
Screening of Omicron Virus Strain by Quantifying the Spike Protein Content
Abstract
1. Introduction
2. Methods
2.1. Animals
2.2. Animal Ethics Statement
2.3. Virus Strains and Potential Vaccine Development
2.4. Lowry Method
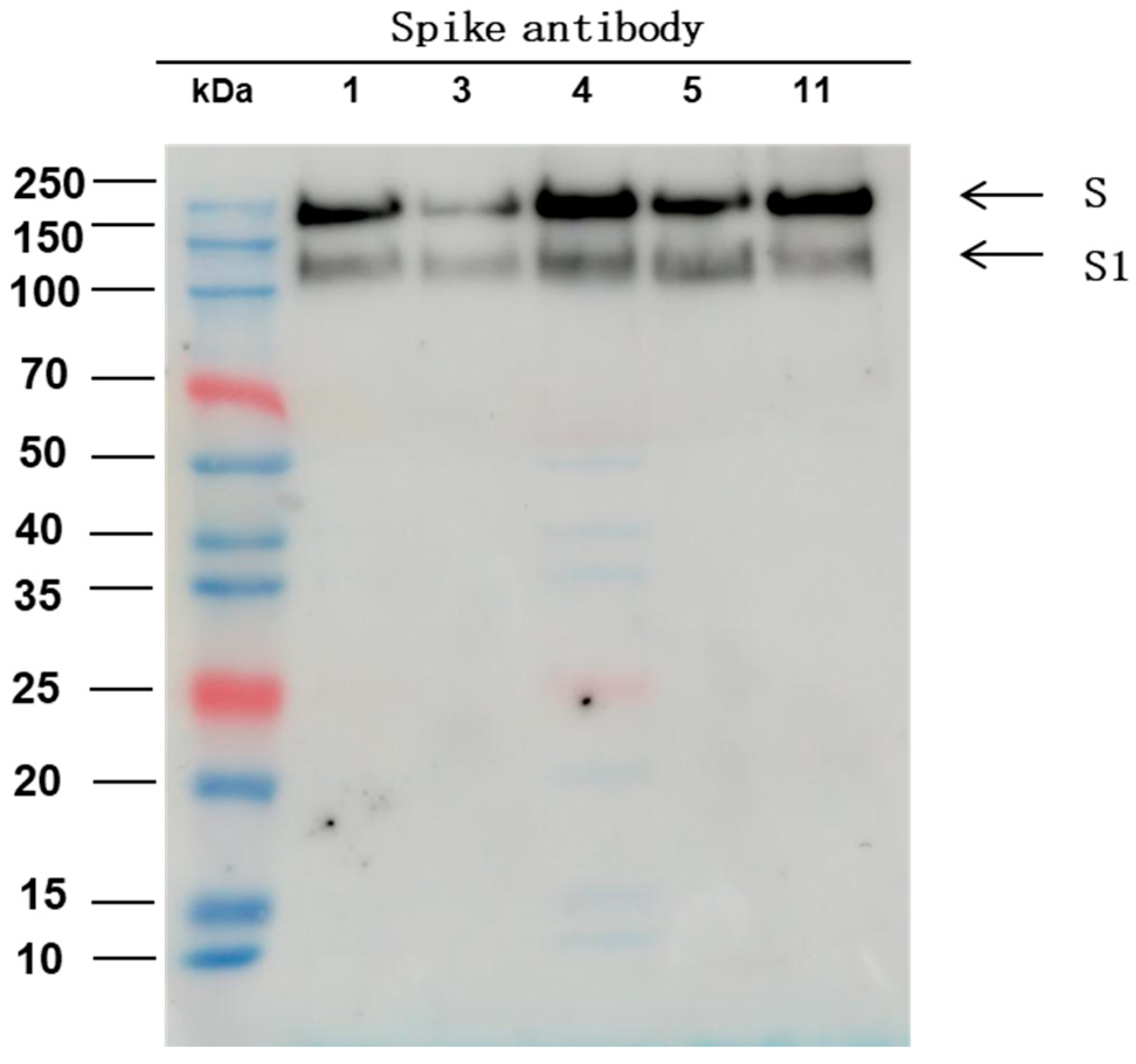
2.5. Western Blot
2.6. Antigen Analysis Based on Multiclonal Antibody ELISA
2.7. Antigen Analysis Based on Monoclonal Antibody ELISA
2.8. Virus Titer Detection
2.9. Surface Plasmon Resonance
2.10. Immunogenicity Analysis
2.11. Statistical Analysis
3. Results
3.1. Spike Protein Content Analysis
3.2. Affinity Analysis
3.3. Virus Titer Analysis
3.4. Neutralizing Antibody Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Viana, R.; Moyo, S.; Amoako, D.G.; Tegally, H.; Scheepers, C.; Althaus, C.L.; Anyaneji, U.J.; Bester, P.A.; Boni, M.F.; Chand, M.; et al. Rapid epidemic expansion of the SARS-CoV-2 Omicron variant in southern Africa. Nature 2022, 603, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Beltran, W.F.; St Denis, K.J.; Hoelzemer, A.; Lam, E.C.; Nitido, A.D.; Sheehan, M.L.; Berrios, C.; Ofoman, O.; Chang, C.C.; Hauser, B.M.; et al. mRNA-based COVID-19 vaccine boosters induce neutralizing immunity against SARS-CoV-2 Omicron variant. medRxiv 2021, 14:2021.12.14.21267755, Update in: Cell 2022, 185, 457–466.e4. https://doi.org/10.1016/j.cell.2021.12.033. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Berkhout, B.; Herrera-Carrillo, E. SARS-CoV-2 Evolution: On the Sudden Appearance of the Omicron Variant. J. Virol. 2022, 96, e0009022. [Google Scholar] [CrossRef] [PubMed]
- Tegally, H.; Moir, M.; Everatt, J.; Giovanetti, M.; Scheepers, C.; Wilkinson, E.; Subramoney, K.; Makatini, Z.; Moyo, S.; Amoako, D.G.; et al. Emergence of SARS-CoV-2 Omicron lineages BA.4 and BA.5 in South Africa. Nat. Med. 2022, 28, 1785–1790. [Google Scholar] [CrossRef]
- Tuekprakhon, A.; Nutalai, R.; Dijokaite-Guraliuc, A.; Zhou, D.; Ginn, H.M.; Selvaraj, M.; Liu, C.; Mentzer, A.J.; Supasa, P.; Duyvesteyn, H.M.; et al. Antibody escape of SARS-CoV-2 Omicron BA.4 and BA.5 from vaccine and BA.1 serum. Cell 2022, 185, 2422–2433.e13. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Song, W.; Wang, L.; Jian, F.; Chen, X.; Gao, F.; Shen, Z.; Wang, Y.; Wang, X.; Cao, Y. ACE2 binding and antibody evasion in enhanced transmissibility of XBB.1.5. Lancet Infect. Dis. 2023, 23, 278–280. [Google Scholar] [CrossRef] [PubMed]
- Parums, D.V. Editorial: The XBB.1.5 (‘Kraken’) Subvariant of Omicron SARS-CoV-2 and its Rapid Global Spread. Med. Sci. Monit. 2023, 29, e939580. [Google Scholar] [CrossRef] [PubMed]
- Kaku, Y.; Okumura, K.; Padilla-Blanco, M.; Kosugi, Y.; Uriu, K.; Hinay, A.A.; Chen, L.; Plianchaisuk, A.; Kobiyama, K.; Ishii, K.J.; et al. Virological characteristics of the SARS-CoV-2 JN.1 variant. Lancet Infect. Dis. 2024, 24, e82. [Google Scholar] [CrossRef]
- Yang, S.; Yu, Y.; Xu, Y.; Jian, F.; Song, W.; Yisimayi, A.; Wang, P.; Wang, J.; Liu, J.; Yu, L.; et al. Fast evolution of SARS-CoV-2 BA.2.86 to JN.1 under heavy immune pressure. Lancet Infect. Dis. 2024, 24, e70–e72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, Y.; Liang, H.; Xu, Y.; Zhou, C.; Yao, Y.; Wang, H.; Yang, X. Innovation-driven trend shaping COVID-19 vaccine development in China. Front. Med. 2023, 17, 1096–1116. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Y.; Huang, B.; Deng, W.; Quan, Y.; Wang, W.; Xu, W.; Zhao, Y.; Li, N.; Zhang, J.; et al. Development of an Inactivated Vaccine Candidate, BBIBP-CorV, with Potent Protection against SARS-CoV-2. Cell 2020, 182, 713–721.e9. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Li, Z.; Zhang, X.; Li, J.; Zhu, F. CoronaVac: A review of efficacy, safety, and immunogenicity of the inactivated vaccine against SARS-CoV-2. Hum. Vaccines Immunother. 2022, 18, 2096970. [Google Scholar] [CrossRef] [PubMed]
- Bravo, L.; Smolenov, I.; Han, H.H.; Li, P.; Hosain, R.; Rockhold, F.; Clemens, S.A.C.; Roa, C.; Borja-Tabora, C.; Quinsaat, A.; et al. Efficacy of the adjuvanted subunit protein COVID-19 vaccine, SCB-2019: A phase 2 and 3 multicentre, double-blind, randomised, placebo-controlled trial. Lancet 2022, 399, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Gao, L.; Tao, L.; Hadinegoro, S.R.; Erkin, M.; Ying, Z.; He, P.; Girsang, R.T.; Vergara, H.; Akram, J.; et al. Efficacy and Safety of the RBD-Dimer-Based COVID-19 Vaccine ZF2001 in Adults. N. Engl. J. Med. 2022, 386, 2097–2111. [Google Scholar] [CrossRef] [PubMed]
- Schlake, T.; Thess, A.; Fotin-Mleczek, M.; Kallen, K.J. Developing mRNA-vaccine technologies. RNA Biol. 2012, 9, 1319–1330. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Soleimanpour, S.; Yaghoubi, A. COVID-19 vaccine: Where are we now and where should we go? Expert. Rev. Vaccines 2021, 20, 23–44. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, M.N.; Minassian, A.M.; Ewer, K.J.; Flaxman, A.L.; Folegatti, P.M.; Owens, D.R.; Voysey, M.; Aley, P.K.; Angus, B.; Babbage, G.; et al. Safety and immunogenicity of ChAdOx1 nCoV-19 vaccine administered in a prime-boost regimen in young and old adults (COV002): A single-blind, randomised, controlled, phase 2/3 trial. Lancet 2021, 396, 1979–1993. [Google Scholar] [CrossRef] [PubMed]
- Sadoff, J.; Gray, G.; Vandebosch, A.; Cárdenas, V.; Shukarev, G.; Grinsztejn, B.; Goepfert, P.A.; Truyers, C.; Fennema, H.; Spiessens, B.; et al. Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against COVID-19. N. Engl. J. Med. 2021, 384, 2187–2201. [Google Scholar] [CrossRef] [PubMed]
- Al Kaabi, N.; Zhang, Y.; Xia, S.; Yang, Y.; Al Qahtani, M.M.; Abdulrazzaq, N.; Al Nusair, M.; Hassany, M.; Jawad, J.S.; Abdalla, J.; et al. Effect of 2 Inactivated SARS-CoV-2 Vaccines on Symptomatic COVID-19 Infection in Adults: A Randomized Clinical Trial. JAMA 2021, 326, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Bueno, S.M.; Abarca, K.; González, P.A.; Gálvez, N.M.S.; Soto, J.A.; Duarte, L.F.; Schultz, B.M.; Pacheco, G.A.; González, L.A.; Vázquez, Y.; et al. Safety and Immunogenicity of an Inactivated Severe Acute Respiratory Syndrome Coronavirus 2 Vaccine in a Subgroup of Healthy Adults in Chile. Clin. Infect. Dis. 2022, 75, e792–e804. [Google Scholar] [CrossRef] [PubMed]
- Stelzer-Braid, S.; Walker, G.J.; Aggarwal, A.; Isaacs, S.R.; Yeang, M.; Naing, Z.; Stella, A.O.; Turville, S.G.; Rawlinson, W.D. Virus isolation of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) for diagnostic and research purposes. Pathology 2020, 52, 760–763. [Google Scholar] [CrossRef] [PubMed]
- Knowles, B.; Bonachela, J.A.; Cieslik, N.; Della Penna, A.; Diaz, B.; Baetge, N.; Behrenfeld, M.J.; Naumovitz, K.; Boss, E.; Graff, J.R.; et al. Altered growth and death in dilution-based viral predation assays. PLoS ONE 2023, 18, e0288114. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, N.; Murayama, A.; Suzuki, R.; Watanabe, N.; Shiina, M.; Liang, T.J.; Wakita, T.; Kato, T. Single strain isolation method for cell culture-adapted hepatitis C virus by end-point dilution and infection. PLoS ONE 2014, 9, e98168. [Google Scholar] [CrossRef] [PubMed]
- Gerganov, G.; Sŭrtmadzhiev, K. [Plaque cloning of the velogenic viscerotropic Newcastle disease virus]. Vet. Med. Nauki 1981, 18, 12–19. [Google Scholar] [PubMed]
- Hirano, N.; Tawara, T.; Nomura, R.; Imai, A.; Ono, K.; Yamaguchi, R. Sensitive plaque assay and propagation of Chuzan (Kasba) virus, a Palyam serogroup orbivirus, in BHK-21 cells. Zentralbl Vet. B 1996, 43, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Flint, J.S.; Enquist, L.W.; Shalka, A.M. Virological methods. In Principles of Virology, 3rd ed.; ASM: Almere, The Netherlands, 2009; Chapter 2. [Google Scholar]
- Smither, S.J.; Lear-Rooney, C.; Biggins, J.; Pettitt, J.; Lever, M.S.; Olinger, G. Comparison of the plaque assay and 50% tissue culture infectious dose assay as methods for measuring filovirus infectivity. J. Virol. Methods 2013, 193, 565–571. [Google Scholar] [CrossRef]
- Cooper, P.D. A method for producing plaques in agar suspensions of animal cells. Virology 1955, 1, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Dulbecco, R. Production of Plaques in Monolayer Tissue Cultures by Single Particles of an Animal Virus. Proc. Natl. Acad. Sci. USA 1952, 38, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, M.T.; Mermelstein, A.G.; Parker Miller, E.; Seth, P.C.; Stancofski, E.S.D.; Fera, D. Structural Analysis of Neutralizing Epitopes of the SARS-CoV-2 Spike to Guide Therapy and Vaccine Design Strategies. Viruses 2021, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Public Law 99-198, Food Security Act of 1985, Subtitle F—Animal Welfare, U.S. DEP’T AGRIC. NAT’L AGRIC. LIBR. [Hereinafter Food Security Act of 1985]; See also Food Security Act of 1985 sec. 1752(a), § 13(2)(A). Available online: https://www.nal.usda.gov/awic/public-law-99-198-foodsecurity-act-1985-subtitle-f-animal-welfare (accessed on 20 October 2016).
- National Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China: Part Three; China Medical Technology Press: Beijing, China, 2020. [Google Scholar]
- Zapata-Cardona, M.I.; Flórez-Álvarez, L.; Gómez-Gallego, D.M.; Moncada-Díaz, M.J.; Hernandez, J.C.; Díaz, F.; Rugeles, M.T.; Aguilar-Jiménez, W.; Zapata, W. Comparison among plaque assay, tissue culture infectious dose (TCID(50)) and real-time RT-PCR for SARS-CoV-2 variants quantification. Iran. J. Microbiol. 2022, 14, 291–299. [Google Scholar] [PubMed]
- Witt, A.S.A.; Trindade, G.d.S.; Gil de Souza, F.; Serafim, M.S.M.; da Costa, A.V.B.; Silva, M.V.F.; Iani, F.C.d.M.; Rodrigues, R.A.L.; Kroon, E.G.; Abrahão, J.S. Ultrastructural analysis of monkeypox virus replication in Vero cells. J. Med. Virol. 2023, 95, e28536. [Google Scholar] [CrossRef] [PubMed]
- Mardanly, S.G.; Kazakov, A.A.; Demkin, V.V.; Zatevalov, A.M.; Mironov, A.Y. Development of a PCR assay for the detection of human herpes virus type 7. Klin. Lab. Diagn. 2022, 67, 658–662. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tan, W.; Lou, Z.; Zhao, Y.; Zhang, J.; Liang, H.; Li, N.; Zhu, X.; Ding, L.; Huang, B.; et al. Vaccination with Omicron Inactivated Vaccine in Pre-vaccinated Mice Protects against SARS-CoV-2 Prototype and Omicron Variants. Vaccines 2022, 10, 1149. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.L.; Garçon, N.; Leo, O.; Friedland, L.R.; Strugnell, R.; Laupèze, B.; Doherty, M.; Stern, P. Vaccine development: From concept to early clinical testing. Vaccine 2016, 34, 6655–6664. [Google Scholar] [CrossRef] [PubMed]
- Delany, I.; Rappuoli, R.; De Gregorio, E. Vaccines for the 21st century. EMBO Mol. Med. 2014, 6, 708–720. [Google Scholar] [CrossRef] [PubMed]
- Vetter, V.; Denizer, G.; Friedland, L.R.; Krishnan, J.; Shapiro, M. Understanding modern-day vaccines: What you need to know. Ann. Med. 2018, 50, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Nuwarda, R.F.; Alharbi, A.A.; Kayser, V. An Overview of Influenza Viruses and Vaccines. Vaccines 2021, 9, 1032. [Google Scholar] [CrossRef] [PubMed]
- Sano, K.; Ainai, A.; Suzuki, T.; Hasegawa, H. Intranasal inactivated influenza vaccines for the prevention of seasonal influenza epidemics. Expert. Rev. Vaccines 2018, 17, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Benfield, T.L.; Helweg-Larsen, J. [SARS-CoV-2 vaccines]. Ugeskr. Laeger 2021, 183, V01210052. [Google Scholar] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Strain | Protein Content (μg/mL) |
|---|---|
| #1 | 2639.79 |
| #3 | 2513.62 |
| #4 | 2429.51 |
| #5 | 2322.45 |
| #11 | 2555.68 |
| Type | 10−2 | 10−3 | 10−4 | 10−5 | Titer (LgCCID50/mL) |
|---|---|---|---|---|---|
| #1 | 8/8 | 5/8 | 0/8 | 0/8 | 4.125 |
| #3 | 8/8 | 5/8 | 0/8 | 0/8 | 4.125 |
| #4 | 8/8 | 8/8 | 1/8 | 0/8 | 4.625 |
| #5 | 8/8 | 5/8 | 0/8 | 0/8 | 4.125 |
| #11 | 8/8 | 5/8 | 0/8 | 0/8 | 4.125 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Z.; Chang, H.; Wang, Y.; Xie, S.; Liu, Y.; Zhao, Y.; Li, N.; Zhang, Y. Screening of Omicron Virus Strain by Quantifying the Spike Protein Content. COVID 2024, 4, 838-847. https://doi.org/10.3390/covid4060056
He Z, Chang H, Wang Y, Xie S, Liu Y, Zhao Y, Li N, Zhang Y. Screening of Omicron Virus Strain by Quantifying the Spike Protein Content. COVID. 2024; 4(6):838-847. https://doi.org/10.3390/covid4060056
Chicago/Turabian StyleHe, Zhenyu, Hengzhen Chang, Yichuan Wang, Siman Xie, Yingwei Liu, Yuxiu Zhao, Na Li, and Yuntao Zhang. 2024. "Screening of Omicron Virus Strain by Quantifying the Spike Protein Content" COVID 4, no. 6: 838-847. https://doi.org/10.3390/covid4060056
APA StyleHe, Z., Chang, H., Wang, Y., Xie, S., Liu, Y., Zhao, Y., Li, N., & Zhang, Y. (2024). Screening of Omicron Virus Strain by Quantifying the Spike Protein Content. COVID, 4(6), 838-847. https://doi.org/10.3390/covid4060056