
Legacy of “New Normal” Plastics and “New Nitrogen” in the Cyanotoxin Footprint in Mangrove Ecosystems


Abstract
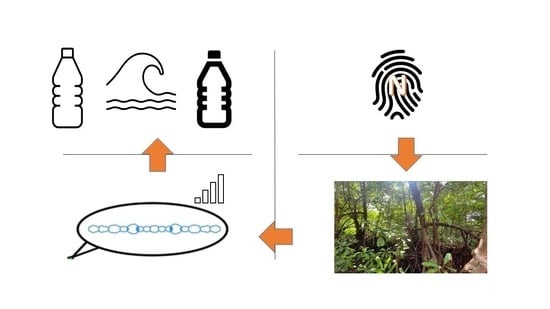
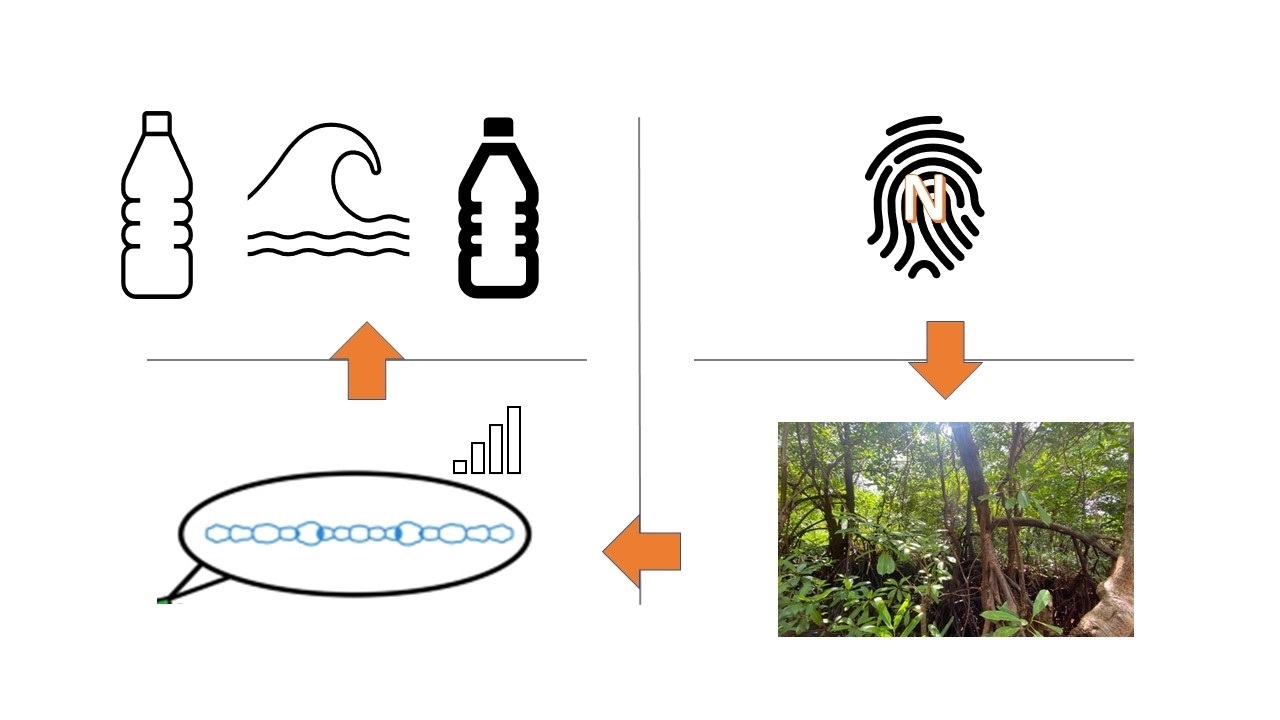
1. Cyanobacteria in Mangrove Ecosystems
- To detail the footprint of cyanotoxin production in mangrove ecosystems.
- To question the ambiguity in cyanotoxin production in relation to N:P ratios and biological nitrogen fixation.
- To elucidate the landscape of plastic-opulent estuarine mangroves with the emerging role of the plastisphere on cyanobacterial biofilm formation, buoyancy, sinking and dispersal.
- To examine the articles of the Ramsar Convention that are impacted by the emergence of diverse types of plastic pollution in wetlands and their downstream reverberations.
- To showcase the nexus between N-fixation and plastic pollution that exacerbates the putative cyanotoxin footprint in mangroves.
- Navigating the path ahead: what science should ideally do in the face of plastic pollution in mangroves and associated wetlands.
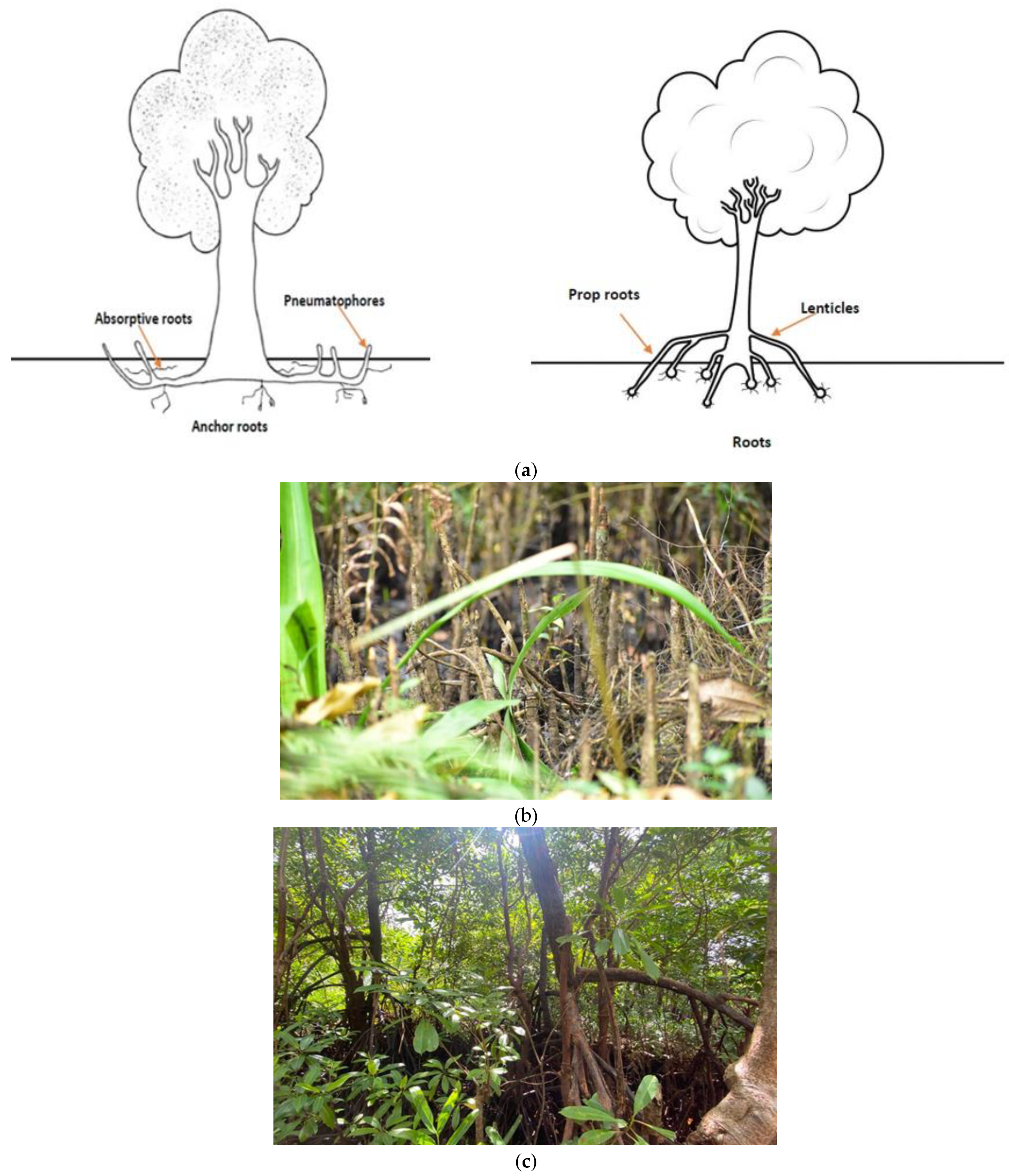
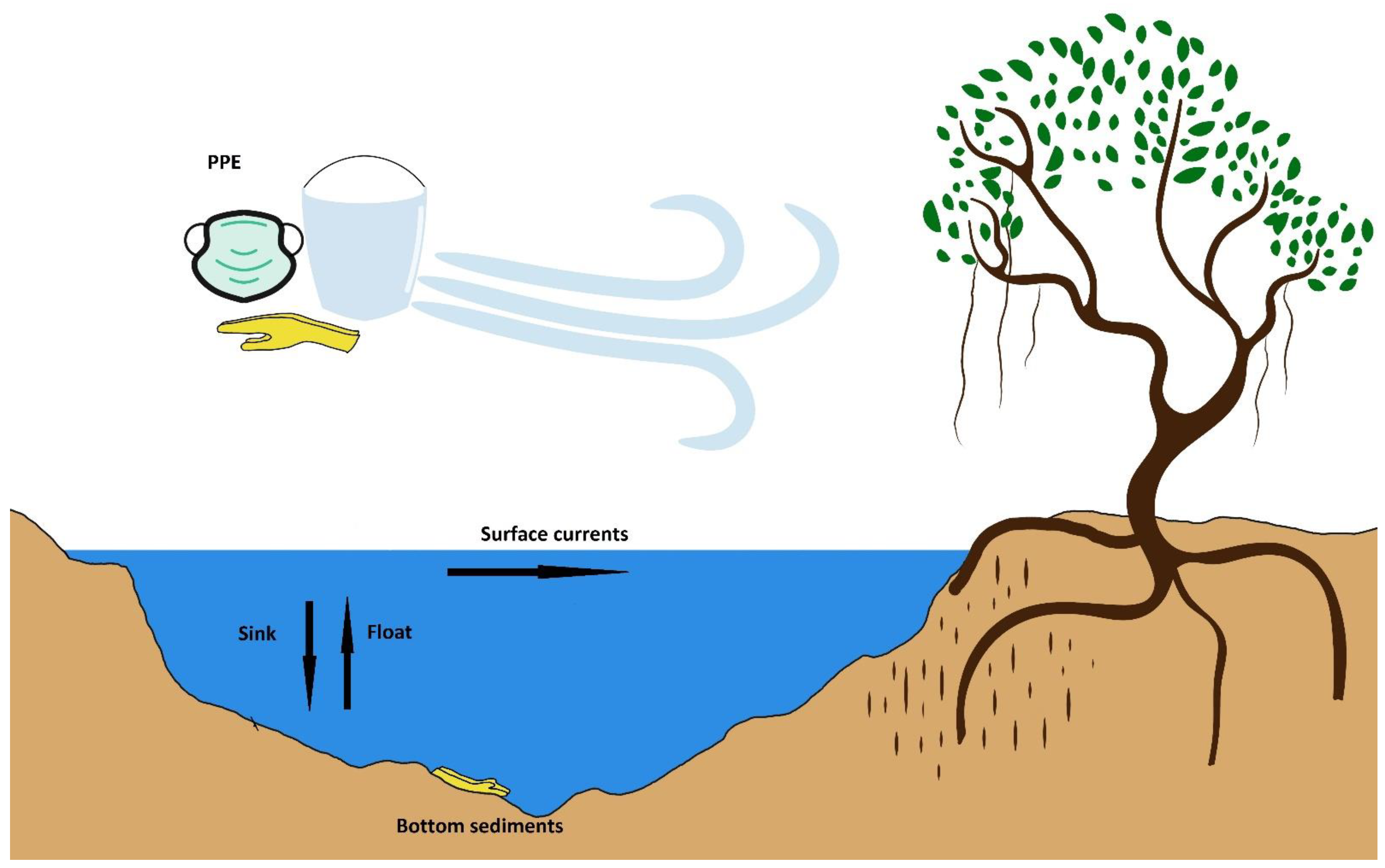
2. Mangrove Cyanotoxins and Their Legacy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/Genera | Habitat | Country | Reference |
|---|---|---|---|
| Gloeothece sp. | Sediment, water | Brazil, Egypt, India | [11] |
| Hydrocoleum sp. | Sediment | Egypt, India, Mexico, Saudi Arabia | [11] |
| Bostrychia sp. | Plant, mud and rock in the mangrove | Southern Africa, Europe | [11,26] |
| Chamaecalyx sp. | Epiphytic | Mozambique, Mexico | [11] |
| Acaryochloris sp. | Epiphytic biofilms on a red alga (Gelidium caulacantheum) colonizing the pneumatophores of a temperate mangrove (Avicennia marina) | Australia | [11] |
| Nostoc sp. | Sediment, water | Brazil, India, Tanzania, Cardoso Island, Bertioga | [11,27] |
| Anabaena sp. | Avicennia pneumatophores, rhizosphere, sediment, water | India, Mexico, Saudi Arabia, Tanzania | [11] |
| Arthrospira sp. | Epiphytic, sediment | Mozambique, Saudi Arabia, Tanzania | [11] |
| Calothrix sp. | Epiphytic, Avicennia marina pneumatophores, Brugiera gymnorrhiza knee roots, rhizosphere, rock, sediment, water, algae- and seagrass-associated | Brazil, Egypt, India, Mexico, Mozambique, Saudi Arabia, South Africa, Tanzania | [11] |
| Chroococcus sp. | Epiphytic, Avicennia pneumatophores, Brugiera gymnorrhiza knee roots, epiphytic, rhizosphere, sediment, water, associated with Bostrychia and Rhizoclonium algae | Brazil, Egypt, India, Mexico, Mozambique, Saudi Arabia, South Africa, Tanzania | [11] |
| Coleofasciculus sp. | Avicennia pneumatophores, Brugiera gymnorrhiza knee roots, Rhizophora mucronata prop roots, ephiphytic, rock surfaces, sediment, algae-associated, among Microcoleus tenerrimus | Brazil, Egypt, India, Mozambique, Saudi Arabia, South Africa, Tanzania | [11,28] |
| Dermocarpa sp. | Epiphytic, Avicennia marina pneumatophores, Brugiera gymnorrhiza knee roots, rhizosphere, sediment, associated with Bostrychia, Caloglossa, Enteromorpha and Rhizoclonium algae | India, Saudi Arabia, South Africa | [11] |
| Dichothrix sp. | Avicennia pneumatophores, Rhizophora prop roots, rhizosphere, sediment | India, Saudi Arabia | [11,29] |
| Gloeocapsa sp. | Epiphytic, Avicennia marina pneumatophores, rhizosphere, sediment, water | Brazil, Egypt, India, Saudi Arabia, Tanzania | [11] |
| Hydrococcus sp. | Epiphytic, associated with Bostrychia, Caloglossa, Enteromorpha and Rhizoclonium algae | Mozambique, South Africa | [11] |
| Lyngbya sp. | Epiphytic, Avicennia pneumatophores, Brugiera gymnorrhiza knee roots, Rhizophora roots/trunks, rhizosphere, rock, sediment, algae- and seagrass-associated, among Coleofasciculus (Microcoleus) chtonoplastes and Porphyrosiphon martensianus | Brazil, Egypt, India, Mexico, Mozambique, Saudi Arabia, South Africa, Tanzania | [11,18] |
| Merismopedia sp. | Rhizosphere, sediment, water, among Oscillatoria | Brazil, India, Saudi Arabia, Tanzania | [11,30] |
| Microcystis sp. | Epiphytic, rhizosphere, sediment, water | India | [11,20] |
| Nodularia sp. | Epiphytic, sediment, water | India, Mozambique, Tanzania | [11,31] |
| Oscillatoria sp. | Epiphytic, Aegiceras corniculatum aerial roots, Avicennia pneumatophores, Brugiera gymnorrhiza knee roots, Suaeda maritima aerial roots, rhizosphere, rock, sediment, water, algae- and seagrass-associated, among Coleofasciculus (Microcoleus) chtonoplastes | Brazil, Egypt, India, Mexico, Mozambique, Saudi Arabia, South Africa, Tanzania | [11,32] |
| Phormidium sp. | Epiphytic, Avicennia pneumatophores, Rhizophora roots/trunks, rhizosphere, rock, sediment, water, algae-associated, among Phormidium simplicissimum or Microcoleus tenerrimus | Brazil, Egypt, India, Mexico, South Africa, Tanzania | [11,33] |
| Pseudanabaena sp. | Epiphytic, sediment, water | India, Mexico, Saudi Arabia | [11,34] |
| Raphidiopsis sp. | Avicennia marina pneumatophores, sediment | Egypt, India | [11] |
| Rivularia sp. | Avicennia pneumatophores, Ceriops tagal bark, Rhizophora prop roots, sediment | Egypt, Saudi Arabia, South Africa, Tanzania | [11] |
| Schizothrix sp. | Aegiceras corniculatum aerial roots, Avicennia marina pneumatophores, Brugiera gymnorrhiza knee roots, Rhizophora mucronata prop roots, sediment, among Scytonema insulare | Brazil, Egypt, India, Mexico, Saudi Arabia, South Africa, Tanzania | [11] |
| Scytonema sp. | Avicennia pneumatophores, Brugiera knee roots, Ceriops tagal bark, Rhizophora roots/ trunks, rhizosphere, rock, sediment, water | Brazil, Egypt, India, Saudi Arabia, South Africa, Tanzania | [11] |
| Synechococcus sp. | Rhizosphere, sediment, water, associated to Bostrychia algae | Brazil, India, Saudi Arabia, Tanzania | [11] |
| Trichodesmium sp. | Rhizosphere, sediment, water | India, Tanzania | [11,35] |
| Xenococcus sp. | Epiphytic, Avicennia schaueriana pneumatophores, Rhizophora roots/trunks, sediment, algae-associated, among Coleofasciculus (Microcoleus) chthonoplastes | Brazil, India, Mexico, Mozambique, Saudi Arabia, South Africa | [11] |
3. Nitrogen Fixation: Can the New Nitrogen Induce Cyanotoxin Production in N-Poor Ecosystems?
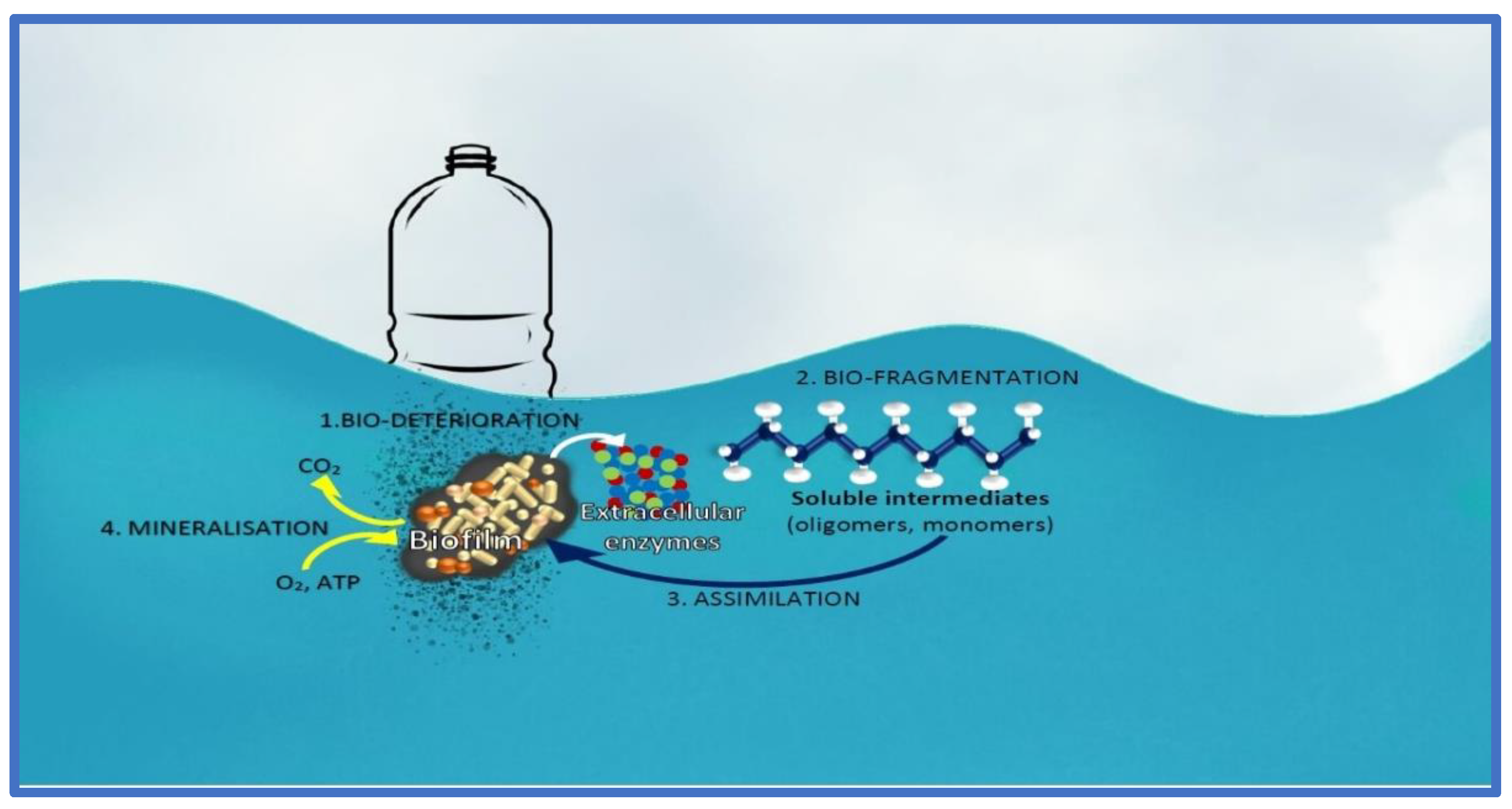
4. Cyanotoxins and the Role of the “Plastisphere”
5. Mangroves in the New Normal: The Ramifications on Ramsar Convention
6. The Path Ahead: Surveillance, Mitigation, Challenges and Newer Technologies
- The inability to decipher harmful and non-harmful bacteria by microscopical means.
- The temporal fluctuation of harmful cyanobacteria between seasons.
- The recalcitrance of harmful toxigenic cyanobacteria to cultivation.
- The false positives in PCR presented by dead cyanobacterial cells and environmental DNA that may not be indicative of cyanobacterial colonization.
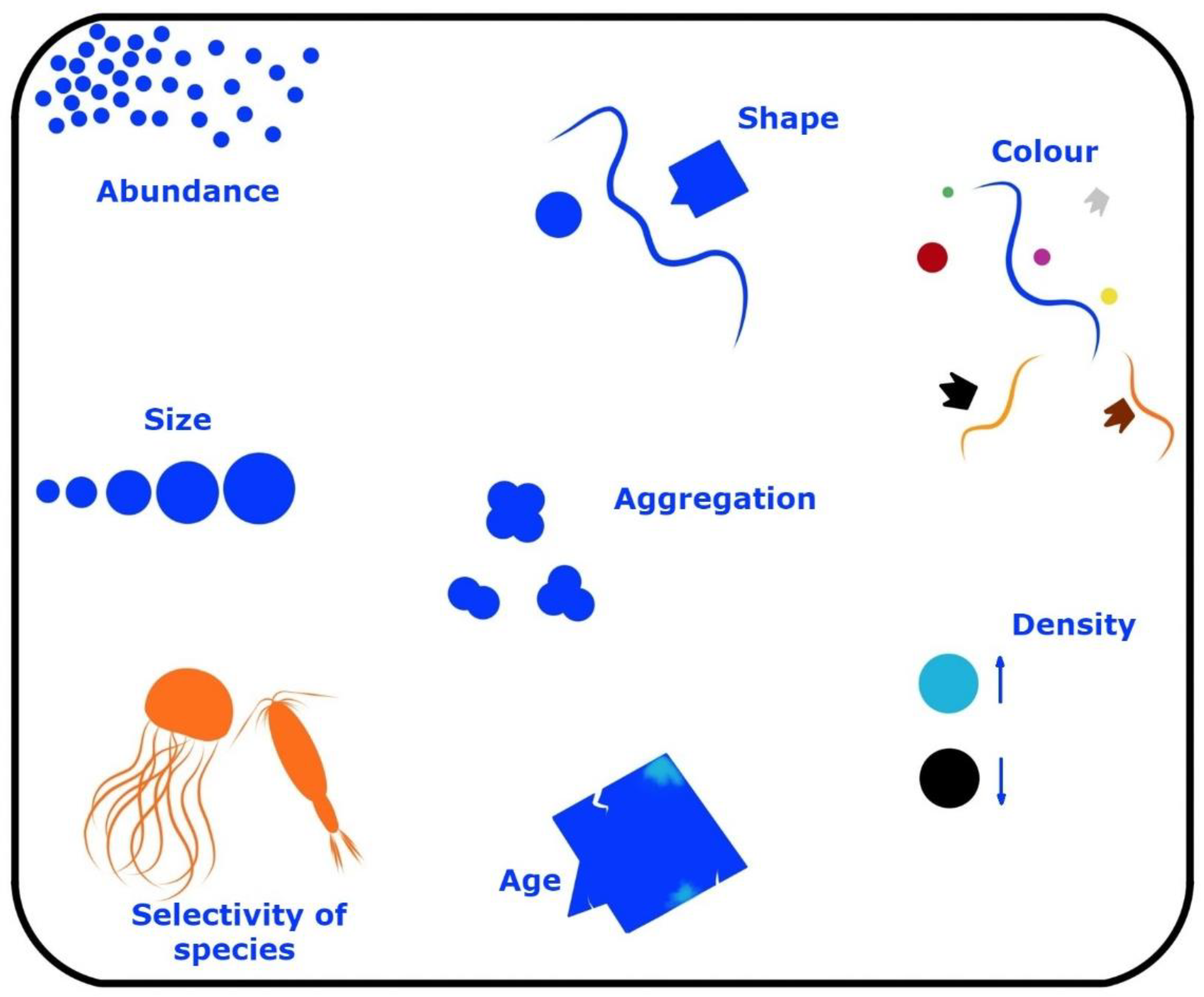
- The differences in plastic fragment sizes, shapes, edges, etc., that specify the cyanobacterial communities.
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuenzer, C.; Bluemel, A.; Gebhardt, S.; Quoc, T.V.; Dech, S. Remote sensing of mangrove ecosystems: A review. Remote Sens. 2011, 3, 878–928. [Google Scholar] [CrossRef]
- Barbier, E.B. The protective service of mangrove ecosystems: A review of valuation methods. Mar. Pollut. Bull. 2016, 109, 676–681. [Google Scholar] [CrossRef]
- Faridah-Hanum, I.; Latiff, A.; Hakeem, K.R.; Ozturk, M. Mangrove Ecosystems of Asia: Status, Challenges and Management Strategies; Springer Science & Business Media: New York, NY, USA, 2013. [Google Scholar]
- Janssen-Stelder, B.; Augustinus, P.; Van Santen, W. Sedimentation in a Coastal Mangrove System, Red River Delta, Vietnam. Proceedings in Marine Science; Elsevier: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Reef, R.; Feller, I.C.; Lovelock, C.E. Nutrition of mangroves. Tree Physiol. 2010, 30, 1148–1160. [Google Scholar] [CrossRef] [PubMed]
- Mearns, A.J.; Reish, D.J.; Oshida, P.S.; Morrison, A.M.; Rempel-Hester, M.A.; Arthur, C.; Rutherford, N.; Pryor, R. Effects of pollution on marine organisms. Water Environ. Res. 2016, 88, 1693–1807. [Google Scholar] [CrossRef] [PubMed]
- Kollimalai, S.; Kandasamy, K. Cyanobacterial diversity from mangrove sediment of south east coast of India. Asian J. Biodivers. 2013, 4. [Google Scholar]
- Lee, R.Y.; Joye, S.B. Seasonal patterns of nitrogen fixation and denitrification in oceanic mangrove habitats. Mar. Ecol. Prog. Ser. 2006, 307, 127–141. [Google Scholar] [CrossRef]
- Bownik, A. Harmful algae: Effects of alkaloid cyanotoxins on animal and human health. Toxin Rev. 2010, 29, 99–114. [Google Scholar] [CrossRef]
- Rejmánková, E.; Komárková, J. A function of cyanobacterial mats in phosphorus-limited tropical wetlands. Hydrobiologia 2000, 431, 135–153. [Google Scholar] [CrossRef]
- Alvarenga, D.O.; Rigonato, J.; Branco, L.H.Z.; Fiore, M.F. Cyanobacteria in mangrove ecosystems. Biodivers. Conserv. 2015, 24, 799–817. [Google Scholar] [CrossRef]
- Alfaro-Espinoza, G.; Ullrich, M.S. Bacterial N2-fixation in mangrove ecosystems: Insights from a diazotroph-mangrove interaction. Front. Microbiol. 2015, 6, 445. [Google Scholar] [CrossRef]
- Barcellos, D.; Queiroz, H.M.; Nóbrega, G.N.; de Oliveira Filho, R.L.; Santaella, S.T.; Otero, X.L.; Ferreira, T.O. Phosphorus enriched effluents increase eutrophication risks for mangrove systems in northeastern Brazil. Mar. Pollut. Bull. 2019, 142, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.A.; Coomes, M.W.; Smith, T.E. Isolation and sequence of the phosphoenolpyruvate carboxylase gene of the marine cyanobacterium Synechococcus PCC 7002. J. Biol. Sci. 2008, 8, 1261–1270. [Google Scholar] [CrossRef]
- Ribeiro, M.S.F.; Tucci, A.; Matarazzo, M.P.; Viana-Niero, C.; Nordi, C.S.F. Detection of Cyanotoxin-Producing Genes in a Eutrophic Reservoir (Billings Reservoir, São Paulo, Brazil). Water 2020, 12, 903. [Google Scholar] [CrossRef]
- Dolman, A.M.; Rücker, J.; Pick, F.R.; Fastner, J.; Rohrlack, T.; Mischke, U.; Wiedner, C. Cyanobacteria and cyanotoxins: The influence of nitrogen versus phosphorus. PLoS ONE 2012, 7, e38757. [Google Scholar] [CrossRef] [PubMed]
- Guidi-Rontani, C.; Jean, M.R.N.; Gonzalez-Rizzo, S.; Bolte-Kluge, S.; Gros, O. Description of new filamentous toxic Cyanobacteria (Oscillatoriales) colonizing the sulfidic periphyton mat in marine mangroves. FEMS Microbiol. Lett. 2014, 359, 173–181. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Al-Shehri, A.M. Biodiversity and toxin production of cyanobacteria in mangrove swamps in the Red Sea off the southern coast of Saudi Arabia. Bot. Mar. 2015, 58, 23–34. [Google Scholar] [CrossRef]
- Silva, C.S.; Genuário, D.B.; Vaz, M.G.; Fiore, M.F. Phylogeny of culturable cyanobacteria from Brazilian mangroves. Syst. Appl. Microbiol. 2014, 37, 100–112. [Google Scholar] [CrossRef]
- Zaheri, A.; Bahador, N.; Yousefzadi, M.; Arman, M. Molecular identification and toxicity effects of cyanobacteria species isolated from the Khoor-e-Khooran mangrove forest, Persian Gulf. Iran. J. Fish. Sci. 2021, 20, 572–589. [Google Scholar]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; CRC Press: Boca Raton, FL, USA; on behalf of the World Health Organization: Geneva, Switzerland, 2021; p. 858. [Google Scholar]
- Wijewickrama, M.M.; Manage, P.M. Accumulation of Microcystin-LR in grains of two rice varieties (Oryza sativa L.) and a leafy vegetable, Ipomoea aquatica. Toxins 2019, 11, 432. [Google Scholar] [CrossRef]
- Muro-Torres, V.M.; Amezcua, F.; Soto-Jiménez, M.; Balart, E.F.; Serviere-Zaragoza, E.; Green, L.; Rajnohova, J. Primary sources and food web structure of a tropical wetland with high density of mangrove forest. Water 2020, 12, 3105. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.D.S.; Kozlowski-Suzuki, B. Cyanotoxins: Bioaccumulation and effects on aquatic animals. Mar. Drugs 2011, 9, 2729–2772. [Google Scholar] [CrossRef]
- Pushpakumara, B.L.D.U.; Gunawardana, D. Preliminary data on the presence of an alternate vanadium nitrogenase in a culturable cyanobiont of Azolla pinnata R. Brown: Implications on Chronic Kidney Disease of an unknown etiology (CKDu). Data Brief 2018, 21, 2590–2597. [Google Scholar] [CrossRef] [PubMed]
- Nedumaran, T. Seaweed: A fertilizer for sustainable agriculture. In Sustainable Agriculture towards Food Security; Springer: Singapore, 2017; pp. 159–174. [Google Scholar]
- Rigonato, J.; Alvarenga, D.O.; Fiore, M.F. Tropical cyanobacteria and their biotechnological applications. In Diversity and Benefits of Microorganisms from the Tropics; Springer: Cham, Switzerland, 2017; pp. 139–167. [Google Scholar]
- Pramanik, P.J.; Mukherjee, J. Euryhalinema mangrovii gen. nov., sp. nov. and Leptoelongatus litoralis gen. nov., sp. nov.(Leptolyngbyaceae) isolated from an Indian mangrove forest. Phytotaxa 2019, 422, 058–074. [Google Scholar]
- Liu, L.; Wu, Y.; Hongxia, J.; Riding, R. Calcified rivulariaceans from the Ordovician of the Tarim Basin, Northwest China, Phanerozoic lagoonal examples, and possible controlling factors. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 448, 371–381. [Google Scholar] [CrossRef]
- Salma, U.; Bengen, D.G.; Kurniawan, F. Impact of mangrove and seagrass ecosystem on marine productivity of Pramuka Island, Seribu Islands, Indonesia. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2022; Volume 1109, p. 012103. [Google Scholar]
- Genuario, D.B.; Vaz, M.G.M.V.; Hentschke, G.S.; Sant’Anna, C.L.; Fiore, M.F. Halotia gen. nov., a phylogenetically and physiologically coherent cyanobacterial genus isolated from marine coastal environments. Int. J. Syst. Evol. Microbiol. 2015, 65, 663–675. [Google Scholar] [CrossRef]
- Ram, A.T.; Shamina, M. Cyanobacterial diversity from seven mangrove environments of Kerala, India. World News Nat. Sci. 2017, 9, 91–97. [Google Scholar]
- Gueye, M.; Ba, N.; Ngom, A.; Mbaye, M.S.; Noba, K. Cyanophytes of the Joal-Fadiouth Lagoon (Senegal). GSC Biol. Pharm. Sci. 2020, 12, 156–161. [Google Scholar] [CrossRef]
- Singh, T.; Bhadury, P. Description of a new marine planktonic cyanobacterial species Synechococcus moorigangaii (Order Chroococcales) from Sundarbans mangrove ecosystem. Phytotaxa 2019, 393, 263–277. [Google Scholar] [CrossRef]
- Briand, M.J.; Bonnet, X.; Goiran, C.; Guillou, G.; Letourneur, Y. Major sources of organic matter in a complex coral reef lagoon: Identification from isotopic signatures (δ13C and δ15N). PLoS ONE 2015, 10, e0131555. [Google Scholar] [CrossRef] [PubMed]
- Monchamp, M.E.; Pick, F.R.; Beisner, B.E.; Maranger, R. Nitrogen forms influence microcystin concentration and composition via changes in cyanobacterial community structure. PLoS ONE 2014, 9, e85573. [Google Scholar] [CrossRef]
- Reis, C.R.G.; Nardoto, G.B.; Oliveira, R.S. Global overview on nitrogen dynamics in mangroves and consequences of increasing nitrogen availability for these systems. Plant and soil. 2017, 2017. 410, 1–19. [Google Scholar] [CrossRef]
- Willis, A.; Chuang, A.W.; Burford, M.A. Nitrogen fixation by the diazotroph Cylindrospermopsis raciborskii (Cyanophyceae). J. Phycol. 2016, 52, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, J.A.; George, S.J.; Rubio, L.M. Molybdenum trafficking for nitrogen fixation. Biochemistry 2009, 48, 9711–9721. [Google Scholar] [CrossRef] [PubMed]
- Alongi, D.M. Nitrogen Cycling and Mass Balance in the World’s Mangrove Forests. Nitrogen 2020, 1, 167–189. [Google Scholar] [CrossRef]
- Beversdorf, L.J.; Miller, T.R.; McMahon, K.D. The role of nitrogen fixation in cyanobacterial bloom toxicity in a temperate, eutrophic lake. PLoS ONE 2013, 8, e56103. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.J.; Langille, M.G.I.; Walker, T.R. Food or just a free ride? A meta-analysis reveals the global diversity of the Plastisphere. ISME J. 2021, 15, 789–806. [Google Scholar] [CrossRef]
- Amaral-Zettler, L.A.; Zettler, E.R.; Mincer, T.J. Ecology of the plastisphere. Nat. Rev. Microbiol. 2020, 18, 139–151. [Google Scholar] [CrossRef]
- Dąbrowska, A. Microbial Degradation of Microplastics. In Recent Advances in Microbial Degradation; Springer: Singapore, 2021; pp. 373–387. [Google Scholar]
- Andrady, A.L. The plastic in microplastics: A review. Mar. Pollut. Bull. 2017, 119, 12–22. [Google Scholar] [CrossRef]
- Razeghi, N.; Hamidian, A.H.; Wu, C.; Zhang, Y.; Yang, M. Scientific studies on microplastics pollution in Iran: An in-depth review of the published articles. Mar. Pollut. Bull. 2021, 162, 111901. [Google Scholar] [CrossRef]
- Astner, A.F.; Hayes, D.G.; O’Neill, H.; Evans, B.R.; Pingali, S.V.; Urban, V.S.; Young, T.M. Mechanical formation of micro-and nano-plastic materials for environmental studies in agricultural ecosystems. Sci. Total Environ. 2019, 685, 1097–1106. [Google Scholar] [CrossRef]
- Botterell, Z.L.; Beaumont, N.; Dorrington, T.; Steinke, M.; Thompson, R.C.; Lindeque, P.K. Bioavailability and effects of microplastics on marine zooplankton: A review. Environ. Pollut. 2019, 245, 98–110. [Google Scholar] [CrossRef]
- Clerico, E.M.; Ditty, J.L.; Golden, S.S. Specialized techniques for site-directed mutagenesis in cyanobacteria. In Circadian rhythms; Humana Press: Totowa, NJ, USA, 2007; pp. 155–171. [Google Scholar]
- De-la-Torre, G.E.; Aragaw, T.A. What we need to know about PPE associated with the COVID-19 pandemic in the marine environment. Marine pollution bulletin 2021, 163, 111879. [Google Scholar] [CrossRef]
- Xu, B.; Liu, F.; Cryder, Z.; Huang, D.; Lu, Z.; He, Y.; Wang, H.; Lu, Z.; Brookes, P.C.; Tang, C.; et al. Microplastics in the soil environment: Occurrence, risks, interactions and fate–a review. Crit. Rev. Environ. Sci. Technol. 2020, 50, 2175–2222. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Codd, G.A. Co-occurrence of cyanobacteria and cyanotoxins with other environmental health hazards: Impacts and implications. Toxins 2020, 12, 629. [Google Scholar] [CrossRef]
- Karakolis, E.G.; Nguyen, B.; You, J.B.; Graham, P.J.; Rochman, C.M.; Sinton, D. Digestible fluorescent coatings for cumulative quantification of microplastic ingestion. Environ. Sci. Technol. Lett. 2018, 5, 62–67. [Google Scholar] [CrossRef]
- Nava, V.; Leoni, B. A critical review of interactions between microplastics, microalgae and aquatic ecosystem function. Water Res. 2021, 188, 116476. [Google Scholar] [CrossRef]
- van Bijsterveldt, C.E.; van Wesenbeeck, B.K.; Ramadhani, S.; Raven, O.V.; van Gool, F.E.; Pribadi, R.; Bouma, T.J. Does plastic waste kill mangroves? A field experiment to assess the impact of macro plastics on mangrove growth, stress response and survival. Sci. Total Environ. 2021, 756, 143826. [Google Scholar] [CrossRef] [PubMed]
- Govender, J.; Naidoo, T.; Rajkaran, A.; Cebekhulu, S.; Bhugeloo, A. Towards characterising microplastic abundance, typology and retention in mangrove-dominated estuaries. Water 2020, 12, 2802. [Google Scholar] [CrossRef]
- Parashar, N.; Hait, S. Plastics in the time of COVID-19 pandemic: Protector or polluter? Sci. Total Environ. 2021, 759, 144274. [Google Scholar] [CrossRef]
- Esterhuizen, M.; Kim, Y.J. Effects of polypropylene, polyvinyl chloride, polyethylene terephthalate, polyurethane, high-density polyethylene, and polystyrene microplastic on Nelumbo nucifera (Lotus) in water and sediment. Environ. Sci. Pollut. Res. 2022, 29, 17580–17590. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, D. The ‘ecological character’ of wetlands: A foundational concept in the Ramsar Convention, yet still cause for debate 50 years later. Marine Freshw. Res. 2021, 73, 1127–1133. [Google Scholar] [CrossRef]
- Garcés-Ordóñez, O.; Mejía-Esquivia, K.A.; Sierra-Labastidas, T.; Patiño, A.; Blandón, L.M.; Díaz, L.F.E. Prevalence of microplastic contamination in the digestive tract of fishes from mangrove ecosystem in Cispata, Colombian Caribbean. Marine pollution bulletin. 2020, 154, 111085. [Google Scholar] [CrossRef]
- Maghsodian, Z.; Sanati, A.M.; Tahmasebi, S.; Shahriari, M.H.; Ramavandi, B. Study of microplastics pollution in sediments and organisms in mangrove forests: A review. Environ. Res. 2022, 208, 112725. [Google Scholar] [CrossRef]
- van Bijsterveldt, C.E.; van Wesenbeeck, B.K.; van der Wal, D.; Afiati, N.; Pribadi, R.; Brown, B.; Bouma, T.J. How to restore mangroves for greenbelt creation along eroding coasts with abandoned aquaculture ponds. Estuarine, coastal and shelf science. 2020, 235, 106576. [Google Scholar] [CrossRef]
- Martin, C.; Baalkhuyur, F.; Valluzzi, L.; Saderne, V.; Cusack, M.; Almahasheer, H.; Krishnakumar, P.K.; Rabaoui, L.; Qurban, M.A.; Arias-Ortiz, A.; et al. Exponential increase of plastic burial in mangrove sediments as a major plastic sink. Sci. Adv. 2020, 6, eaaz5593. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Guo, J.; Huang, F.; Massey, I.Y.; Huang, R.; Li, Y.; Wen, C.; Ding, P.; Zeng, W.; Liang, G. Removal of Microcystin-LR by a Novel Native Effective Bacterial Community Designated as YFMCD4 Isolated from Lake Taihu. Toxins 2018, 10, 363. [Google Scholar] [CrossRef]
- Howard, M.D.; Nagoda, C.; Kudela, R.M.; Hayashi, K.; Tatters, A.; Caron, D.A.; Busse, L.; Brown, J.; Sutula, M.; Stein, E.D. Microcystin prevalence throughout lentic waterbodies in coastal southern California. Toxins 2017, 9, 231. [Google Scholar] [CrossRef] [PubMed]
- Leiser, R.; Wu, G.; Neu, T.R.; Potthoff, K.W. Biofouling, metal sorption and aggregation are related to sinking of microplastics in a stratified reservoir. Water Res. 2020, 176, 115748. [Google Scholar] [CrossRef]
- Chen, X.; Xiong, X.; Jiang, X.; Shi, H.; Wu, C. Sinking of floating plastic debris caused by biofilm development in a freshwater lake. Chemosphere 2019, 222, 856–864. [Google Scholar] [CrossRef]
- Henao, E.; Rzymski, P.; Waters, M.N. A Review on the Study of Cyanotoxins in Paleolimnological Research: Current Knowledge and Future Needs. Toxins 2019, 12, 6. [Google Scholar] [CrossRef]
- Österholm, J.; Rafael, P.; David, F.; Kaarina, S. Phylogenomic Analysis of Secondary Metabolism in the Toxic Cyanobacterial Genera Anabaena, Dolichospermum and Aphanizomenon. Toxins 2020, 12, 248. [Google Scholar] [CrossRef]
- Mlouka, A.K.; Comte, A.M.; Castets, C.; Bouchier, N. Tandeau de Marsac. The gas vesicle gene cluster from Microcystis aeruginosa and DNA rearrangements that lead to loss of cell buoyancy. J. Bacteriol. 2014, 186, 2355–2365. [Google Scholar] [CrossRef] [PubMed]
- Barone, G.D.; Ferizović, D.; Biundo, A.; Lindblad, P. Hints at the Applicability of Microalgae and Cyanobacteria for the Biodegradation of Plastics. Sustainability 2020, 12, 10449. [Google Scholar] [CrossRef]
- Jacquin, J.; Cheng, J.; Odobel, C.; Conan, P.; Pujo-pay, M.; Meistertzheim, A.; Jean-francois, G. Microbial ecotoxicology of marine plastic debris: A review on colonization and biodegradation by the ‘plastisphere’. Front. Microbiol. 2018, 10, 865. [Google Scholar] [CrossRef]
- Hoiczyk, E.; Hansel, A. Cyanobacterial cell walls: News from an unusual prokaryotic envelope. J. Bacteriol. 2000, 182, 1191–1199. [Google Scholar] [CrossRef] [PubMed]
- Palm, G.J.; Reisky, L.; Böttcher, D.; Müller, H.; Michels, E.A.; Walczak, M.C.; Berndt, L.; Weiss, M.S.; Bornscheuer, U.T.; Weber, G. Structure of the plastic-degrading Ideonella sakaiensis MHETase bound to a substrate. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Idroos, E.S.; Manage, P.M. Bioremediation of Microcystins by Two Native Bacteria; Bacillus Cereus and Rahnella Aquatilis. Asian J. Microbiol. Biotechnol. Environ. Sci. 2018, 20, 24–32. [Google Scholar]





| Type of Plastic | Effects on the Aquatic Vegetation |
|---|---|
| polyurethane (PU) | Reduction of seed germination. Growth inhibition. Significant effects on anti-oxidative enzyme activity. |
| polypropylene (PP) | Reduction of seed germination. Growth inhibition. Significant effects on anti-oxidative enzyme activity. |
| polycarbonate (PC) | Data not available. |
| high-density polyethylene (HDPE) | No influence on seed germination. Growth inhibition. Effects on anti-oxidative enzyme activity. |
| polyvinyl chloride (PVC) | Reduction of seed germination Growth inhibition. Significant effects on anti-oxidative enzyme activity. |
| low-density polyethylene (LDPE) | Data not available. |
| polystyrene (PS) | No influence on seed germination. Growth inhibition. Significant effects on anti-oxidative enzyme activity. |
| polyethylene terephthalate (PET) | No influence on seed germination. No growth inhibition. Significant effects on anti-oxidative enzyme activity. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gunawardana, D.; Abeysiri, S.; Manage, P. Legacy of “New Normal” Plastics and “New Nitrogen” in the Cyanotoxin Footprint in Mangrove Ecosystems. Phycology 2023, 3, 106-126. https://doi.org/10.3390/phycology3010007
Gunawardana D, Abeysiri S, Manage P. Legacy of “New Normal” Plastics and “New Nitrogen” in the Cyanotoxin Footprint in Mangrove Ecosystems. Phycology. 2023; 3(1):106-126. https://doi.org/10.3390/phycology3010007
Chicago/Turabian StyleGunawardana, Dilantha, Sashika Abeysiri, and Pathmalal Manage. 2023. "Legacy of “New Normal” Plastics and “New Nitrogen” in the Cyanotoxin Footprint in Mangrove Ecosystems" Phycology 3, no. 1: 106-126. https://doi.org/10.3390/phycology3010007
APA StyleGunawardana, D., Abeysiri, S., & Manage, P. (2023). Legacy of “New Normal” Plastics and “New Nitrogen” in the Cyanotoxin Footprint in Mangrove Ecosystems. Phycology, 3(1), 106-126. https://doi.org/10.3390/phycology3010007