceae) from the Sargasso Sea Off the Coast of Brazil, Western Atlantic Ocean. Checklist 2012, 8, 638–641. [Google Scholar] [CrossRef]
- Smetacek, V.; Zingone, A. Green and Golden Seaweed Tides on the Rise. Nature 2013, 504, 84–88. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. In World-Wide Electronic Publication; National University of Irlande: Galway, Ireland, 2013; Available online: http://www.algaebase.org (accessed on 18 April 2024).
- Stiger-Pouvreau, V.; Mattio, L.; De Ramon N’Yeurt, A.; Uwai, S.; Dominguez, H.; Flórez-Fernández, N.; Connan, S.; Critchley, A.T. A Concise Review of the Highly Diverse Genus Sargassum C. Agardh with Wide Industrial Potential. J. Appl. Phycol. 2023, 35, 1453–1483. [Google Scholar] [CrossRef]
- Amaral-Zettler, L.A.; Dragone, N.B.; Schell, J.; Slikas, B.; Murphy, L.G.; Morrall, C.E.; Zettler, E.R. Comparative Mitochondrial and Chloroplast Genomics of a Genetically Distinct Form of Sargassum Contributing to Recent “Golden Tides” in the Western Atlantic. Ecol. Evol. 2017, 7, 516–525. [Google Scholar] [CrossRef]
- Dibner, S.; Martin, L.; Thibaut, T.; Aurelle, D.; Blanfuné, A.; Whittaker, K.; Cooney, L.; Schell, J.M.; Goodwin, D.S.; Siuda, A.N.S. Consistent Genetic Divergence Observed among Pelagic Sargassum Morphotypes in the Western North Atlantic. Mar. Ecol. 2022, 43, e12691. [Google Scholar] [CrossRef]
- Schell, J.M.; Goodwin, D.S.; Siuda, A.N.S. Recent Sargassum Inundation Events in the Caribbean: Shipboard Observations Revea Dominance of a Previously Rare Form. Oceanography 2015, 28, 8–10. [Google Scholar] [CrossRef]
- Sissini, M.N.; De Barros Barreto, M.B.B.; Széchy, M.T.M.; De Lucena, M.B.; Oliveira, M.C.; Gower, J.; Liu, G.; De Oliveira Bastos, E.; Milstein, D.; Gusmão, F.; et al. The Floating Sargassum (Phaeophyceae) of the South Atlantic Ocean—Likely Scenarios. Phycologia 2017, 56, 321–328. [Google Scholar] [CrossRef]
- van Tussenbroek, B.I.; Hernández Arana, H.A.; Rodríguez-Martínez, R.E.; Espinoza-Avalos, J.; Canizales-Flores, H.M.; González-Godoy, C.E.; Barba-Santos, M.G.; Vega-Zepeda, A.; Collado-Vides, L. Severe impacts of brown tides caused by Sargassum spp. on near-shore Caribbean seagrass communities. Mar. Pollut. Bull. 2017, 122, 272–281. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, R.E.; Medina-Valmaseda, A.E.; Blanchon, P.; Monroy-Velázquez, L.V.; Almazán-Becerril, A.; Delgado-Pech, B.; Vásquez-Yeomans, L.; Francisco, V.; García-Rivas, M.C. Faunal mortality associated with massive beaching and decomposition of pelagic Sargassum. Mar. Pollut. Bull. 2019, 146, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Resiere, D.; Valentino, R.; Nevière, R.; Banydeen, R.; Gueye, P.; Florentin, J.; Cabié, A.; Lebrun, T.; Mégarbane, B.; Guerrier, G.; et al. Sargassum seaweed on Caribbean islands: An international public health concern. Lancet 2018, 392, 2691. [Google Scholar] [CrossRef] [PubMed]
- Changeux, T. Sargasses transatlantiques cruise, RV Yersin. Fr. Oceanogr. Cruises 2017. [Google Scholar] [CrossRef]
- Thibaut, T. Sargasses. Fr. Oceanogr. Cruises 2017. [Google Scholar] [CrossRef]
- Winge, Ø. The Sargasso Sea, Its Boundaries and Vegetation. Report on the Danish Oceanographical Expeditions 1908–1910 to the Mediterranean and Adjacent Seas; A.F. Høst&søn.: Copenhagen, Denmark, 1923. [Google Scholar]
- Parr, A.E. Quantitative Observations on the Pelagic Sargassum Vegetation of the Western North Atlantic. Bull. Bingham Oceanogr. Collect. 1939, 6, 1–94. [Google Scholar]
- Siuda, A.N.S.; Blanfuné, A.; Dibner, S.; Verlaque, M.; Boudouresque, C.-F.; Connan, S.; Goodwin, D.S.; Stiger-Pouvreau, V.; Viard, F.; Rousseau, F.; et al. Morphological and Molecular Characters Differentiate Common Morphotypes of Atlantic Holopelagic Sargassum. Phycology 2024, 4, 256–275. [Google Scholar] [CrossRef]
- Pellegrini, M.; Valls, R.; Pellegrini, L. Chemotaxonomy and chemical markers in brown algae (in French). Lagascalia 1997, 19, 145–164. [Google Scholar]
- Le Lann, K.; Rumin, J.; Cérantola, S.; Culioli, G.; Stiger-Pouvreau, V. Spatiotemporal variations of diterpene production in the brown macroalga Bifurcaria bifurcata from the western coasts of Brittany (France). J. Appl. Phycol. 2014, 26, 1207–1214. [Google Scholar] [CrossRef]
- Jégou, C.; Culioli, G.; Stiger-Pouvreau, V. Meroditerpene from Cystoseira nodicaulis and its taxonomic significance. Biochem. Syst. Ecol. 2012, 44, 202–204. [Google Scholar] [CrossRef]
- Hernández-Bolio, G.I.; Fagundo-Mollineda, A.; Caamal-Fuentes, E.E.; Robledo, D.; Freile-Pelegrin, Y.; Hernández-Núñez, E. NMR metabolic profiling of Sargassum species under different stabilization/extraction processes. J. Phycol. 2021, 57, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Simon, G.; Kervarec, N.; Cérantola, S. HRMAS NMR analysis of algae and identification of molecules of interest via conventional 1D and 2D NMR: Sample preparation and optimization of experimental conditions. Methods Mol. Biol. 2015, 1308, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Le Lann, K.; Kraffe, E.; Kervarec, N.; Cérantola, S.; Payri, C.E.; Stiger-Pouvreau, V. Isolation of turbinaric acid as a chemomarker of Turbinaria conoides (J. Agardh) Kützing from South Pacific Islands. J. Phycol. 2014, 50, 1048–1057. [Google Scholar] [CrossRef]
- Le Lann, K.; Kervarec, N.; Payri, C.E.; Deslandes, E.; Stiger-Pouvreau, V. Discrimination of allied species within the genus Turbinaria (Fucales, Phaeophyceae) using HRMAS NMR spectroscopy. Talanta 2008, 74, 1079–1083. [Google Scholar] [CrossRef]
- Jégou, C.; Culioli, G.; Kervarec, N.; Simon, G.; Stiger-Pouvreau, V. LC/ESI-MSn and 1H HR-MAS NMR analytical methods as useful taxonomical tools within the genus Cystoseira C. Agardh (Fucales; Phaeophyceae). Talanta 2010, 83, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Tanniou, A.; Vandanjon, L.; Gonçalves, O.; Kervarec, N.; Stiger-Pouvreau, V. Rapid geographical differentiation of the European spread brown macroalga Sargassum muticum using HRMAS NMR and Fourier-Transform Infrared spectroscopy. Talanta 2015, 132, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Jégou, C.; Kervarec, N.; Cérantola, S.; Bihannic, I.; Stiger-Pouvreau, V. NMR use to quantify phlorotannins. The case of Cystoseira tamariscifolia, a phloroglucinol-producing brown macroalga in Brittany (France). Talanta 2015, 135, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Gobert, T.; Gautier, A.; Connan, S.; Rouget, M.L.; Thibaut, T.; Stiger-Pouvreau, V.; Waeles, M. Trace metal content from holopelagic Sargassum spp. sampled in the tropical North Atlantic Ocean: Emphasis on spatial variation of arsenic and phosphorus. Chemosphere 2022, 308, 136186. [Google Scholar] [CrossRef] [PubMed]
- Lalegerie, F.; Lajili, S.; Bedoux, G.; Taupin, L.; Stiger-Pouvreau, V.; Connan, S. Photo-protective compounds in red macroalgae from Brittany: Considerable diversity in mycosporine-like amino acids (MAAs). Mar. Environ. Res. 2019, 147, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Stengel, D.B. Intra-thallus differentiation of fatty acid and pigment profiles in some temperate Fucales and Laminariales. J. Phycol. 2015, 51, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Mathieu-Resuge, M.; Le Grand, F.; Brosset, P.; Lebigre, C.; Soudant, P.; Vagner, M.; Pecquerie, L.; Sardenne, F. Red muscle of small pelagic fishes’ fillets are high-quality sources of essential fatty acids. J. Food Compos. Anal. 2023, 120, 105304. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Sardenne, F.; Puccinelli, E.; Vagner, M.; Pecquerie, L.; Bideau, A.; Le Grand, F.; Soudant, P. Post-mortem storage conditions and cooking methods affect long-chain omega-3 fatty acid content in Atlantic mackerel (Scomber scombrus). Food Chem. 2021, 359, 129828. [Google Scholar] [CrossRef]
- Mathieu-Resuge, M.; Kraffe, E.; Le Grand, F.; Boens, A.; Bideau, A.; Lluch-Cota, S.E.; Racotta, I.S.; Schaal, G. Trophic ecology of suspension-feeding bivalves inhabiting a north-eastern Pacific coastal lagoon: Comparison of different biomarkers. Mar. Environ. Res. 2019, 145, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Ar Gall, E.; Lelchat, F.; Hupel, M.; Jégou, C.; Stiger-Pouvreau, V. Extraction and purification of phlorotannins from brown algae. Methods Mol. Biol. 2015, 1308, 131–143. [Google Scholar] [CrossRef]
- Stiger-Pouvreau, V.; Jégou, C.; Cérantola, S.; Guérard, F.; Le Lann, K. Chapter 13—Phlorotannins in Sargassaceae species from Brittany (France): Interesting molecules for ecophysiological and valorisation purposes. In Advances in Botanical Research; Bourgougnon, N., Ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: Cambridge, MA, USA, 2014; pp. 379–411. [Google Scholar]
- Zubia, M.; Fabre, M.S.; Kerjean, V.; Le Lann, K.; Stiger-Pouvreau, V.; Fauchon, M.; Deslandes, E. Antioxidant and antitumoural activities of some Phaeophyta from Brittany coasts. Food Chem. 2009, 116, 693–701. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, J.; Shen, J.; Silva, A.; Dennis, D.A.; Barrow, C.J. A simple 96-well microplate method for estimation of total polyphenol content in seaweeds. J. Appl. Phycol. 2006, 18, 445–450. [Google Scholar] [CrossRef]
- Cérantola, S.; Breton, F.; ar Gall, E.; Deslandes, E. Co-occurrence and antioxidant activities of fucol and fucophlorethol classes of polymeric phenols in Fucus spiralis. Bot. Mar. 2006, 49, 347–351. [Google Scholar] [CrossRef]
- Jégou, C.; Connan, S.; Bihannic, I.; Cérantola, S.; Guérard, F.; Stiger-Pouvreau, V. Phlorotannin and Pigment Content of Native Canopy-Forming Sargassaceae Species Living in Intertidal Rockpools in Brittany (France): Any Relationship with Their Vertical Distribution and Phenology? Mar. Drugs 2021, 19, 504–523. [Google Scholar] [CrossRef]
- Posit Team. RStudio: Integrated Development Environment for R; Posit: Hong Kong, China, 2023. [Google Scholar]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K. Cluster: Cluster Analysis Basics and Extensions; R Package Version; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Bache, S.M.; Wickham, H. Magrittr: A Forward-Pipe Operator for R. 2020. Available online: https://cran.r-project.org/web/packages/magrittr/index.html (accessed on 18 April 2024).
- Wickham, H. Tidyr: Tidy Messy Data. 2021. Available online: https://tidyr.tidyverse.org/ (accessed on 18 April 2024).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O'Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. 2020. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 18 April 2024).
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in {R}. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef] [PubMed]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. 2020. Available online: https://rpkgs.datanovia.com/factoextra/ (accessed on 18 April 2024).
- Lê, S.; Josse, J.; Husson, F. {FactoMineR}: A package for multivariate analysis. J. Stat. Software 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Mangiafico, S. Rcompanion: Functions to Support Extension Education Program Evaluation. 2022. Available online: https://www.researchgate.net/publication/307608990_rcompanion_Functions_to_support_extension_education_program_evaluation_R_statistical_package (accessed on 18 April 2024).
- Dinno, A. Dunn.test: Dunn’s Test of Multiple Comparisons Using Rank Sums. 2017. Available online: https://cran.r-project.org/web/packages/dunn.test/dunn.test.pdf (accessed on 18 April 2024).
- Davis, D.; Simister, R.; Campbell, S.; Marston, M.; Bose, S.; McQueen-Mason, S.J.; Gomez, L.D.; Gallimore, W.A.; Tonon, T. Biomass composition of the golden tide pelagic seaweeds Sargassum fluitans and S. natans (morphotypes I and VIII) to inform valorisation pathways. Sci. Total Environ. 2021, 762, 143134. [Google Scholar] [CrossRef]
- Robledo, D.; Vázquez-Delfín, E.; Freile-Pelegrín, Y.; Vásquez-Elizondo, R.M.; Qui-Minet, Z.N.; Salazar-Garibay, A. Challenges and opportunities in relation to Sargassum events along the Caribbean Sea. Front. Mar. Sci. 2021, 8, 1000–1012. [Google Scholar] [CrossRef]
- Franks, J.S.; Johnson, D.R.; Ko, D.S. Pelagic Sargassum in the Tropical North Atlantic. Gulf Carib. Res. 2016, 27, 6–11. [Google Scholar] [CrossRef]
- Podlejski, W.; Berline, L.; Nerini, D.; Doglioli, A.; Lett, C. A new Sargassum drift model derived from features tracking in MODIS images. Mar. Poll. Bull. 2023, 188, 114629. [Google Scholar] [CrossRef]
- Putman, N.F.; Lumpkin, R.; Olascoaga, M.J.; Trinanes, J.; Goni, G.J. Improving transport predictions of pelagic Sargassum. J. Exp. Mar. Biol. Ecol. 2020, 529, 151398. [Google Scholar] [CrossRef]
- Stiger, V.; Horiguchi, T.; Yoshida, T.; Coleman, A.W.; Masuda, M. Phylogenetic Relationships of Sargassum (Sargassaceae, Phaeophyceae) with Reference to a Taxonomic Revision of the Section Phyllocystae Based on ITS-2 nrDNA Sequences. Phycol. Res. 2000, 48, 251–260. [Google Scholar] [CrossRef]
- Yoshida, T.; Stiger, V.; Horiguchi, T. Sargassum boreale sp. Nov. (Fucales, Phaeophyceae) from Hokkaido, Japan. Phycol. Res. 2000, 48, 125–131. [Google Scholar] [CrossRef]
- Salaün, S.; Kervarec, N.; Potin, P.; Haras, D.; Piotto, M.; La Barre, S. Whole-cell spectroscopy is a convenient tool to assist molecular identification of cultivatable marine bacteria and to investigate their adaptive metabolism. Talanta 2010, 80, 1758–1770. [Google Scholar] [CrossRef]
- Obando, C.Z.; Linossier, I.; Kervarec, N.; Zubia, M.; Turquet, J.; Faÿ, F.; Rehel, K. Rapid identification of osmolytes in tropical microalgae and cyanobacteria by 1H HR-MAS NMR spectroscopy. Talanta 2016, 153, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Alcantara, G.B.; Barison, A.; Ferreira, A.G.; Honda, N.K.; Ferreira, M.M.C. Chemotaxonomic Discrimination of Lichens by 1H NMR, solution and HR-MAS, and Chemometric Analysis. Ann. Magn. Reson. 2006, 5, 1–4. [Google Scholar]
- Dutra, L.M.; da Conceicao Santos, A.D.; Lourenço, A.V.F.; Nagata, N.; Heiden, G.; Campos, F.R.; Barison, A. 1H HR-MAS NMR and chemometric methods for discrimination and classification of Baccharis (Asteraceae): A proposal for quality control of Baccharis trimera. J. Pharm. Biomed. Anal. 2020, 184, 113200. [Google Scholar] [CrossRef]
- Stengel, D.B.; Connan, S.; Popper, Z.A. Algal chemodiversity and bioactivity: Sources of natural variability and implications for commercial application. Biotechnol. Adv. 2011, 29, 483–501. [Google Scholar] [CrossRef]
- Van Ginneken, V.J.T.; Helsper, J.P.F.G.; De Visser, W.; van Keulen, H.; Brandenburg, W.A. Polyunsaturated fatty acids in various macroalgal species from north Atlantic and tropical seas. Lipids Health Dis. 2011, 10, 104. [Google Scholar] [CrossRef] [PubMed]
- Milledge, J.J.; Maneein, S.; López, E.A.; Bartlett, D. Sargassum inundations in Turks and Caicos: Methane potential and proximate, ultimate, lipid, amino acid, metal and metalloid analyses. Energies 2020, 13, 1523. [Google Scholar] [CrossRef]
- Turner, J.P.; Rooker, J.R. Fatty acid composition of flora and fauna associated with Sargassum mats in the Gulf of Mexico. Mar. Biol. 2006, 149, 1025–1036. [Google Scholar] [CrossRef]
- Lalegerie, F.; Gager, L.; Stiger-Pouvreau, V.; Connan, S. The stressful life of red and brown seaweeds on the temperate intertidal zone: Effect of abiotic and biotic parameters on the physiology of macroalgae and content variability of particular metabolites. Adv. Bot. Res. 2020, 95, 247–287. [Google Scholar] [CrossRef]
- Stiger-Pouvreau, V.; Deslandes, E.; Payri, C.E. Phenolic contents of two brown algae, Turbinaria ornata and Sargassum mangarevense on Tahiti (French Polynesia): Interspecific, ontogenic and spatio-temporal variations. Bot. Mar. 2004, 47, 402–409. [Google Scholar] [CrossRef]
- Nakai, M.; Kageyama, N.; Nakahara, K.; Miki, W. Phlorotannins as radical scavengers from the extract of Sargassum ringgoldianum. Mar. Biotechnol. 2006, 8, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Shen, P.; Gu, Y.; Zhang, C.; Sun, C.; Qin, L.; Yu, C.; Qi, H. Metabolomic approach for characterization of polyphenolic compounds in Laminaria japonica, Undaria pinnatifida, Sargassum fusiforme and Ascophyllum nodosum. Foods 2021, 10, 192. [Google Scholar] [CrossRef]
- Taniguchi, R.; Ito, C.; Keitoku, S.; Miyake, Y.; Itoigawa, M.; Matsui, T.; Shibata, T. Analysis on the structure of phlorethols isolated from the warm-temperate brown seaweed Sargassum carpophyllum and their antioxidant properties. Nat. Prod. Com. 2022, 17, 1934578X221109406. [Google Scholar] [CrossRef]
- Urrea-Victoria, V.; Furlan, C.M.; dos Santos, D.Y.A.C.; Chow, F. Antioxidant potential of two Brazilian seaweeds in response to temperature: Pyropia spiralis (red alga) and Sargassum stenophyllum (brown alga). J. Exp. Mar. Biol. Ecol. 2022, 549, 151706. [Google Scholar] [CrossRef]
- Agregán, R.; Munekata, P.E.S.; Franco, D.; Dominguez, R.; Carballo, J.; Lorenzo, J.M. Phenolic compounds from three brown seaweed species using LC-DAD–ESI-MS/MS. Food Res. Internat. 2017, 99, 979–985. [Google Scholar] [CrossRef]
- Glombitza, K.W.; Forster, M.; Farnham, W.F. Antibiotics from algae—Part 25: Polyhydroxyphenyl ethers from the brown alga Sargassum muticum (Yendo) Fensholt, Part II. Bot. Mar. 1982, 25, 449–454. [Google Scholar] [CrossRef]
- Montero, L.; Sánchez-Camargo, A.P.; García-Cañas, V.; Tanniou, A.; Stiger-Pouvreau, V.; Russo, M.; Rastrelli, L.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Anti-proliferative activity and chemical characterization by comprehensive two-dimensional liquid chromatography coupled to mass spectrometry of phlorotannins from the brown macroalga Sargassum muticum collected on North-Atlantic coasts. J. Chrom. A 2016, 1428, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Tanniou, A.; Vandanjon, L.; Incera, M.; Serrano Leon, E.; Husa, V.; Le Grand, J.; Nicolas, J.-L.; Poupart, N.; Kervarec, N.; Stiger-Pouvreau, V.; et al. Assessment of the spatial variability of phenolic contents and associated bioactivities in the invasive alga Sargassum muticum sampled along its European range from Norway to Portugal. J. Appl. Phycol. 2014, 26, 1215–1230. [Google Scholar] [CrossRef]
- Catarino, M.D.; Pires, S.M.G.; Silva, S.; Costa, F.; Braga, S.S.; Pinto, D.C.G.A.; Silva, A.M.S.; Cardoso, S.M. Overview of Phlorotannins’ Constituents in Fucales. Mar. Drugs 2022, 20, 754. [Google Scholar] [CrossRef] [PubMed]
- Chouh, A.; Nouadri, T.; Catarino, M.D.; Silva, A.M.; Cardoso, S.M. Phlorotannins of the brown algae Sargassum vulgare from the Mediterranean Sea coast. Antioxidants 2022, 11, 1055. [Google Scholar] [CrossRef] [PubMed]
- Glombitza, K.W.; Keusgen, M.; Hauperich, S. Fucophlorethols from the brown algae Sargassum spinuligerum and Cystophora torulosa. Phytochemistry 1997, 46, 1417–1422. [Google Scholar] [CrossRef]
- Glombitza, K.W.; Keusgen, M. Fuhalols and deshydroxyfuhalols from the brown alga Sargassum spinuligerum. Phytochemistry 1995, 38, 987–995. [Google Scholar] [CrossRef]
- Keusgen, M.; Glombitza, K.W. Pseudofuhalols from the brown alga Sargassum spinuligerum. Phytochemistry 1997, 46, 1403–1415. [Google Scholar] [CrossRef]
- Keusgen, M.; Glombitza, K.W. Phlorethols, fuhalols and their derivatives from the brown alga Sargassum spinuligerum. Phytochemistry 1995, 38, 975–985. [Google Scholar] [CrossRef]
- Glombitza, K.W.; Hauperich, S.; Keusgen, M. Phlorotannins from the brown algae Cystophora torulosa and Sargassum spinuligerum. Nat. Tox. 1997, 5, 58–63. [Google Scholar] [CrossRef]
- Li, Y.; Fu, X.; Duan, D.; Liu, X.; Xu, J.; Gao, X. Extraction and identification of phlorotannins from the brown alga, Sargassum fusiforme (Harvey) Setchell. Mar. Drugs 2017, 15, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Changeux, T.; Berline, L.; Podlejski, W.; Guillot, T.; Stiger-Pouvreau, V.; Connan, S.; Thibaut, T. Variability in growth and tissue composition (CNP, natural isotopes) of the three morphotypes of holopelagic Sargassum. Aq. Bot. 2023, 187, 103644. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Environment | Latitude | Longitude |
|---|---|---|---|
| Station 4 | Pelagic raft in open water | 08°51.950 N | 49°08.380 W |
| Station 9 | Pelagic raft in open water | 12°46.160 N | 55°31.040 W |
| Mangrove | Stranded in Martinique Mangrove | 14°33.002 N | 61°00.371 W |
| Station GP | Pelagic raft in open water | 15°04.161 N | 34°06.223 W |
| Station 12 | Pelagic raft in open water | 15°57.270 N | 61°59.300 W |
| Station 23 | Pelagic raft in open water | 16°07.366 N | 61°57.353 W |
| Station 15 | Pelagic raft in open water | 17°19.134 N | 59°36.044 W |
| Station 19 | Pelagic raft in open water | 23°01.740 N | 59°13.367 W |
| (A) | |
| Chemical shift (ppm) | Carbon signals |
| 95–100 | Methine carbons |
| 100–105 | Aryl-aryl carbons |
| 125–130 | Diaryl-ether carbons |
| 145–150 | Additional hydroxyl functions |
| 150–165 | Phenolic carbons |
| (B) | |
| Type of bond | Class of phlorotannin |
| Ether | Phlorethols Fuhalols, if additional hydroxyl groups |
| Phenyl | Fucols |
| Ether and Phenyl | Fucophlorethol |
| Dibenzodioxin | Eckols |
| Sargassum natans var. wingei | Sargassum natans var. natans | Sargassum fluitans var. fluitans | |
|---|---|---|---|
| C14:0 | 2.96 ± 0.20% | 2.63 ± 0.30% | 4.17 ± 0.32% |
| C15:0 | 0.36 ± 0.11% | 0.40 ± 0.06% | 0.52 ± 0.11% |
| C16:0 | 47.91 ± 3.31% | 45.80 ± 3.57% | 47.53 ± 3.90% |
| C16:1n-7 | 0.43 ± 0.09% | 0.26 ± 0.05% | 0.28 ± 0.07% |
| C16:1n-7 | 10.00 ± 1.53% | 4.53 ± 0.46% | 5.22 ± 0.54% |
| C17:0 | 0.38 ± 0.05% | 0.14 ± 0.03% | 0.12 ± 0.04% |
| C17:1n-7 | 0.17 ± 0.06% | 0.50 ± 0.18% | 0.42 ± 0.23% |
| C18:0 | 1.36 ± 0.51% | 2.35 ± 0.71% | 2.32 ± 0.86% |
| C18:1n-9 | 11.85 ± 0.57% | 13.75 ± 1.05% | 15.08 ± 1.76% |
| C18:1n-7 | 0.65 ± 0.45% | 0.82 ± 0.46% | 0.81 ± 0.55% |
| C18:2n-6 | 6.41 ± 0.91% | 5.37 ± 0.67% | 4.65 ± 0.55% |
| C18:4n-3 | 0.53 ± 0.35% | 1.08 ± 0.44% | 0.53 ± 0.28% |
| C18:3n-3 | 2.21 ± 0.53% | 3.69 ± 0.65% | 1.82 ± 0.46% |
| C20:0 | 0.44 ± 0.21% | 0.78 ± 0.21% | 0.83 ± 0.30% |
| C20:1n-9 | 0.83 ± 0.14% | 1.05 ± 0.27% | 1.60 ± 0.21% |
| C20:2n-6 | 0.33 ± 0.03% | 0.34 ± 0.09% | 0.37 ± 0.07% |
| C20:3n-6 | 0.54 ± 0.17% | 0.48 ± 0.07% | 0.52 ± 0.16% |
| C20:4n-6 | 7.52 ± 2.66% | 8.75 ± 1.50% | 6.98 ± 1.78% |
| C20:4n-3 | 0.32 ± 0.16% | 0.53 ± 0.13% | 0.26 ± 0.12% |
| C20:5n-3 | 0.66 ± 0.53% | 1.55 ± 0.97% | 0.69 ± 0.63% |
| C22:0 | 0.61 ± 0.10% | 1.24 ± 0.26% | 1.40 ± 0.29% |
| C22:1n-9 | 1.37 ± 0.29% | 1.08 ± 0.34% | 1.35 ± 0.40% |
| C22:1n-7 | 0.17 ± 0.08% | 0.19 ± 0.10% | 0.26 ± 0.16% |
| C22:6n-3 | 0.55 ± 0.17% | 0.62 ± 0.16% | 0.84 ± 0.19% |
| C24:0 | 0.61 ± 0.57% | 1.34 ± 1.63% | 1.10 ± 1.45% |
| SFA | 54.64 ± 2.44% | 54.68 ± 1.66% | 58.00 ± 2.31% |
| MUFA | 25.49 ± 2.25% | 22.18 ± 1.50% | 25.03 ± 2.07% |
| PUFA | 19.08 ± 4.37% | 22.42 ± 3.02% | 16.67 ± 4.15% |
| Station (See Table 1) | Morphotype | TPC (mg·g−1 DW) |
|---|---|---|
| 4 | S1 | 50.31 ± 6.60 |
| S2 | 27.52 ± 3.59 | |
| S3 | 13.15 ± 1.43 | |
| 9 | S1 | 31.60 ± 2.45 |
| S2 | 15.42 ± 0.77 | |
| S3 | 11.45 ± 0.76 | |
| Mangrove | S1 | 2.35 ± 0.41 |
| S2 | 2.26 ± 0.81 | |
| S3 | 2.12 ± 0.67 | |
| GP | S1 | 7.55 ± 0.47 |
| S2 | 9.73 ± 1.18 | |
| S3 | 9.48 ± 1.35 | |
| 12 | S1 | 26.12 ± 3.88 |
| S2 | 11.35 ± 1.79 | |
| S3 | 4.98 ± 0.42 | |
| 23 | S1 | 19.53 ± 0.46 |
| S2 | 19.05 ± 2.81 | |
| S3 | 14.82 ± 1.56 | |
| 15 | S1 | 11.35 ± 1.79 |
| S2 | 17.75 ± 1.73 | |
| S3 | 6.53 ± 0.31 | |
| 19 | S1 | 41.56 ± 2.69 |
| S2 | 32.34 ± 3.44 | |
| S3 | 6.53 ± 1.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kergosien, N.; Helias, M.; Le Grand, F.; Cérantola, S.; Simon, G.; Nirma, C.; Thibaut, T.; Berline, L.; Changeux, T.; Blanfuné, A.; et al. Morpho- and Chemotyping of Holopelagic Sargassum Species Causing Massive Strandings in the Caribbean Region. Phycology 2024, 4, 340-362. https://doi.org/10.3390/phycology4030018
Kergosien N, Helias M, Le Grand F, Cérantola S, Simon G, Nirma C, Thibaut T, Berline L, Changeux T, Blanfuné A, et al. Morpho- and Chemotyping of Holopelagic Sargassum Species Causing Massive Strandings in the Caribbean Region. Phycology. 2024; 4(3):340-362. https://doi.org/10.3390/phycology4030018
Chicago/Turabian StyleKergosien, Nolwenn, Mathieu Helias, Fabienne Le Grand, Stéphane Cérantola, Gaëlle Simon, Charlotte Nirma, Thierry Thibaut, Léo Berline, Thomas Changeux, Aurélie Blanfuné, and et al. 2024. "Morpho- and Chemotyping of Holopelagic Sargassum Species Causing Massive Strandings in the Caribbean Region" Phycology 4, no. 3: 340-362. https://doi.org/10.3390/phycology4030018
APA StyleKergosien, N., Helias, M., Le Grand, F., Cérantola, S., Simon, G., Nirma, C., Thibaut, T., Berline, L., Changeux, T., Blanfuné, A., Connan, S., & Stiger-Pouvreau, V. (2024). Morpho- and Chemotyping of Holopelagic Sargassum Species Causing Massive Strandings in the Caribbean Region. Phycology, 4(3), 340-362. https://doi.org/10.3390/phycology4030018